Introduction
Spinal cord injury (SCI) is a common trauma that
threatens human health seriously and can occur at any age (1). It frequently results in failure in limb
sensation, motor function and automatic nervous function. The high
disability and mortality rates of SCI often have a huge
physiological and psychological effect on individuals, their
families and society (2). However,
there is no effective treatment due to the complicated
pathophysiological mechanism of SCI, although numerous clinical and
animal studies have been conducted (3,4).
SCI involves a series of cytological and molecular
biology cascade reactions (5). It
can be divided into primary and secondary injury (1). Primary injury of the spinal cord may
result in irreversible death of a large number of neurons in the
damaged tissue (6), with no
effective methods to regenerate the neurons available to date.
Primary injury may then result in secondary injury after several
min or hfrom SCI. Thus, it further results in neuronal injury and
expansion of the SCI area (5,6). The
early application of drugs prevents secondary injury; thus, the
condition of spinal cord tissues can be improved to preserve the
anatomical structure for functional recovery (6). Therefore, early drug administration is
important for acute SCI treatment.
The inflammatory reaction is a major component of
secondary SCI and is a key constituent of the pathophysiological
mechanism of acute SCI. Inflammation may promote a series of
molecular biological events, resulting in the activation of
inflammatory cells from the spinal cord tissues and infiltration of
the circulatory system, the release of various pro-inflammatory
mediators and neurotoxins, and the generation of oxygen radicals
and nitroso compounds, and consequently causing cell damage
(7,8).
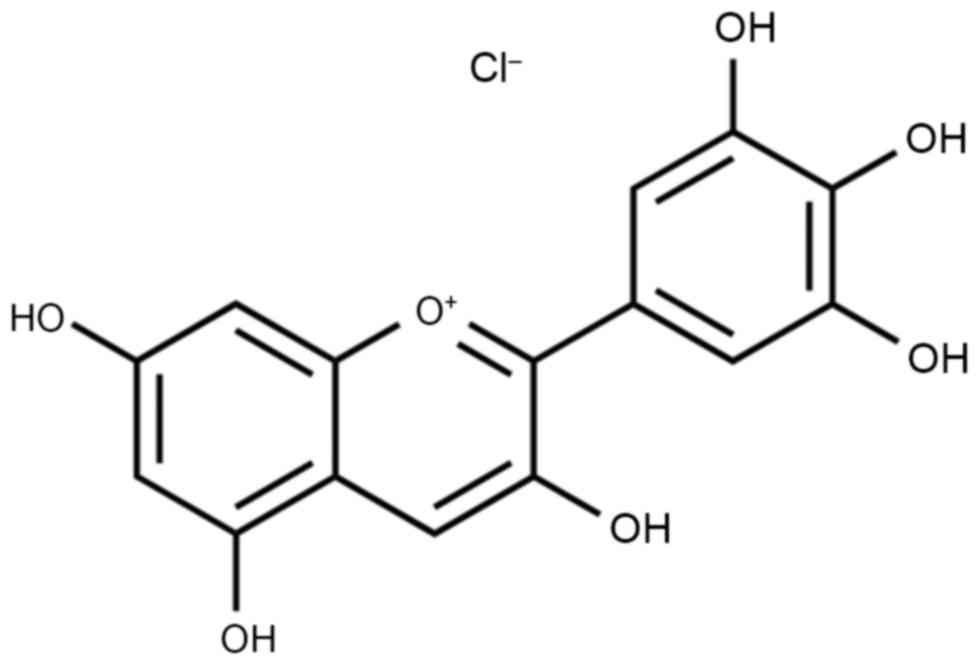
Delphinidin (Fig. 1)
is a polyphenolic substance that is derived from catechinic acid
and epicatechin (9). It is widely
obtained from the pericarp, testa and bark of various plants.
Delphinidin extracted from grape seeds has a high purity and
oligomer content. As a representative of the flavonoid polyphenolic
substances, delphinidin is widely found in plant-based food
(10). It has powerful antibiosis,
anti-inflammatory, anti-allergenic and anti-oxygenation activities
(11).
Delphinidin has been extensively used as a
complementary anti-inflammation and trauma treatment in the form of
a food supplement (12,13). The present study investigated whether
the anti-inflammatory effect of delphinidin alleviates the
SCI-induced inflammation in a rat model through intramedullary
spinal pressure.
Materials and methods
Animal care and groups
All experimental procedures were performed in
accordance with the Guidelines for the Care and Use of Laboratory
Animals published by Liaocheng People's Hospital, Liaocheng
Clinical School of Taishan Medical University (Liaocheng, China).
The study was approved by the Ethics Committee of Liaocheng
People's Hospital. Adult male Sprague-Dawley rats (n=40, weight,
220–250 g, 8–9 weeks-old) were purchased from the Laboratory Animal
Center of the Taishan Medical University, and raised in the
specific-pathogen-free Laboratory Animal Center at a constant
environment with a temperature of 22±1°C, 50–60% humidity, an
alternating 12 h light-dark cycle and free access to food/water.
The rats were randomly divided into four groups (n=10 rats per
group): Sham, SCI model, and two SCI + delphinidin-treated (40 or
200 mg/kg) groups. A total of 24 h after SCI, the delphinidin group
rats were treated with 40 or 200 mg/kg delphinidin (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) for 2 weeks.
