Introduction
Lower back pain is an endemic problem that causes
substantial disability (1–3).
It is estimated that 60–80% of individuals are affected by lower
back pain at some point in their lives (4–7).
The causes of lower back pain are multifactorial, although 40% of
all cases involve the degeneration of the intervertebral disc (IVD)
(8). However, conservative and
surgical treatments only target the alleviation of the symptoms
rather than the repair or deceleration of the underlying
degenerative process. Therapies, such as gene therapy, to attenuate
or reverse the degenerative process within the IVD are being
developed (9).
The IVD consists of two regions: the inner nucleus
pulposus (NP) and the outer anulus fibrosus (AF). The normal NP is
mainly composed of aggrecan and collagen type II, whereas the
normal AF contains significant amounts of collagen type I (10–12). Previous studies have demonstrated
that the number of NP cells decreases and the composition of the
extracellular matrix associated with these cells is altered in
degenerative discs (13–15). In vitro and in vivo
studies have suggested that the cellular loss attributed to the
excessive apoptosis of disc cells plays an important role in IVD
degeneration (16,17).
Among the apoptotic signaling pathways, two main
caspase-dependent pathways have been observed: the intrinsic and
extrinsic pathways, which are mediated by the mitochondria and
death receptor, respectively (18). Several studies have demonstrated
that the mechanism of apoptosis involves two pathways in NP cells
derived from degenerative human disc which vary among patients.
However, these two pathways ultimately induce caspase-3 to initiate
apoptosis (19–22).
Survivin is a unique inhibitor of apoptosis (IAP)
that deters the activation of intrinsic and extrinsic pathways,
with a focus on the former. Survivin inhibits apoptosis by binding
to caspase-9 or by blocking second mitochondria-derived activator
of caspases (SMAC; a pro-apoptotic protein that binds IAPs and thus
prevents them from inhibiting caspases) and thus prevents the
pro-apoptotic protein from blocking IAP proteins (Fig. 1) (23). The expression of survivin is
strictly controlled in embryonic tissues and in the majority of
tumors, but not during tissue differentiation and maturation
(23,24). Thus, survivin presents an
attractive target for cancer therapy (25), and has been extensively studied in
cell cycle and apoptotic assays for cancer cells (26,27). Studies have demonstrated that the
oncofetal gene, survivin, is re-expressed in osteoarthritis and
rheumatoid arthritis (28–30).
Moreover, preliminary studies have indicated that survivin is
expressed in fetal disc tissue and have noted the differential
expression of survivin between NP tissue derived from degenerative
disc and that derived from a relatively normal disc (31,32). However, to our knowledge, limited
research has been conducted on the effects of lentivirus
(LV)-mediated survivin transfection on NP cells derived from
degenerative human disc in vitro.
The present study was conducted to determine the
effects of the LV-mediated survivin transfection on the morphology
and apoptosis of NP cells in vitro. Our results provide
fundamental information on the effectiveness of gene therapy aimed
at the attenuation of disc degeneration in vitro.
Materials and methods
Experimental materials
NP samples from herniated IVDs of the lumbar spine
were collected from 10 patients, including 6 males and 4 females,
with a mean age of 51 years (46–62 years) (Table I). All patients were diagnosed
with IVD degeneration (lumbar disc herniation) by magnetic
resonance imaging and had undergone spinal fusion to relieve
chronic lower back pain. This study was approved by the Ethics
Committee of the Affiliated Hospital of Qingdao University Medical
College (Shandong, China). Written informed consent was obtained
from all patients prior to enrollment; all patients agreed to the
publication of their clinical data.
 | Table IDemographic data of the patients. |
Table I
Demographic data of the patients.
| Patient no. | Gender | Age (years) | Location of lumbar
disc herniation | Pfirrmann
grade |
|---|
| 1 | Male | 46 | L4/L5 | IV |
| 2 | Female | 50 | L5/S1 | V |
| 3 | Male | 57 | L4/L5 | V |
| 4 | Male | 61 | L5/S1 | V |
| 5 | Female | 62 | L4/L5 | V |
| 6 | Female | 55 | L4/L5 | IV |
| 7 | Male | 48 | L5/S1 | IV |
| 8 | Male | 53 | L5/S1 | V |
| 9 | Female | 51 | L4/L5 | IV |
| 10 | Male | 55 | L4/L5 | V |
The tissue material was harvested and collected
under sterile conditions. A phosphate-buffered saline (PBS)
solution at 4°C was used as a transport medium. All biopsies were
delivered to the laboratory for the processing of cell culture on
the day of harvest.
LV with survivin and an empty LV were purchased from
Shanghai Genechem Co. Ltd. (Shanghai, China) The titers of the LV
carrying survivin and the empty LV were 2×108 and
1×109 TU/ml, respectively.
Cell isolation and culture
The tissues were weighed and washed twice in PBS.
The NP and AF were separated based on their macroscopic
morphologies by omitting the transitional zone. Cells from each
patient were isolated and separately cultured. The NP tissue was
cut into small sections (approximately 1 mm2) and then
digested with 0.25% trypsinase (HyClone, Logan, UT, USA) at 37°C
under gentle agitation. After 20 min, the digestion was terminated
using DMEM/F12 medium (HyClone) with 15% fetal calf serum
(Gibco-BRL, Carslbad, CA, USA) and the tissues were centrifuged at
1,000 rpm for 5 min. Subsequently, 0.5% collagenase type II (MP
Biomedicals, LLC, Santa Ana, CA, USA) was used at 37°C for
approximately 4 h, after which the tissues were centrifuged at
1,000 rpm for 5 min and washed thrice with DMEM/F12 medium
containing 15% fetal calf serum.
