Introduction
The incidence of low back pain (LBP) caused by
intervertebral disc degeneration (IDD) has exhibited an increasing
trend across the world (1,2). It was previously reported that the
total number of years lived by individuals with disability caused
by low back pain increased by 54% worldwide between 1990 and
2015(1). The condition impairs the
quality of life of the affected individuals and imposes substantial
socioeconomic and medical costs (1,2). The
estimated indirect and direct cost of LBP in the United States can
reach 100 billion dollars per year (3,4).
Senescence of the nucleus pulposus (NP) cells is one
of the most important features of IDD (5). Collagen-2 is the major component of
the extracellular matrix (ECM), which aids in the maintenance of
the disc height and responds to external mechanical stress
(5,6). NP cells ensure long-term matrix
renewal to maintain the homeostasis of ECM. Decreased content of
collagen-2 in senescent NP cells has been indicated to eventually
lead to IDD (5). Previous studies
have reported a positive association of the degree of cellular
senescence with the IDD grade (6,7). IDD
has long been considered an age-related disease. However, recent
studies have documented a trend of occurrence of IDD in
increasingly younger individuals (8). This phenomenon may be related to
stress-induced premature senescence (SIPS) of NP cells (6).
Production of reactive oxygen species (ROS) is a
normal physiological phenomenon in the intervertebral disc due to
the oxygen-utilizing metabolism of disc cells (9). ROS is a key factor, which increases
survival stress and triggers SIPS at the cellular level (10,11).
It has been indicated that excessive ROS may induce senescence and
apoptosis of NP cells (10).
Antioxidant therapies have also produced good results in the
treatment of IDD (12).
p-Coumaric acid (PCA) is a phenolic acid, which is
widely distributed in numerous plants, as well as human diets
(13,14). Previous studies have demonstrated
the antioxidant, cardioprotective, anti-melanogenic,
anti-mutagenic, anti-platelet and anti-inflammatory effects of PCA
(15-18).
In addition, in a recent study, PCA was indicated to attenuate
IL-1β-induced inflammatory response and cellular senescence in rat
chondrocytes (19). Although
potential antioxidant effects have been suggested for PCA, an
in-depth investigation of its antioxidant and anti-senescence
potential in NP cells has so far not been performed, to the best of
our knowledge. Thus, the present study used an in vitro
model to investigate the inhibitory effect of PCA on ROS-induced
senescence in NP cells.
Materials and methods
Reagents
PCA was purchased from Sigma-Aldrich (Merck KGaA).
It was dissolved in DMSO and the final concentration of DMSO in the
medium was <0.05%. The same volume of DMSO was added to the
control and H2O2 groups in all experiments.
The concentration of PCA used in the present study was based on a
previous study (19).
Cell isolation and culture
NP tissue was isolated from patients with lumbar
fracture who had undergone lumbar interbody fusion surgery. NP
tissues used in the present study were obtained from a total of
five patients (three male and two female; age, 31.5±6.5 years old).
The nucleus pulposus (NP) tissue was collected between January 2020
and December 2020 through surgery from The Second Affiliated
Hospital of Zhejiang University School of Medicine (Hangzhou,
China). The inclusion criterion was that patients underwent lumbar
interbody fusion or discectomy. The exclusion criteria were:
Systemic immune diseases; malignant tumor of the spine; or
infection of intervertebral disc (IVD). The degeneration grade was
evaluated based on modified Pfirrmann grading (20). The degeneration grade was grade 2
in 3 patients and grade 1 in 2 patients. NP tissue was digested
with 0.2% collagenase II (Sigma-Aldrich; Merck KGaA) for 4 h at
37˚C and 0.25% trypsin (Sigma-Aldrich; Merck KGaA) for 15 min at
37˚C. NP cells were collected by centrifugation at 300 x g for 5
min at room temperature. Subsequently, NP cells were cultured in
high-glucose DMEM (Hyclone; Cytiva) mixed with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) and antibiotics (100 U/ml
penicillin, 100 µg/ml streptomycin; Sigma-Aldrich; Merck KGaA).
Cells were cultured at 37˚C with 5% CO2 in fully
humidified air.
Cell proliferation assay
A commercial kit [Cell Counting Kit-8 (CCK-8);
Dojindo Laboratories, Inc.] was used to evaluate the proliferation
of NP cells. NP cells were seeded in 96-well plates at a density of
3x103 cells per well. NP cells were pretreated with
different concentrations of PCA (10 or 50 µg/ml) for 2 h at 37˚C
and then stimulated with 200 µM H2O2, based
on a previous study (21), for 2 h
at 37˚C in the presence or absence of PCA (10 or 50 µg/ml). After
being cultured for 0, 1, 3, 5, 7 or 9 days, the medium was replaced
with fresh culture medium, 10 µl CCK-8 solution was added and the
cells were incubated for an additional 2 h. The optical density
(OD) of each well was measured at 450 nm using a microplate reader
(Thermo Fisher Scientific, Inc.). Culture medium only was used as a
blank control. Cell proliferation (day x) was calculated using the
following equation: Cell proliferation=[OD (day x)-OD (blank)]/[OD
(day 0)-OD (blank)].
Cell cycle assay
Cell cycle analysis was performed using a cell cycle
measurement kit (Beyotime Institute of Biotechnology). NP cells
were seeded at a density of 1x105/well in a six-well
plate, pretreated with different concentrations of PCA (10 or 50
µg/ml) for 2 h and then stimulated with 200 µM
H2O2 for 2 h in the presence or absence of
PCA (10 or 50 µg/ml) at 37˚C. The medium was replaced with fresh
culture medium, and cells were cultured for 72 h. Subsequently, the
cells were washed with cold PBS and fixed with 70% ethanol at 4˚C
overnight. In total, 1x105 cells were resuspended with
RNase A solution (100 µl, Sigma-Aldrich; Merck KGaA) and PI (400
µl; Sigma-Aldrich; Merck KGaA) and incubated for 30 min at 37˚C.
