Introduction
Human mitochondrial DNA (mtDNA) is a 16.6 kb
circular double-stranded DNA molecule, which is present at a high
copy number per cell. Thirteen of the 85 subunits comprising the
oxidative phosphorylation system are encoded by mtDNA, including
subunits within complex I (ND1, ND2, ND3, ND4, ND4L, ND5, ND6),
complex III (CytB), complex IV (COI, COII, COIII) and complex V
(ATPase6, ATPase8). Human mtDNA also contains 24 genes encoding 22
transfer RNAs (tRNAs) and 2 ribosomal RNAs (rRNAs) which are
essential for protein synthesis within the mitochondria. Another
important region in mtDNA is termed the D-loop, which is the
initial site of heavy chain replication and the promoters for heavy
and light chain transcription (1,2).
The mitochondrion functions as a powerhouse to
generate adenosine triphosphate (ATP) through oxidative
phosphorylation (OXPHOS). The mitochondrial OXPHOS also produces
most of the cellular reactive oxygen species (ROS) at complexes I
and III. It is believed that mtDNA mutations in the subunits of
complexes I or III cause aberrant ROS production which may damage
mtDNA, as well as nuclear DNA (3). In particular, the mitochondrion has
relatively less sophisticated DNA protection and repair systems,
and thus it is vulnerable to high mutation rates (4). The mitochondrion also plays central
role in apoptosis (5,6), cell proliferation (7) and calcium signaling (8).
Germline and somatic mtDNA mutations have been found
in primary breast cancer. One mtDNA population polymorphism has
been found to be associated with an increased risk of breast
cancer, suggesting that germline mtDNA mutations may be important
in the etiology of breast cancer. This mutation occurs in the ND3
gene at nt 10398 in which the 10398A allele was linked to an
increased risk of invasive breast cancer in African-American women,
compared with African-American women with the 10398G allele. The
10398 allele was an independent risk factor for breast cancer in
African-American women but no association was detectable in
Caucasian women. It was thought that this mutation results in
increased oxidative stress (9).
However, Setiawan et al argued that there was no association
between the 10398 allele and breast cancer in African-American
women; however, their study did not provide detail information on
the materials and methods that were used (10). Parrella et al reported that
mtDNA mutations were detected in 61% of patients using direct
sequencing. The affected genes included ND4, ND5 and CytB (11). Tan et al reported that 14
of the 19 breast tumors (74%) displayed at least one somatic mtDNA
mutation. Twenty-seven somatic mutations were found and 22 of them
occurred in the D-loop region. The affected genes included ND2,
16srRNA and ATPase6 (12).
Gallardo et al reported mtDNA mutations of the COI gene in
primary breast cancer. The mutation was expected to impair the
interaction between subunit I and II of cytochrome c oxidase
and the mutant had a reduced complex IV activity by 50% (13). Other studies have reported mtDNA
mutations in the ND1, CoIII, tRNA-I and tRNA-T genes (11,14). It is believed that mtDNA mutations
in structure proteins of the mitochondria cause impaired electron
transport function and lead to electron leakage and increased ROS
production, which in turn increases oxidative stress and oxidative
damage to the mitochondria in the process of transformation and
cancer progression. There is no direct evidence on breast cancer to
support the ROS theory thus far; however, there is experimental
evidence on prostate cancer to support this theory. Petros et
al reported that cybrids with pathogenic mtDNA ATP6 T8993G
generated tumors 7-fold larger than the wild-type (T8993T) cybrids.
The mutant tumors generated significantly more ROS (15).
Studies in this field have established an
association between mtDNA mutations and breast cancer. However,
evidence for a direct association between these mtDNA mutations and
breast cancer is still lacking in terms of the function of the
mutant and the development of breast cancer. There has even been
some debate between different research groups (9,10,16). Moreover, there is no reliable
prediction of breast cancer predisposition or progression based on
mtDNA mutation patterns identified thus far.