The cells were transferred to a 12.5-cm2
culture flask at a density of 105 cells/cm2.
The cells were then cultured in a CO2 incubator (Sanyo
Electric Co., Ltd., Osaka, Japan) at 37°C with humidity and then
grew in DMEM/F12 containing 15% fetal calf serum and 1%
penicillin/streptomycin (Invitrogen Life Technologies, Tokyo,
Japan). The growth medium was changed every 3 days after cell
adhesion. Cell samples from different patients were kept separate
from one another. All experiments were carried out in duplicate and
were conducted with human NP cells from passages 2 to 3.
Immunohistochemistry
NP cells from passage 2 were cultured on glass
slides and were then fixed for 10 min with 4% paraformaldehyde
followed by permeabilization for 5 min with 0.1% Triton X-100 in
PBS. For antigen retrieval, the slides were boiled for 20 min (10
mM citrate buffer, pH 6.0) and then endogenous peroxidase was
Affiliated Hospital of Qingdao University Medical College blocked
using hydrogen peroxide. The cells were pre-incubated for 1 h in a
solution of PBS containing 10% goat serum in order to prevent
non-specific antibody binding. The NP cells were incubated for 16 h
at 4°C with anti-human collagen type II antibody [1:100; Cat. no.
ab3092) and anti-human aggrecan antibody (1:100; Cat. no. ab3778)
[both from Abcam (Hong Kong) Ltd., Hong Kong, China]. Following
incubation, the cells were washed 3 times with PBS, incubated for 1
h at room temperature with rabbit anti-mouse IgG and then rinsed
with PBS. This was followed by coloration with DAB and hematoxylin
staining and observation under a microscope (Olympus GX51; Olympus
Corp., Tokyo, Japan).
Gene transfection
To quantify the percentage of successfully
transfected NP cells at a given multiplicity of infection (MOI), an
identical procedure was performed with the LV-green fluorescent
protein (GFP) (Shanghai Genechem Co., Ltd.) for each MOI assessed.
NP cells from passage 2 were plated as a monolayer in 96-well
plates at 4×104 cells/ml and were incubated for 24 h.
Solutions of the viral particles equal to 30, 40, 60, 80 and 100
MOI were pre-mixed with DMEM/F12 medium and were added to the
96-well plates. After 48 h, the NP cells were examined under a
fluorescence microscope (Olympus CKX41SF; Olympus Corp.), and the
percentage of NP cells synthesizing GFP was determined.
NP cells from passage 2 were divided into 3 groups
(the positive, negative control and blank control groups), which
were transfected with LV with survivin, the empty LV or an equal
amount of DMEM/F12 medium, respectively. The transfection procedure
was performed with an MOI value of 50. The transfected NP cells
were incubated in a CO2 incubator at 37°C. After 8 h,
the growth medium was changed.
Observation of cell morphology
The morphology and growth of the NP cells were
observed daily under an inverted microscope (Olympus CKX41; Olympus
Corp.). The cell morphological changes induced by transfection with
LV carrying survivin were recorded and images were obtained using a
microscope.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
To measure the expression of survivin following
transfection with LV in the transfected cells and subsequent
generation cells, RT-qPCR was performed. mRNA was extracted from
the NP cells using TRIzol reagent (Invitrogen Life Technologies,
San Diego, CA, USA) according to the one-step method. A total of 1
µg mRNA was reverse transcribed into cDNA using PrimeScript
RT Reagent (Takara DRR037A; Takara Bio, Inc., Shiga, Japan), and
the reaction product was treated with RNase-Free DNase I. The
absorbance at 260 and 280 nm was measured for quantification and
quality control.
qPCR was conducted using the following cycling
conditions (LightCycler 480II; Roche Diagnostics GmbH, Mannheim,
Germany): 95°C for 5 min followed by 33 cycles of 94°C for 45 sec,
56°C for 45 sec and 72°C for 45 sec and a final extension at 72°C
for 10 min. Primers and probes were designed using Primer Express
Software (Applied Biosystems, Ltd., Warrington, UK). Total gene
specificity was confirmed by BLAST searches (GenBank database
sequences). Primers were purchased from Sangon Biotech Co., Ltd.
(Shanghai, China) (Table II).
Another specific primer pair for human glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) was used as an internal control. In each
experiment, samples were analyzed in duplicate. The normalized
target gene expression was determined through the comparative Ct
(ΔΔCt) method.
| Table IINucleotide sequences of sense and
antisense primers and product size. |
Table II
Nucleotide sequences of sense and
antisense primers and product size.
| Gene | Primer | Product size
(bp) |
|---|
| Survivin | F:
CAGATGACGACCCCATAGAGGA | 141 |
| R:
CCTTTGCAATTTTGTTCTTGGC |
| GAPDH | F:
GGATTTGGTCGTATTGGG | 205 |
| R:
GGAAGATGGTGATGGGATT |
Detection of apoptosis by flow cytometry
and measurement of caspase-3 activity
Second-generation NP cells transfected with LV were
placed in 6-well culture plates at 1×105 cells/well and
treated as above. The apoptotic rate of the NP cells was detected
by Annexin V/propidium iodide (PI) (Nanjing KeyGen Biotech. Co.,
Ltd., Jiangsu, China) double staining according to the
manufacturer’s instructions and as previously described (17,33). Briefly, the cells of the different
groups were collected by trypsinization and centrifugation, and
then washed with ice-cold PBS twice and resuspended in 500
µl binding buffer. A total of 5 µl of
fluorescein-conjugated Annexin V and 5 µl of PI was added
followed by further incubation in the dark for 15 min at room
temperature. The apoptotic rate was analyzed by flow cytometry (BD
LSR II flow cytometer) using FACSDiva software (both from
Becton-Dickinson, Franklin Lakes, NJ, USA). The cells stained
positive for Annexin V and negative for PI were identified as early
apoptotic cells and those positive for double staining were
identified as late apoptotic cells in each sample. They were
counted and represented as a percentage of the total cell
population.
