Introduction
Radiation therapy is widely utilized for the
treatment of pelvic tumors. The rectum is a high-risk organ for
radiation-induced injury, which is a dose-limiting factor of pelvic
radiotherapy. Chronic damage characterized by connective tissue
growth, tissue remodeling and fibrosis impair rectal function and
worsen the quality of life of cancer patients (1).
Previous studies have demonstrated that radiation
can induce hypoxia via vessel damage and tissue remodeling, which
then promote radiation-induced injury to the lungs, brain and
rectum (2–4). Further studies have indicated that
hypoxic conditions exist in normal rectal tissues following
irradiation. Hypoxia increases hypoxia inducible factor-1α (HIF-1α)
and vascular endothelial growth factor (VEGF) expression, which
aggravates inflammation and injury (4,5).
Furthermore, hypoxia can stabilize HIF-1α expression and stimulate
the transforming growth factor (TGF)-β-dependent synthesis of
collagen and extracellular matrix (ECM) proteins (6). Hypoxia is also an inducer of
reactive oxygen species (ROS) and increased leukocyte migration and
vascular permeability, and it upregulates TGF-β and promotes
collagen formation, which are of vital importance in the
development of fibrosis (7,8).
In addition, ROS exert deleterious effects on neurological organs
through damage to cellular macromolecules and membranes, which is
associated with low levels of antioxidants, high concentrations of
unsaturated fatty acids and the availability of redox-active iron
(9).
Telomeres are specialized structures that protect
chromosomal ends from degradation, end fusion and rearrangement,
and preserve chromosomal integrity and stability (10). Telomerase consists of telomerase
reverse transcriptase (TERT), telomerase-associated proteins, and
the telomerase RNA (TR) component (TERC) (11). The catalytic subunit of the
telomerase holoenzyme, TERT, which is essential to the maintenance
of telomerase, contributes to cell proliferation (12,13), preventing cellular senescence
(14,15), and promoting the repair of damaged
tissue and wound healing (16,17). Hypoxic conditions induce a DNA
damage response by causing telomere damage. In response to this
damage, HIF-1α induces telomerase in order to heal the damaged
chromosome ends (18). TERT also
undergoes subcellular localization to the mitochondria in neurons
upon cellular stress in Alzheimer's disease (AD) and protects
against brain injury (9).
Furthermore, TERT reduces intracellular ROS production and inhibits
ROS-mediated apoptosis (6,19).
However, the role of TERT in radiation-induced
injury to normal tissue and the associatoin between TERT and
hypoxia remain unclear. In this study, we examined TERT expression
in rectal tissue following irradiation and examined its effects on
hypoxia-induced fibroblast apoptosis.
Materials and methods
Cell culture and animals
Human skin fibroblasts (FBs) were purchased from the
Cell Bank of the Chinese Academy of Sciences (Shanghai, China).
Both the FBs and human TERT (hTERT)-overexpressing fibroblasts
(FB-TERT fibroblasts) that were used in this study were not used
for more than 3 months and/or 20 passages. The cells were cultured
in Dulbecco's modified Eagle's medium (DMEM; Biowest, Nuaillé,
France), supplemented with 1% penicillin/streptomycin and 10% fetal
bovine serum (FBS; Biowest) in free gas exchange with the
atmospheric air at 37°C. Exposure to hypoxia (for 24 or 48 h) was
carried out using a tri-gas incubator (37°C, 5% CO2, 93%
N2, 2% O2; YCP-50S; Changsha Huaxi Electronic
Technology Co., Ltd., Hunan, China). We also used the autophagy
inhibitor, 3-methyladenine (3-MA; Selleckchem, Houston, TX, USA),
at the concentration of 5 mM, the ROS scavenger, N-acetylcysteine
(NAC; Sigma, St. Louis, CA, USA) at the concentration of 3 mM, the
p-NF-κB inhibitor, BAY 11-7082 (Selleckchem) at the concentration
of 10 µM and DMSO (Sigma) at the concentration of 10
µM. Thirty female C57BL/6N mice (age, 5 weeks; weight, 15–25
g) were purchased from Shanghai Sippr/BK Laboratory Animals Co.,
Ltd. (Shanghai, China). Prior to irradiation, the mice were housed
in a pathogen-free room under controlled temperature and humidity
for 1 week. Following irradiation, all mice were divided into 6
groups (the control group, and the post-RT group for 1, 7, 14, 28
and 90 days).
