Introduction
Bladder cancer is one of the most common urinary
malignancies worldwide and has a high recurrence rate. In the
United States, the numbers of new cases and deaths due to bladder
cancer in 2019 were reported to be 80,470 and 17,670, respectively
(1). Muscle-invasive bladder
cancers represent 30% of all cases and are associated with a poor
prognosis (2). Despite the recent
progress in the treatment of bladder cancer, including surgery,
radiation therapy, chemotherapy and immunotherapy, the prognosis of
patients remains unsatisfactory, particularly for muscle-invasive
and metastatic bladder cancer (3-5).
The prognosis of bladder cancer is closely associated with the
tumor stage; however, patients do not exhibit specific symptoms at
the early stage (6). Therefore,
identifying potential early detection markers is important for
effective treatment.
Long non-coding (lnc)RNA represents a type of
non-coding RNA >200 nucleotides long that lacks protein-coding
capacity (7). To date, lncRNAs have
been demonstrated to be associated with various intracellular and
extracellular activities, such as gene transcription, mRNA splicing
and tumorigenesis (8). In our
previous studies, dysregulated lncRNAs, including growth
arrest-specific 5 and sprouty4-intron 1, have been demonstrated to
serve crucial roles in the development and progression of bladder
cancer (9,10). However, the roles and the underlying
mechanisms of the action of lncRNAs in the progression of bladder
cancer remain unknown.
The prostate cancer-associated transcript 6 (PCAT6),
a 1.0-kb intergenic lncRNA located at chromosome 1q32.1, was first
detected in cervical cancer cells (11) and has been reported to be
responsible for the regulation of radiosensitivity, cell
proliferation and chemoresistance in various types of cancer,
including breast (12), gastric
(13) and cervical (14) cancer. However, the functions
performed by PCAT6 in bladder cancer have not been reported to
date. Therefore, the present study aimed to clarify the role of
PCAT6 in the progression of bladder cancer and the potential
underlying mechanism.
Materials and methods
Tissue specimens
A total of 32 pairs of bladder cancer and adjacent
normal tissues (3 cm from the tumor margin) were obtained from
patients who underwent radical cystectomy (age range, 41-84 years
old; mean age, 62.1±11.7 years) at Wuhan Central Hospital (Wuhan,
China) between January 2016 and September 2019. The patient
clinical data are presented in Table
I. Patients who had been treated with chemotherapy, laser
therapy, cryotherapy or other anticancer modalities prior to the
surgical procedure were excluded from the study. The specimens were
classified according to the 2004 World Health Organization
Consensus Classification and Staging System (15). The research protocol was approved by
the Ethics Committee of the Central Hospital of Wuhan, and written
informed consent was obtained from each patient before enrollment.
The expression of PCAT6 in bladder cancer tissues was analyzed by
Gene Expression Profiling Interactive Analysis (GEPIA 1.0;
https://gepia.cancer-pku.cn/index.html) tools based on
public data from the TCGA database (16).
 | Table IAssociations between PCAT6 expression
levels and the clinicopathological characteristics of patients with
bladder cancer. |
Table I
Associations between PCAT6 expression
levels and the clinicopathological characteristics of patients with
bladder cancer.
| | Expression of
PCAT6, n | |
|---|
| Parameter | Cases, n | Low | High | P-value |
|---|
| Sex | | | | 0.3702 |
|
Male | 23 | 19 | 4 | |
|
Female | 9 | 6 | 3 | |
| Age | | | | 0.6833 |
|
<55 | 12 | 10 | 2 | |
|
≥55 | 20 | 15 | 5 | |
| Pathological
stage | | | | 0.0101 |
|
pTa-T1 | 23 | 21 | 2 | |
|
pT2-T4 | 9 | 4 | 5 | |
| Grade | | | | 0.6317 |
|
Low | 7 | 5 | 2 | |
|
High | 25 | 20 | 5 | |
| Total | 32 | 25 | 7 | |
Cell transfection
The human metastatic bladder cancer cell line T24T,
which is a lineage-related lung metastatic variant of invasive
bladder cancer cell line T24, was provided by Dr Dan Theodorescu
(Departments of Urology, University of Virginia, Charlottesville,
VA, USA) in 2010 and maintained at our laboratory as previously
described (17-19).
The cell line was subjected to DNA tests and authenticated in our
previous study (20). Human bladder
cancer cell lines EJ, UMUC3, 5637 and the human immortalized
uroepithelium cell line (SV-HUC-1) were purchased from ATCC in 2015
and maintained at our laboratory. Cells were cultured in RPMI-1640
medium (HyClone; Cytiva) containing 10% fetal bovine serum
(HyClone; Cytiva) in humidified air containing 5% CO2 at
37˚C. Small interfering (si)RNAs designed to target PCAT6
(siPCAT6), negative control siRNA (siNC), miR-143-3p mimic
(miR-143-3p), miRNA mimic negative control (mimic-NC), miR-143-3p
inhibitor (anti-miR143) and control inhibitor (control) were
obtained from Guangzhou RiboBio Co., Ltd.