Apoptosis was evaluated by measuring caspase-3
activity using a Caspase-3 Colorimetric Assay kit (BioVision, Inc.,
Milpitas, CA, USA). Second-generation NP cells were counted and
pelleted at 1.5×106 cells for 48 h following
transfection with LV carrying survivin or the empty LV. The cells
were then resuspended in cell lysis buffer, and 50 µl of 2X
reaction buffer (containing 10 mM DTT) and 5 µl of DEVE-pNA
were added. The samples were incubated for 90 min at 37°C, and the
absorbance was read at 405 nm using a Microtiter Plate Reader
(Sunrise™; Tecan Group, Ltd., Männedorf, Switzerland).
Statistical analysis
All values were presented as the means ± SEM.
One-way ANOVA with Fisher’s least significant difference (LSD) post
hoc test were applied to reveal the statistical significance of the
differences. A value of P<0.05 was considered to indicate a
statistically significant difference. Statistical analyses were
performed using SPSS software for Windows, version 19 (SPSS, Inc.,
Chicago, IL, USA).
Results
Transfection efficiency of LVs
Following transfection of the NP cells with the LVs
for 48 h, all the transfected cells expressed GFP successfully. For
NP cells derived from degenerative human disc, the changes in the
transfection rate were demonstrated by the changes in MOI
(transfection with LV-GFP): 60–70% of cells with an MOI of 30 and
40, 70–85% of cells with an MOI of 60 and 80–100% of cells with an
MOI of 80 and 100 (Fig. 2).
Observation of cell morphology under a
light microscope
The primary NP cells derived from degenerative human
disc were round at the moment of isolation (Fig. 3A) and they had attached to the
culture dish after 5–7 days of culture. The cells gradually became
elongated and triangular or polygonal in shape, and the cytoplasm
became plump and equally distributed. The number of attached cells
exponentially increased. After 15–20 days, 90% of the cells had
formed colonies. The passaged NP cells derived from degenerative
