Introduction
Prostate cancer is on of the most common cancer
types worldwide and is the second leading cause of
cancer-associated mortalities in men in the United States (1). Hormonal therapy remains the standard
therapy for patients with advanced prostate cancer by targeting
androgen signaling; however, despite initial short-term clinical
responses, most of the patients recur with castrate-resistant
prostate cancer (2). Thus, there
is a requirement to develop novel therapeutic agents to treat
prostate cancer.
Heat shock protein 90 (Hsp90) is an adenosine
triphosphatase (ATPase)-dependent molecular chaperone that is
required for protein folding and maturation, and can interact with
numerous client proteins to prevent their aggregation (3). With the client proteins,
overexpression of Hsp90 is associated with the progression of tumor
cells, including their survival, proliferation, invasion and
metastasis (4). The androgen
receptor (AR) is a nuclear receptor that has a key role in prostate
cancer carcinogenesis and progression, and ARs translocate from the
cytoplasm into the nucleus after activation by androgenic hormones.
The AR has been shown to be a client protein of Hsp90, and Hsp90
inhibition blocks the androgen-induced nuclear import of ARs
(5). Therefore, Hsp90 has been
utilized as a molecular target of anti-cancer drugs and the
development of an Hsp90 inhibitors has become an active area of
research.
As Hsp90 is ubiquitously expressed in various cell
types, selectively inhibiting cancer cell proliferation and
progression by using Hsp90 inhibitors was initially considered
questionable; however, it was demonstrated that tumor cells are
more sensitive to Hsp90 inhibitors than normal cells (3). The reasons for therapeutic
selectivity for cancer versus normal cells can be summarized as
follows: Cancer cells are addicted to the oncogenic processes that
are induced by oncoproteins (6).
As numerous oncoproteins are Hsp90 client proteins, Hsp90
inhibition can deplete these oncoproteins and cause a greater
impact on cancer cells than on normal cells (7,8).
Furthermore, hypoxic, acidic and nutrient-deprived conditions are
common in the tumor microenvironment and may further increase the
number of denatured proteins in tumors. In order to cope with these
environmental stresses, cancer cells become more dependent on Hsp90
than normal cells (9). Finally,
Hsp90 inhibitors can selectively accumulate in tumor tissue while
being rapidly cleared from the circulation and normal tissue
(3,10), partly because Hsp90 isolated from
tumor cells has a higher affinity to Hsp90 inhibitors than Hsp90
isolated from normal cells (3).
Geldanamycin (GA) was identified as the first
natural product inhibitor of Hsp90 that binds to the N-terminal
ATPase domain of Hsp90 to inhibit its chaperone function, and
significantly induces tumor cell death via an apoptotic mechanism
(11,12). However, GA exhibits hepatotoxicity
at its effective concentrations, thus limiting its clinical
application (13). The
modification of position 17 of GA not only leads to the retention
of its the excellent anti-tumor activity but also to a reduction of
its hepatotoxicity (14).
According to this effect, numerous GA derivatives with reduced
hepatotoxicity have been designed, and several of them have entered
clinical trials to treat patients with prostate cancer (15–17). In previous studies by our group
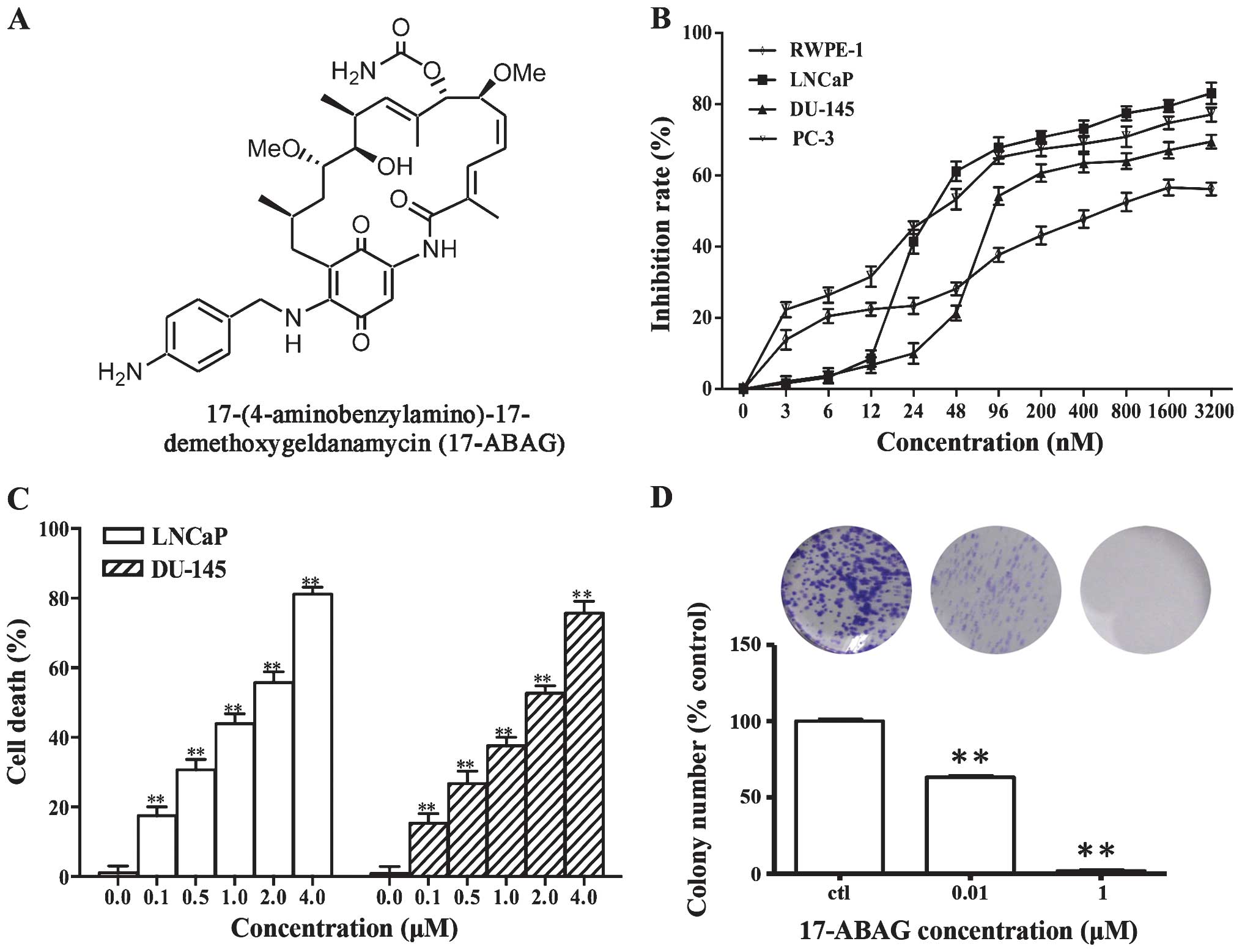
>200 GA derivatives have been designed and synthesized (14,18,19). After screening, 17-ABAG (Fig. 1A) was further examined for its
in vitro and in vivo anti-cancer activities. The
present study further examined the activity and mechanism of action
of 17-ABAG, which showed potent anti-tumor activity against
prostate cancer and low hepatotoxicity in vivo.