X-ray irradiation
X-ray irradiation (RT) was performed with a single
dose of 25 Gy and at a dose rate of 3.69 Gy/min using the Small
Animal Radiation Research Platform (SARRP; Baltimore, MD, USA). The
mice were anesthetized by pentobarbitol sodium (60–80 µl)
and were then irradiation was applied locally to the rectums (10 mm
in diameter). Following irradiation, the mice were housed in cages
and supplied with a standard laboratory diet and water. The study
was performed in strict accordance with the recommendations in the
Guide for the Care and Use of Laboratory Animals of Fudan
University. The protocol was approved by the Committee on the
Ethics of Animal Experiments of Fudan University.
Lentiviral vector production
For construction of the hTERT-expressing lentiviral
vector, a cDNA-coding hTERT (NF-198253.2, 3399-bp cDNA) was
synthesized and cloned into the BamHI and XbaI
restriction endonuclease sites of the pLenO-DCE vector (Invabio
Biotechnology Ltd., Shanghai, China). The pLenO-DCE-hTERT plasmid
was then acquired, which also encodes green fluorescent protein
(GFP). The products were transformed into competent fibroblasts.
The emerged cell colonies were cultured, plasmid DNA was isolated,
digested and sequenced to confirm that the correct clone had been
made. After the correct sequence was confirmed, lentiviral vector
particles were produced in accordance with the manufacturer's
instructions (Invitrogen, Carlsbad, CA, USA). A control vector
driving the expression of GFP (pLenO-DCE-GFP) was also generated.
pRsv-REV, Pmd1g-Prre, Pmd2g and pLenO-DCE-hTERT were co-transfected
into 293T cells (American Type Culture Collection; ATCC, Manassas,
VA, USA) and viral supernatants were harvested 48 and 72 h
post-transfection and concentrated using 4 rounds of
ultracentrifugation. The cell surpernatant, which contained viral
particles, was collected and stored at −80°C. Viral titers were
detected by the double dilution method on 293T cells.
Tissue sampling, histological analysis
and immunohistochemistry
The mice were sacrificed by cervical dislocation.
Rectal tissue samples were collected on days 1, 7, 14, 28, and 90
post-irradiation. The sections were fixed using 4% paraformaldehyde
and then embedded in paraffin. The sections were then cut to 4
µm thickness for hematoxylin and eosin (H&E) staining,
Masson's staining and immunohistochemical analysis. For the
analysis of the severity of irradiation-induced injury and
fibrosis, the levels of mucosal depletion and collagen deposition
were analyzed under a microscope (Nikon, Tokyo, Japan). For
immunohistochemical analysis, the samples were incubated with
anti-HIF-1α polyclonal antibody (1:150; Medical & Biological
Laboratories Co., Ltd., Nagoya, Japan) or anti-hTERT polyclonal
antibody (1:1500; ab183105; Abcam, Cambridge, UK) as the primary
antibody overnight at 4°C. The sections were then incubated with
the secondary antibody (1:500; Abcam) and examined under a
microscope (Nikon, Tokyo, Japan).
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
RNA was isolated using a High Pure RNA Isolation kit
(RNAprep pure cell/bacteria kit; Tiangen, Beijing, China) and
treated with DNase to eliminate genomic DNA contamination. Total
RNA (500 ng) was used as a template for reverse transcription into
cDNA using the PrimeScript™ RT reagent kit (Perfect Real Time;
Takara Bio Inc., Shiga, China), which was then used for qPCR. All
mRNA expression levels were normalized to β-actin. The following
primers were used for TERT: sense, 5′-CGGAAGAGTGTCTGGAGCAA-3′ and
antisense, 5′-GGATGAAGCGGAGTCTGGA-3′. qPCR was performed using
LightCycler 480 SYBR-Green I Master (Roche Diagnostics, Basel,
Switzerland). The ΔΔCt method was used for relative
quantification.
Western blot analysis
Cells were lysed with RIPA lysis buffer (Roche
Diagnostics, Basel, Switzerland), and total protein was quantified
using the Pierce BCA Protein Assay kit (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). For western blot analysis, equal amounts
of protein extracts were separated by sodium dodecyl sulfate
(SDS)-polyacrylamide gel electrophoresis (PAGE) on a 10% (w/v)
polyacrylamide gel, followed by electrotransfer onto a BioTrace NC
Membrane (Pall Corporation, Pensacola, FL, USA). The blots were
blocked for 1 h with blocking buffer [5% (w/v) bovine serum
albumin, 0.1% (v/v) Tween-20 in phosphate-buffered saline (PBS)].