Materials and methods
All blood samples were collected according to
procedures approved by the Institutional Review Board of the
Guangzhou General Hospital of Guangzhou Military Command. There
were 58 blood samples from breast cancer patients and 58 samples
from age-matched healthy individuals used in this study. mtDNA from
total cellular DNA was enriched by PCR-based strategies.
In PCR-based enrichment, two sets of primers were
designed to amplify amplicons that cover the mtDNA genome. The
primers used in this study were as follos: Mito-8kb-A-Fwd,
GACGGGCTCACATCACCCCATAA/Mito-8kb-A-Rev, GCG TACGGCCAGGGCTATTGGT and
Mito-8kb-B-Fwd, GGT GGCTGGCACGAAATTGACC/Mito-8kb-B-Rev, GCC
ACAACTAACCTCCTCGGACTCCT.
Purified, blunt-ended PCR products were subsequently
fragmented by sonication. Fragmented DNA was then end-repaired,
A-tailed and ligated with an adaptor oligonucleotide from the Ion
Torrent genomic DNA library preparation kit (Life Technologies,
South San Francisco, USA) in accordance with the manufacturer’s
instructions. Adaptor-ligated products were then size-selected by
gel purification and sequenced by Ion Torrent PGM. The Mitomap
(http://www.mitomap.org) and mtDB (http://www.genpat.uu.se/mtDB/) databases were used to
identify sequence variants.
Results
High-throughput sequencing and quality control were
performed. Quality scores were all above 200 and the sequence
content was steady (Fig. 1).
Thus, the sequence data were ready for use in further analysis.
In the following step, we compared data from the
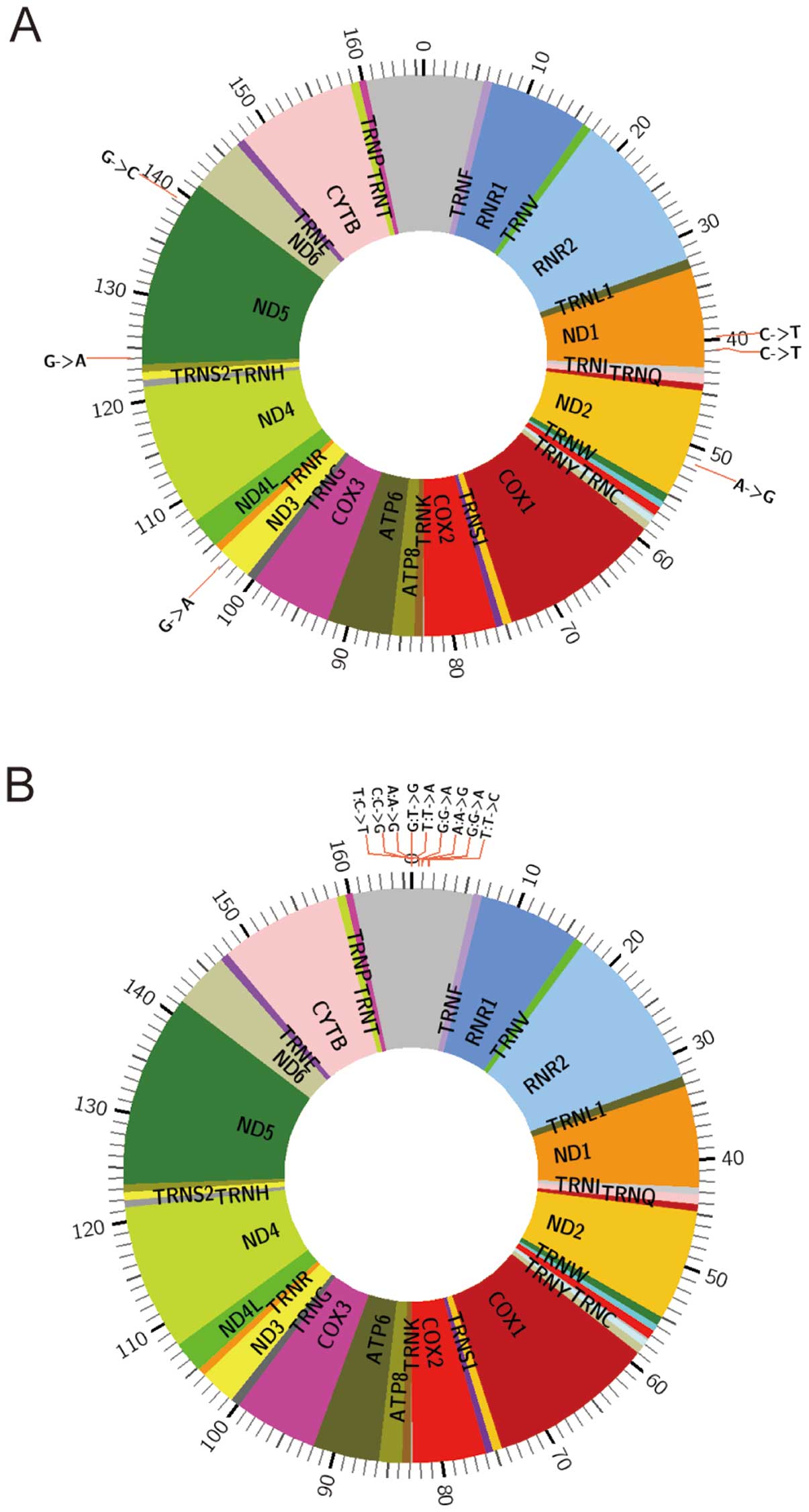
blood of breast cancer patients and the controls in 4 pairs of
samples (Figs. 2–5). There were two pools of 28 blood
samples from patients or controls in pair 1 and pair 2. For pair 3
and pair 4, there was one patient sample and one age-matched
control. The reported variants were confirmed. In addition, we
found a new mtDNA variant (13564 A>G). In this study,
mitochondrial genomes were fully sequenced from blood DNA obtained
from the patients with breast cancer and the normal controls. The
new mtDNA variants may be potential biomarkers for the early
detection of breast cancer. There are several advantages of using
mtDNA as a potential biomarker for cancer-specific mutation