When the cell confluence reached 80%, a total of 100
nM siPCAT6, 100 nM siNC, 50 nM miR-143-3p mimics, 50 nM anti-miR143
or 50 nM of their corresponding negative controls were transfected
into cells at 37˚C for 24 h using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) respectively. The
sequences were as follows: siPCAT6, 5'-UGCAGCUCCGCUAUGGCCU-3';
siNC, 5'-UUCUCCGAACGUGUCACGUTT-3'; miR-143-3p,
5'-UGAGAUGAAGCACUGUAGCUC-3'; mimic-NC, 5'-UUCUCCGAACGUGUCACGUTT-3';
anti-miR143, 5'-GAGCUACAGUGCUUCAUCUCA-3'; inhibitor control,
5'-CAGUACUUUUGUGUAGUA-3'. After 48 h incubation at 37˚C,
transfected cells were harvested and utilized for further
experiments. The transfection efficiency was determined by
RT-qPCR.
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
Total RNA was extracted from tissues and cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and reverse-transcribed to cDNA using the Prime Script™ RT
Reagent kit (cat. no. RR037A; Takara Biotechnology Co., Ltd.). The
temperature protocol using for RT was as follows: 37˚C for 15 min
and 85˚C for 5 sec. For microRNA (miRNA) reverse transcription,
miRNA First-Strand cDNA Synthesis kit (cat. no. B532451; Sangon
Biotech Co., Ltd.) was used according to the manufacturer's
instructions. qPCR was performed using SYBR® Premix Ex
Taq (cat. no. RR041A; Takara Biotechnology Co., Ltd.) on the
StepOnePlus Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The thermocycling conditions were as follows:
Initial denaturation for 30 sec at 95˚C, followed by 40 cycles for
5 sec at 95˚C and 35 sec at 60˚C. GAPDH was used as an endogenous
control for lncRNAs and mRNAs. The expression levels of miRNA were
normalized to those of U6. The primers used were as follows: PCAT6
forward, 5'-CCCCTCCTTACTCTTGGACAAC-3' and reverse,
5'-GACCGAATGAGGATGGAGACAC-3'; miR-143-3p forward,
5'-GGGGTGAGATGAAGCACTG-3' and reverse, 5'-CAGTGCGTGTCGTGGAGT-3';
PDIA6 forward, 5'-GGAGGTCAGTATGGTGTTCAGGGAT-3'; and reverse,
5'-CTGCCACCTTGGTAATCTTCTGGTC-3'; GAPDH forward,
5'-TGCACCACCAACTGCTTAGC-3' and reverse,
5'-GGCATGGACTGTGGTCATGAG-3'; U6 forward, 5'-CTCGCTTCGGCAGCACA-3'
and reverse, 5'-AACGCTTCACGAATTTGCGT-3'.
Fluorescence in situ
hybridization
Cy3-labeled mRNA probes for PCAT6 were designed and
synthesized using cDNA as a template by Guangzhou RiboBio Co., Ltd
(cat. no. lnc1cm001). Total RNA was isolated with RNAiso Plus (cat.
no. 9109; Takara Biotechnology Co., Ltd.) from T24T cells and then
reverse transcribed into cDNA with PrimeScript™ RT reagent Kit
(cat. no. RR037A; Takara Biotechnology Co., Ltd.) as
aforementioned. The probe sequence was is not publicly available.
T24T and EJ cells were fixed in 4% paraformaldehyde for 30 min at
37˚C and incubated with 0.1% Triton X-100 on ice for 10 min. In
total, 2.5 µl PCAT6 probes (20 µM) were hybridized with the cells
for 5 h in the dark at 37˚C. Sodium citrate buffer was then added
to wash the cell for 5 min at 42˚C for three times in the dark. 10
µl DAPI (5 µg/ml) was used to stain the nucleus for 10 min in the
dark at room temperature. After washed with PBS for three times,
images were captured using a Nikon A1Si Laser Scanning Confocal
Microscope (magnification, x400; Nikon Instruments, Inc.).
MTT assay
The proliferative capacity of bladder cancer cells
was evaluated by the MTT assay. At 48 h post-transfection, T24T and
EJ cells were plated into 96-well plates at a density of 2,000
cells/well and incubated for 24, 48, 72 or 96 h. Subsequently, 20
µl MTT (5 mg/ml; EMD Millipore) was added in each well and cultured
for 4 h. The culture medium was removed, and 150 µl DMSO was to
dissolve the crystals. The absorbance was measured at 570 nm using
a microplate reader.
5-Ethynyl-2'-deoxyuridine (EdU)
assay
Transfected T24T and EJ cells (1x105
cells/well) were seeded in 96-well plates. EdU (100 mmol/l;
Guangzhou RiboBio Co., Ltd.) was added to the medium and incubated
for 2 h at room temperature in the dark. Subsequently, the DNA
contents of the cells were stained with 100 µl Hoechst 33342 (2
µg/ml; Guangzhou RiboBio Co., Ltd.) for 30 min at room temperature
in the dark. Images were captured with a fluorescent microscope
(magnification, x200; Olympus Corporation). The ratio of
EdU-stained cells to Hoechst-stained cells was calculated to
evaluate the cell proliferation.
Wound healing assay
At 48 h post-transfection, T24T and EJ cells were
seeded in a 6-well culture dish (5x105 cells/well) in
RPMI-1640 medium (HyClone; Cytiva) containing 10% FBS (HyClone;
Cytiva) at 37˚C. When the cell confluence reached 90%, cells were
mechanically wounded by passing a sterile 200 µl plastic pipette
tip through the monolayer with one stroke and incubated at 37˚C in
serum-free medium. The cells were washed with PBS to wash away the
floating cells. The wound healing processes were observed under a
light microscope (magnification, x100; Olympus Corporation) at 0
and 48 h after the scratch, and the distance was analyzed with
ImageJ software (National Institutes of Health). The relative
migration rate (%)=(wound width at 0 h-wound width at 48 h)/wound
width at 0 h x100.