human disc only required 3 h to attach to the culture dish, and 90%
of the cells formed colonies after 7–10 days of culture. Cell
morphology was similar to that of primary cells (Fig. 3B).
To identify the NP cell phenotype,
immunohistochemistry for type II collagen and aggrecan was
performed (Fig. 3C and D). The
results revealed that these cells expressed both type II collagen
and aggrecan, which confirmed the NP cell phenotype.
In the second-passage exponential growth phase,
survivin was transfected into the cells using LV. At 3 days after
transfection, the morphology of the NP cells derived from
degenerative human disc, which were transfected with LV carrying
survivin significantly changed compared with that of the control
groups. In addition, cytomorphosis, the reduction of the cytoplasm
and cell shrinkage were evident. The cell volume changed
significanlty, and pseudopodia became longer (Fig. 4A–C). The morphology of the
third-passage cells was similar to that of the second-passage cells
(Fig. 4D–F).
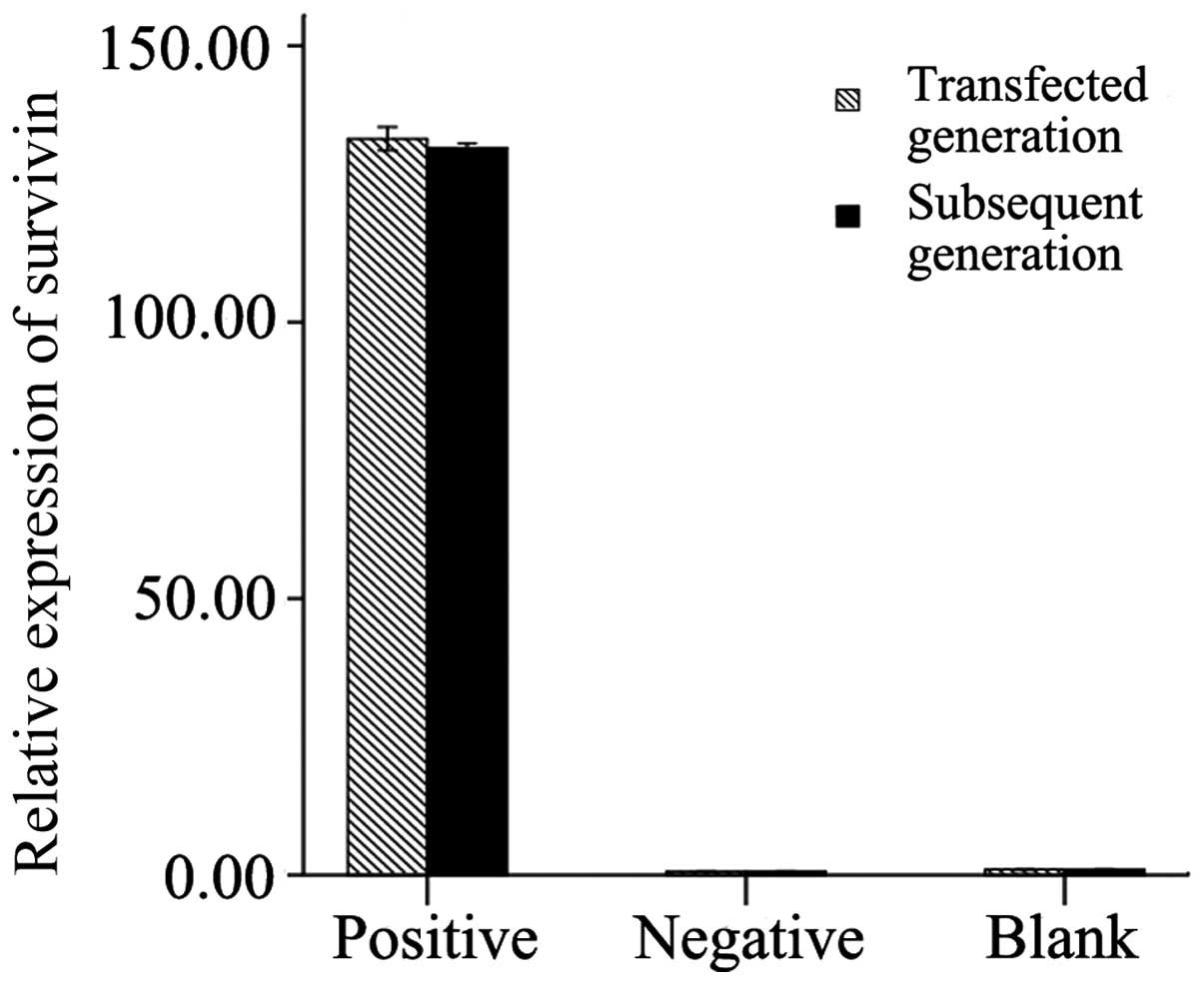
Expression levels of surviving
The survivin mRNA expression levels in the NP cells
derived from degenerative human disc were measured in the
transfected cells and subsequent generation cells. The results
revealed that in the transfected cells and subsequent generation
cells, the expression of survivin following transfection with LV
carrying survivin was significantly higher than that in the control
groups (p<0.01). No significant difference in the survivin
expression levels was observed between the transfected cells and
subsequent generation cells (transfected generation + LV-survivin
vs. subsequent generation + LV-survivin, p=0.242; transfected
generation + empty LV vs. subsequent generation + empty LV,
p=0.996; transfected generation + equal DMEM/F12 medium vs. equal
DMEM/F12 medium + equal DMEM/F12 medium, p=0.999). In additon, no
significant differences were observed between the negative control