studies. The genome is well characterized, with 16,568 bp harboring
37 genes. Secondly, a high copy number is an advantage over nuclear
DNA for the detection of sequence variants. In addition, DNA repair
is less efficient in the mitochondria than nuclear genomes;
therefore, mutations are more easily identified.
Discussion
Evidence indicates an association between mtDNA
mutations and breast cancer; however, there is no reliable
prediction of breast cancer predisposition or progression based on
mtDNA mutation patterns thus far. In this study, we compared mtDNA
sequence data from the blood of breast cancer patients and
controls. We confirmed the reported variants and found a new mtDNA
variant (13564 A>G). As some studies have suggested, there may
be two classes of cancer mtDNA mutations: tumorigenic mutations and
adaptive mutations. Tumorigenic mutations would be advantageous in
the initial phases of tumor growth, while the adaptive mutations
would be advantageous in the late phases of tumor growth when the
tumor becomes vascularized (17).
Furthermore, since the two classes of mtDNA mutations have
different functions in cancer cells, they may be expected to arise
and be lost from the tumor cells at different phases during tumor
growth. This could be the reason that the heteroplasmid 294 nt ND1
deletion mtDNA has been shown to be present in 50% of the mtDNAs in
primary renal cell carcinoma but absent in the subsequent
metastatic tumors from the same patient (17,18). The same may occur in breast
cancer. Moreover, Rajasimha et al reported that the
selection against pathogenic mtDNA mutations occurs in a stem cell
population. They found that the percentage of pathogenic mtDNA
mutations in the blood decreases exponentially over time compared
with that in hematopoietic stem cells and leukocyte precursors
(19). Therefore, it can be
argued that primary breast cancer cells may not be the right cell
population to detect tumorigenic mtDNA mutations. In other words,
studies carried out on primary breast cancer cells to date have
been unable to detect tumorigenic mtDNA mutations. Thus, perhaps it
would be more effective to detect tumorigenic mtDNA mutations in
breast cancer stem cells.