SCI model
In order to establish an SCI model, the rats were
anesthetized with 35 mg/kg pentobarbital sodium by intraperitoneal
injection. The spinal cord was aseptically exposed following
laminectomy at T9-10. Next, a 2-mm diameter hammer was implanted at
25 mm from the T9-10 spinal cord and removed after 1 min.
Subsequently, manual bladder expression was performed three times
daily.
Basso, Beattie, Bresnahan (BBB) open
field locomotor assessment
Following delphinidin treatment, the BBB scores of
rats were assessed to evaluate the recovery condition of the motor
function. The BBB scores ranged between 0 and 21, with the lowest
point (score of 0) indicating complete paralysis and the maximum
point (score of 21) implying normal function.
ELISAs for determining the activities
of various inflammatory factors
Following delphinidin treatment, rats was sacrificed
using decapitation under anesthetization (35 mg/kg pentobarbital
sodium) and spinal cord (2 mm cephalic and caudally around the
injury epicenter) was removed. The tissues were dissolved in RIPA
lysis buffer (Beyotime Institute of Biotechnology, Jiangsu, China),
and a BCA kit (Beyotime Institute of Biotechnology) was used to
quantify the protein concentration. In accordance with the
specifications of the ELISA kits, protein samples (10 µg) were used
to measure the activities of tumor necrosis factor-α (TNF-α; cat.
no. H052), interleukin (IL)-6 (cat. no. H007), cyclooxygenase-2
(COX-2; cat. no. H200), prostaglandin E2 (PGE2; cat. no. H099) and
caspase-3 (cat. no. G015; all Jiancheng Bioengineering Institute,
Nanjing, China) using a luminometer (MicroLumat Plus LB 96V;
Berthold Technologies, Bad Wildbach, Germany).
Western blot analysis
Following delphinidin treatment, protein samples
were obtained from spinal cord tissues. The protein samples (40 µg)
were separated by 8–10% SDS-PAGE and transferred into
polyvinylidene difluoride membranes (EMD Millipore, Bedford, MA,
USA). The membranes were blocked in 5% fat-free milk for 1 h and
then incubated at 4°C overnight with primary antibodies, including
antibodies against nuclear factor-κB (NF-κB)/p65, activator protein
1 (AP-1), phosphorylated-p38-MAPK and β-actin. Subsequently, the
membranes were incubated with secondary antibodies for 2 h at room
temperature, and images were captured using ChemiDoc-It TS2 Imager
(UVP, LLC, Upland, CA, USA). The relative protein expression was
determined using ImageJ2× software (National Institutes of Health,
Bethesda, MD, USA).
Statistical analysis
Data are expressed as the mean ± standard deviation
and the Student's t-test was used for single statistical
comparisons using SPSS version 13.0 (SPSS, Inc., Chicago, IL, USA)
software package. P<0.05 was considered to indicate a
statistically significant difference.
Results
Anti-inflammatory effect of
delphinidin improves the BBB scores in the rat model of SCI
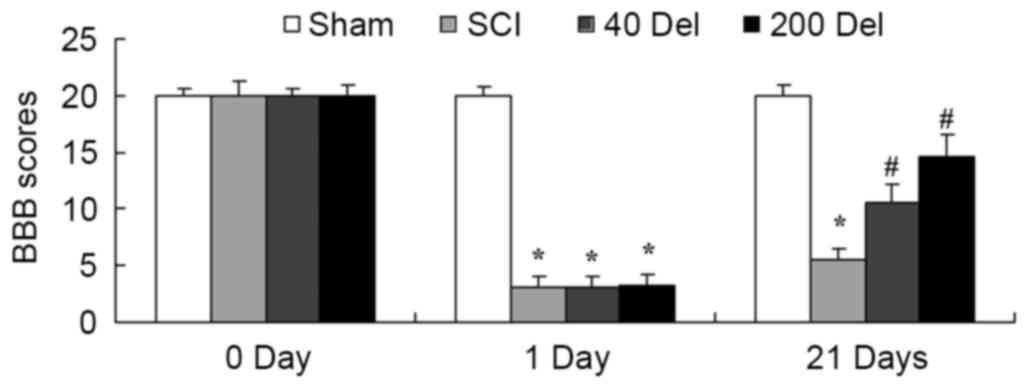
The present study investigated whether the
anti-inflammatory effect of delphinidin was able to improve the BBB
scores in a rat model of SCI. As shown in Fig. 2, BBB scores in the SCI model group
were markedly reduced compared with those of the sham group
(P=0.0023). However, treatment with 40 or 200 mg/kg delphinidin
significantly increased the SCI-induced inhibition of BBB scores at
21 days after SCI (Fig. 2; P=0.0079
or 0.0056, respectively).