Cell invasion and migration assay
Transwell chambers uncoated or coated with Matrigel
(cat. no. 353097 for migration and cat. no. 354480 for invasion;
8-µm pore size filter; BD Biosciences) were used to determine the
cell migratory or invasive abilities, respectively. Transfected
T24T and EJ cells were seeded in the upper chamber of the Transwell
insert with serum-free medium at a density of 1ⅹ104
cells/well. Medium supplemented with 10% fetal bovine serum was
added to the lower chamber. Following 24-h incubation at 37˚C with
5% CO2, cells on the upper surface of the membrane were
removed with a cotton swab. The migratory cells in the lower
chamber were fixed with 4% paraformaldehyde for 15 min at room
temperature and stained for 15 min at room temperature using 0.1%
crystal violet. The numbers of migrated and invasive cells were
counted under a light microscope (magnification, x100; Olympus
Corporation) and the cell numbers were counted in five random
fields of view.
Dual-luciferase reporter assay
The binding sites between miR-143-3p and PCAT6 were
predicted and analyzed using Starbase 2.0 (https://starbase.sysu.edu.cn/) and Lncbase v.2
(https://carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=site%2Findex).
The binding sites between miR-143-3p and PDIA6 3'-untranslated
regions (UTR) were predicted by TargetScan 7.1 (https://www.targetscan.org/vert_71/). The
sequences of PCAT6 and PDIA6 3'UTR containing the potential
wild-type (WT) or mutant (MUT) binding sites of miR-143-3p were
synthesized by Tsingke Biological Technology Co., Ltd. and then
inserted into the pGL3 vector (Guangzhou RiboBio Co., Ltd.) to
construct the luciferase reporter. The T24T and EJ cells were
seeded at a density of 1x104 into 96-well plates. The
luciferase reporter (0.1 µg) and miR-143-3p mimics (40 nM) or
mimic-NC (40 nM) were co-transfected into cells using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After 48 h of incubation at 37˚C, the
transfected cells were harvested and assayed for luciferase
activity using the dual-luciferase reporter assay system (Promega
Corporation). Firefly luciferase activity was normalized to
Renilla luciferase activity.
RNA immunoprecipitation (RIP)
assay
The EZMagna RIP kit (cat. no. 17-701; EMD Millipore)
was used to evaluate the target relationship between miR143-3p and
PCAT6. T24T and EJ cells were harvested, resuspended with RIP lysis
buffer supplemented with RNase Inhibitor (Promega Corporation) on
ice for 5 min and centrifuged at 22,000 x g at 4˚C for 10 min. In
total, 40 µl protein A/G beads and 5 µg human anti-Ago2 antibody
(cat. no. ab32381; Abcam) or 5 µg negative control normal IgG (cat.
no. ab188776; Abcam) were incubated at 4˚C for 8 h in 900 µl RIP
buffer, before 2 mg total protein in 100 µl supernatant was added
and incubated at 4˚C overnight. After brief centrifugation at 1,000
x g for 2 min at 4˚C, the samples were placed on a magnetic rack
for 30 min at 4˚C. The supernatant was discarded and 500 µl RIP
wash buffer was added to resuspend the beads, which was repeated
five times. The supernatant was removed and samples were treated
with proteinase K (cat. no. ST532; Beyotime Institute of
Biotechnology.) on a shaker at 58˚C for 30 min. After
centrifugation at 1,000 x g for 5 min at 4˚C, the supernatant was
collected and 250 µl RIP buffer was added. A total of 400 µl
mixture (phenol: Chloroform: Isoamyl alcohol, 125:24:1) was added
to isolate the immunoprecipitated RNAs and the purified RNAs were
subjected to RT-qPCR analysis.
Western blot assay
T24T and EJ cells were collected at 48 h
post-transfection and resuspended in RIPA lysis buffer (Beyotime
Institute of Biotechnology). The concentration of protein was
determined using a bicinchoninic acid Protein Assay kit (Beyotime
Institute of Biotechnology). In total, 50 µg of each protein
samples were separated by 10% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes (EMD Millipore). Following
blocking with 5% skimmed milk (Beyotime Institute of Biotechnology)
for 2 h at room temperature, the membranes were incubated with
primary antibodies against PDIA6 (1:2,000; cat. no. ab227545;
Abcam) or GAPDH (1:5,000; cat. no. ab9485; Abcam) at 4˚C overnight.
After being washed three times in TBS-T (0.1% Tween-20), the
membrane was incubated with the HRP-conjugated Goat Anti-Rabbit IgG
H&L secondary antibody (1:10,000; cat. no. ab97051; Abcam) at
room temperature for 1 h before developing with an ECL kit
(Beyotime Institute of Biotechnology). Data analysis was performed
using ImageJ Software version 1.8.0 (National Institutes of Health)
to evaluate the expression levels of proteins.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analysis was performed by SPSS 17.0 (SPSS, Inc.) and
GraphPad Prism 7.0 (GraphPad Software, Inc.). All experiments were
independently repeated in triplicate. χ2 test was
applied to determine the associations between PCAT6 expression and
the clinical parameters of patients with bladder cancer. Pearson
correlation analysis was used to analyze correlation between the
expression. Comparison between tumor and adjacent non-cancerous
tissues was analyzed using a paired Student's t-test. Comparison
between two independent groups was analyzed via unpaired Student's
t-test, whilst comparisons among multiple groups were calculated by
one-way ANOVA followed by Tukey's test. P<0.05 was considered to
indicate a statistically significant difference.