Finally, the DNA content was detected via flow cytometry (BD
LSRFortessa; BD Biosciences) and the percentage of cells in each
phase was analyzed by ModFit LT 5.1 (Verity Software House).
Reverse transcription-quantitative
(RT-q)PCR assay
After treating the cells with
H2O2 and different concentrations of PCA as
aforementioned, total RNA of NP cells was isolated using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) following the manufacturer's instructions. RT was performed
using the PrimeScript™ II 1st Strand cDNA Synthesis Kit (cat. no.
6210A; Takara Bio, Inc.), using the temperature protocol of 37˚C
for 15 min and 85˚C for 5 sec. DNA amplification was performed
using SYBR Premix Ex Taq kit (Takara Bio, Inc.) followed by qPCR.
The thermocycling conditions were set as follows: Initial
denaturation at 95˚C for 3 min; followed by 40 cycles of 95˚C for
30 sec, 55˚C for 20 sec and 72˚C for 20 sec. The primers were
designed and checked using BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Relative
gene expression was calculated using the 2-ΔΔCq method
and GAPDH as the internal reference (22). The primer sequences are presented
in Table I.
 | Table IPrimer sequences used for reverse
transcription-quantitative PCR. |
Table I
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Direction | Primer sequence
(5'-3') |
|---|
| Cox-2 | Forward |
ACCCGTGGAGCTCACATTAACTAT |
| | Reverse |
ATACTGTTCTCCGTACCTTCACCC |
| iNOS | Forward |
CACTTCCAACGCAACATGGG |
| | Reverse |
CTTTGACCCAGTAGCTGCCA |
| p53 | Forward |
GAGGATTCACAGTCGGAT |
| | Reverse |
ATCATCTGGAGGAAGAAGTT |
| p16 | Forward |
GCTGCCCAACGCACCGAATA |
| | Reverse |
ACCACCAGCGTGTCCAGGAA |
| Aggrecan | Forward |
TGAGCGGCAGCACTTTGAC |
| | Reverse |
TGAGTACAGGAGGCTTGAGG |
| Collagen-2 | Forward |
GAACTGGTGGAGCAGCAAGA |
| | Reverse |
AGCAGGCGTAGGAAGGTCAT |
| GAPDH | Forward |
CCACCAACTGCTTAGCCCCC |
| | Reverse |
GCAGTGATGGCATGGACTGTGG |
Immunofluorescence
NP cells were seeded at a density of
3x104/well in a 24-well plate, pretreated with different
concentrations of PCA (10 or 50 µg/ml) for 2 h and then stimulated
with 200 µM H2O2 for 2 h in the presence or
absence of PCA (10 or 50 µg/ml)at 37˚C. The medium was replaced
with fresh culture medium, and cells were cultured for 72 h.
Subsequently, cells were fixed with 4% paraformaldehyde for 20 min
at room temperature, treated with 0.1% Triton X-100 for 10 min and
blocked with 5% bovine serum albumin (Beyotime Institute of
Biotechnology) for 1 h at room temperature. After washing with PBS
three times, NP cells were incubated with primary antibodies
against collagen-2 (cat. no. ab34712; 1:100 dilution; Abcam), p53
(cat. no. 2527; 1:100 dilution; Cell Signaling Technology, Inc.)
and p16 (cat. no. 18769S; 1:100 dilution; Cell Signaling
Technology, Inc.) overnight at 4˚C, before being exposed to Alexa
Fluor® 488-conjugated secondary antibodies (cat. no.
4412; 1:1,000 dilution; Cell Signaling Technology, Inc.) for 60 min
at room temperature. After washing with PBS three times, NP cells
were counterstained with DAPI (cat. no. 40728ES03; 5 mg/ml; 5 min)
and phalloidin conjugated with iFluor™ 555 (cat. no. 40737ES75;
1:10,000 dilution; 30 min) at room temperature (Shanghai Yeasen
Biotechnology Co., Ltd.). A fluorescence microscope (Olympus
Corporation) was used for observation and imaging (x200
magnification). Integrated optical density or positive cell numbers
in each image were measured using Image-Pro Plus 6.0 software
(Media Cybernetics, Inc.). For every sample, six fields of view
were taken before they were averaged.
Senescence-associated β-galactosidase
(SA-β-gal) staining
SA-β-gal staining was performed using the
β-galactosidase senescence staining kit by following the
manufacturer's instructions (cat. no. C0602; Beyotime Institute of
Biotechnology). NP cells were seeded at a density of
1x105/ well in a six-well plate, then washed with PBS
and fixed with 4% paraformaldehyde for 15 min at room temperature.
Subsequently, NP cells were incubated overnight with the SA-β-gal
staining solution at 37˚C. Subsequently, cells with positive
staining were observed and counted under an inverted light
microscope (x200 magnification, Olympus Corporation). For every
sample, six fields of view were taken before they were
averaged.
Statistical analysis
All experiments were performed ≥ three times before
the results are presented as the mean ± standard deviation.