The antibodies used for western blot analysis were as follows:
nuclear factor-κB (NF-κB)/p-NF-κB p65 (2884-1; Epitomics,
Burlingame, CA, USA), LC3 (M152-3; Medical & Biological
Laboratories Co., Ltd.) and TERT (ab183105; Abcam). This was
followed by incubation with horseradish peroxidase-coupled
secondary anti-rabbit (Millipore, Billerica, MA, USA) or anti-mouse
antibodies (GeneTex Inc., Irvine, CA, USA). Protein bands were
detected using the ECL Blotting Detection Reagents (Thermo Fisher
Scientific, Inc.) and imaged and quantified using the Chemioscope
Mini system (Bioshine, Shanghai, China). Nuclear-cytoplasmic
fractionation was conducted using the NE-PER Nuclear and
Cytoplasmic Extraction Reagents kit (Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions.
Silencing of hTERT by RNA
interference
The cells (FB-TERT fibroblasts) were plated prior to
transfection for a period of 24 h. They were then transfected with
hTERT siRNA using X-treme gene siRNA reagent (Roche Diagnostics,
Basel, Switzerland). The siRNAs were synthesized by Shanghai
GenePharma Co., Ltd. (Zhangjiang Hi-Tech Park, Shanghai). The siRNA
sequences were as follows: hTERT siRNA sense,
5′-GGAAGAGUGUCUGGAGCAATT-3′ and antisense,
5′-UUGCUCCAGACACUCUUCCTT-3′. The negative control (scramble) siRNA
sequence was as follows: sense, 5′-UUCU CCGAACGUGUCACGUTT-3′ and
antisense, 5′-ACGUGACACGUUCGGAGAATT-3′.
Telomerase activity assay
Telomerase activity was measured using the
telomerase repeat amplification protocol enzyme-linked
immunosorbent assay (ELISA). The PCR-ELISA kit was provided by
Roche Diagnostics GmbH (Mannheim, Germany). The PCR product was
detected with an antibody against digoxigenin that was conjugated
to peroxidase. The probe was visualized by virtue of
peroxidase-metabolizing TMB color liquid to form a colored reaction
product, and the absorbance of the samples was measured at 450 nm
using an ELISA microtiter reader (Epoch; BioTek Instruments,
Winooski, VT, USA).
Detection of autophagy
The cells were digested and washed twice with PBS
(pH 7.4; HyClone, Logan, UT, USA), centrifuged for 5 min at 37°C,
and resuspended in a binding buffer. Autophagosomes were marked
using the Cyto-ID Autophagy Detection kit (Enzo Life Sciences,
Inc., Farmingdale, NY, USA). Marked cells were photographed under a
fluorescence microscope (Nikon), and the fluorescence intensity was
detected using a flow cytometer (FC500 MPL; Beckman Coulter, Inc.,
Brea, CA, USA).
Detection of apoptosis
The digested cells were washed twice with PBS (pH
7.4; HyClone), centrifuged for 5 min at 37°C, and resuspended in
binding buffer. They were then stained using the Annexin V-FITC
Apoptosis Detection kit, followed by the addition of propidium
iodide (PI) (both from BD Biosciences, San Jose, CA, USA). The
samples were then analyzed by flow cytometry.
ROS generation assay
Intracellular ROS production was measured using
2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) (Beyotime
Institute of Biotechnology, Shanghai, China). The fibroblasts were
seeded in 60-mm culture dishes. The cells were then subjected to
hypoxic conditions for 24 and 48 h and were then incubated with
DCFH-DA (20 mM) for 20 min at 37°C in a dark place. Following
incubation, the cells were harvested using trypsin-EDTA solution.
The cell suspensions were then centrifuged for 5 min at room
temperature and the supernatant was removed. The fluorescence
intensity of DCFH-DA was measured and calculated using a flow
cytometer (FC500 MPL; Beckman Coulter, Inc.).