Collectively, the present study provided a theoretical foundation
for targeted therapies for prostate cancer and indicated that
17-ABAG is a potent, novel inhibitor of Hsp90.
Materials and methods
17-ABAG was dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich, St. Louis, MO, USA) to prepare 10-mmol/l stock
solutions that were stored at −20°C.
Details of antibodies and reagents
Anti-β-actin antibody (sc-47778, monoclonal, raised
in mouse, 1:10,000) was from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA), MTT, Hoechst 33258 and RNaseA were all purchased
from Sigma-Aldrich. Anti-AKT (#9272, polyclonal, raised in rabbit,
1:1,000), anti-phospho (p)-AKT (ser473; #4058, monoclonal, raised
in rabbit, 1:1,000), anti-human epidermal growth factor receptor 2
(Her2; #2165, monoclonal, raised in rabbit, 1:1,000),
anti-epidermal growth factor receptor (EGFR; sc-03, polyclonal,
raised in rabbit, 1:500), anti-c-Raf (#9422, polyclonal, raised in
rabbit, 1:1,000), anti-B-cell lymphoma 2 (Bcl-2; #2876, polyclonal,
raised in rabbit, 1:1,000) and anti-Bcl-2- associated X protein
(Bax; #2772, polyclonal, raised in rabbit, 1:1,000) antibodies were
from Cell Signaling Technology (Beverly, MA, USA).
Anti-cyclin-dependent kinase 4 (Cdk4; sc-260, polyclonal, raised in
rabbit, 1:500), and anti-prostate-specific antigen (PSA; sc-7316,
monoclonal, raised in mouse, 1:500), anti-AR (sc-7305, monoclonal,
raised in mouse, 1:500), anti-Hsp70 (sc-24; monoclonal, raised in
mouse, 1:500), Hsp90 (sc-69703, monoclonal, raised in mouse, 1:500)
and NKX-3.1 (sc-15022, polyclonal, raised in goat, 1:500)
antibodies were purchased from Santa Cruz Biotechnology, Inc. The
Annexin V fluorescein isothiocyanate (FITC) Apoptosis Detection kit
was purchased from BD Pharmingen (San Diego, CA, USA).
Cells and cell culture
The human androgen-dependent prostate cancer cell
line, LNCaP, two human androgen-independent prostate cancer cell
lines, DU145 and PC-3, and the normal human prostate cell line,
RWPE-1, were purchased from the American Type Culture Collection
(Manassas, VA, USA). The cells were all maintained in Roswell Park
Memorial Institute (RPMI)-1640 medium (Gibco/Invitrogen, Mount
Waverley VIC, Australia). The medium was supplemented with 10%
fetal bovine serum (FBS, Gibco) 2 mM L-glutamine (Gibco),
penicillin (100 units/ml) and streptomycin (100 μg/ml,
Biowest LLC, Kansas City, MO, USA). The cells were incubated at
37°C in an atmosphere of 5% CO2 and 95% air.
Cytotoxicity assays
The cytotoxicity of the compounds was measured using
an MTT assay (Sigma-Aldrich) as previously described (20). DNA content was detected using a
FCM-FC500 system (Beckman Coulter, Brea, CA, USA)
Measurement of cell death
Cell death induced by compounds was determined by
evaluating the plasma membrane integrity by examining the
permeability of cells to propidium iodide (PI). Cells were
trypsinized, collected and centrifuged at 300 × g at 4°C for 5 min,
washed once with phosphate-buffered saline (PBS) and re-suspended
in PBS containing 5 μg/ml PI. The level of PI incorporation
was quantified by flow cytometry using a FACScan flow cytometer
(Beckman Coulter EPICS XL; Beckman Coulter).
Colony formation assay
Cells were cultured in six-well plates (1,000/well)
overnight, followed by replacement of the medium with added
17-ABAG. The plates were then incubated at 37°C with 5%
CO2 for 10 days. On the last day, the medium was
removed, and after washing with PBS and fixing with methanol, the
colonies were stained with crystal violet solution (Sangon Biotech,
Shanghai, China) for 3 h at room temperature. The cells were
observed under a microscope (Leica DMIL; Leica Microsystems,
Wetzlar, Germany) and images were captured with a scanner (Leica
Application Suite version 4.40; Leica Microsystems) and visible
colony numbers were counted after washing and air-drying.
Detection of apoptosis by DAPI
staining
Following treatment with 17-ABAG, the cells were
collected and washed once with 2 ml of ice-cold PBS, fixed with 1
ml 4% paraformaldehyde for 20 min and washed once again with 2 ml
ice-cold PBS. The cells were incubated in 1 ml PBS containing DAPI
at 50 μg/ml and 100 μg/ml RNase A (both from
Sigma-Aldrich). This mixture was incubated for 30 min at 37°C.
After washing with 2 ml PBS three times, the cells were observed
using a fluorescence microscope (Leica DM2500; Leica Microsystems)
at 340 nm (excitation) and 488 nm (emission).
Assessment of apoptosis by Annexin V/PI
staining
Cells were seeded in a 6-well plate 1 day prior
treatment with the compounds. After 17-ABAG treatment for 24 h,
cells were stained with Annexin V and PI following the
manufacturer’s instructions (Annexin V-FITC Apoptosis Detection
kit; BD Pharmingen). Subsequently, cells were analyzed by flow
cytometry and BD CellQuest Pro software (BD Pharmingen) using the
FL1 channel for FITC and FL3 detector for PI.
Western blot analysis
After treatment with or without 17-ABAG for
different durations, cells were harvested and lysed in ice-cold
lysis buffer [20 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM EDTA, 1 mM
ethylene glycol tetraacetic acid, 1% Triton X-100, 2.5 mM sodium
pyrophosphate, 1 mM β-glycerolphosphate, 1 mM sodium orthovanadate,
1 mg/ml leupeptin and 1 mM phenylmethylsulfonyl fluoride; Beyotime
Biotechnology, Shanghai, China]. The lysate was mixed with an equal
volume of 2X loading buffer [4% SDS, 10% 2-mercaptoethanol, 20%
glycerol and 0.2 mg/ml bromophenol blue in 0.1 M Tris-HCl (pH 6.8)]
and boiled for 10 min immediately. The boiled lysates were
separated by 8–12% SDS-PAGE at 100 V and then were transferred onto
Immobilon-P membranes (Millipore, Billerica, MA, USA). After
blocking the membranes with 5% skimmed milk in PBS with 0.1%
Tween-20 for 1 h, they were incubated overnight with the
corresponding primary antibodies in blocking solution at 4°C.