Anti-inflammatory effect of
delphinidin alleviates the intramedullary spinal pressure in the
SCI rat model
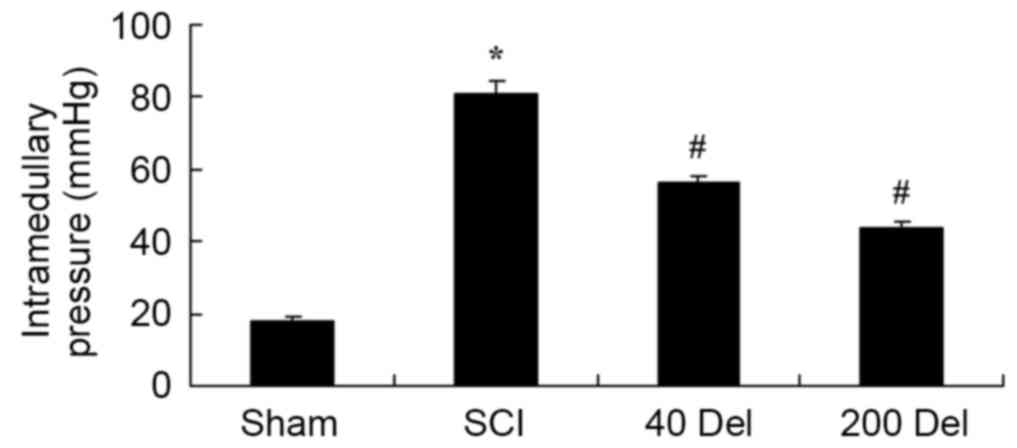
The effect of delphinidin treatment on the
intramedullary spinal pressure in SCI rats was investigated. There
was a significant increase in the intramedullary spinal pressure in
SCI rats when compared with the sham group (Fig. 3; P=0.0037). However, 40 or 200 mg/kg
delphinidin treatment significantly reduced the induction of
intramedullary spinal pressure when compared with the untreated SCI
rats (Fig. 3; P=0.0072 or 0.0044,
respectively).
Delphinidin treatment reduces the
activities of inflammatory factors in the rat model of SCI
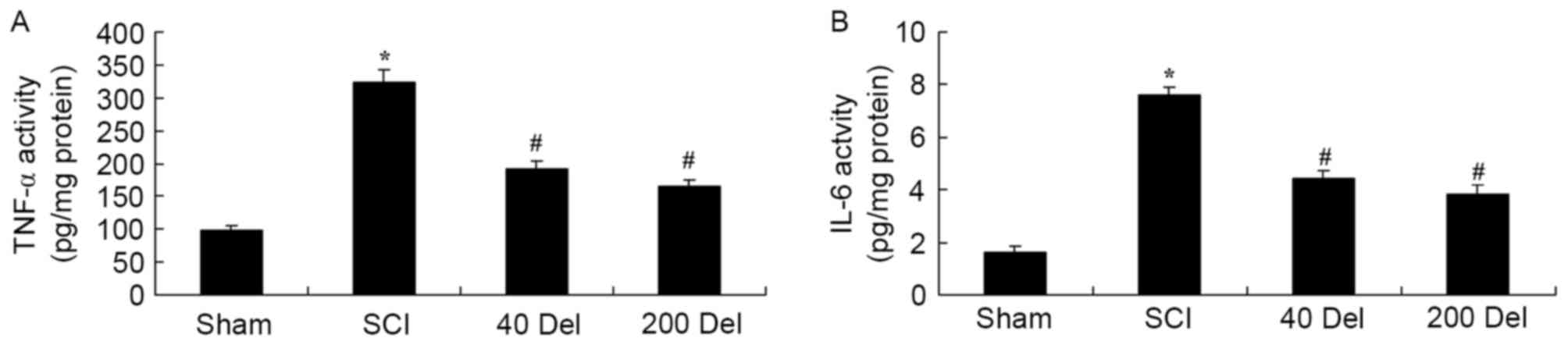
To examine the anti-inflammatory effect of
delphinidin in SCI rats, the activities of anti-inflammatory
factors TNF-α and IL-6 were analyzed by ELISA. The TNF-α and IL-6
activities were found to be markedly increased in SCI rats,
compared with the sham group (Fig.
4; P=0.0021 or 0.0014, respectively). However, 40 or 200 mg/kg
delphinidin treatment significantly suppressed the promotion of
TNF-α and IL-6 activities in the SCI rats (Fig. 4; P=0.0055 or 0.0038 and P=0.0051 or
0.0024, respectively). Taken together, these results indicated
delphinidin has an anti-inflammatory effect in SCI.