Measurement of mitochondrial membrane
potential (ΔΨm)
Loss of ΔΨm was assessed by flow cytometry with
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazole-carbocya-nide
iodine (mitochondrial membrane potential assay kit with JC-1;
Beyotime Institute of Biotechnology) staining. The cells were
collected into 1.5 ml tubes and incubated with JC-1 for 20 min at
37°C. The fluorescence was detected at excitation and emission
wavelengths of 525 and 590 nm, respectively. Red emission from the
dye is attributed to a potential-dependent aggregation of JC-1 in
the mitochondrial. Green fluorescence reflects the monomeric form
of JC-1, appearing in the cytoplasm after mitochondrial membrane
depolarization. The ΔΨm of the cells in each treatment group was
calculated as the fluorescence ratio of red to green.
Carbonylcyanide-m-chlorop henylhydrazone (CCCP) is a protonophore,
which can cause the dissipation of ΔΨm and was used as a positive
control.
Assessment of intracellular glutathione
to glutathione disulfide (GSH/GSSG) levels
The cells were seeded for 24 h in a 6-well plate
prior to transfection at 37°C. Following transfection under hypoxic
or normoxic conditions (21% O2), the cells were washed
once with chilled PBS. The GSH assay was optimized from the
protocol of the GSH and GSSG assay kit (Beyotime Institute of
Biotechnology).
Statistical analysis
All experiments were performed at least 3 times for
statistical significance. Numerical data are expressed as the means
± SD. Statistical analysis was performed using the paired Student
t-test. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
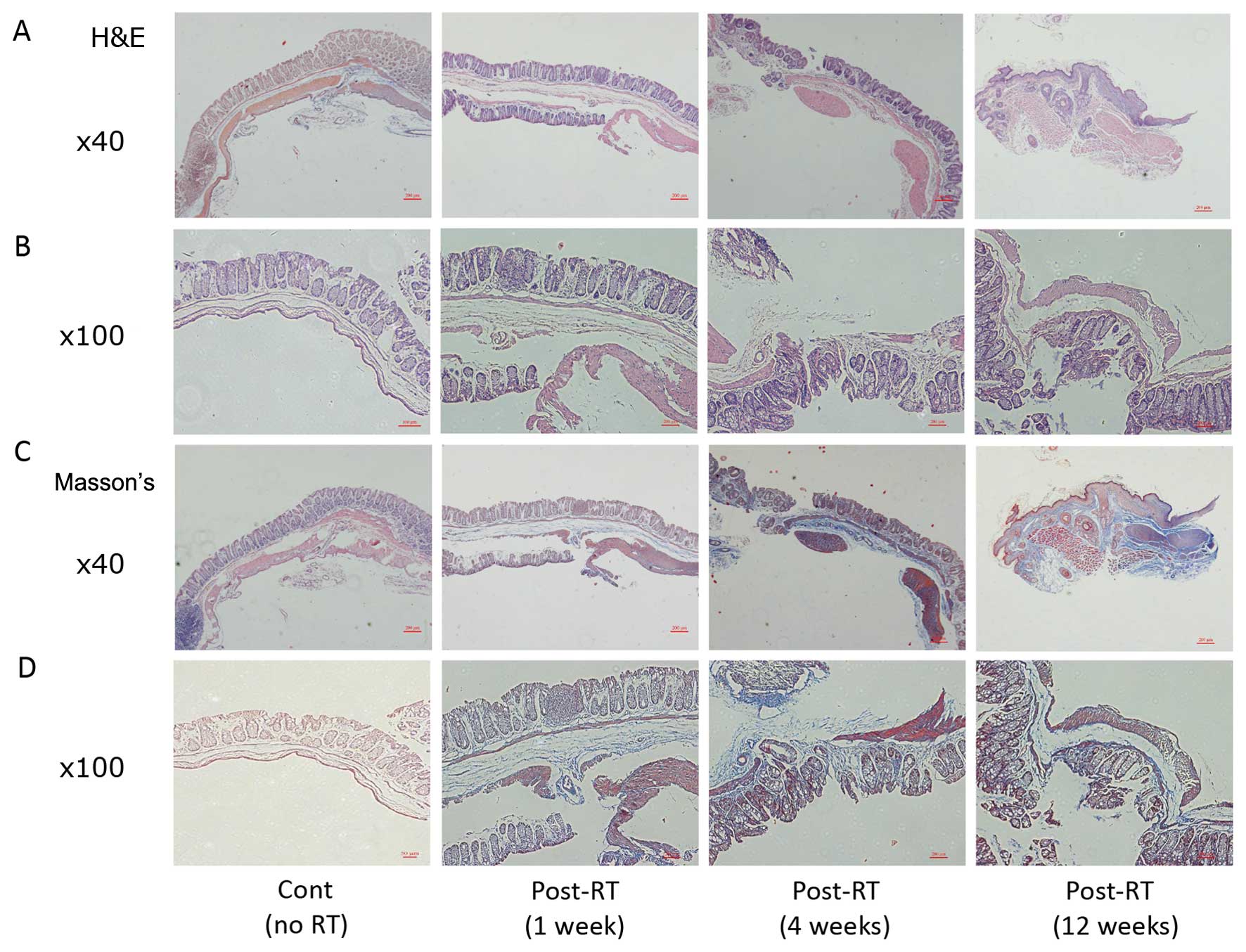
Histological changes in rectal tissue
following radiation
The histological changes following irradiation were
characterized by the loss of the surface epithelium, deep erosion
of the mucosa, thickening of the lamina propria and submucosa
(Fig. 1A and B), collagen
deposition and fibrosis (Fig. 1C and
D). The severity of the damage increased with time. In the
unirradiated control rectal tissue, no such lesions were
observed.