and blank control groups (transfected cells, p=0.786; subsequent
generation cells, p=0.790; Fig.
5).
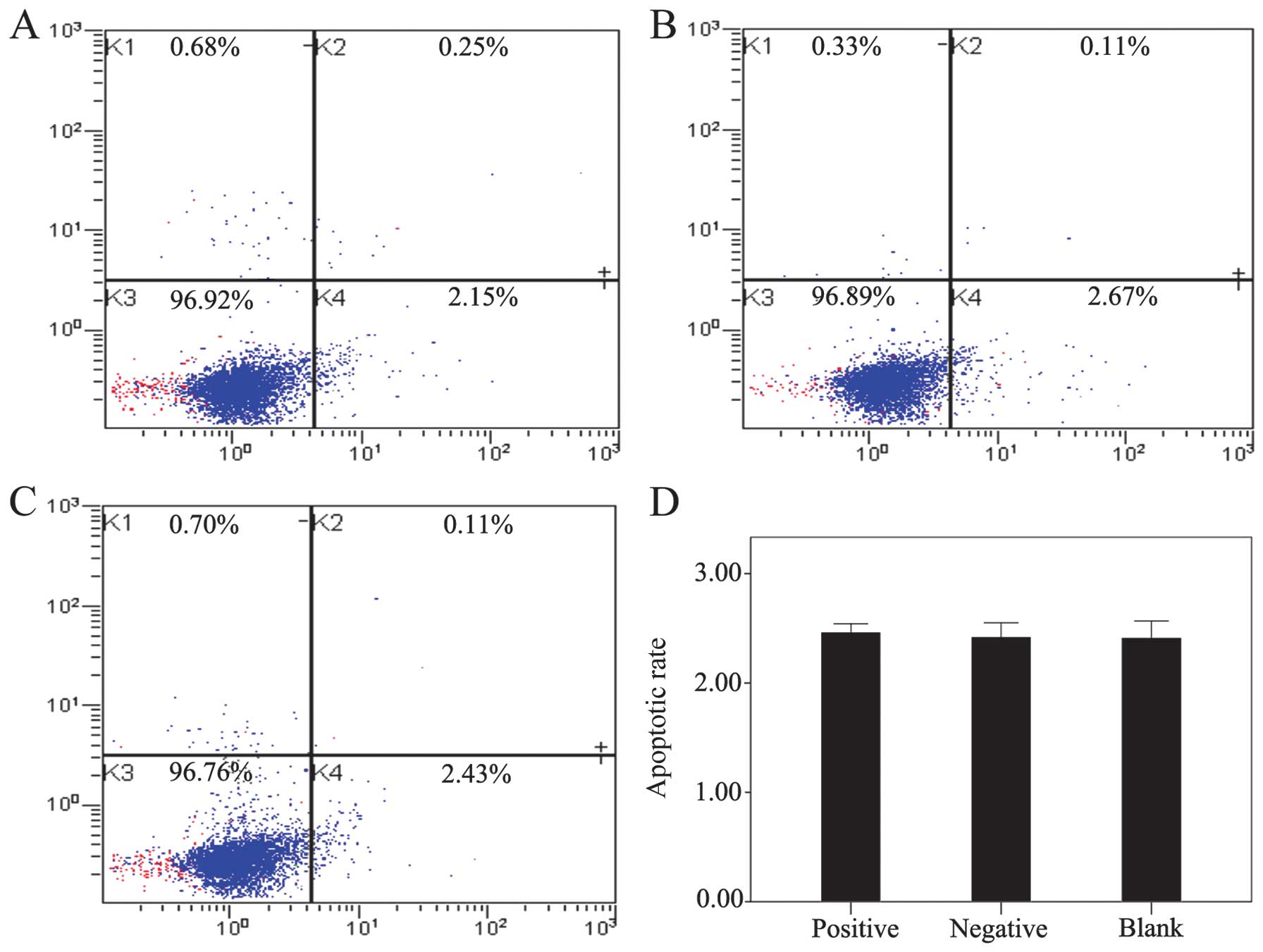
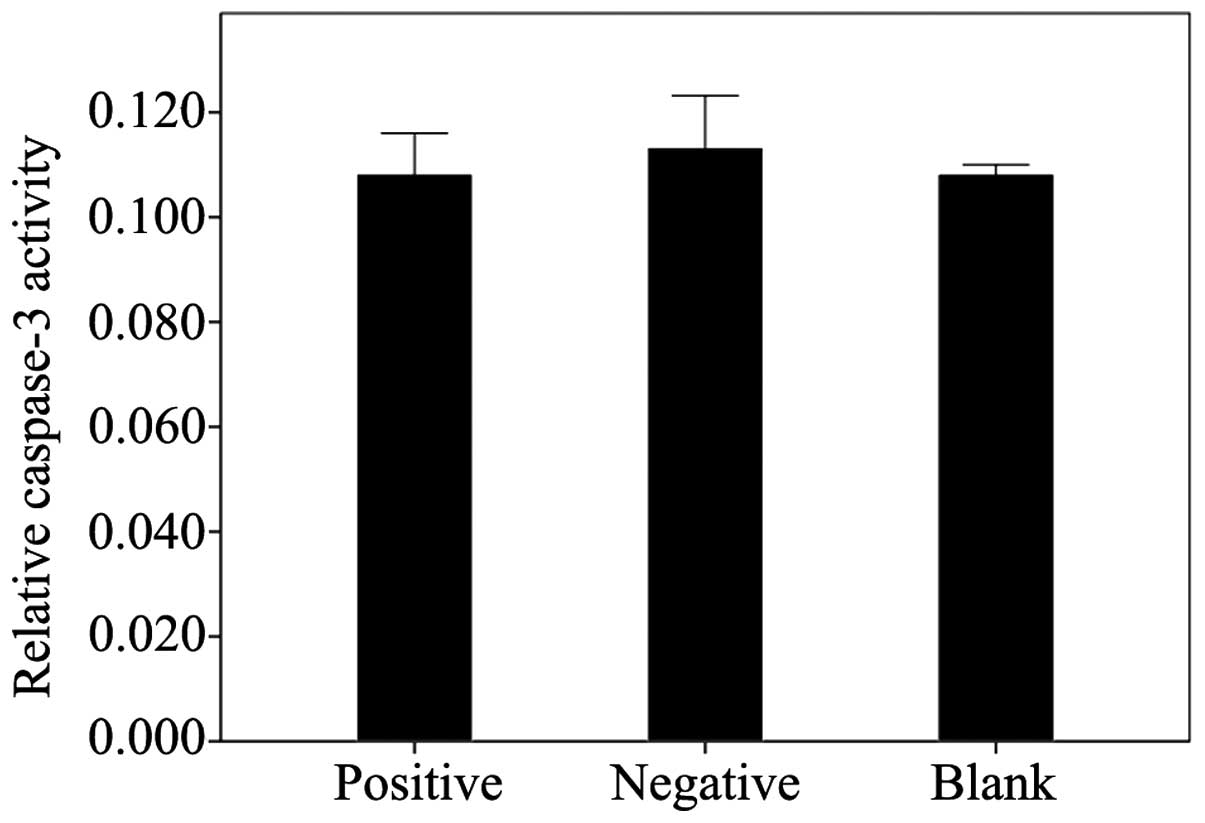
Measurement of apoptotic rate and
caspase-3 activity
The effect of LV-mediated survivin transfection on
the apoptosis of the NP cells derived from degenerative human disc
was examined by flow cytometry and by measuring caspase-3 activity.
The results revealed that the transfection of survivin into the NP
cells did not contribute to decreasing the apoptotic rate and
caspase-3 activity. No significant differences in the apoptotic
rate (Fig. 6) and in caspase-3
activity (Fig. 7) were observed
between the positive (survivin-transfected cells and the control
groups (negative and blank; apoptotic rate, p=0.952; caspase-3
activity, p=0.858).
Discussion
Degenerative disc disease is a serious healthcare
problem. Traditional methods focus on the treatment of multiple
symptoms simultaneously. Gene therapy is a developing technology
with great potential. The in vitro culture of NP cells
derived from degenerative human disc serves as the foundation of
gene therapy and aids in the study of cell proliferation, cell
morphology, gene and protein expression, as well as in cell
function. Therefore, a better understanding of the in vitro
culture of NP cells and the identification of NP cells are
important for future research.
Disc tissue consists of NP and AF cells. No
significant differences have been observed in the morphology of NP
and AF cells during monolayer expansion (34). However, significant differences
have been found between the matrices of NP and AF cells. NP cells
have a gelatinous structure that is primarily composed of aggrecan
and collagen type II. The outer AF cells contain substantial
amounts of collagen type I (35–37). Therefore, the current study
confirmed the NP cell phenotype by using immunohistochemistry for
type II collagen and aggrecan.