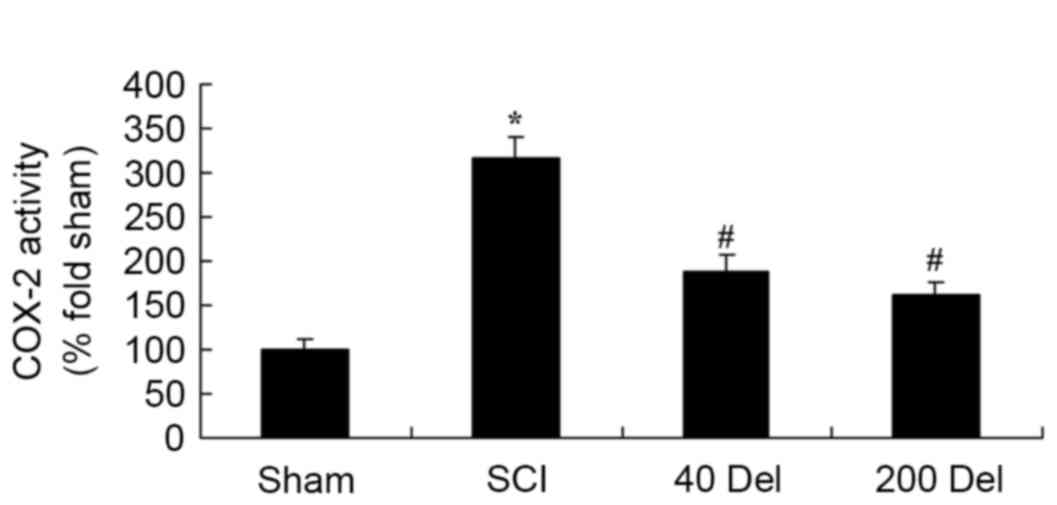
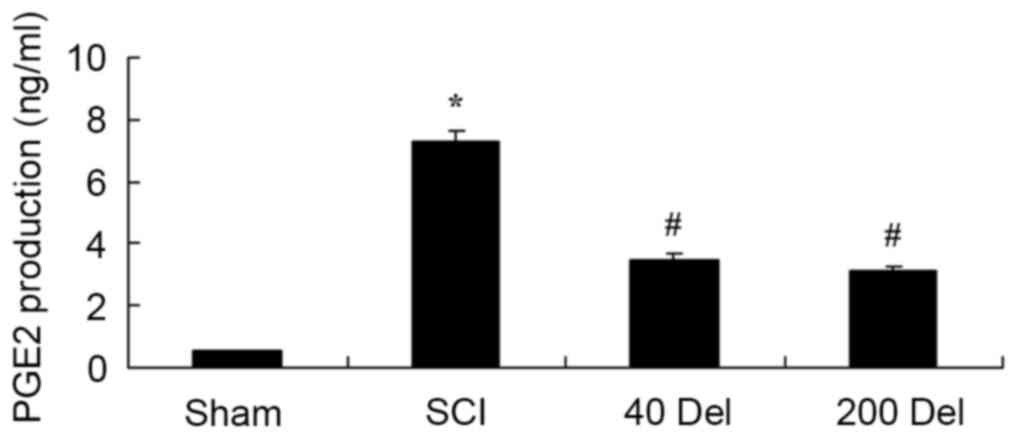
Delphinidin inhibits the COX-2
activity and PGE2 production in the rat model of SCI
In order to investigate the anti-inflammatory effect
of delphinidin in SCI rats, the COX-2 and PGE2 levels were
investigated by ELISA. The induction of SCI in the rats resulted in
enhanced COX-2 activity (Fig. 5;
P=0.0025) and PGE2 production (Fig.
6; P=0.0012) when compared with the levels in the sham group.
By contrast, upon treatment with different concentrations of
delphinidin for 2 weeks, the activity of COX-2 in the SCI rats was
significantly inhibited when compared with the untreated SCI model
rats (Fig. 5). In addition, 40 or
200 mg/kg delphinidin treatment markedly inhibited the activation
of PGE2 production in SCI rats (Fig.
6; P=0.0031 or 0.0026, respectively).
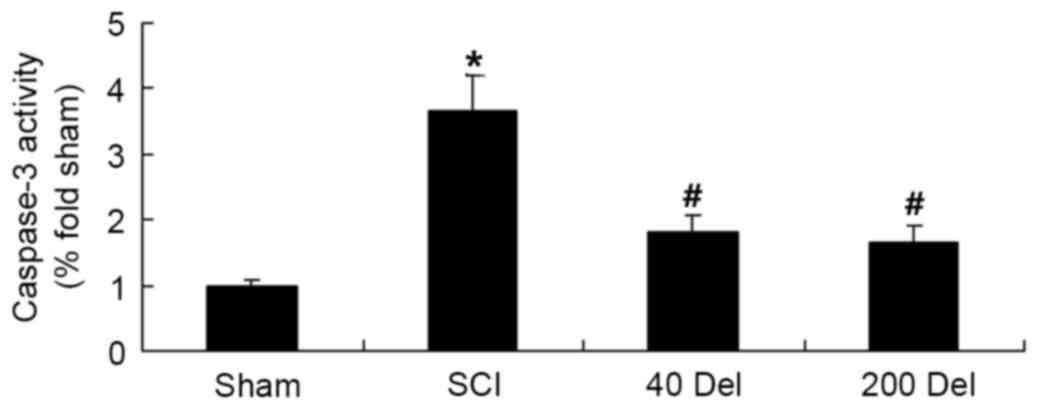
Delphinidin treatment reduces
caspase-3 activity in the rat model of SCI
To further determine the effect of delphinidin
treatment on apoptosis in the SCI rats, caspase-3 activity was
detected in the present study using ELISA. As shown in Fig. 7, the caspase-3 activity was evidently
activated in SCI rats compared with the sham group (P=0.0049).
However, 40 or 200 mg/kg delphinidin treatment significantly
inhibited the caspase-3 activity induced by SCI in the rat model of
SCI (Fig. 7; P=0.0058 or 0.0045,
respectively).