Antibodies against the following proteins were obtained from Santa
Cruz Biotechnology, Inc.: EGFR, Hsp70, AR, Hsp90, PSA, NKX-3.1 and
Cdk4. Antibodies against the following proteins were from Cell
Signaling Technology (Danvers, MA): Bax, Bcl-2, HER2, phospho-Akt,
Akt and c-Raf. The primary antibodies were detected using either a
peroxidase-conjugated ImmunoPure goat anti-rabbit immunoglobulin G
(IgG) (H+L) or peroxidase-conjugated ImmunoPure goat anti-mouse IgG
(H+L) secondary antibody and enhanced chemiluminescence (Western
ECL reagent, WBKL0500, Millipore). TBST was used for washing
between the addition of the primary and secondary antibodies. The
Fluor Chem-E western imaging system (ProteinSimple, Santa Clara,
CA) was used to capture images.
Reverse transcription quantitative
polymerase chain reaction (PCR)
Total RNA from 2×106 cells for each cell
line was isolated using TRIzol reagent (Invitrogen Life
Technologies, Carlsbad, CA, USA). Two micrograms of total RNA were
reverse transcribed using the Transcriptor First Strand cDNA
synthesis kit (Roche Applied Science, Basel, Switzerland). To
synthesize thecDNA, 0.5 mM deoxynucleoside triphosphate, 50 pmol
random hexamers, 50 U ExScript reverse transcriptase (200 U/μl), 10
U RNase inhibitor, 500 ng total RNA, and 1X reaction buffer were
mixed in each reaction tube (10 μl per reaction) and then
incubated at 42°C for 15 min, followed by a 2-min incubation at
95°C to inactivate the ExScript reverse transcriptase. Real-time
monitoring of PCR amplification of the cDNA was performed using DNA
primers and the ABI PRISM 7500 HT Sequence Detection system
(Applied Biosystems; Foster City, CA, USA) with SYBR PCR Master Mix
(Thermo Fisher Scientific, Waltham, MA, USA) using the following
program: 95°C for 10 sec, 1 cycle; 95°C for 5 sec, 62°C for 31 sec,
40 cycles; followed by a 30-min melting curve collection to verify
the primer dimers. Target gene expression was normalized to GAPDH
levels in the respective samples as an internal standard, and the
comparative cycle threshold (Ct) method was used to calculate
relative amount of target mRNAs, as previously described (21). Oligonu cleotide primers used for
PCR amplification of human GAPDH, PSA, NKX3.1 and FKBP5 were as
follows: GAPDH sense, 5′-TCCTGTTCGACAGTCAGCCGCA-3′ and antisense,
5′-ACCAGGCGCCCAATACGACCA-3′; PSA sense, 5′-CACAGCCTGTTTCATCCTGA-3′
and antisense, 5′-AGGTCCATGACCTTCACAGC-3′; NKX3.1 sense,
5′-GGACTGAGTGAGCCTTTTGC-3′ and antisense,
5′-CAGCCAGATTTCTCCTTTGC-3′; FKBP5 sense, 5′-TCCCTCGAATGCAACTCTCT-3′
and antisense, 5′-GCCACATCTCTGCAGTCAAA-3′. Each PCR reaction was
carried out in triplicate.
Immunofluorescence
LNCaP cells were grown on coverslips and treated
with or without 0.2 μM 17-ABAG for 24 h followed by
treatment with 1 nmol/l R1881 for 6 h. After treatment, cells were
fixed with 4% formaldehyde, permeabilized for 10 min in 0.2% Triton
X-100 in PBS and then incubated for 1 h in blocking buffer (5%
bovine serum albumin in PBS). Next, the cells were incubated with
AR (1:250) antibody overnight, and were then visualized with
Cy3-conjugated addinipure goat anti-mouse IgG (H+L). Nuclei were
stained by incubating the cells with 10 μg/ml Hoechst 33258
(Sigma-Aldrich) in PBS and then washing extensively with PBS.
Images were captured using a fluorescence microscope (Leica DM2500;
Leica Microsystems).
In vivo anti-tumor assays
Six- to eight-week-old male athymic nude mice
(BALB/c-nu; n=14) were obtained from Slac Laboratory Animal
(Shanghai, China). The mice were kept in cages (97
cm2×12.7 cm) in an environment with a temperature of
26–28°C, a humidity of 40–60% and a 10 hlight/14 h dark cycle and
were allowed free access to food and water (every other day). LNCaP
or DU-145 cells (1×106 cells/animal) were injected
subcutaneously to generate orthotopic xenografts. Next, the mice
bearing tumor cells were randomly divided into treatment and
control groups (7 mice per group). The drug was injected via the
caudal vein every three days at a dose of 10 mg/kg body weight,
whereas the blank control group received an equal volume of 5%
glucose (Tianjin Pacific Pharmaceutical, Tianjin, China) injection
containing 1% DMSO and 2% lecithin (Sangon Biotech). During
treatment, subcutaneous tumors were measured with a vernier caliper
every three days, and body weight was monitored regularly (the mean
weight on day 0 was 25.287 (control) vs. 24.646 (treated group),
and on day 21 it was 25.621 (control) vs. 25.468 (treated group).
The tumor volume was calculated by the formula (V=ab2/2,
where a and b represent the longest and shortest diameters of the
tumor, respectively). After treatment for 21 days with the drug,
the animals were sacrificed by spinal dislocation and solid tumors
were removed. All of the animal protocols of the present study were
approved by the Shanghai Medical Experimental Animal Care
Commission (Shanghai, China).
Immunohistochemical staining
Sections of the heart, liver, spleen, lung, kidney
and tumor were prepared for immunohistochemical analysis from
sacrificed mice with tumor xenografts as previously described
(22). The expression of Ki67
(Santa Cruz Inc, sc-15402) were detected by immunohistochemical
staining. The paraffin-embedded sections were pre-treated and
stained with antibodies. The secondary antibodies against rabbit
IgG were supplied in an IHC kit (#CW2069) from Beijing Cowin
Bioscience Co., Ltd., Beijing, China, and the sections were
observed using a Leica DM2500 microscope (Leica Microsystems).
Statistical analysis
Values are expressed as the means ± standard error
of the mean. Student’s t-test (SPSS 19.0, IBM) was used to
determine the significant differences between the treatment and
control groups. P<0.05 was considered to indicate a
statistically significant difference between values. All of the
experiments were conducted in triplicate.