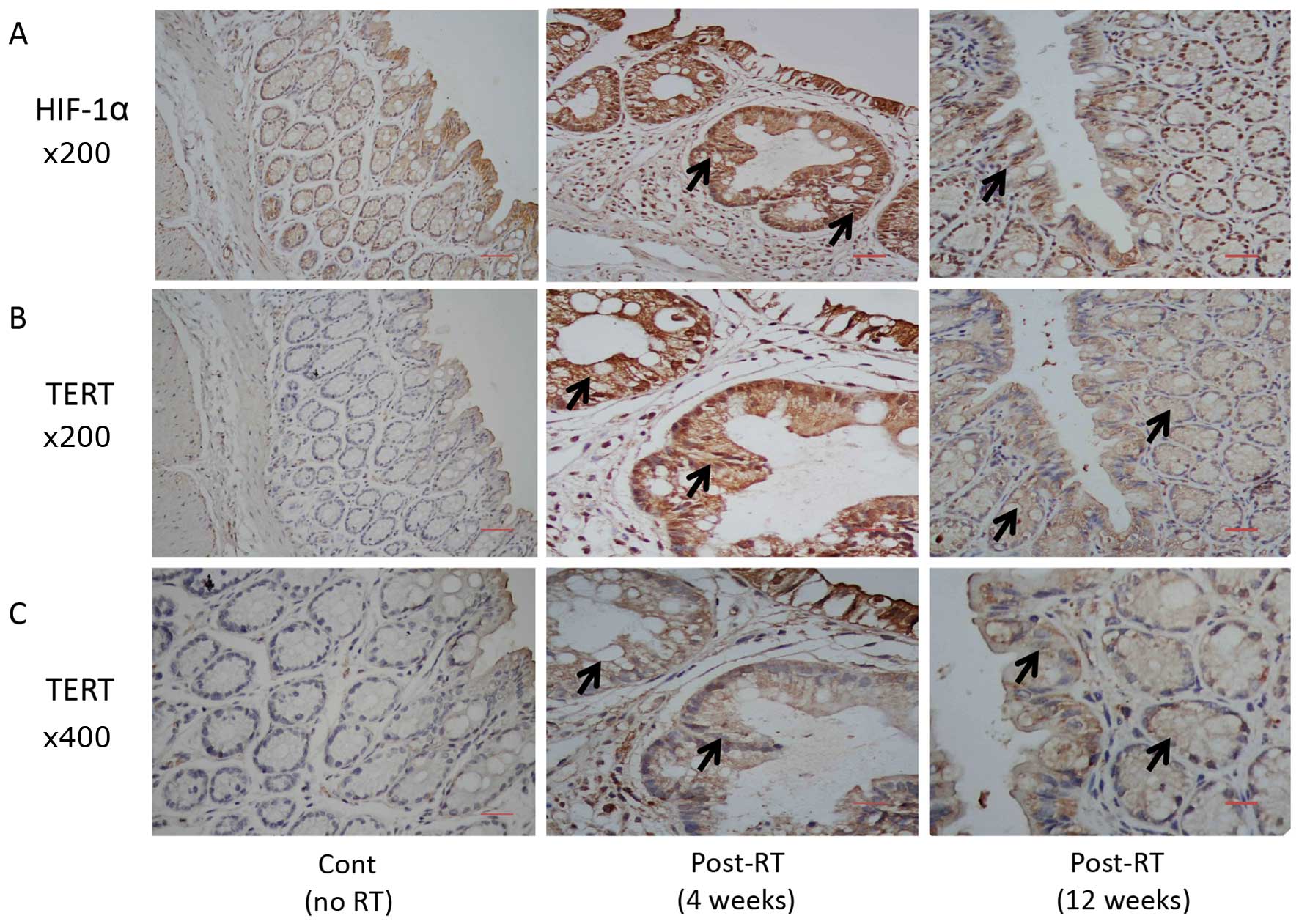
HIF-1α and TERT protein levels in
irradiated rectal tissue
The HIF-1α and TERT protein levels in the irradiated
rectal tissue were analyzed. As shown in Fig. 2, the HIF-1α and TERT protein
levels increased significantly 4 and 12 weeks post-irradiation.