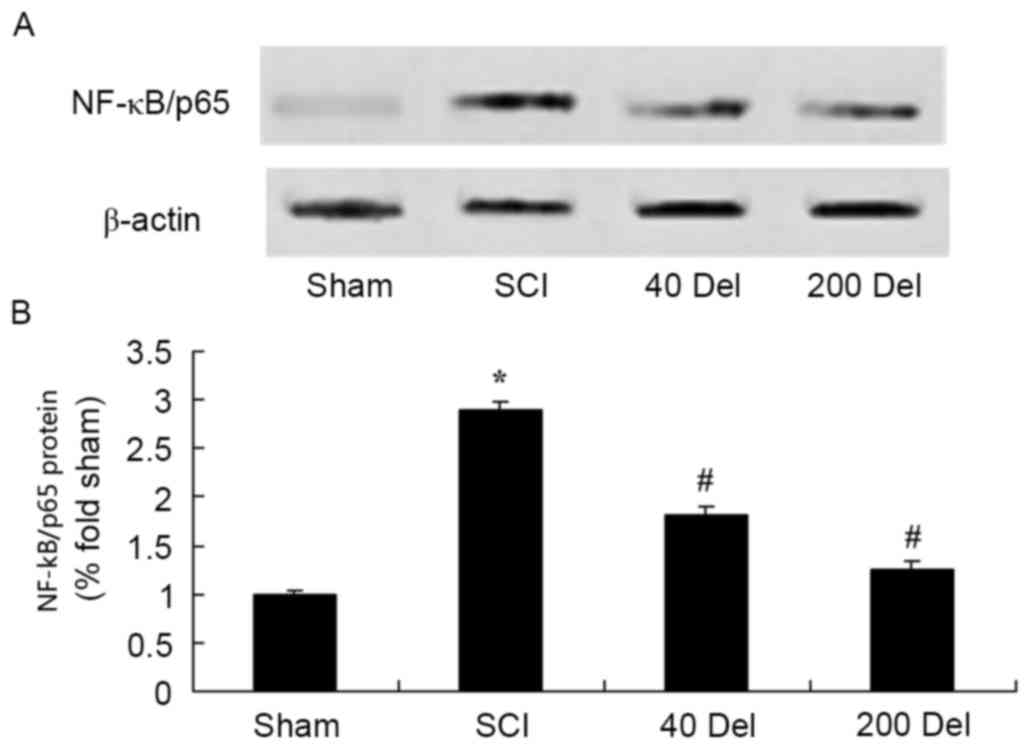
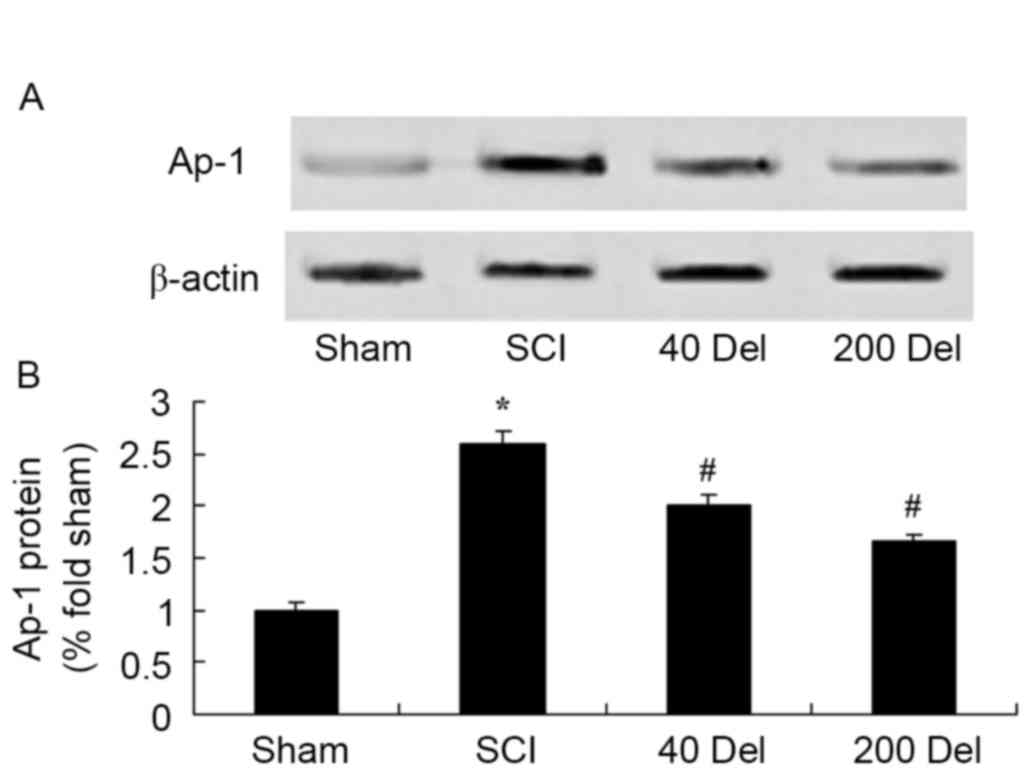
Delphinidin reduces NF-κB and Ap-1
protein levels in a rat model of SCI
To further investigate the anti-inflammatory effect
of delphinidin in the rat model of SCI, NF-κB/p65 and Ap-1 protein
expression levels were detected using western blot analysis. As
shown in Figs. 8 and 9, the induction of SCI evidently increased
the NF-κB/p65 (P=0.0012 or 0.0009, respectively) and Ap-1 protein
expression levels when compared with those in the sham group rats.
However, 40 or 200 mg/kg delphinidin treatment significantly
inhibited the protein expression levels of NF-κB/p65 (Fig. 8; P=0.0044 or 0.0032, respectively)
and Ap-1 (Fig. 9; P=0.0052 or
0.0041) in SCI rats (Fig. 8).