Solid evidence supports the hypothesis that breast
cancer follows a cancer stem cell model. In 2003, Al-Hajj et
al reported that they were able to identify and isolate a
minority group of breast cancer cells using the surface marker,
CD44+/CD24−/low/Lin−. Only a few
of these cells (only 100 cells) were able to form tumors in mice,
whereas of the rest cell population (tens of thousands cells)
failed to form tumors. Furthermore, this tumorigenic subpopulation
could be serially passaged and the subsequent tumors had a
heterogeneity similar to the parental tumor:
CD44+/CD24−/low/Lin− tumorigenic
cells, as well as the phenotypically diverse mixed populations of
non-tumorigenic cells (20).
There may be some concern in terms of species incompatibilities
since they used a model in which human breast cancer cells were
grown in immunocompromised mice. However, breast cancer stem cells
were confirmed in following studies using mouse models of breast
cancer. In 2008, Cho et al isolated and characterized cancer
stem cells in MMTV-Wnt-1 murine breast tumors. They demonstrated
that Thy1+CD24+ cancer cells were highly
enriched cells capable of regenerating new tumors compared with the
cells of the tumor that did not have this marker profile (21). Similarly, Zhang et al
identified breast cancer stem cells in a p53-null mouse model of
breast cancer using the
Lin−/CD29high/CD24high marker
(22). Vaillant et al
identified breast cancer stem cells in MMTV-Wnt-1, as well as in
p53+/− mouse models using the luminal epithelial
progenitor marker, CD61/β3 integrin (23). It seems that several sets of
markers can be used to isolate breast stem cells, although the
functions of these markers in stem cells remain unclear. In 2007,
Ginestier et al isolated normal and malignant human mammary
stem cells using aldehyde dehydrogenase (ALDH)1 as a marker. They
demonstrated that normal and cancer human mammary epithelial cells
with increased ALDH activity had stem cell/progenitor properties.
In breast tumors, a high ALDH activity identified the tumorigenic
cell fraction which was capable of self-renewal and generating
tumors. The subsequent tumors recapitulated the heterogeneity of
the parental tumor. Furthermore, the expression of ALDH1 detected
by immunostaining correlated with a poor prognosis in a series of
577 breast carcinomas (24). In
2009, Charafe-Jauffret et al isolated breast cancer stem
cells in 23 breast cancer cell lines using ALDH assay followed by
FACS. They confirmed the stem cell properties of ALDH-positive
populations in vitro and in NOD/SCID xenografts (25). Taken together, using
high-throughput sequencing, the findings from our study provide new
evidence to support the association between mtDNA mutations and
breast cancer. As a next step, it would be helpful to detect
tumorigenic mtDNA mutations in breast cancer stem cells to expand
these findings.
Acknowledgements
This study was funded by grants from Guangdong
Province, the China Science and Technology Development Project of
Guangdong Province (2010B011300018-7) and the Natural Science
Foundation of Guangdong Province (8451051501000491).
References
|
1
|
Attardi G and Schatz G: Biogenesis of
mitochondria. Annu Rev Cell Biol. 4:289–333. 1988. View Article : Google Scholar
|
|
2
|
Lightowlers RN, Chinnery PF, Turnbull DM
and Howell N: Mammalian mitochondrial genetics: heredity,
heteroplasmy and disease. Trends Genet. 13:450–455. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wallace DC: A mitochondrial paradigm of
metabolic and degenerative diseases, aging, and cancer: a dawn for
evolutionary medicine. Annu Rev Genet. 39:359–407. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
DiMauro S and Schon EA: Mitochondrial DNA
mutations in human disease. Am J Med Genet. 106:18–26. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang X: The expanding role of mitochondria
in apoptosis. Genes Dev. 15:2922–2933. 2001.PubMed/NCBI
|
|
6
|
Kroemer G and Reed JC: Mitochondrial
control of cell death. Nat Med. 6:513–519. 2000. View Article : Google Scholar
|
|
7
|
Rustin P: Mitochondria, from cell death to
proliferation. Nat Genet. 30:352–353. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Babcock DF and Hille B: Mitochondrial
oversight of cellular Ca2+ signaling. Curr Opin
Neurobiol. 8:398–404. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Canter JA, Kallianpur AR, Parl FF and
Millikan RC: Mitochondrial DNA G10398A polymorphism and invasive
breast cancer in African-American women. Cancer Res. 65:8028–8033.