Statistical analysis was performed using one-way analysis of
variance followed by Student-Newman-Keuls post-hoc analysis using
SPSS 21.0 (IBM Corp.) and GraphPad Prism 9.0 (GraphPad Software,
Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
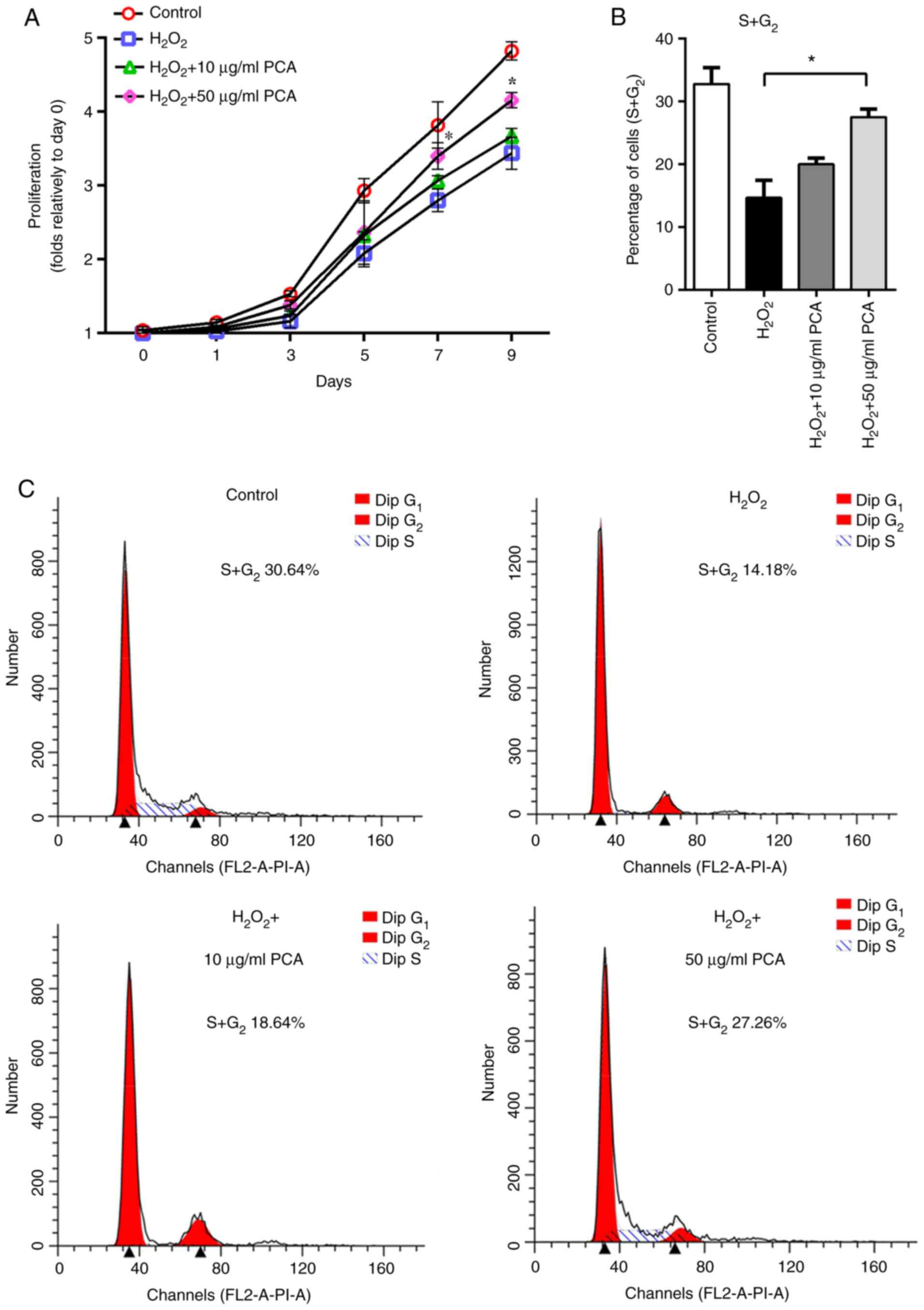
PCA reduces
H2O2-induced anti-proliferative effects and
cell cycle arrest in NP cells
The results of the cell viability assay indicated
anti-proliferative effects of H2O2 on NP
cells. However, pre-treatment with PCA reduced the
anti-proliferative effects of H2O2, where
only the 50 µg/ml group exhibited significantly greater
proliferative capability on days 7 and 9 compared with that in the
H2O2 group (Fig.
1A). To explore whether the ROS-induced anti-proliferative
effects were associated with cell cycle arrest, the cell cycle
distribution of NP cells was detected using flow cytometry. It was
indicated that the percentage of NP cells in S and G2
phases was notably decreased after treatment with
H2O2 compared with that in the control group
(Fig. 1B and C). Treatment with 50 µg/ml PCA reversed
the cell cycle arrest induced by H2O2
(Fig. 1B and C). Furthermore, 10 µg/ml PCA also partly
increased the percentage of NP cells in S and G2 phases
compared with that in the H2O2 group,
although the difference between 10 µg/ml PCA and
H2O2 groups in this respect was not
statistically significant (Fig. 1B
and C).
PCA reduces ROS and senescence-related
gene expression changes in NP cells
Stimulation with H2O2 led to
notable upregulation of the expression of ROS and
senescence-related genes in NP cells (Fig. 2A and B). PCA markedly inhibited the mRNA
expression of cyclooxygenase 2, inducible nitric oxide synthase,
p53 and p16 in a concentration-dependent manner (Fig. 2C and D). Furthermore,
H2O2 stimulation also downregulated the
expression of aggrecan and collagen-2 (Fig. 2E and F). Only 50 µg/ml PCA treatment was able
to counteract the ROS-induced downregulation of collagen-2 and
aggrecan (Fig. 2E and F).