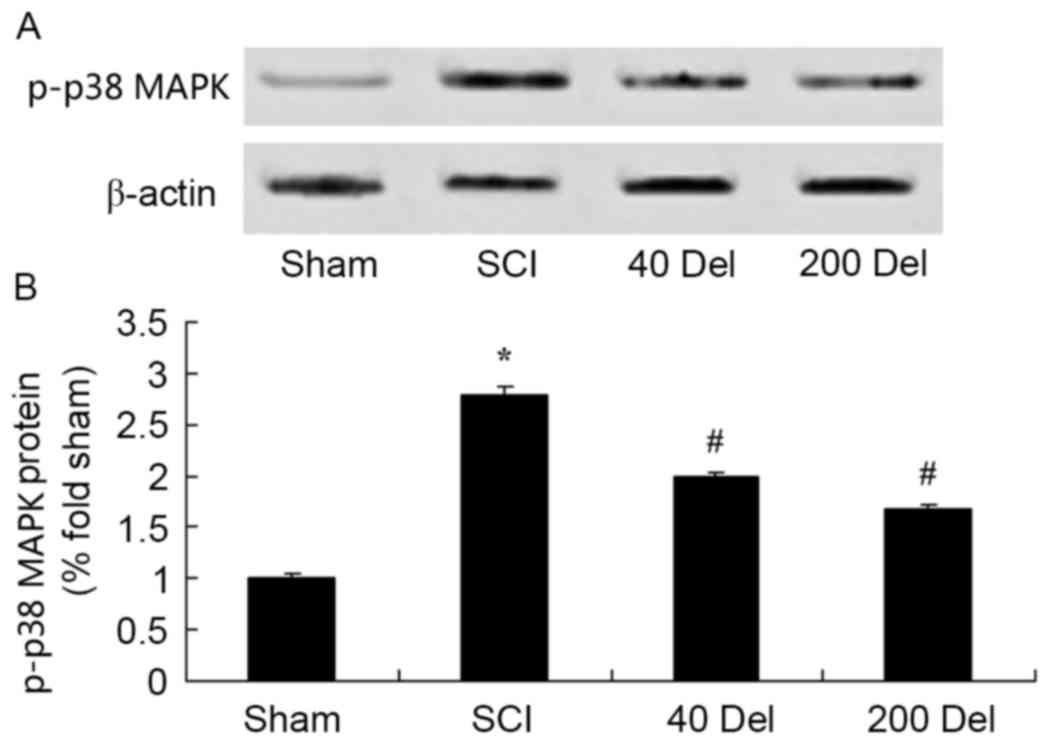
Anti-inflammatory effect of
delphinidin blocks the activation of the p38-MAPK signaling pathway
in the rat model of SCI
To further study the molecular and cellular effects
of delphinidin on SCI, the expression of phosphorylated p38-MAPK in
the rat model of SCI was detected. At 2 weeks after SCI, the
p-p38-MAPK protein expression in the SCI model group was markedly
higher in comparison with that of the sham group (Fig. 10; P=0.0029). However, treatment with
40 or 200 mg/kg delphinidin significantly suppressed the
SCI-induced activation of p-p38-MAPK protein expression in SCI rats
(Fig. 10; P=0.0047 or 0.0035,
respectively).
Discussion
Acute SCI is a trauma that may result in disability
(2). One of the most important
targets of early treatment for SCI is to prevent the secondary
injury. The pathological development process of secondary injury in
the spinal cord involves capillary changes, edema, altered energy
metabolism, various biochemical changes and apoptosis (14). Inhibiting the inflammatory reaction
and lipid peroxidation is also regarded as one of the most
important targets of early treatment for SCI (15). Following SCI, the early inflammatory
reaction serves a role in protecting the spinal cord nerves and
promoting functional recovery (16).
For these reasons, strategies for relieving the inflammatory
reaction after SCI, and promoting structural reconstruction and
functional recovery of the damaged areas is the focus of research
in the fields of neuroscience, orthopedics and sports medicine
(17,18). In the present study, delphinidin
treatment significantly increased the SCI-inhibited BBB scores and
reduced the induction of intramedullary spinal pressure in SCI
rats. These findings suggest that delphinidin may be used to treat
SCI and to reduce intramedullary spinal pressure.
The inflammatory reaction is mainly regulated by the
expression of several genes, while the NF-κB family is the major
regulatory factor of inflammatory gene expression (7,19). NF-κB
can regulate expression of many cellular factors and regulate
inflammatory reactions. Abnormally activated NF-κB may launch the
apoptosis of neurons in trauma of central nervous system,
excitotoxic injury, ischemic damage and neurodegenerative disease
(12). NF-κB is abnormally activated
following SCI, activating inducible nitric oxide synthase in
neurons (7). The expression of
various genes in SCI is regulated by NF-κB, including the
expression of pro-inflammatory cell factors, such as TNF-α, IL-1β
and IL-6, as well as proteases, such as matrix metalloproteinase
(7). Pal et al (12) reported that delphinidin reduces
psoriasiform lesions by inducing epidermal differentiation and
inhibiting inflammation in flaky skin mice. Cho et al
(9) suggested that delphinidin
inhibited extracellular matrix production in nasal polyp-derived
fibroblasts through the MAPK/NF-κB signaling pathway induced by
TGF-β1 stimulation. In addition, the molecular mechanism of
delphinidin regulating SCI-induced inflammation through the
suppression of NF-κB after SCI was initially observed in the
present study.