2005.
|
|
10
|
Setiawan VW, Chu LH, John EM, et al:
Mitochondrial DNA G10398A variant is not associated with breast
cancer in African-American women. Cancer Genet Cytogenet.
181:16–19. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Parrella P, Xiao Y, Fliss M, et al:
Detection of mitochondrial DNA mutations in primary breast cancer
and fine-needle aspirates. Cancer Res. 61:7623–7626.
2001.PubMed/NCBI
|
|
12
|
Tan DJ, Bai RK and Wong LJ: Comprehensive
scanning of somatic mitochondrial DNA mutations in breast cancer.
Cancer Res. 62:972–976. 2002.PubMed/NCBI
|
|
13
|
Gallardo ME, Moreno-Loshuertos R, Lopez C,
et al: m.6267G>A: a recurrent mutation in the human
mitochondrial DNA that reduces cytochrome c oxidase activity
and is associated with tumors. Hum Mutat. 27:575–582. 2006.
|
|
14
|
Zhu W, Qin W, Bradley P, Wessel A, Puckett
CL and Sauter ER: Mitochondrial DNA mutations in breast cancer
tissue and in matched nipple aspirate fluid. Carcinogenesis.
26:145–152. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Petros JA, Baumann AK, Ruiz-Pesini E, et
al: mtDNA mutations increase tumorigenicity in prostate cancer.
Proc Natl Acad Sci USA. 102:719–724. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Salas A, Yao YG, Macaulay V, Vega A,
Carracedo A and Bandelt HJ: A critical reassessment of the role of
mitochondria in tumorigenesis. PLoS Med. 2:e2962005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brandon M, Baldi P and Wallace DC:
Mitochondrial mutations in cancer. Oncogene. 25:4647–4662. 2006.
View Article : Google Scholar
|
|
18
|
Horton TM, Petros JA, Heddi A, et al:
Novel mitochondrial DNA deletion found in a renal cell carcinoma.
Genes Chromosomes Cancer. 15:95–101. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rajasimha HK, Chinnery PF and Samuels DC:
Selection against pathogenic mtDNA mutations in a stem cell
population leads to the loss of the 3243A-->G mutation in blood.
Am J Hum Genet. 82:333–343. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cho RW, Wang X, Diehn M, et al: Isolation
and molecular characterization of cancer stem cells in MMTV-Wnt-1
murine breast tumors. Stem Cells. 26:364–371. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang M, Behbod F, Atkinson RL, et al:
Identification of tumor-initiating cells in a p53-null mouse model
of breast cancer. Cancer Res. 68:4674–4682. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vaillant F, Asselin-Labat ML, Shackleton
M, Forrest NC, Lindeman GJ and Visvader JE: The mammary progenitor
marker CD61/beta3 integrin identifies cancer stem cells in mouse
models of mammary tumorigenesis. Cancer Res. 68:7711–7717. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ginestier C, Hur MH, Charafe-Jauffret E,
et al: ALDH1 is a marker of normal and malignant human mammary stem
cells and a predictor of poor clinical outcome. Cell Stem Cell.
1:555–567. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Charafe-Jauffret E, Ginestier C, Iovino F,
et al: Breast cancer cell lines contain functional cancer stem
cells with metastatic capacity and a distinct molecular signature.
Cancer Res. 69:1302–1313. 2009. View Article : Google Scholar : PubMed/NCBI
|