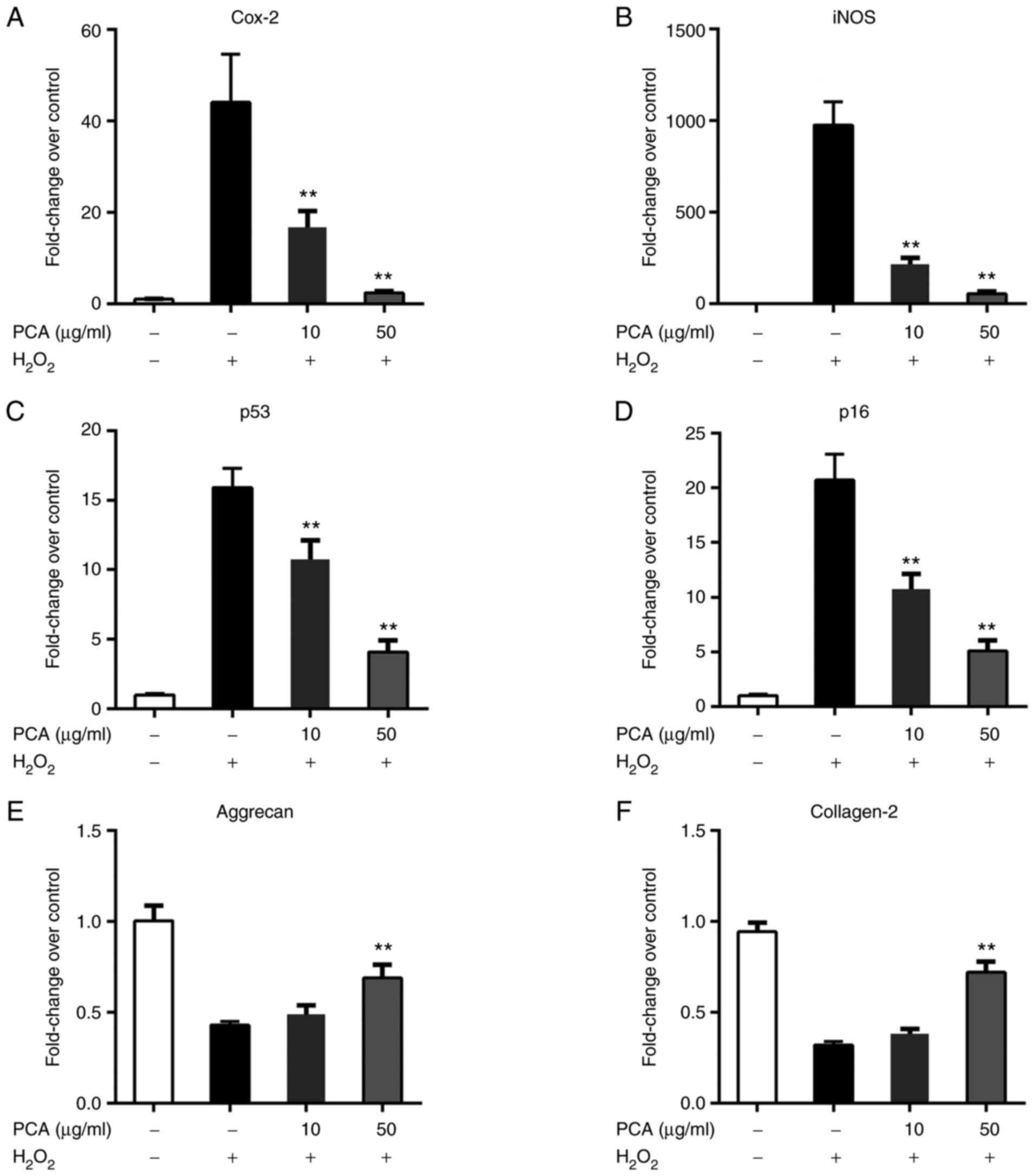 | Figure 2PCA reduces ROS- and
senescence-related gene expression changes in NP cells. NP cells
were treated with H2O2 and different
concentrations of PCA and then cultured for 72 h. Subsequently,
ROS- and senescence-related gene expression was measured via
reverse transcription-quantitative PCR. (A) Cox-2, (B) iNOS, (C)
p53, (D) p16, (E) aggrecan and (F) collagen-2. Values are expressed
as the mean ± standard deviation. **P<0.01 vs.
H2O2 treatment group. NP, nucleus pulposus;
PCA, p-coumaric acid; ROS, reactive oxygen species; Cox-2,
cyclooxygenase 2; iNOS, inducible nitric oxide synthase. |
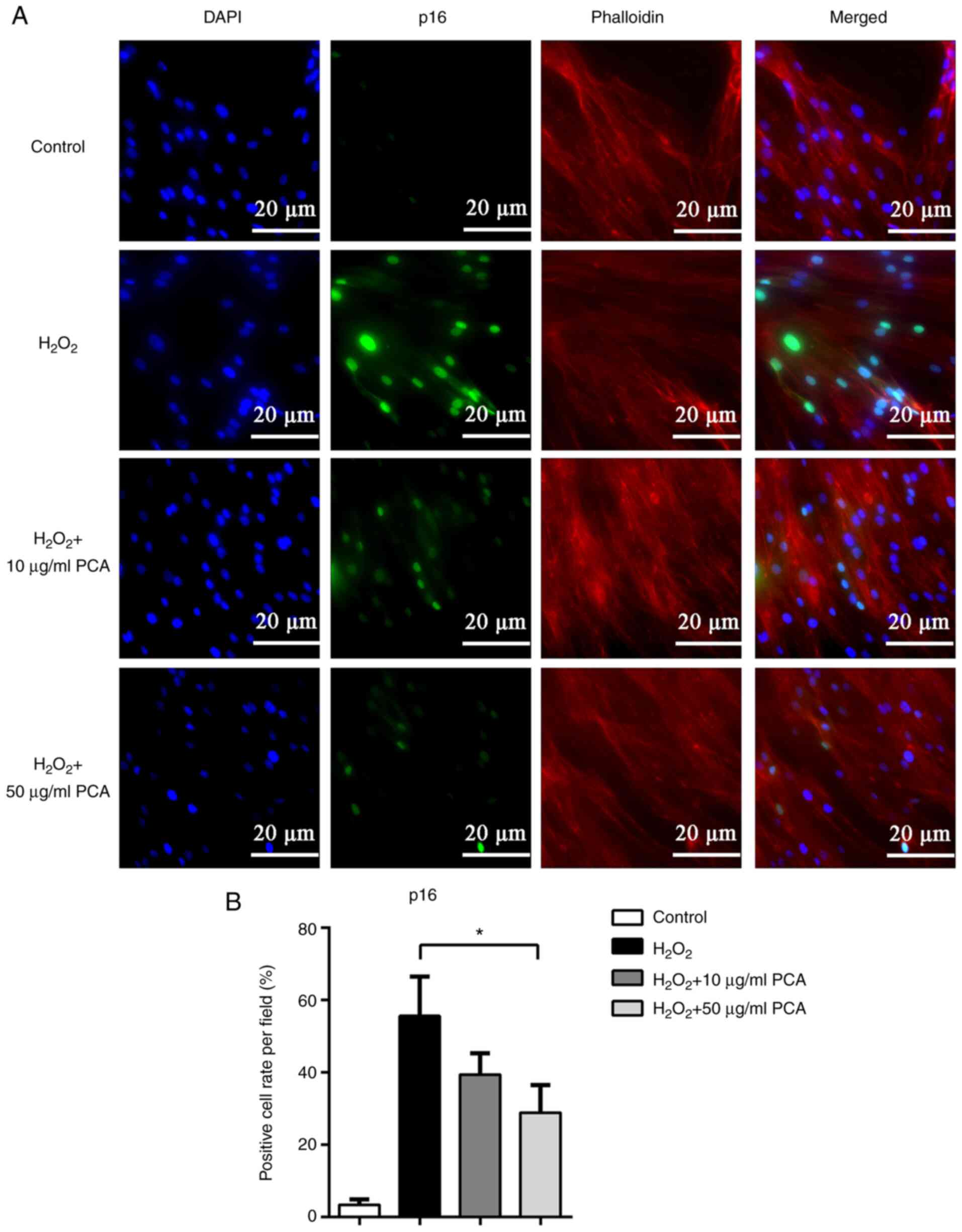