Results
17-ABAG inhibits LNCaP cell
proliferation
To evaluate the in vitro anti-tumor effects
of 17-ABAG, MTT assays were performed to examine the proliferative
inhibitory activity of 17-ABAG against the normal human prostate
cell line RWPE-1 and the three human prostate cancer cell lines
LNCaP, DU-145 and PC-3. In all of the cell lines tested, 17-ABAG
inhibited the cell growth in a dose-dependent manner. In the
prostate cancer cell lines, 17-ABAG displayed potent cytotoxicity
with half maximal inhibitory concentration (IC50) values
ranging from 30.15 to 102.63 nmol/l (LNCaP, 30.15 nM; DU-145,
102.63 nM; PC-3, 44.27 nM) at 72 h (Fig. 1B). However, 17-ABAG showed lower
cytotoxicity to RWPE-1 cells, with an IC50 value of 589
nM (Fig. 1B). These results
indicated that 17-ABAG possesses high activity against LNCaP cells
but lower cytotoxicity against normal prostate cells (RWPE-1).
To assess the ability of 17-ABAG to induce cell
death, membrane integrity was assessed using PI staining. The
results showed that 17-ABAG induced cell death of LNCaP and DU-145
cells in a dose-dependent manner (Fig. 1C), suggesting that cell death is
the main contributor to the anti-proliferative activity of 17-ABAG.
Consistently, a colony formation assay showed that the numbers of
colonies formed by the cells treated with 17-ABAG significantly
decreased compared with that of the control LNCaP (Fig. 1D) and Du-145 cells (data not
shown).
17-ABAG induces LNCaP cell apoptosis
Induction of apoptosis is one of the important
mechanisms via which chemotherapeutic drugs kill tumor cells. DAPI
staining revealed that 17-ABAG induced morphological changes in the
cells within 24 h of exposure (Fig.
2A). The cells shrank, became rounded and contained fragmented
nuclei, all of which are characteristic morphological features
(i.e., condensed nuclei) of stressed cells moving into apoptosis.
These observations led to the hypothesis that 17-ABAG induces
apoptosis of LNCaP cells. Flow cytometry and western blot analyses
were utilized to explore whether the anti-proliferative activity of
17-ABAG is associated with apoptosis. The Annexin V-PI assay
revealed that the number of cells undergoing apoptosis
significantly increased following treatment with 17-ABAG compared
with that in the control LNCaP cells (Fig. 2B). In addition, 17-ABAG increased
the expression levels of apoptosis-associated protein Bcl-2 and
reduced the expression levels of Bax (Fig. 2C). These results suggested that
17-ABAG induces apoptosis via the classic apoptotic pathway.
 | Figure 217-ABAG induces prostate cancer cell
apoptosis. (A) LNCaP cells were treated for 24 h with or without 1
μM 17-ABAG. The images were captured using fluorescence
microscopy (magnification, ×600) to observe nuclear staining with
DAPI. Apoptotic cells with condensed nuclei were identified using
nuclear staining as indicated by the white arrows. (B) Detection of
apoptosis using Annexin V-FITC/PI double staining. After treatment
with or without 1 μM 17-ABAG for 24 h, LNCaP cells were
analyzed using a FACSCalibur flow cytometer. The horizontal axis
represents Annexin V intensity, and the vertical axis shows PI
staining. The lines divide each plot into four quadrants: Lower
left quadrant, live cells; lower right quadrant, early apoptotic
cells; upper left quadrant, necrotic cells; upper right quadrant,
late apoptotic cells. (C) Western blot analysis of
apoptosis-associated proteins after treatment with 17-ABAG. DU-145
cells were treated for the indicated times with 1 μM
17-ABAG. The DU-145 cells were treated for the indicated periods of
time with 1 μM 17-ABAG. The expression levels of Bcl-2 and
Bax were determined by western blot analysis. β-actin was used for
normalization and verification of protein loading. PI, propidium
iodide; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X protein;
FITC, fluorescein isothiocyanate. |
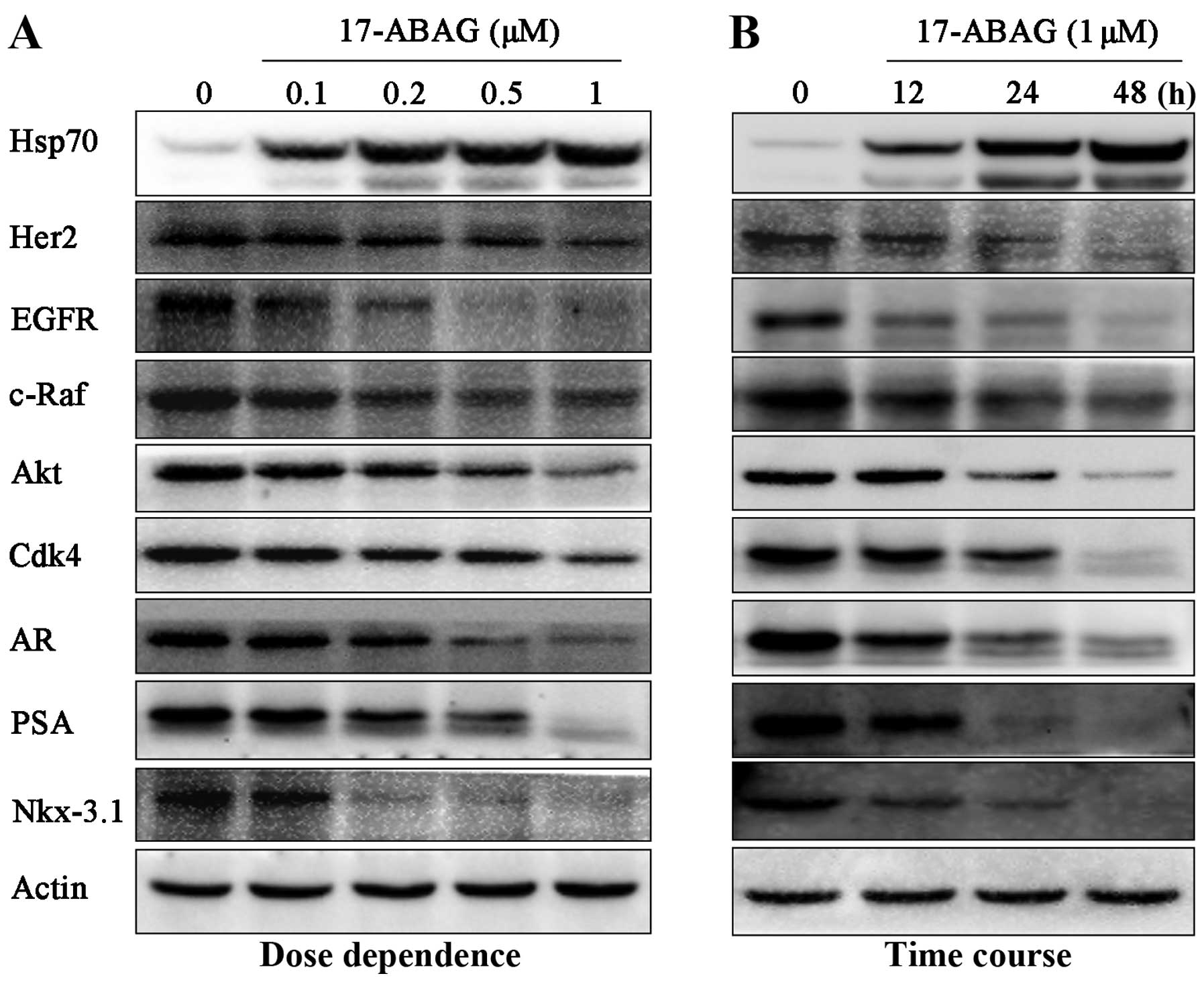
17-ABAG downregulates Hsp90 client
proteins
GA has been identified as an Hsp90 inhibitor via
facilitating the degradation of Hsp90 client proteins required for
tumor growth (11,12). To determine whether 17-ABAG can
also regulate Hsp90, the client proteins of Hsp90 were examined,
including Her2, EGFR, c-Raf, AKT, p-AKT and Cdk4. Western blot
analysis demonstrated that these well-known client proteins were
significantly downregulated following treatment with 17-ABAG in a
time- and dose-dependent manner (Fig.