TERT protein translocated from the nucleus to the cytoplasm
following irradiation, which was more evident in the 12 weeks
post-RT group (Fig. 2C). Brown
staining indicated positive staining and increased expression in
the irradiated tissues.
Effect of TERT on apoptosis and ROS
production under hypoxic conditions
The TERT-overexpressing fibroblasts were
successfully generated. The FB-TERT fibroblasts were smaller
(Fig. 3A) than the original
fibroblasts (FBs). To assess the efficiency of transfection, the
relative mRNA expression of hTERT was determined in the FBs and
FB-TERT fibroblasts; hTERT mRNA expression was significantly higher
in the FB-TERT fibroblasts (Fig.
3A). The fibroblasts transfected with TERT siRNA (TERTi)
exhibited a decreased TERT mRNA expression and telomerase activity
(Fig. 3B and C). To determine
whether TERT affects apoptosis under hypoxic conditions, we
examined the apoptosis level following different durations of
exposure to hypoxia. As shown in Fig.
3D, exposure to hypoxic conditions (low oxygen concentrations)
induced cell apoptosis in a time-dependent manner compared to the
cells exposed to normoxic conditions. The TERT-overexpressing
fibroblasts exhibited a significantly reduced percentage of
apoptotic cells under normoxic and hypoxic conditions compared to
the FBs transfected with the empty vector and to those transfected
with hTERT siRNA (Fig. 3D). The
TERT-overexpressing fibroblasts transfected with the negative
control siRNA exhibited a significant reduction in ROS production,
as compared to the cells transfected with hTERT siRNA (Fig. 3E).
Silencing of TERT reduces autophagy
levels in fibroblasts
The activation of autophagy was analyzed on the
basis of the fluorescence intensity of autophagosomes (Fig. 4A) and the expression of LC-3 in
fibroblasts (Fig. 4B). The
fibroblasts transfected with hTERT siRNA exhibited reduced
autophagy levels under hypoxic conditions. Similarly, they also had
a lower ratio of LC3-II to LC3-I expression than that observed in
the FB-TERT fibroblasts transfected with the negative control
siRNA. To evaluate the association between autophagy, ROS and
apoptosis, the autophagy inhibitor, 3-MA, and a ROS scavenger were
used. As shown in Fig. 4C, the
fibroblasts treated with the autophagy inhibitor exhibited
increased ROS production under hypoxic conditions, which indicates
that autophagy leads to the scavenging of redundant ROS. The
fibroblasts treated with the ROS scavenger exhibited moderate
apoptosis. However, the inhibition of autophagy markedly increased
apoptosis, which indicates a protective effect of autophagy against
cell apoptosis by ROS scavenging (Fig. 4D).