In our study, in primary culture the NP cells had a
polygonal shape with short pseudopodia during early proliferation.
However, these cells later became increasingly elongated. This
change in cell morphology was even more pronounced when the cells
were passaged. Compared with the primary NP cells, the passaged
cells had reduced adhesiveness and mostly assumed a short
spindle-shaped appearance. After the fourth passage, cells have
been shown to develop slowly (34). For the negative and blank control
groups, our observations were consistent with the results of a
previous study (34).
However, in the positive group (survivin-transfected
cells), the cell morphology was significantly altered compared with
the other 2 control groups in terms of the reduction in the
cytoplasm, cell shrinkage, lengthening of pseudopodia and increased
intercellular space. Nevertheless, the cells did not die, but
rather remained attached to the culture dish. After passage, these
cells sequentially attached to the dish but did not develop. This
phenomenon suggested that the cell morphology was affected by
transfection.
The results obtained by RT-qPCR revealed the stable
overexpression of survivin following transfection with LV carrying
survivin in the transfected cells and subsequent generation cells.
No significant differences were observed between the NP cells
derived from degenerative human disc transfected with the empty LV
and an equal amount of culture fluid (DMEM/F12). Furthermore,
during our research, LV-TGFβ3, LV-TIMP1, LV-TGFβ3-TIMP1,
LV-survivin-TIMP1, LV-survivin-TGFβ3, and LV-survivin-TGFβ3-TIMP1
were simultaneously transfected into NP cells derived from
degenerative human disc. The results revealed that similar
morphological changes occurred in the NP cells following
transfection with LV-survivin-TIMP1, LV-survivin-TGFβ3, and
LV-survivin-TGFβ3-TIMP1. No changes were observed in the NP cells
following transfection with LV-TGFβ3, LV-TIMP1, LV-TGFβ3-TIMP1
without survivin (data not shown). These results suggest that
survivin contributes to these morhological changes.
Cell morphology is at least partly determined by the
cytoskeleton. It has been demonstrated that some factors may alter
actin filaments through the activation or inhibition of distinct
mitogen-activated protein kinase (MAPK) pathways (38). MAPK pathways have been implicated
in G2/M phase regulation and apoptosis (39–42). The regulation of survivin is
closely associated with MAPK pathways (23,43). Therefore, the overexpression of
survivin may reversely affect MAPK pathways, similar to the
inhibition of different tyrosine kinases in the actin signal
transduction pathways, which may subsequently result in alterations
in cytoskeleton dynamics. To confirm this phenomenon, further
studies are required to examine the structure of the actin
filaments in cells, in which following transfection with LV
carrying survivin, the phosphorylation state of p38 MAPK,
extracellular signal-regulated kinase (ERK) and JNK in response to
transfection needs to be analyzed.
In previous studies, using immunohistochemical
staining, survivin expression was observed in 20-, 26- and 28-week
fetal-age IVDs and the differences were not statistically
significant (31). Survivin was
strongly expressed in NP tissue from degenerative human disc,
whereas it was weakly expressed in NP cells from relatively normal
disc, the difference being statistically significant (p=0.048).
These previous data demonstrate that survivin plays an important
role in fetal IVD growth and is extremely likely to be involved in
the regulation of apoptosis and cell proliferation in the
degeneration process of NP tissue (31,32). We designed the present study based
on the fact that the main function of survivin (mitosis regulation
and apoptosis inhibition) contributes to an increased number of NP
cells, which attenuates IVD degeneration. However, the results did
not meet our expectations, namely that the apoptotic rate and
caspase-3 activity would decrease following LV-survivin
transfection. However, no significant decrease was observed.
No significant decrease was observed in the
apoptotic rate and caspase-3 activity following transfection with
LV carrying survivin. This result may be attributed to the culture
conditions (sufficient oxygen and glucose). As the IVD is the
largest avascular organ in the body and does not directly supply
blood to NP cells (44,45), material and gas exchange mainly
depends on diffusion from the nearest blood supply. Owing to the
progressive age-related degeneration and calcification of the
cartilage end-plate (46), the
number of arteries that supply the periphery of the disc decreases.
This decrease impairs the diffusion function of the IVD and,
therefore, nutrition to the disc and oxygen supply deteriorate.
Thus, NP cells derived from degenerative human disc are under a
relative ischemic condition. Thus, further studies are required to
analyze the function of survivin in in vitro culture under
ischemic conditions.
In the present study, we used LV vectors to
transfect survivin into NP cells and analyzed the gene expression
of survivin by RT-qPCR. Changes in cell morphology were closely
observed under a microscope and recorded. Apoptosis was evaluated
assessed by performing flow cytometry and by measuring caspase-3
activity. It is our intention to further study cell morphology
using electron microscopy in order to confirm the changes in cell
ultrastructure. and explore the effect of transfection with LV
carrying survivin on the apoptosis of NP cells derived from
degenerative human disc in ischemia culture for comparison with
normal culture conditions.
In conclusion, we demonstrated that LV carrying
survivin may be used to effectively deliver the survivin gene into
NP cells derived from degenerative human disc. However, cell
morphology was evidently altered, whereas apoptosis was not
decreased following transfection. Further research is required to
determine whether survivin may be used as a candidate for gene
therapy and, more specifically, for the deceleration of the
degeneration of NP cells, as well as to elucidate the mechanisms
involved in this process.
Acknowledgments
We would like to thank all the members of our
research group for their enthusiastic participation in the present
study. The present study was supported by a research grant awarded
from the National Natural Science Foundation of China (no.
81171758).