3), demonstrating that 17-ABAG induces Hsp90 client protein
degradation.
17-ABAG inhibits AR signaling in LNCaP
cells
AR has been reported to be a client protein of Hsp90
(5). Therefore, the present study
next evaluated the effects of 17-ABAG on AR signaling in LNCaP
cells. 17-ABAG was shown to induce AR downregulation in LNCaP cells
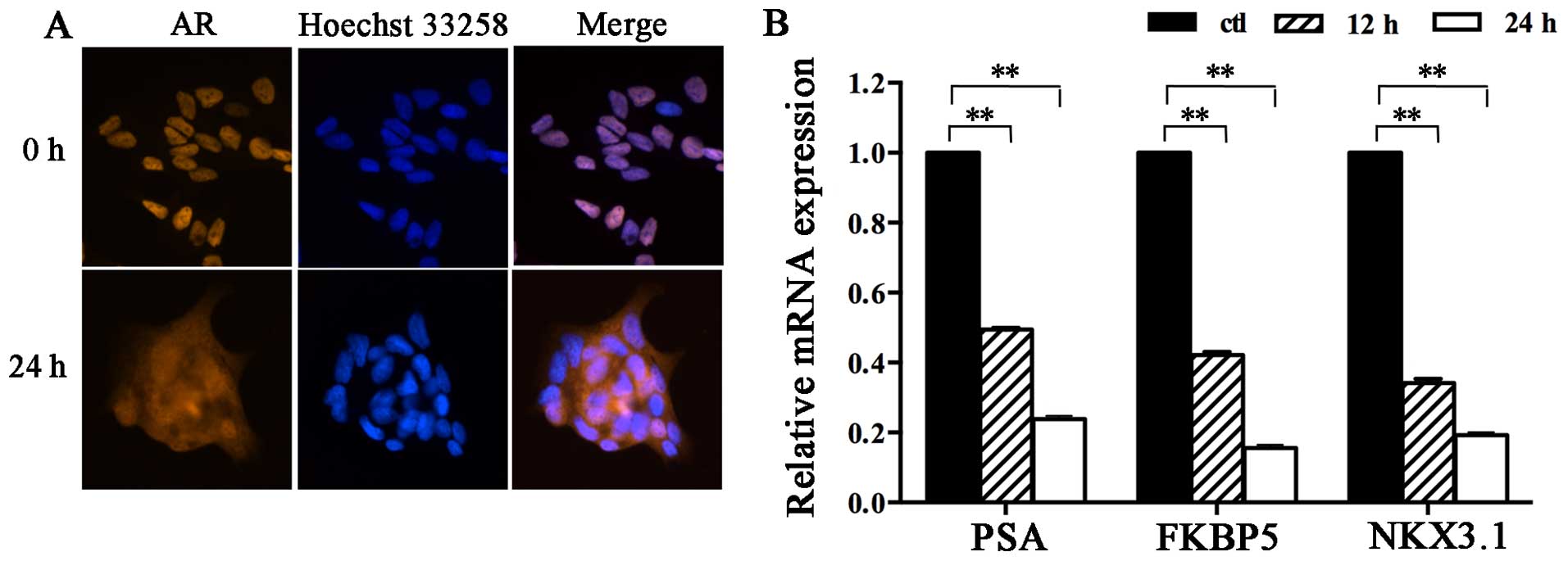
(Fig. 3). Furthermore,
immunofluorescence staining indicated that AR nuclear translocation
was blocked after treatment with 17-ABAG. Abundant AR residues were
observed in the nucleus following staining with R1881, a
photoaffinity label of AR, while the nuclear translocation of ARs
was significantly inhibited by pre-treatment with 17-ABAG (Fig. 4A). This observation indicated that
17-ABAG inhibited the nuclear localization of AR in LNCaP cells
in vitro, which was consistent with the results of a
previous study (5).
To further confirm the effect of 17-ABAG on AR
function, the mRNA levels of several well-characterized
androgen-regulated genes were measured in LNCaP cells. RT-qPCR
indicated that 17-ABAG significantly decreased the mRNA levels of
AR target genes, including PSA, NKX3.1 and FKBP5 (Fig. 4B), suggesting that AR
transcriptional activity was blocked by 17-ABAG treatment. The
protein levels of the AR target genes PSA and NKX3.1 were also
examined. Western blot analysis demonstrated that the expression
levels of these AR target genes were significantly downregulated
following treatment with 17-ABAG in a time- and dose-dependent
manner (Fig. 3).