Results
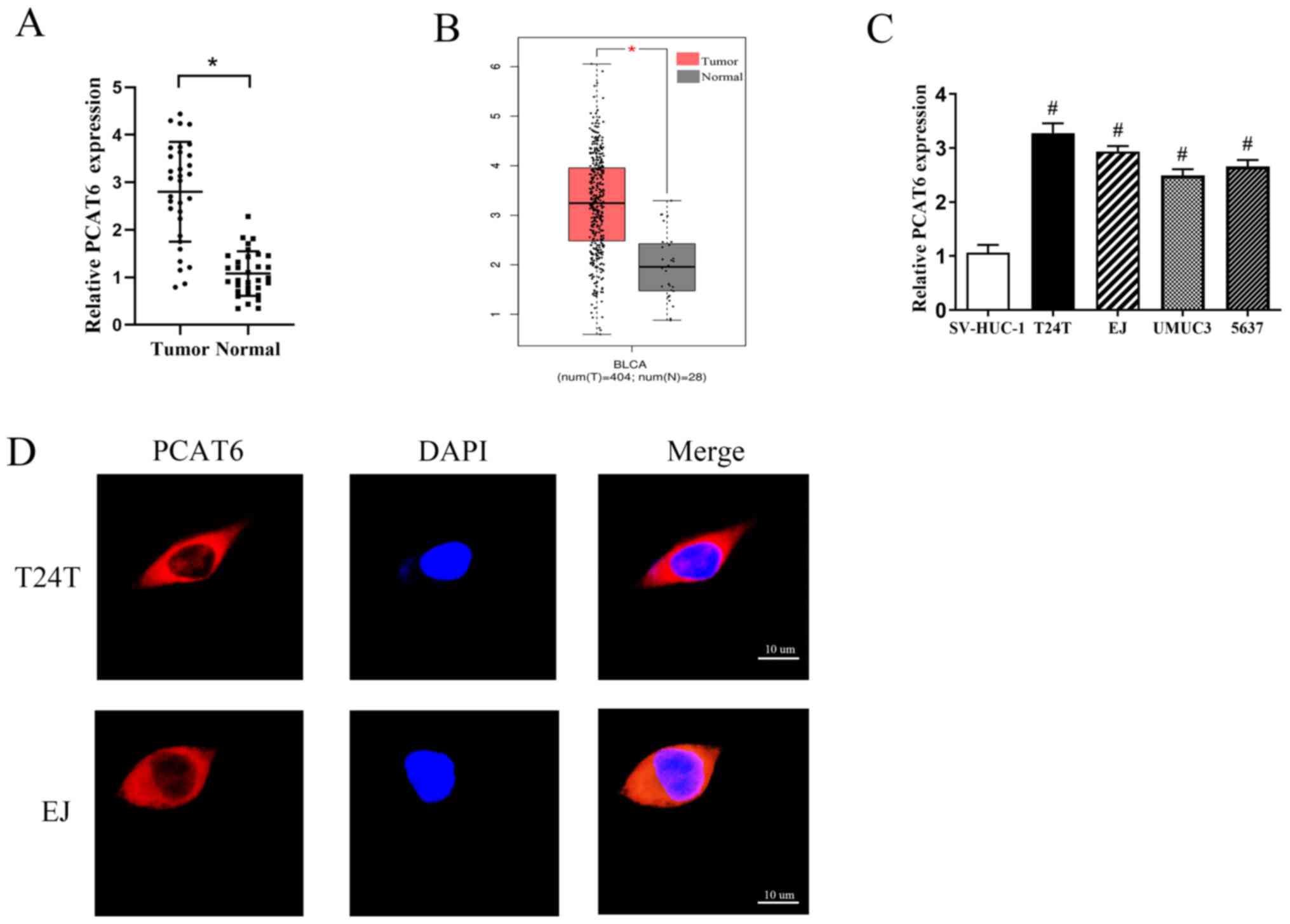
PCAT6 is upregulated in bladder cancer
tissues and cell lines
In order to determine the functions of PCAT6 in
bladder cancer, the expression levels of PCAT6 in 32 pairs of
bladder cancer and adjacent normal tissues were first determined.
The results demonstrated that the expression levels of PCAT6 were
markedly increased in bladder cancer tissues compared with those in
the matched adjacent normal tissues (Fig. 1A). This was consistent with data
from GEPIA (Fig. 1B). Additionally,
the expression levels of PCAT6 were associated with the
pathological stage, but not with other parameters such as sex, age
or tumor grade (Table I).
Subsequently, the expression levels of PCAT6 in four bladder cancer
cell lines (T24T, EJ, UMUC3 and 5637) and one human normal bladder
epithelial cell line (SV-HUC-1) were detected; as demonstrated in
Fig. 1C, the levels of PCAT6
expression were significantly higher in the bladder cancer cells
compared with those in SV-HUC-1 cells. Two cell lines, T24T and EJ,
that exhibited the highest levels of PCAT6 expression were selected
for further experiments.
Emerging studies have suggested that lncRNAs
function through various regulatory mechanisms depending on their
specific location in the cell (21,22).
Therefore, the subcellular localization of PCAT6 in T24T and EJ
cells was analyzed by fluorescent in situ hybridization. The
results revealed that PCAT6 was mainly located in the cytoplasm
(Fig. 1D).