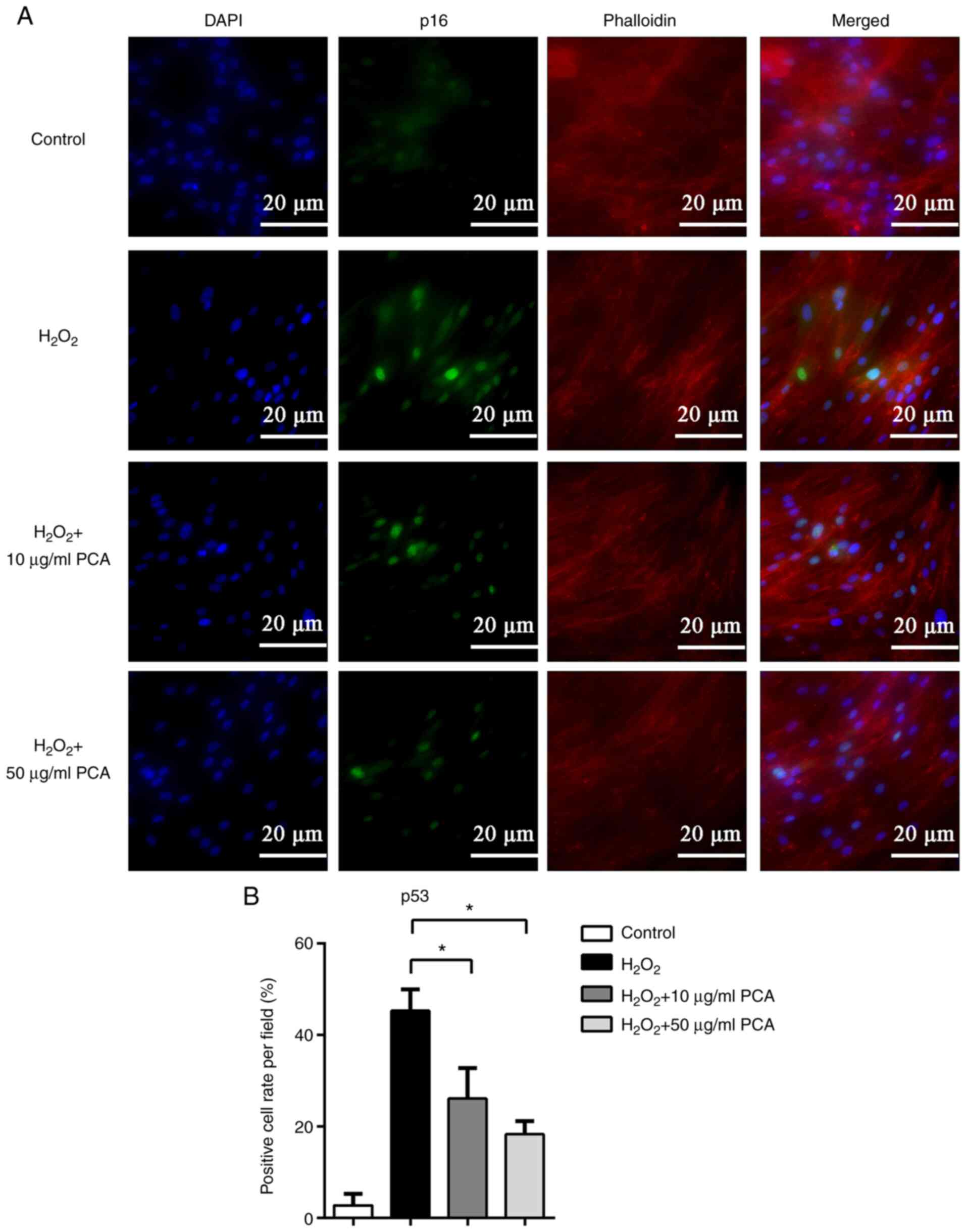
PCA reduces p16 and p53 protein
expression induced by H2O2 in NP cells
To further investigate the changes in the expression
of senescence-related proteins, immunofluorescence staining was
used to locate and measure the expression of p16 and p53. Both p16
and p53 were observed to be expressed in the nucleus and the
expression of p16 and p53 was markedly upregulated after
stimulation with H2O2 (Figs. 3 and 4). PCA treatment markedly reduced p16 and
p53 expression in NP cells compared with that in the
H2O2 group (Figs. 3 and 4). PCA treatment at a concentration of 10
µg/ml resulted in significantly less p53 expression compared with
that in the H2O2 group (Fig. 4).