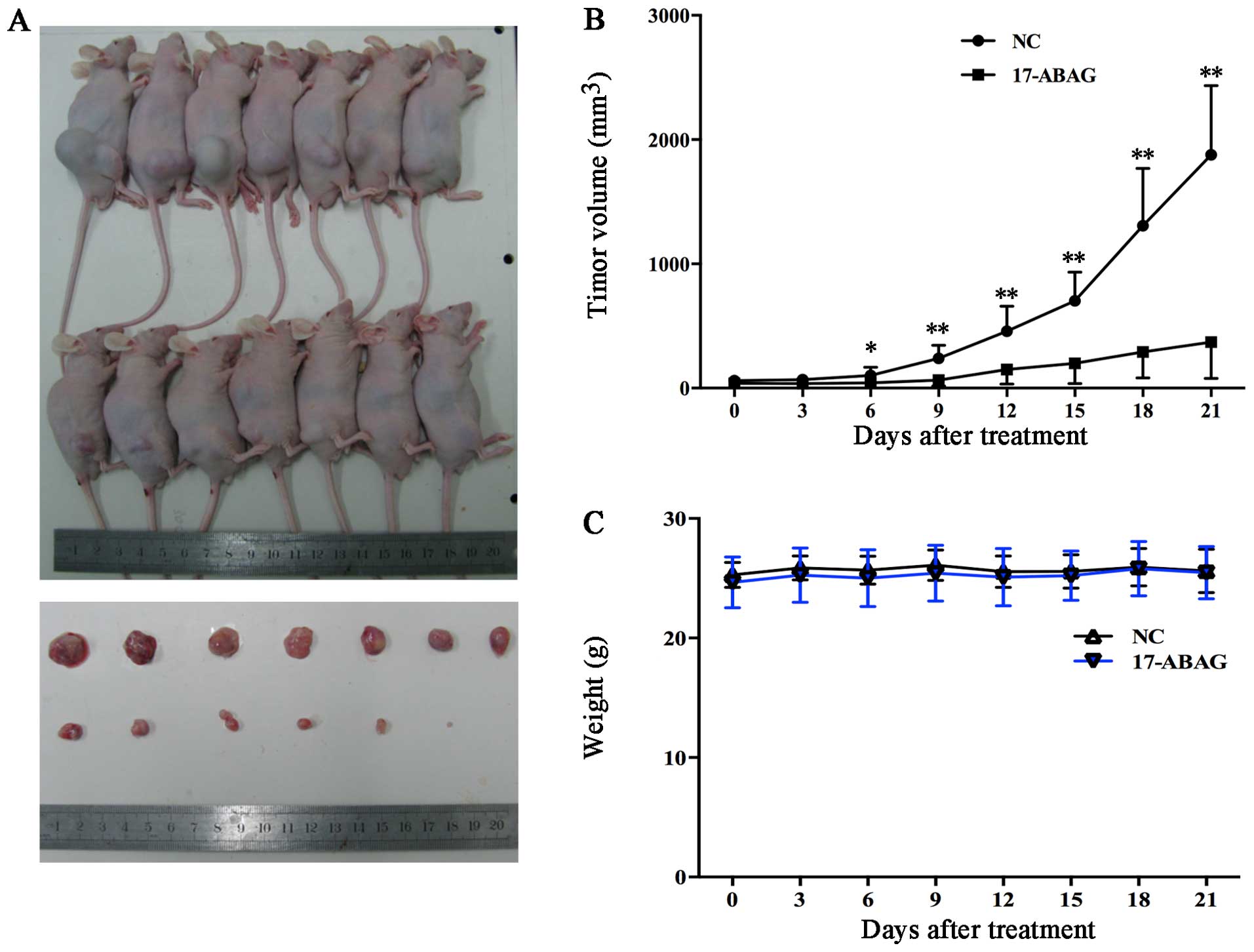
17-ABAG is a promising anti-tumor agent
in vivo
Based on the potent inhibitory in vitro
effects of 17-ABAG on the LNCaP cell line, the in vivo
anti-tumor activity of 17-ABAG was evaluated using prostate cancer
xenografts of LNCaP cells.
LNCaP cells were sub-cutaneously inoculated into
male nude mice, which received an injection of either vehicle
control or 17-ABAG (10 mg/kg every three days). The animals treated
with 17-ABAG (n=7) exhibited a significantly lower average tumor
volume compared with the control mice from day six onwards (42.68
vs. 102.08 mm3, respectively; P<0.05) (Fig. 5A). After 21 days’ treatment, the
average tumor volume was 370.09 mm3 for treated mice
compared with 1,876.87 mm3 for control mice (Fig. 5A and B). When each animal was
considered individually, the incidence of mice progressing with a
tumor volume of 900 mm3 or greater was significantly
diminished by day 21 in 17-ABAG-treated animals (0/7; 0%) compared
with controls (7/7; 100%). No obvious side-effects or body weight
loss were observed (Fig. 5C).
Immunohistochemical analysis indicated decreased Ki67 expression
after treatment with 17-ABAG in the tumors in vivo (Fig. 5D). The observed inhibition of
tumor progression by 17-ABAG may have resulted from decreased
proliferation (reduced Ki67, the proliferation marker). More
importantly, no damage to the organs was detected, including the
heart, liver, spleen, lung and kidney (Fig. 5E). These results showed that
17-ABAG was successful in suppressing the growth of LNCaP xenograft
tumors. These findings suggested that 17-ABAG suppressed prostate
tumor growth in vivo without any observable
side-effects.
Discussion
Prostate cancer is one of the most common cancer
types in males worldwide, occurring more commonly in the developed
world and at increasing rates in developing countries (23,24). Beginning with Huggins and Hodges
(25) first reporting the
susceptibility of prostate cancer to androgen withdrawal, hormonal
therapy remains the most effective therapy for patients with
advanced prostate cancer. However, after 12–18 months on average, a
large percentage of prostate cancer patients will eventually
progress to a castration-resistant stage and succumb to the disease
(median survival time, ~1–2 years) (2,26).
There are limited therapeutic options available for
castrate-resistant prostate cancer (CRPC). Chemotherapy drugs,
immunotherapy and vaccine therapy exhibit limited efficacy and
limited improvement in survival (27). Thus, novel approaches for the
treatment of patients with advanced disease are still urgently
required. With a deeper understanding of the molecular mechanisms
of the tumorigenesis and progression of prostate cancer, numerous
approaches, preferably based on selective targeting of
mechanistically relevant cancer proteins, are currently being
evaluated to improve the treatment of prostate cancer (28).
Hsp90 is a molecular chaperone that maintains the
normal activity of cells through ensuring the proper folding,
maturation, conformational stabilization and location of its client
proteins (3,29). However, through a variety of
oncoproteins, Hsp90 can regulate the progression of tumor cells and
affect their survival, proliferation, invasion and metastasis
(4,29–31). Previous studies have shown that in
prostate cancer, Hsp90 regulates the nuclear localization and
activation of ARs (5,32,33), which has a key role in prostate
carcinogenesis and progression (34,35). The mechanisms of Hsp90 action in
prostate cancer are likely to be complex, as AKT and Her-2
signaling are also important pathways in prostate cancer and, of
note, are Hsp90 client proteins (5,36).
Cancer has six major hallmarks (37), and Hsp90 inhibitors appear to be
the only cancer chemotherapeutic agents known to produce strong
combinatorial effects on all of the hallmarks of cancer
simultaneously (38). In
addition, the multiple downstream effects markedly reduce the
opportunities for cancer cells to develop resistance to Hsp90
inhibitors (39,40). Although Hsp90 is widely expressed
in various cell types, the inhibition of Hsp90 can selectively kill
cancer cells with little effect on normal cells (3,41).