At present, TGF-β1 is increasingly investigated due
to its association with the majority of fiber hyperplastic diseases
(20). In addition, TGF-β1 is
recognized as one of the most powerful cell factors that cause
fibrosis (21). The present study
results revealed that delphinidin significantly inhibited the
activation of PGE2 production, and suppressed the protein
expression levels of AP-1 and p38-MAPK in SCI rats. This data
suggests that delphinidin prevents SCI through regulation of PGE2
production, AP-1 and p38-MAPK expression.
COX-2 is typically considered to be harmful under
pathological conditions, such as inflammation, pain, cellular
damage and tumor. However, recent studies have proven that COX-2
also has an effect on the normal physiological function of humans
(22). In the central nervous
system, COX-2 is expressed in the glutamate neurons of the
hippocampus and cerebral cortex, and is critical to neurovascular
coupling during the period of synaptic activity, long-term synaptic
plasticity and functional congestion (23). Previous data indicated that COX-2
expression is associated with hypoxia, peroxidation and neuronal
cell death induced by excitatory amino acids after SCI (24). The application of COX-2 inhibitors
subsequent to SCI serves a certain neuroprotective effect,
indicating that COX-2 is harmful after SCI (25). Hwang et al (26) suggested that delphinidin inhibited
COX-2 expression via TNF-α in JB6 P+ mouse epidermal cells. In the
present study, delphinidin treatment significantly reduced COX-2
activity in SCI rats.
It is known that p-p38-MAPK participates in signal
transduction in the process of nerve cell apoptosis. Thus, strong
or weak expression of p-p38 predicts whether nerve cells are alive
(27). As a result of the apoptosis
process of neurons and colloid cells subsequent to SCI, p-p38
expression increases, indicating that the p-p38 signal transduction
pathway is altered after SCI (28).
SCI results in the activation of p-p38, which then increases NOS
activity and promotes apoptosis (29). The results of the present study
demonstrated that the anti-inflammatory effect of delphinidin
reduced the p-p38-MAPK protein expression and caspase-3 activity in
a rat model of SCI. Furthermore, Oak et al (30) reported that delphinidin prevents the
activation of p38-MAPK and JNK in vascular smooth muscle cells.
Overall, these data suggest that administration of delphinidin
alleviates the SCI-induced inflammation through suppression of the
p38-MAPK signaling pathway.
In conclusion, the anti-inflammatory effect of
delphinidin was able to alleviate the SCI-induced inflammation and
intramedullary spinal pressure in a rat model of SCI. The present
study confirms that the anti-inflammatory effect of delphinidin was
achieved in SCI rats through theinhibition of COX-2 and PGE2
production, and the suppression of NF-κB and p38 MAPK signaling
pathways.
References
|
1
|
de Rivero Vaccari JP, Marcillo A, Nonner
D, Dietrich WD and Keane RW: Neuroprotective effects of bone
morphogenetic protein 7 (BMP7) treatment after spinal cord injury.
Neurosci Lett. 465:226–229. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lim JH, Muguet-Chanoit AC, Smith DT, Laber
E and Olby NJ: Potassium channel antagonists 4-aminopyridine and
the T-butyl carbamate derivative of 4-aminopyridine improve hind
limb function in chronically non-ambulatory dogs; a blinded,
placebo-controlled trial. PLoS One. 9:e1161392014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DePaul MA, Palmer M, Lang BT, Cutrone R,
Tran AP, Madalena KM, Bogaerts A, Hamilton JA, Deans RJ, Mays RW,
et al: Intravenous multipotent adult progenitor cell treatment
decreases inflammation leading to functional recovery following
spinal cord injury. Sci Rep. 5:167952015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Galluzzi F, De Rensis F, Saleri R and
Spattini G: Effect of urethral infusion of atracurium besylate on
manual bladder expression in dogs and cats with spinal cord
injuries: A randomised trial. Vet Rec. 176:5452015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wilson JR, Grossman RG, Frankowski RF,
Kiss A, Davis AM, Kulkarni AV, Harrop JS, Aarabi B, Vaccaro A,
Tator CH, et al: A clinical prediction model for long-term
functional outcome after traumatic spinal cord injury based on
acute clinical and imaging factors. J Neurotrauma. 29:2263–2271.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rosety-Rodriguez M, Camacho A, Rosety I,
Fornieles G, Rosety MA, Diaz AJ, Bernardi M, Rosety M and Ordonez
FJ: Low-grade systemic inflammation and leptin levels were improved
by arm cranking exercise in adults with chronic spinal cord injury.
Arch Phys Med Rehabil. 95:297–302. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oudega M: Inflammatory response after
spinal cord injury. Exp Neurol. 250:151–155. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fan QQ, Li L, Wang WT, Yang X, Suo ZW and
Hu XD: Activation of α2 adrenoceptors inhibited NMDA
receptor-mediated nociceptive transmission in spinal dorsal horn of
mice with inflammatory pain. Neuropharmacology. 77:185–192. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cho JS, Kang JH, Shin JM, Park IH and Lee
HM: Inhibitory effect of delphinidin on extracellular matrix
production via the MAPK/NF-κB pathway in nasal polyp-derived
fibroblasts. Allergy Asthma Immunol Res. 7:276–282. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dayoub O, Andriantsitohaina R and Clere N:
Pleiotropic beneficial effects of epigallocatechin gallate,
quercetin and delphinidin on cardiovascular diseases associated
with endothelial dysfunction. Cardiovasc Hematol Agents Med Chem.