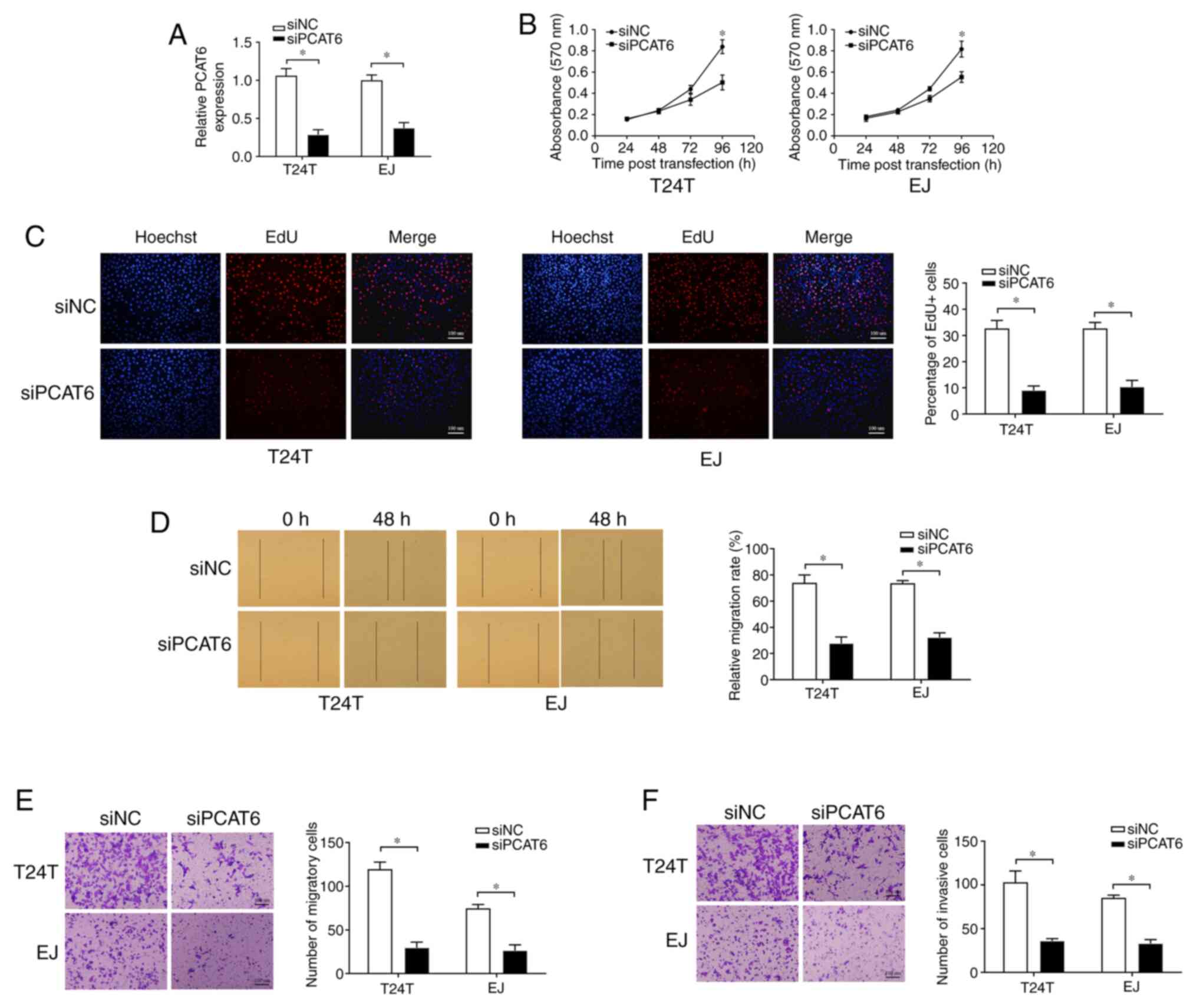
Knockdown of PCAT6 suppresses bladder
cancer cell proliferation, migration and invasion
To determine whether PCAT6 may affect the malignancy
of bladder cancer, T24T and EJ cells were transfected with siPCAT6,
and RT-qPCR analysis demonstrated that the expression of PCAT6 was
effectively inhibited (Fig. 2A).
MTT and EdU assays were conducted to detect cell proliferation,
which revealed that the knockdown of PCAT6 was suppressed cell
proliferation compared with that in the siNC group (Fig. 2B and C). Subsequently, wound healing and
Transwell assays were performed to determine the effects of PCAT6
on bladder cancer cell migratory capacity. As presented in Fig. 2D and E, knockdown of PCAT6 contributed to a
sharp decline in the migration of T24T and EJ cells compared with
that observed in the corresponding siNC groups. In addition, PCAT6
depletion also contributed to the significant suppression of cell
invasion compared with that in the siNC groups, as demonstrated by
Transwell invasion assay (Fig. 2F).
Taken together, these results demonstrated that PCAT6 may serve an
oncogenic role in bladder cancer.
PCAT6 acts as a sponge for miR-143-3p
in bladder cancer cells
To predict the possible downstream targets of PCAT6,
the online bioinformatics tools starBase and Lncbase were used and
the results demonstrated that miR-143-3p contained a binding site
for PCAT6 (Fig. 3A). starBase was
further used to determine that the expression levels of miR-143-3p
were significantly downregulated in bladder cancer tissues compared
with those in the adjacent normal tissues (Fig. 3B). These results were confirmed by
RT-qPCR assay in tissues from the patients included in the present
study (Fig. 3C). In addition,
Pearson's correlation analysis revealed a fair negative correlation
between the expression levels of PCAT6 and miR-143-3p in the
bladder cancer tissues (Fig. 3D).