Thus, Hsp90 has been considered as a novel molecular target in
prostate cancer (5,42).
GA was the first reported natural product inhibitor
of Hsp90; however, it exhibits hepatotoxicity at its effective
concentrations, thus limiting its clinical use. A number of Hsp90
inhibitors have demonstrated significant anti-tumor effects in
multiple cancer models, many of which have been evaluated in
clinical trials (15–17). However, studies have suggested
that most of these inhibitors are not efficacious in monotherapy
and have certain disadvantages (15–17,43–45). For example, IPI-504, a derivative
of geldanamycin, showed a minimal effect on the PSA level or tumor
burden and was associated with unacceptable toxicity (17). In previous studies by our group,
>200 GA derivatives were designed and synthesized (14,18,19), among which 17-ABAG inhibited the
proliferation and induced apoptosis of LNCaP cells in vitro
as well as in vivo.
The present study demonstrated that 17-ABAG can
selectively inhibit androgen-dependent (LNCaP) and
androgen-independent (DU-145 and PC-3) prostate cancer cells with
markedly lower cytotoxicity against normal prostate cells (RWPE-1)
in vitro. In addition, the present study showed that 17-ABAG
was able to induce LNCaP cell apoptosis through the regulation of
apoptosis-associated proteins. As an Hsp90 inhibitor, 17-ABAG, also
downregulated the protein levels of Hsp90 client proteins,
including Her2, EGFR, AKT, c-Raf and Cdk4 in a time- and
dose-dependent manner, which may partly account for the mechanism
of the anti-proliferative activity of 17-ABAG against
androgen-dependent prostate cancer cells.
Previous studies have demonstrated that in prostate
cancer, Hsp90 regulates the nuclear localization and activation of
ARs (5,32,33), which have a key role in prostate
carcinogenesis and progression (34,35). The present study demonstrated that
the localization of ARs shifted from predominantly nuclear to
cytoplasmic after treatment with 17-ABAG, consistent with the
results of previous studies. 17-ABAG also decreased the protein
levels of ARs and decreased the mRNA and protein levels of AR
target genes, including PSA, NKX3.1 and FKBP5.
To determine whether 17-ABAG shows any anti-tumor
effects in vivo, LNCaP cells were subcutaneously inoculated
into male nude mice, and tumor-bearing animals were treated via
vein injection with 17-ABAG after the development of visible
tumors. It was observed that 17-ABAG showed significant anti-tumor
effects compared with the vehicle control treatment. In addition,
no obvious side effects or organ damage were detected, suggesting
that 17-ABAG treatment may be a potential safe treatment for
prostate cancer.
In conclusion, 17-ABAG inhibited the proliferation
of LNCaP cells in vitro and in vivo by decreasing the
expression of Hsp90 client proteins, inhibiting the AR signaling
pathway and inducing apoptosis. The results of the present study
suggested that 17-ABAG may be a potential safe treatment for
prostate cancer; however, its clinical potential requires
validation in prospective studies.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (no. 81302214), the Shanghai Nature
Science Foundation of Shanghai Science and Technology Committee
(no. 13ZR1432700), the Shanghai Key Medical Specialty Program (no.
ZK2012A22) and the Shanghai Minhang Health Bureau Foundation (no.
2012MW01).
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moul JW and Dawson N: Quality of life
associated with treatment of castration-resistant prostate cancer:
A review of the literature. Cancer Invest. 30:1–12. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kamal A, Thao L, Sensintaffar J, Zhang L,
Boehm MF, Fritz LC and Burrows FJ: A high-affinity conformation of
Hsp90 confers tumour selectivity on Hsp90 inhibitors. Nature.
425:407–410. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gorska M, Popowska U, Sielicka-Dudzin A,
Kuban-Jankowska A, Sawczuk W, Knap N, Cicero G and Wozniak F:
Geldanamycin and its derivatives as Hsp90 inhibitors. Front Biosci
(Landmark Ed). 17:2269–2277. 2012. View
Article : Google Scholar
|
|
5
|
Ischia J, Saad F and Gleave M: The promise
of heat shock protein inhibitors in the treatment of castration
resistant prostate cancer. Curr Opin Urol. 23:194–200. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Weinstein IB and Joe AK: Mechanisms of
disease: Oncogene addiction - a rationale for molecular targeting
in cancer therapy. Nat Clin Pract Oncol. 3:448–457. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
da Rocha Dias S, Friedlos F, Light Y,
Springer C, Workman P and Marais R: Activated B-RAF is an Hsp90
client protein that is targeted by the anticancer drug
17-allylamino-17-demethoxygel-danamycin. Cancer Res.
65:10686–10691. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Grbovic OM, Basso AD, Sawai A, Ye Q,
Friedlander P, Solit D and Rosen N: V600E B-Raf requires the Hsp90
chaperone for stability and is degraded in response to Hsp90
inhibitors. Proc Natl Acad Sci USA. 103:57–62. 2006. View Article : Google Scholar :
|
|
9
|
Whitesell L, Bagatell R and Falsey R: The
stress response: Implications for the clinical development of hsp90
inhibitors. Curr Cancer Drug Targets. 3:349–358. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chiosis G and Neckers L: Tumor selectivity
of Hsp90 inhibitors: The explanation remains elusive. ACS Chem
Biol. 1:279–284. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Scheibel T and Buchner J: The Hsp90
complex - a super-chaperone machine as a novel drug target. Biochem
Pharmacol. 56:675–682. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Stebbins CE, Russo AA, Schneider C, Rosen
N, Hartl FU and Pavletich NP: Crystal structure of an
Hsp90-geldanamycin complex: Targeting of a protein chaperone by an
antitumor agent. Cell. 89:239–250. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fukuyo Y, Hunt CR and Horikoshi N:
Geldanamycin and its anti-cancer activities. Cancer Lett.