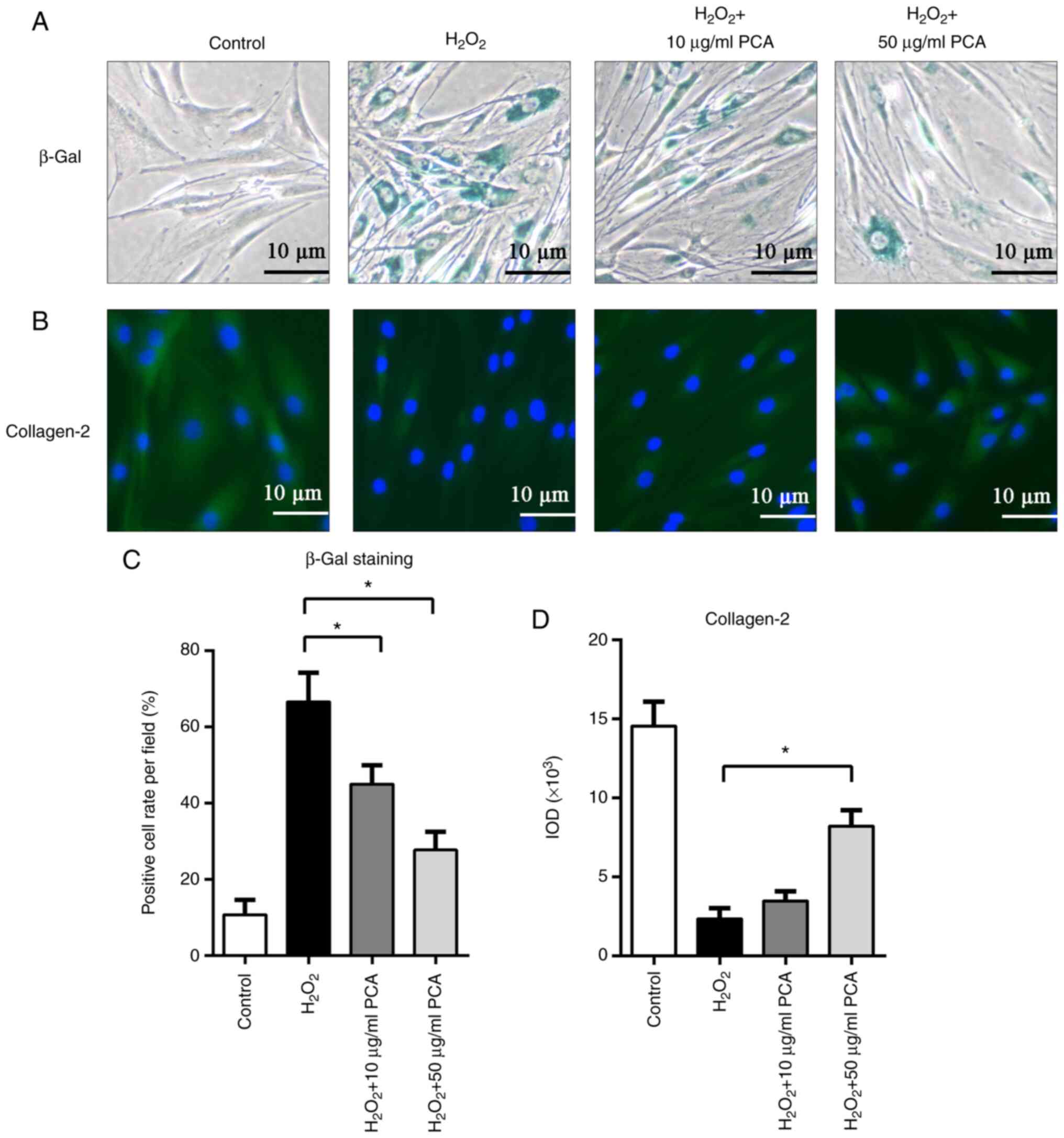
PCA inhibits senescence of NP cells
induced by H2O2
As presented in Fig.
5, β-gal staining was used to identify senescent NP cells.
After stimulation with H2O2, NP cells
exhibited increased β-gal staining. Of note, PCA treatment (10 and
50 µg/ml) significantly reduced β-gal staining after
H2O2 treatment (P<0.05; Fig. 5A and C). Furthermore, immunofluorescence
staining for collagen-2 also indicated that
H2O2 inhibited collagen-2 expression in NP
cells, while PCA (50 µg/ml) restored collagen-2 expression after
H2O2 stimulation (P<0.05; Fig. 5B and C). However, treatment with 10 µg/ml PCA
did not result in any significant difference in collagen-2
expression compared with that in the H2O2
group (P>0.05; Fig. 5B and
C).
Discussion
Cell senescence is considered a key factor in the
pathogenesis of IDD. In normal IVDs, NP cells achieve a balance
between ECM anabolism and catabolism, thus maintaining long-term
matrix renewal (6). Senescent NP
cells are characterized by proliferative arrest and physiological
function abnormalities. For example, senescent NP cells exhibit
decreased ECM synthesis and increased secretion of matrix
degradation enzymes, which perturbs the ECM homeostasis (6). Senescent cells also have the ability
to exhibit senescence-associated secretory phenotype (SASP)
(23). SASP was indicated to
contain proinflammatory factors, cytokines, enzymes and other
bioactive factors (24). SASP may
trigger senescence of the surrounding healthy cells, inducing a
vicious cycle (24). Certain SASP
components, including matrix-degrading enzymes and proinflammatory
factors, were also indicated to be associated with a decrease in
ECM (25). Furthermore, the
avascularity of IVD limits the immune-mediated clearance of
senescent cells (24,26). In a previous study, less
senescence-associated changes were observed in the end plate (EP)
than in the annulus fibrosus and NP cells (27). This may be attributable to the
better blood supply in the EP.
Various signaling pathways have been indicated to be
associated with cell senescence and most of these eventually
regulate cell senescence via interaction with the
p53/p21/retinoblastoma (RB) and p16/RB pathways (28,29).