In addition, higher levels of miR-143-3p were observed in T24T and
EJ cells following PCAT6 knockout compared with those in the
siNC-transfected cells (Fig.
3E).
To confirm the relationship between PCAT6 and
miR-143-3p, a dual-luciferase reporter assay was performed. The WT
and MUT sequences of PCAT6 were designed, and their binding sites
to miR-143-3p are presented in Fig.
3F. As demonstrated in Fig. 3G,
transfection with the miR-143-3p mimics induced a significant
inhibition of the relative luciferase activity of WT-PCAT6, but not
MUT-PCAT6, compared with that in the corresponding mimic-NC groups.
Furthermore, the RIP assay revealed that PCAT6 and miR-143-3p were
enriched in the Ago2-containing immunoprecipitants from T24T and EJ
cells compared with the control IgG groups (Fig. 3H). These results suggested that
miR-143-3p was a direct target of PCAT6 and negatively modulated by
PCAT6.
PCAT6 upregulates the expression of
PDIA6 via miR-143-3p
In order to determine the underlying mechanism of
miR-143-3p-mediated progression of bladder cancer, the downstream
targets of miR-143-3p were predicted using TargetScan software. As
presented in Fig. 4A, PDIA6
contained a putative target sequence for miR-143-3p in its
3'-untranslated region. Therefore, we hypothesized that PCAT6 may
exert its roles in bladder cancer via the miR-143-3p/PDIA6 axis. To
verify this hypothesis, dual-luciferase reporter assay was
performed, and the results demonstrated that the luciferase
activity was suppressed following co-transfection of WT-PDIA6 and
miR-143-3p compared with that in cells transfected with the
mimic-NC (Fig. 4B), indicating that
PDIA6 was a direct target of miR-143-3p in bladder cancer cells. In
addition, PCAT6 knockdown led to a significant reduction of PDIA6
mRNA and protein expression levels, whereas this effect was
reversed by the miR-134-3p inhibitors (Fig. 4C-E). These results suggested that
PDIA6 was a direct target of miR-143-3p, and the expression levels
of PDIA6 were positively modulated by PCAT6.
Inhibition of miR-143-3p reverses the
siPCAT6-mediated suppressive effects on the malignant phenotypes of
bladder cancer cells
In order to determine whether PCAT6 affected bladder
cancer cells via miR-143-3p, T24T and EJ cells were co-transfected
with the miR-143-3p inhibitor and siPCAT6. As demonstrated in
Fig. 5A and B, knockdown of PCAT6 led to a significant
inhibition of cell proliferation compared with that in the
siNC-transfected cells, whereas co-transfection with miR-143-3p
inhibitor reversed this effect. Similarly, the results of the wound
healing and Transwell assays revealed that the inhibition of
miR-143-3p reduced the inhibitory effect of PCAT6 knockdown on cell
migration and invasion (Fig. 5C-E).
These results suggested that PCAT6 was effective in promoting the
malignant behaviors of bladder cancer cells in a miR-143-3p
dependent manner.
Discussion
Bladder cancer is one of the most common urinary
malignancies worldwide with high morbidity and mortality rates
(23). Recent studies have
suggested that lncRNAs may serve essential roles in the development
and malignant progression of bladder cancer (24,25).
Therefore, the study of bladder cancer-related lncRNAs may help
identify early therapeutic targets. lncRNA PCAT6 is a lncRNA that
has been reported to be upregulated in osteosarcoma and colorectal
cancer (26,27) and has emerged as a critical
regulator in tumorigenesis and cancer progression. For example, Bai
et al (28) have
demonstrated that PCAT6 promotes gastrointestinal stromal tumor
cell proliferation and stemness, but inhibits apoptosis through the
ceRNA mechanism and the activation of the Wnt signaling pathway. Ma
et al (14) have reported
that PCAT6 facilitates the proliferation, migration and cisplatin
resistance of cervical cancer cells and impedes their apoptosis via
the PCAT6/miR-543/zinc finger E-box-binding homeobox 1 axis.
However, the understanding of the effects of PCAT6 on bladder
cancer carcinogenesis is limited. In the present study, the
expression levels of PCAT6 were enhanced in bladder cancer tissues
compared with that in adjacent normal tissues. The knockdown of
PCAT6 suppressed bladder cancer cell proliferation, migration and
invasion. According to these results, PCAT6 may be a suitable
putative diagnostic biomarker and therapeutic target for bladder
cancer. However, further studies are required to determine whether
PCAT6 may regulate other biological behaviors of bladder cancer,
such as apoptosis and chemoresistance.
To date, the ceRNA hypothesis has been extensively
studied, and there increasing studies have analyzed the involvement
of a complex network of lncRNA/miRNA/mRNA pathways in various types
of human cancer, including bladder cancer (29-31).