290:24–35. 2010. View Article : Google Scholar
|
|
14
|
Li Z, Jia L, Wang J, Wu X, Hao H, Xu H, Wu
Y, Shi G, Lu C and Shen Y: Design, synthesis and biological
evaluation of 17-arylmethylamine-17-demethoxygeldanamycin
derivatives as potent Hsp90 inhibitors. Eur J Med Chem. 85:359–370.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Heath EI, Hillman DW, Vaishampayan U,
Sheng S, Sarkar F, Harper F, Gaskins M, Pitot HC, Tan W, Ivy SP, et
al: A phase II trial of 17-allylamino-17-demethoxygeldanamycin in
patients with hormone-refractory metastatic prostate cancer. Clin
Cancer Res. 14:7940–7946. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pacey S, Wilson RH, Walton M, Eatock MM,
Hardcastle A, Zetterlund A, Arkenau HT, Moreno-Farre J, Banerji U,
Roels B, et al: A phase I study of the heat shock protein 90
inhibitor alvespimycin (17-DMAG) given intravenously to patients
with advanced solid tumors. Clin Cancer Res. 17:1561–1570. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Oh WK, Galsky MD, Stadler WM, Srinivas S,
Chu F, Bubley G, Goddard J, Dunbar J and Ross RW: Multicenter phase
II trial of the heat shock protein 90 inhibitor, retaspimycin
hydrochloride (IPI-504), in patients with castration-resistant
prostate cancer. Urology. 78:626–630. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Z, Jia L, Wang J, Wu X, Shi G, Lu C and
Shen Y: Discovery of novel 17-phenylethyla minegeldanamycin
derivatives as potent Hsp90 inhibitors. Chem Biol Drug Des.
85:181–188. 2015. View Article : Google Scholar
|
|
19
|
Wu Y, Li Z, Wang Z, Xu H, Wu X, Lu C and
Shen Y: Synthesis of novel 17-[3,6-Dioxa-8-N-(substituted
cinnamyol)- octanediamino]-17-demethoxygeldanamycin derivatives.
Chin J Org Chem. 34:1870–1874. 2014.In Chinese. View Article : Google Scholar
|
|
20
|
Mosmann T: Rapid colorimetric assay for
cellular growth and survival: Application to proliferation and
cytotoxicity assays. J Immunol Methods. 65:55–63. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mahdavinezhad A, Mousavi-Bahar SH,
Poorolajal J, Yadegarazari R, Jafari M, Shabab N and Saidijam M:
Evaluation of miR-141, miR-200c, miR-30b Expression and
Clinicopathological Features of Bladder Cancer. Int J Mol Cell Med.
4:32–39. 2015.PubMed/NCBI
|
|
22
|
Shen P, Sun J, Xu G, Zhang L, Yang Z, Xia
S, Wang Y, Liu Y and Shi G: KLF9, a transcription factor induced in
flutamide-caused cell apoptosis, inhibits AKT activation and
suppresses tumor growth of prostate cancer cells. Prostate.
74:946–958. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Baade PD, Youlden DR and Krnjacki LJ:
International epidemiology of prostate cancer: Geographical
distribution and secular trends. Mol Nutr Food Res. 53:171–184.
2009. View Article : Google Scholar
|
|
24
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huggins C and Hodges CV: Studies on
prostatic cancer. I. The effect of castration, of estrogen and
androgen injection on serum phosphatases in metastatic carcinoma of
the prostate. CA Cancer J Clin. 22:232–240. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Asmane I, Céraline J, Duclos B, Rob L,
Litique V, Barthélémy P, Bergerat JP, Dufour P and Kurtz JE: New
strategies for medical management of castration-resistant prostate
cancer. Oncology. 80:1–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lassi K and Dawson NA: Update on
castrate-resistant prostate cancer: 2010. Curr Opin Oncol.
22:263–267. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Corcoran NM and Gleave ME: Targeted
therapy in prostate cancer. Histopathology. 60:216–231. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pearl LH and Prodromou C: Structure and
mechanism of the Hsp90 molecular chaperone machinery. Annu Rev
Biochem. 75:271–294. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Whitesell L and Lindquist SL: HSP90 and
the chaperoning of cancer. Nat Rev Cancer. 5:761–772. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Takayama S, Reed JC and Homma S:
Heat-shock proteins as regulators of apoptosis. Oncogene.
22:9041–9047. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lamoureux F, Thomas C, Yin MJ, Kuruma H,
Fazli L, Gleave ME and Zoubeidi A: A novel HSP90 inhibitor delays
castrate-resistant prostate cancer without altering serum PSA
levels and inhibits osteoclastogenesis. Clin Cancer Res.
17:2301–2313. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Saporita AJ, Ai J and Wang Z: The Hsp90
inhibitor, 17-AAG, prevents the ligand-independent nuclear
localization of androgen receptor in refractory prostate cancer
cells. Prostate. 67:509–520. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cano LQ, Lavery DN and Bevan CL:
Mini-review: Foldosome regulation of androgen receptor action in
prostate cancer. Mol Cell Endocrinol. 369:52–62. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang CK, Luo J, Lee SO and Chang C:
Concise review: Androgen receptor differential roles in
stem/progenitor cells including prostate, embryonic, stromal, and
hematopoietic lineages. Stem Cells. 32:2299–2308. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Johnson VA, Singh EK, Nazarova LA,
Alexander LD and McAlpine SR: Macrocyclic inhibitors of hsp90. Curr
Top Med Chem. 10:1380–1402. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang H and Burrows F: Targeting multiple
signal transduction pathways through inhibition of Hsp90. J Mol Med
(Berl). 82:488–499. 2004. View Article : Google Scholar
|
|
39
|
Workman P, Burrows F, Neckers L and Rosen
N: Drugging the cancer chaperone HSP90: Combinatorial therapeutic
exploitation of oncogene addiction and tumor stress. Ann NY Acad
Sci. 1113:202–216. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pearl LH, Prodromou C and Workman P: The
Hsp90 molecular chaperone: An open and shut case for treatment.
Biochem J. 410:439–453. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Eskew JD, Sadikot T, Morales P, Duren A,
Dunwiddie I, Swink M, Zhang X, Hembruff S, Donnelly A, Rajewski RA,
et al: Development and characterization of a novel C-terminal
inhibitor of Hsp90 in androgen dependent and independent prostate
cancer cells. BMC Cancer. 11:4682011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Centenera MM, Fitzpatrick AK, Tilley WD
and Butler LM: Hsp90: Still a viable target in prostate cancer.
Biochim Biophys Acta. 1835:211–218. 2013.PubMed/NCBI
|
|
43
|
Porter JR, Fritz CC and Depew KM:
Discovery and development of Hsp90 inhibitors: A promising pathway
for cancer therapy. Curr Opin Chem Biol. 14:412–420. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Soga S, Akinaga S and Shiotsu Y: Hsp90
inhibitors as anti-cancer agents, from basic discoveries to
clinical development. Curr Pharm Des. 19:366–376. 2013. View Article : Google Scholar
|
|
45
|
Price JT, Quinn JM, Sims NA, Vieusseux J,
Waldeck K, Docherty SE, Myers D, Nakamura A, Waltham MC, Gillespie
MT and Thompson EW: The heat shock protein 90 inhibitor,
17-allylamino-17-demethoxygeldanamycin, enhances osteoclast
formation and potentiates bone metastasis of a human breast cancer
cell line. Cancer Res. 65:4929–4938. 2005. View Article : Google Scholar : PubMed/NCBI
|