References
|
1
|
McMeeken J, Tully E, Stillman B, Nattrass
C, Bygott IL and Story I: The experience of back pain in young
Australians. Man Ther. 6:213–220. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Waddell G: Low back disability. A syndrome
of Western civilization. Neurosurg Clin N Am. 2:719–738.
1991.PubMed/NCBI
|
|
3
|
Waddell G: Low back pain: A twentieth
century health care enigma. Spine (Phila Pa 1976). 21:2820–2825.
1996. View Article : Google Scholar
|
|
4
|
Hillman M, Wright A, Rajaratnam G, Tennant
A and Chamberlain MA: Prevalence of low back pain in the community:
Implications for service provision in Bradford, UK. J Epidemiol
Community Health. 50:347–352. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hoy D, Brooks P, Blyth F and Buchbinder R:
The Epidemiology of low back pain. Best Pract Res Clin Rheumatol.
24:769–781. 2010. View Article : Google Scholar
|
|
6
|
Juniper M, Le TK and Mladsi D: The
epidemiology, economic burden, and pharmacological treatment of
chronic low back pain in France, Germany, Italy, Spain and the UK:
A literature-based review. Expert Opin Pharmacother. 10:2581–2592.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rossignol M, Rozenberg S and Leclerc A:
Epidemiology of low back pain: What’s new? Joint Bone Spine.
76:608–613. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Luoma K, Riihimäki H, Luukkonen R,
Raininko R, Viikari-Juntura E and Lamminen A: Low back pain in
relation to lumbar disc degeneration. Spine (Phila Pa 1976).
25:487–492. 2000. View Article : Google Scholar
|
|
9
|
Levicoff EA, Gilbertson LG and Kang JD:
Gene therapy for disc repair. Spine J. 5(Suppl 6): 287S–296S. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Roughley PJ: Biology of intervertebral
disc aging and degeneration: Involvement of the extracellular
matrix. Spine (Phila Pa 1976). 29:2691–2699. 2004. View Article : Google Scholar
|
|
11
|
Singh K, Masuda K, Thonar EJ, An HS and
Cs-Szabo G: Age-related changes in the extracellular matrix of
nucleus pulposus and anulus fibrosus of human intervertebral disc.
Spine (Phila Pa 1976). 34:10–16. 2009. View Article : Google Scholar
|
|
12
|
Sivan SS, Hayes AJ, Wachtel E, Caterson B,
Merkher Y, Maroudas A, Brown S and Roberts S: Biochemical
composition and turnover of the extracellular matrix of the normal
and degenerate intervertebral disc. Eur Spine J. 23(Suppl 3):
S344–S353. 2014. View Article : Google Scholar
|
|
13
|
Freemont AJ: The cellular pathobiology of
the degenerate intervertebral disc and discogenic back pain.
Rheumatology (Oxford). 48:5–10. 2009. View Article : Google Scholar
|
|
14
|
Le Maitre CL, Freemont AJ and Hoyland JA:
The role of interleukin-1 in the pathogenesis of human
intervertebral disc degeneration. Arthritis Res Ther. 7:R732–R745.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Smith LJ, Nerurkar NL, Choi KS, Harfe BD
and Elliott DM: Degeneration and regeneration of the intervertebral
disc: Lessons from development. Dis Model Mech. 4:31–41. 2011.
View Article : Google Scholar :
|
|
16
|
Rannou F, Lee TS, Zhou RH, Chin J, Lotz
JC, Mayoux-Benhamou MA, Barbet JP, Chevrot A and Shyy JY:
Intervertebral disc degeneration: The role of the mitochondrial
pathway in annulus fibrosus cell apoptosis induced by overload. Am
J Pathol. 164:915–924. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao CQ, Liu D, Li H, Jiang LS and Dai LY:
Interleukin-1beta enhances the effect of serum deprivation on rat
annular cell apoptosis. Apoptosis. 12:2155–2161. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Scaffidi C, Schmitz I, Zha J, Korsmeyer
SJ, Krammer PH and Peter ME: Differential modulation of apoptosis
sensitivity in CD95 type I and type II cells. J Biol Chem.
274:22532–22538. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ha KY, Kim BG, Kim KW, Oh IS and Seo JY:
Apoptosis in the sequestrated nucleus pulposus compared to the
remaining nucleus pulposus in the same patient. Spine (Phila Pa
1976). 36:683–689. 2011. View Article : Google Scholar
|
|
20
|
Kaneyama S, Nishida K, Takada T, Suzuki T,
Shimomura T, Maeno K, Kurosaka M and Doita M: Fas ligand expression
on human nucleus pulposus cells decreases with disc degeneration
processes. J Orthop Sci. 13:130–135. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kuo YJ, Wu LC, Sun JS, Chen MH, Sun MG and
Tsuang YH: Mechanical stress-induced apoptosis of nucleus pulposus
cells: An in vitro and in vivo rat model. J Orthop Sci. 19:313–322.
2014. View Article : Google Scholar
|
|
22
|
Park JB, Lee JK, Park EY and Riew KD:
Fas/FasL interaction of nucleus pulposus and cancer cells with the
activation of caspases. Int Orthop. 32:835–840. 2008. View Article : Google Scholar
|
|
23
|
Johnson ME and Howerth EW: Survivin: A
bifunctional inhibitor of apoptosis protein. Vet Pathol.