In the present study, miR-143-3p was demonstrated to be a direct
target of PCAT6 in bladder cancer. PDIA6 belongs to the protein
disulfide isomerase (PDI) family and is widely expressed in various
types of human cancer, including breast cancer (32) and ovarian cancer (33). Increasing evidence suggests that
PDIA6 is associated with the progression of various types of cancer
(34-35). For example, Kim et al
(36) reported that inhibition of
PDIA6 may transduce EGFR signaling by activating and inducing A
disintegrin and metalloprotease 17 during the migration and
invasion of U87MG glioblastoma cells. Bai et al (37) have reported that PDIA6 is
upregulated in NSCLC and inhibits cisplatin-induced NSCLC cell
apoptosis and autophagy via the MAP4K1/JNK/c-Jun signaling pathway.
In addition, PDIA6 has been demonstrated to be upregulated in
bladder cancer, and its knockdown inhibits cell proliferation and
invasion via the Wnt/b-catenin signaling pathway (38). In the present study, the expression
levels of PDIA6 were suppressed by PCAT6 silencing in bladder
cancer cells. Furthermore, rescue experiments revealed that
miR-143-3p knockdown reduced the anticancer effects of PCAT6
silencing on cell proliferation, migration and invasion, suggesting
that miR-143-3p/PDIA6 axis may be crucial for the tumorigenic roles
of PCAT6 in bladder cancer.
In conclusion, the results of the present study
revealed that PCAT6 downregulation inhibited cell proliferation,
migration and invasion by targeting the miR-134-3p/PDIA6 axis.
These results may provide a new theoretical basis to explore the
mechanism underlying the regulation of bladder cancer and a novel
target for its diagnosis and treatment.
Acknowledgements
Not applicable.
Funding
Funding: This study was supported by The National Natural
Science Foundation of China (grant no. 81802538) and The Health and
Family Planning Commission of Wuhan (grant no. S201802260064).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GL designed the study, wrote and revised the
manuscript. YZ and LC performed the experiments. YZ analyzed the
data. GL, YZ and LC confirmed the authenticity of all the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
the Central Hospital of Wuhan (2021-011) and written informed
consent was obtained from each patient before enrollment.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Dettlaff K, Stawny M, Ogrodowczyk M,
Jelińska A, Bednarski W, Wątróbska-Świetlikowska D, Keck RW, Khan
OA, Mostafa IH and Jankun J: Formulation and characterization of
EGCG for the treatment of superficial bladder cancer. Int J Mol
Med. 40:329–336. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Babjuk M, Bohle A, Burger M, Capoun O,
Cohen D, Compérat EM, Hernández V, Kaasinen E, Palou J, Rouprêt M,
et al: EAU guidelines on non-muscle-invasive urothelial carcinoma
of the bladder: Update 2016. Eur Urol. 71:447–461. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Clark PE, Spiess PE, Agarwal N, Bangs R,
Boorjian SA, Buyyounouski MK, Efstathiou JA, Flaig TW, Friedlander
T, Greenberg RE, et al: NCCN guidelines insights: Bladder cancer,
version 2.2016. J Natl Compr Canc Netw. 14:1213–1224.
2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Mohammed AA, El-Tanni H, El-Khatib HM,
Mirza AA, Mirza AA and Alturaifi TH: Urinary bladder cancer:
Biomarkers and target therapy, new era for more attention. Oncol
Rev. 10(320)2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Babjuk M: Bladder cancer in the elderly.
Eur Urol. 73:51–52. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Batista PJ and Chang HY: Long noncoding
RNAs: Cellular address codes in development and disease. Cell.
152:1298–1307. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Isin M and Dalay N: LncRNAs and neoplasia.
Clin Chim Acta. 444:280–288. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wang M, Guo C, Wang L, Luo G, Huang C, Li
Y, Liu D, Zeng F, Jiang G and Xiao X: Long noncoding RNA GAS5
promotes bladder cancer cells apoptosis through inhibiting EZH2
transcription. Cell Death Dis. 9(238)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Liu D, Li Y, Luo G, Xiao X, Tao D, Wu X,
Wang M, Huang C, Wang L, Zeng F and Jiang G: LncRNA SPRY4-IT1
sponges miR-101-3p to promote proliferation and metastasis of
bladder cancer cells through up-regulating EZH2. Cancer Lett.
388:281–291. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Orom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Shi R, Wu P, Liu M, Chen B and Cong L:
Knockdown of lncRNA PCAT6 Enhances radiosensitivity in
triple-negative breast cancer cells by regulating miR-185-5p/TPD52
axis. Onco Targets Ther. 13:3025–3037. 2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Dong D, Lun Y, Sun B, Sun H, Wang Q, Yuan
G and Quan J: Silencing of long non-coding RNA PCAT6 restrains
gastric cancer cell proliferation and epithelial-mesenchymal
transition by targeting microRNA-15a. Gen Physiol Biophys. 39:1–12.
2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ma Z, Gu G, Pan W and Chen X: LncRNA PCAT6
accelerates the progression and chemoresistance of cervical cancer
through up-regulating ZEB1 by sponging miR-543. Onco Targets Ther.
13:1159–1170. 2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Miyamoto H, Miller JS, Fajardo DA, Lee TK,
Netto GJ and Epstein JI: Non-invasive papillary urothelial
neoplasms: The 2004 WHO/ISUP classification system. Pathol Int.
60:1–8. 2010.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zheng F, Wang M, Li Y, Huang C, Tao D, Xie
F, Zhang H, Sun J, Zhang C, Gu C, et al: CircNR3C1 inhibits
proliferation of bladder cancer cells by sponging miR-27a-3p and
downregulating cyclin D1 expression. Cancer Lett. 460:139–151.