ROS are a key factor that increases cellular survival stress and
triggers SIPS, and ROS production was indicated to accelerate the
onset and progress of senescence via activation of p21(10). The feedback loop between ROS and
p21 is also important in SIPS: ROS production activates p21
expression, while p21 also induces mitochondrial dysfunction and
ROS production (10).
p21-dependent SIPS is not restricted to proliferative tissues and
the brain neurons also acquire intense DNA damage caused by ROS
production (30).
p53 also serves a critical role in cellular response
to stress (31). p53 mainly
functions as a transcription factor, regulating the expression of
specific target genes (32). These
genes are involved in the regulation of autophagy, DNA damage
repair, senescence and apoptosis (33). The p53 level is positively
associated with the severity and duration of the stress stimulus
and different levels of p53 may have different consequences
(34). In a study by Chen et
al (21), sublethal doses of
H2O2 induced senescent-like growth arrest in
human diploid fibroblasts, while higher doses induced apoptosis. At
higher doses of H2O2, the p53 levels were two
times higher compared with those at sublethal doses of
H2O2 (21).
In the present study, NP cells were treated with 200 µM
H2O2 for 2 h, which has been reported as a
sublethal dose (21). The results
also indicated that after treatment with
H2O2, NP cells exhibited senescent features
instead of undergoing apoptosis.
The anti-inflammatory and anti-oxidative effects of
PCA have been reported in the context of various diseases,
including kidney diseases (35),
liver diseases (36) and
osteoarthritis (19). The dosage
of PCA used in different diseases ranged from 100 to 164 µg/ml
(37). Huang et al
(19) indicated that PCA
attenuated IL-1β-induced inflammatory response and cellular
senescence in chondrocytes. The dosage of PCA used in their study
was 10, 20 and 40 µg/ml, while 40 µg/ml PCA treatment achieved the
best results (19). Considering
that the biological characteristics of NP cells are similar to
those of chondrocytes, 10 and 50 µg/ml PCA were used in the present
study. Treatment with 50 µg/ml PCA significantly inhibited the
cellular senescence of NP cells without affecting cell
proliferation.
The present study was a preliminary study on PCA and
cellular senescence and certain limitations require to be
acknowledged. Firstly, the mechanisms by which PCA regulates
cellular senescence in NP cells were not explored and additional
studies are required to investigate the underlying mechanisms.
Furthermore, animal studies are required to verify the therapeutic
effect of PCA in vivo.
In conclusion, in the present in vitro
experiments, PCA suppressed ROS-induced senescence in NP cells via
both the p16 and p53 pathways. These results suggest that PCA may
be a potential agent for the treatment of IDD.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by a grant from the
National Natural Science Foundation of China (grant no.
81802228).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KS, YL, ZW, KH and ZY designed the study. KS, YL, ZW
and KH performed the research. KS and ZY analyzed the data. KS, YL
and ZY wrote the manuscript. KS and ZY confirm the authenticity of
all the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Institutional Review Board approval for the present
study was provided by the Ethics Committee of The Second Affiliated
Hospital, Zhejiang University School of Medicine (Hangzhou, China;
approval no. 2020-1184). Written informed consent was obtained from
each donor.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hartvigsen J, Hancock MJ, Kongsted A, Louw
Q, Ferreira ML, Genevay S, Hoy D, Karppinen J, Pransky G, Sieper J,
et al: What low back pain is and why we need to pay attention.
Lancet. 391:2356–2367. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Gille O, Bouloussa H, Mazas S, Vergari C,
Challier V, Vital JM, Coudert P and Ghailane S: A new
classification system for degenerative spondylolisthesis of the
lumbar spine. Eur Spine J. 26:3096–3105. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Vos T, Flaxman AD, Naghavi M, Lozano R,
Michaud C, Ezzati M, Shibuya K, Salomon JA, Abdalla S, Aboyans V,
et al: Years lived with disability (YLDs) for 1160 sequelae of 289
diseases and injuries 1990-2010: A systematic analysis for the
global burden of disease study 2010. Lancet. 380:2163–2196.
2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Andersson GB: Epidemiological features of
chronic low-back pain. Lancet. 354:581–585. 1999.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Feng C, Liu H, Yang M, Zhang Y, Huang B
and Zhou Y: Disc cell senescence in intervertebral disc
degeneration: Causes and molecular pathways. Cell Cycle.
15:1674–1684. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhang Y, Yang B, Wang J, Cheng F, Shi K,
Ying L, Wang C, Xia K, Huang X, Gong Z, et al: Cell senescence: A
nonnegligible cell state under survival stress in pathology of
intervertebral disc degeneration. Oxid Med Cell Longev.
2020(9503562)2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Tang N, Dong Y, Chen C and Zhao H:
Anisodamine maintains the stability of intervertebral disc tissue
by inhibiting the senescence of nucleus pulposus cells and
degradation of extracellular matrix via interleukin-6/janus
kinases/signal transducer and activator of transcription 3 pathway.
Front Pharmacol. 11(519172)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Machino M, Yukawa Y, Imagama S, Ito K,
Katayama Y, Matsumoto T, Inoue T, Ouchida J, Tomita K, Ishiguro N
and Kato F: Age-related and degenerative changes in the osseous
anatomy, alignment, and range of motion of the cervical spine: A
comparative study of radiographic data from 1016 patients with
cervical spondylotic myelopathy and 1230 asymptomatic subjects.
Spine (Phila Pa 1976). 41:476–482. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lee DC, Adams CS, Albert TJ, Shapiro IM,
Evans SM and Koch CJ: In situ oxygen utilization in the rat
intervertebral disc. J Anat. 210:294–303. 2007.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Passos JF, Nelson G, Wang C, Richter T,
Simillion C, Proctor CJ, Miwa S, Olijslagers S, Hallinan J, Wipat
A, et al: Feedback between p21 and reactive oxygen production is
necessary for cell senescence. Mol Syst Biol. 6(347)2010.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Deniz GY and Altun S: Evaluation of
nickel-induced brain injuries in rats via oxidative stress and
apoptosis: Attenuating effects of hyperoside. Turk J Zool.