41:599–607. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sah NK, Khan Z, Khan GJ and Bisen PS:
Structural, functional and therapeutic biology of survivin. Cancer
Lett. 244:164–171. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Andersen MH, Svane IM, Becker JC and
Straten PT: The universal character of the tumor-associated antigen
survivin. Clin Cancer Res. 13:5991–5994. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ghadimi MP, Young ED, Belousov R, et al:
Survivin is a viable target for the treatment of malignant
peripheral nerve sheath tumors. Clin Cancer Res. 18:2545–2557.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Romagnoli M, Séveno C, Bataille R and
Barillé-Nion S: Survivin in cancerology : Molecular aspects and
therapeutic applications. Med Sci (Paris). 24:821–827. 2008.In
French. View Article : Google Scholar
|
|
28
|
Lechler P, Balakrishnan S, Schaumburger J,
Grässel S, Baier C, Grifka J, Straub RH and Renkawitz T: The
oncofetal gene survivin is re-expressed in osteoarthritis and is
required for chondrocyte proliferation in vitro. BMC Musculoskelet
Disord. 12:1502011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Baran M, Möllers LN, Andersson S, Jonsson
IM, Ekwall AK, Bjersing J, Tarkowski A and Bokarewa M: Survivin is
an essential mediator of arthritis interacting with urokinase
signalling. J Cell Mol Med. 13:3797–3808. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bokarewa M, Tarkowski A and Magnusson M:
Pathological survivin expression links viral infections with
pathogenesis of erosive rheumatoid arthritis. Scand J Immunol.
66:192–198. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang KS, Yue B, Ma XX, et al: The
expression of survivin and its significance in fetal intervertebral
disc. Qingdao Daxue Yixueyuan Xuebao. 49:205–206. 2013.In
Chinese.
|
|
32
|
Yang KS: The expression of survivin and
its significance in intervertebral disc. PhD thesis. Qingdao
University; 2013
|
|
33
|
Zucchini-Pascal N, de Sousa G and Rahmani
R: Lindane and cell death: At the crossroads between apoptosis,
necrosis and autophagy. Toxicology. 256:32–41. 2009. View Article : Google Scholar
|
|
34
|
Kluba T, Niemeyer T, Gaissmaier C and
Gründer T: Human anulus fibrosis and nucleus pulposus cells of the
intervertebral disc: Effect of degeneration and culture system on
cell phenotype. Spine (Phila Pa 1976). 30:2743–2748. 2005.
View Article : Google Scholar
|
|
35
|
Le Maitre CL, Pockert A, Buttle DJ,
Freemont AJ and Hoyland JA: Matrix synthesis and degradation in
human intervertebral disc degeneration. Biochem Soc Trans.
35:652–655. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nerlich AG, Schleicher ED and Boos N: 1997
Volvo Award winner in basic science studies. Immunohistologic
markers for age-related changes of human lumbar intervertebral
discs. Spine (Phila Pa 1976). 22:2781–2795. 1997. View Article : Google Scholar
|
|
37
|
Sive JI, Baird P, Jeziorsk M, Watkins A,
Hoyland JA and Freemont AJ: Expression of chondrocyte markers by
cells of normal and degenerate intervertebral discs. Mol Pathol.
55:91–97. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cui S, Wienhoefer N and Bilitewski U:
Genistein induces morphology change and G2/M cell cycle arrest by
inducing p38 MAPK activation in macrophages. Int Immunopharmacol.
18:142–150. 2014. View Article : Google Scholar
|
|
39
|
Astuti P, Pike T, Widberg C, Payne E,
Harding A, Hancock J and Gabrielli B: MAPK pathway activation
delays G2/M progression by destabilizing Cdc25B. J Biol Chem.
284:33781–33788. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gil-Araujo B, Toledo Lobo MV,
Gutiérrez-Salmerón M, Gutiérrez-Pitalúa J, Ropero S, Angulo JC,
Chiloeches A and Lasa M: Dual specificity phosphatase 1 expression
inversely correlates with NF-κB activity and expression in prostate
cancer and promotes apoptosis through a p38 MAPK dependent
mechanism. Mol Oncol. 8:27–38. 2014. View Article : Google Scholar
|
|
41
|
Pereira L, Igea A, Canovas B, Dolado I and
Nebreda AR: Inhibition of p38 MAPK sensitizes tumour cells to
cisplatin-induced apoptosis mediated by reactive oxygen species and
JNK. EMBO Mol Med. 5:1759–1774. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang H, Ye Y, Chui JH, Zhu GY, Li YW, Fong
DW and Yu ZL: Oridonin induces G2/M cell cycle arrest and apoptosis
through MAPK and p53 signaling pathways in HepG2 cells. Oncol Rep.
24:647–651. 2010.PubMed/NCBI
|
|
43
|
Carter BZ, Mak DH, Schober WD,
Cabreira-Hansen M, Beran M, McQueen T, Chen W and Andreeff M:
Regulation of survivin expression through Bcr-Abl/MAPK cascade:
Targeting survivin overcomes imatinib resistance and increases
imatinib sensitivity in imatinib-responsive CML cells. Blood.
107:1555–1563. 2006. View Article : Google Scholar
|
|
44
|
Anderson DG and Tannoury C: Molecular
pathogenic factors in symptomatic disc degeneration. Spine J.
5(Suppl 6): 260S–266S. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kim KW, Lim TH, Kim JG, Jeong ST, Masuda K
and An HS: The origin of chondrocytes in the nucleus pulposus and
histologic findings associated with the transition of a notochordal
nucleus pulposus to a fibrocartilaginous nucleus pulposus in intact
rabbit intervertebral discs. Spine (Phila Pa 1976). 28:982–990.
2003. View Article : Google Scholar
|
|
46
|
Buckwalter JA: Aging and degeneration of
the human intervertebral disc. Spine (Phila Pa 1976). 20:1307–1314.
1995.
|