2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Liu F, Zhang H, Xie F, Tao D, Xiao X,
Huang C, Wang M, Gu C, Zhang X and Jiang G: Hsa_circ_0001361
promotes bladder cancer invasion and metastasis through
miR-491-5p/MMP9 axis. Oncogene. 39:1696–1709. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Li Y, Zheng F, Xiao X, Xie F, Tao D, Huang
C, Liu D, Wang M, Wang L, Zeng F and Jiang G: CircHIPK3 sponges
miR-558 to suppress heparanase expression in bladder cancer cells.
EMBO Rep. 18:1646–1659. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Jiang G, Wu AD, Huang C, Gu J, Zhang L,
Huang H, Liao X, Li J, Zhang D, Zeng X, et al: Isorhapontigenin
(ISO) inhibits invasive bladder cancer formation in vivo and human
bladder cancer invasion in vitro by targeting STAT1/FOXO1 axis.
Cancer Prev Res (Phila). 9:567–580. 2016.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Carlevaro-Fita J and Johnson R: Global
positioning system: Understanding long noncoding RNAs through
subcellular localization. Mol Cell. 73:869–883. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Chen LL: Linking long noncoding RNA
localization and function. Trends Biochem Sci. 41:761–772.
2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Kamat AM, Hahn NM, Efstathiou JA, Lerner
SP, Malmström PU, Choi W, Guo CC, Lotan Y and Kassouf W: Bladder
cancer. Lancet. 388:2796–2810. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Guo Y, Chen D, Su X, Chen J and Li Y: The
lncRNA ELF3-AS1 promotes bladder cancer progression by interaction
with Kruppel-like factor 8. Biochem Biophys Res Commun.
508:762–768. 2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wang F, Zu Y, Zhu S, Yang Y, Huang W, Xie
H and Li G: Long noncoding RNA MAGI2-AS3 regulates CCDC19
expression by sponging miR-15b-5p and suppresses bladder cancer
progression. Biochem Biophys Res Commun. 507:231–235.
2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Guan X, Xu Y and Zheng J: Long non-coding
RNA PCAT6 promotes the development of osteosarcoma by increasing
MDM2 expression. Oncol Rep. 44:2465–2474. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Wu H, Zou Q, He H, Liang Y, Lei M, Zhou Q,
Fan D and Shen L: Long non-coding RNA PCAT6 targets miR-204 to
modulate the chemoresistance of colorectal cancer cells to
5-fluorouracil-based treatment through HMGA2 signaling. Cancer Med.
8:2484–2495. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Bai F, Zhang N, Fang W, He X, Zheng Y and
Gu D: PCAT6 mediates cellular biological functions in
gastrointestinal stromal tumor via upregulation of PRDX5 and
activation of Wnt pathway. Mol Carcinog. 59:661–669.
2020.PubMed/NCBI View
Article : Google Scholar
|
|
29
|
Yu X, Wang ZL, Han CL, Wang MW, Jin Y, Jin
XB and Xia QH: LncRNA CASC15 functions as an oncogene by sponging
miR-130b-3p in bladder cancer. Eur Rev Med Pharmacol Sci.
23:9814–9820. 2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zhou K, Yang J, Li X and Chen W: Long
non-coding RNA XIST promotes cell proliferation and migration
through targeting miR-133a in bladder cancer. Exp Ther Med.
18:3475–3483. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wang F, Zu Y, Huang W, Chen H, Xie H and
Yang Y: LncRNA CALML3-AS1 promotes tumorigenesis of bladder cancer
via regulating ZBTB2 by suppression of microRNA-4316. Biochem
Biophys Res Commun. 504:171–176. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ramos FS, Serino LT, Carvalho CM, Lima RS,
Urban CA, Cavalli IJ and Ribeiro EM: PDIA3 and PDIA6 gene
expression as an aggressiveness marker in primary ductal breast
cancer. Genet Mol Res. 14:6960–6967. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Samanta S, Tamura S, Dubeau L,
Mhawech-Fauceglia P, Miyagi Y, Kato H, Lieberman R, Buckanovich RJ,
Lin YG and Neamati N: Expression of protein disulfide isomerase
family members correlates with tumor progression and patient
survival in ovarian cancer. Oncotarget. 8:103543–103556.
2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Mao L, Wu X, Gong Z, Yu M and Huang Z:
PDIA6 contributes to aerobic glycolysis and cancer progression in
oral squamous cell carcinoma. World J Surg Oncol.
19(88)2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Yan C, Song X, Wang S, Wang J and Li L:
Knockdown of PDIA6 inhibits cell proliferation and enhances the
chemosensitivity in gastric cancer cells. Cancer Manag Res.
12:11051–11062. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Kim TW, Ryu HH, Li SY, Li CH, Lim SH, Jang
WY and Jung S: PDIA6 regulation of ADAM17 shedding activity and
EGFR-mediated migration and invasion of glioblastoma cells. J
Neurosurg. 126:1829–1838. 2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Bai Y, Liu X, Qi X, Liu X, Peng F, Li H,
Fu H, Pei S, Chen L, Chi X, et al: PDIA6 modulates apoptosis and
autophagy of non-small cell lung cancer cells via the MAP4K1/JNK
signaling pathway. EBioMedicine. 42:311–325. 2019.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Cheng HP, Liu Q, Li Y, Li XD and Zhu CY:
The inhibitory effect of PDIA6 downregulation on bladder cancer
cell proliferation and invasion. Oncol Res. 25:587–593.
2017.PubMed/NCBI View Article : Google Scholar
|