44:104–113. 2020.
|
|
12
|
Tian Y, Bao Z, Ji Y, Mei X and Yang H:
Epigallocatechin-3-gallate protects H2O2-induced nucleus pulposus
cell apoptosis and inflammation by inhibiting cGAS/sting/NLRP3
activation. Drug Des Devel Ther. 14:2113–2122. 2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Tesanovic K, Pejin B, Sibul F, Matavulj M,
Rašeta M, Janjušević L and Karaman M: A comparative overview of
antioxidative properties and phenolic profiles of different fungal
origins: Fruiting bodies and submerged cultures of Coprinus comatus
and Coprinellus truncorum. J Food Sci Technol. 54:430–438.
2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Janjušević L, Karaman M, Šibul F,
Tommonaro G, Iodice C, Jakovljević D and Pejin B: The lignicolous
fungus trametes versicolor (L.) Lloyd (1920): A promising natural
source of antiradical and AChE inhibitory agents. J Enzyme Inhib
Med Chem. 32:355–362. 2017.PubMed/NCBI View Article : Google Scholar
|
|
15
|
An SM, Koh JS and Boo YC: p-Coumaric acid
not only inhibits human tyrosinase activity in vitro but also
melanogenesis in cells exposed to UVB. Phytother Res. 24:1175–1180.
2010.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Luceri C, Giannini L, Lodovici M,
Antonucci E, Abbate R, Masini E and Dolara P: p-Coumaric acid, a
common dietary phenol, inhibits platelet activity in vitro and in
vivo. Br J Nutr. 97:458–463. 2007.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Navaneethan D and Rasool M: p-Coumaric
acid, a common dietary polyphenol, protects cadmium
chloride-induced nephrotoxicity in rats. Ren Fail. 36:244–251.
2014.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Pragasam SJ, Venkatesan V and Rasool M:
Immunomodulatory and anti-inflammatory effect of p-coumaric acid, a
common dietary polyphenol on experimental inflammation in rats.
Inflammation. 36:169–176. 2013.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Huang X, You Y, Xi Y, Ni B, Chu X, Zhang R
and You H: p-Coumaric acid attenuates IL-1β-induced inflammatory
responses and cellular senescence in rat chondrocytes.
Inflammation. 43:619–628. 2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Griffith JF, Wang YX, Antonio GE, Choi KC,
Yu A, Ahuja AT and Leung PC: Modified pfirrmann grading system for
lumbar intervertebral disc degeneration. Spine (Phila Pa 1976).
32:E708–E712. 2007.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Chen QM, Liu J and Merrett JB: Apoptosis
or senescence-like growth arrest: Influence of cell-cycle position,
p53, p21 and bax in H2O2 response of normal human fibroblasts.
Biochem J. 347:543–551. 2000.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Childs BG, Durik M, Baker DJ and van
Deursen JM: Cellular senescence in aging and age-related disease:
From mechanisms to therapy. Nat Med. 21:1424–1435. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Kirkland JL and Tchkonia T: Cellular
senescence: A translational perspective. EBioMedicine. 21:21–28.
2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kepler CK, Ponnappan RK, Tannoury CA,
Risbud MV and Anderson DG: The molecular basis of intervertebral
disc degeneration. Spine J. 13:318–330. 2013.PubMed/NCBI View Article : Google Scholar
|
|
26
|
de Magalhaes JP and Passos JF: Stress,
cell senescence and organismal ageing. Mech Ageing Dev. 170:2–9.
2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Xing QJ, Liang QQ, Bian Q, Ding DF, Cui
XJ, Shi Q and Wang YJ: Leg amputation accelerates senescence of rat
lumbar intervertebral discs. Spine (Phila Pa 1976). 35:E1253–E1261.
2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Munoz-Espin D and Serrano M: Cellular
senescence: From physiology to pathology. Nat Rev Mol Cell Biol.
15:482–496. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Chicas A, Wang X, Zhang C, McCurrach M,
Zhao Z, Mert O, Dickins RA, Narita M, Zhang M and Lowe SW:
Dissecting the unique role of the retinoblastoma tumor suppressor
during cellular senescence. Cancer Cell. 17:376–387.
2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Rass U, Ahel I and West SC: Defective DNA
repair and neurodegenerative disease. Cell. 130:991–1004.
2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liu D and Xu Y: p53, oxidative stress, and
aging. Antioxid Redox Signal. 15:1669–1678. 2011.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Mijit M, Caracciolo V, Melillo A,
Amicarelli F and Giordano A: Role of p53 in the regulation of
cellular senescence. Biomolecules. 10(420)2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Vousden KH and Prives C: Blinded by the
light: The growing complexity of p53. Cell. 137:413–431.
2009.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Spallarossa P, Altieri P, Aloi C,
Garibaldi S, Barisione C, Ghigliotti G, Fugazza G, Barsotti A and
Brunelli C: Doxorubicin induces senescence or apoptosis in rat
neonatal cardiomyocytes by regulating the expression levels of the
telomere binding factors 1 and 2. Am J Physiol Heart Circ Physiol.
297:H2169–H2181. 2009.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Rafiee Z, Moaiedi MZ, Gorji AV and
Mansouri E: P-coumaric acid mitigates doxorubicin-induced
nephrotoxicity through suppression of oxidative stress,
inflammation and apoptosis. Arch Med Res. 51:32–40. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Xie W, Zhang S, Lei F, Ouyang X and Du L:
Ananas comosus L. Leaf phenols and p-coumaric acid regulate liver
fat metabolism by upregulating CPT-1 expression. Evid Based
Complement Alternat Med. 2014(903258)2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Abazari MF, Nasiri N, Karizi SZ, Nejati F,
Haghi-Aminjan H, Norouzi S, Piri P, Estakhr L, Faradonbeh DR and
Kohandani M: An updated review of various medicinal applications of
p-coumaric acid: From antioxidative and anti-inflammatory
properties to effects on cell cycle and proliferation. Mini Rev Med
Chem. 21:2187–2201. 2021.PubMed/NCBI View Article : Google Scholar
|