11:249–264. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yuan B, Okusumi S, Yoshino Y, Moriyama C,
Tanaka S, Hirano T, Takagi N and Toyoda H: Delphinidin induces
cytotoxicity and potentiates cytocidal effect in combination with
arsenite in an acute promyelocytic leukemia NB4 cell line. Oncol
Rep. 34:431–438. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pal HC, Chamcheu JC, Adhami VM, Wood GS,
Elmets CA, Mukhtar H and Afaq F: Topical application of delphinidin
reduces psoriasiform lesions in the flaky skin mouse model by
inducing epidermal differentiation and inhibiting inflammation. Br
J Dermatol. 172:354–364. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hafeez BB, Siddiqui IA, Asim M, Malik A,
Afaq F, Adhami VM, Saleem M, Din M and Mukhtar H: A dietary
anthocyanidin delphinidin induces apoptosis of human prostate
cancer PC3 cells in vitro and in vivo: Involvement of nuclear
factor-kappaB signaling. Cancer Res. 68:8564–8572. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sutbeyaz ST, Koseoglu BF and Gokkaya NK:
The combined effects of controlled breathing techniques and
ventilatory and upper extremity muscle exercise on cardiopulmonary
responses in patients with spinal cord injury. Int J Rehabil Res.
28:273–276. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schulz R, Czaja SJ, Lustig A, Zdaniuk B,
Martire LM and Perdomo D: Improving the quality of life of
caregivers of persons with spinal cord injury: A randomized
controlled trial. Rehabil Psychol. 54:1–15. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Whiteneck GG, Gassaway J, Dijkers MP,
Lammertse DP, Hammond F, Heinemann AW, Backus D, Charlifue S,
Ballard PH and Zanca JM: Inpatient and postdischarge rehabilitation
services provided in the first year after spinal cord injury:
Findings from the SCIRehab study. Arch Phys Med Rehabil.
92:361–368. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ellenbroek D, Kressler J, Cowan RE, Burns
PA, Mendez AJ and Nash MS: Effects of prandial challenge on
triglyceridemia, glycemia, and pro-inflammatory activity in persons
with chronic paraplegia. J Spinal Cord Med. 38:468–475. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jorgensen V, Elfving B and Opheim A:
Assessment of unsupported sitting in patients with spinal cord
injury. Spinal Cord. 49:838–843. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tyagi P, Kadekawa K, Kashyap M, Pore S and
Yoshimura N: Spontaneous recovery of reflex voiding following
spinal cord injury mediated by anti-inflammatory and
neuroprotective factors. Urology. 88:57–65. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ji H, Tang H, Lin H, Mao J, Gao L, Liu J
and Wu T: Rho/Rock cross-talks with transforming growth
factor-β/Smad pathway participates in lung fibroblast-myofibroblast
differentiation. Biomed Rep. 2:787–792. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rathore KI, Redensek A and David S: Iron
homeostasis in astrocytes and microglia is differentially regulated
by TNF-α and TGF-β1. Glia. 60:738–750. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Banovac K, Williams JM, Patrick LD and
Levi A: Prevention of heterotopic ossification after spinal cord
injury with COX-2 selective inhibitor (rofecoxib). Spinal Cord.
42:707–710. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang G, Huang C, Wang Y, Guo Q, Jiang H
and Wen J: Changes in expression of cyclooxygenase-2 in the spinal
dorsal horn after intrathecal p38MAPK inhibitor SB203580 on
neuropathic pain in rats. Ann Palliat Med. 2:124–129.
2013.PubMed/NCBI
|
|
24
|
Quan HH, Kang KS, Sohn YK and Li M: Tempol
reduces injury area in rat model of spinal cord contusion injury
through suppression of iNOS and COX-2 expression. Neurol Sci.
34:1621–1628. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Coronel MF, Labombarda F, De Nicola AF and
González SL: Progesterone reduces the expression of spinal
cyclooxygenase-2 and inducible nitric oxide synthase and prevents
allodynia in a rat model of central neuropathic pain. Eur J Pain.
18:348–359. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hwang MK, Kang NJ, Heo YS, Lee KW and Lee
HJ: Fyn kinase is a direct molecular target of delphinidin for the
inhibition of cyclooxygenase-2 expression induced by tumor necrosis
factor-alpha. Biochem Pharmacol. 77:1213–1222. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He BR, Xie ST, Wu MM, Hao DJ and Yang H:
Phagocytic removal of neuronal debris by olfactory ensheathing
cells enhances neuronal survival and neurite outgrowth via p38MAPK
activity. Mol Neurobiol. 49:1501–1512. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Taves S, Berta T, Liu DL, Gan S, Chen G,
Kim YH, Van de Ven T, Laufer S and Ji RR: Spinal inhibition of p38
MAP kinase reduces inflammatory and neuropathic pain in male but
not female mice: Sex-dependent microglial signaling in the spinal
cord. Brain Behav Immun. 55:70–81. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gwak YS, Unabia GC and Hulsebosch CE:
Activation of P-38alpha MAPK contributes to neuronal
hyperexcitability in caudal regions remote from spinal cord injury.
Exp Neurol. 220:154–161. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oak MH, Bedoui JE, Madeira SV, Chalupsky K
and Schini-Kerth VB: Delphinidin and cyanidin inhibit
PDGF(AB)-induced VEGF release in vascular smooth muscle cells by
preventing activation of p38 MAPK and JNK. Br J Pharmacol.
149:283–290. 2006. View Article : Google Scholar : PubMed/NCBI
|