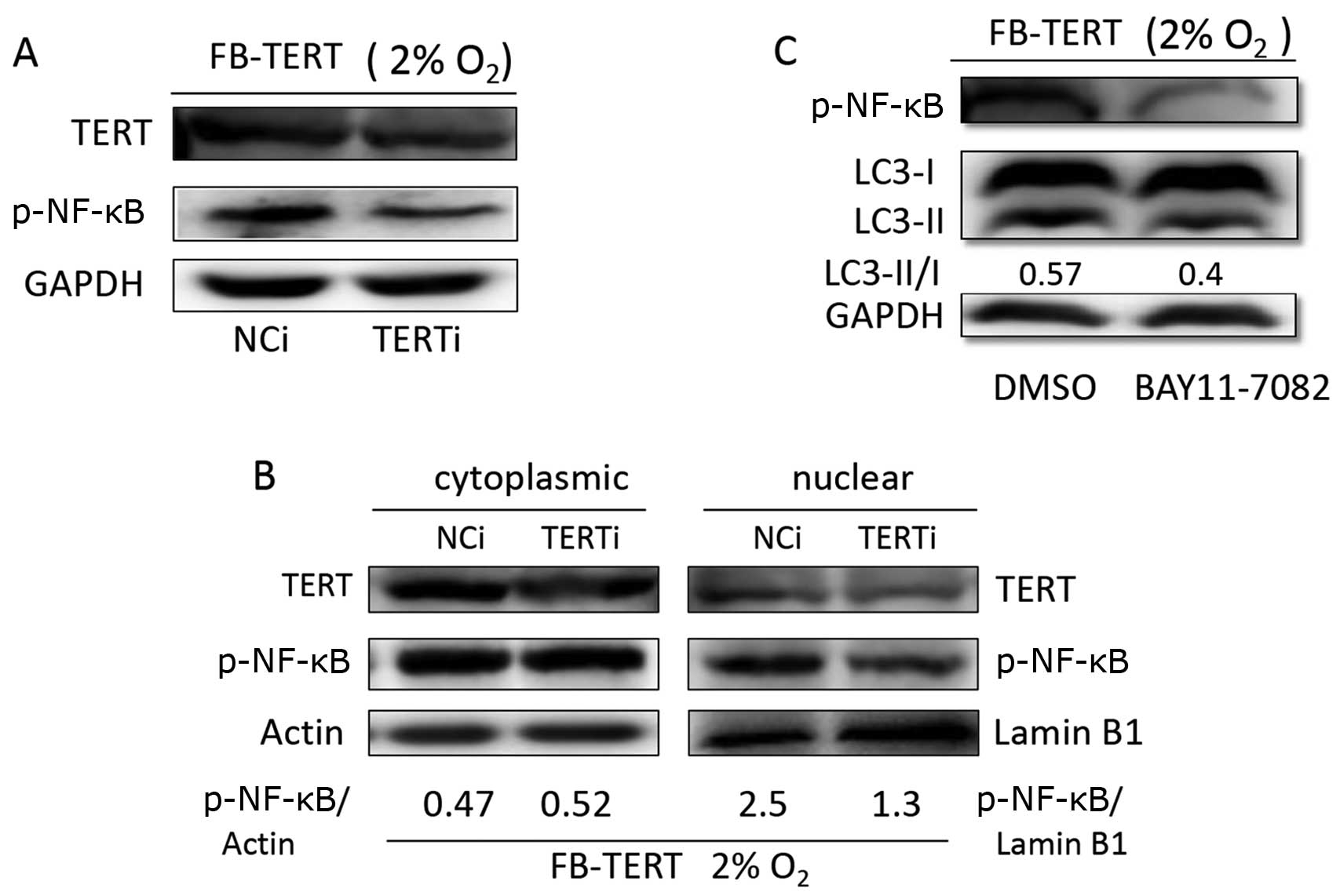
TERT promotes autophagy through p-NF-κB
activation
The fact that TERT promotes autophagy was
demonstrated (Fig. 4A and B), and
thus NF-κB was examined as a medium between TERT and autophagy.
p-NF-κB p65 expression was assessed in the FB-TERT fibroblasts
transfected with hTERT siRNA or negative control siRNA. An evident
reduction in p-NF-κB p65 expression was observed in the fibroblasts
in which hTERT expression was silenced (Fig. 5A). In addition, in the
TERT-overexpressing fibroblasts, p-NF-κB p65 was activated and
translocated to the nucleus, which could be significantly assayed,
while no obvious changes were observed in the cytoplasm (Fig. 5B). At the same time, the
inhibition of p-NF-κB p65 activation by BAY11-7082 reduced the
ratio of LC3-II to LC3-I expression compared to that observed in
the dimethyl sulphoxide (DMSO)-treated group (Fig. 5C), which indicates that TERT
promotes autophagy through the activation of NF-κB.
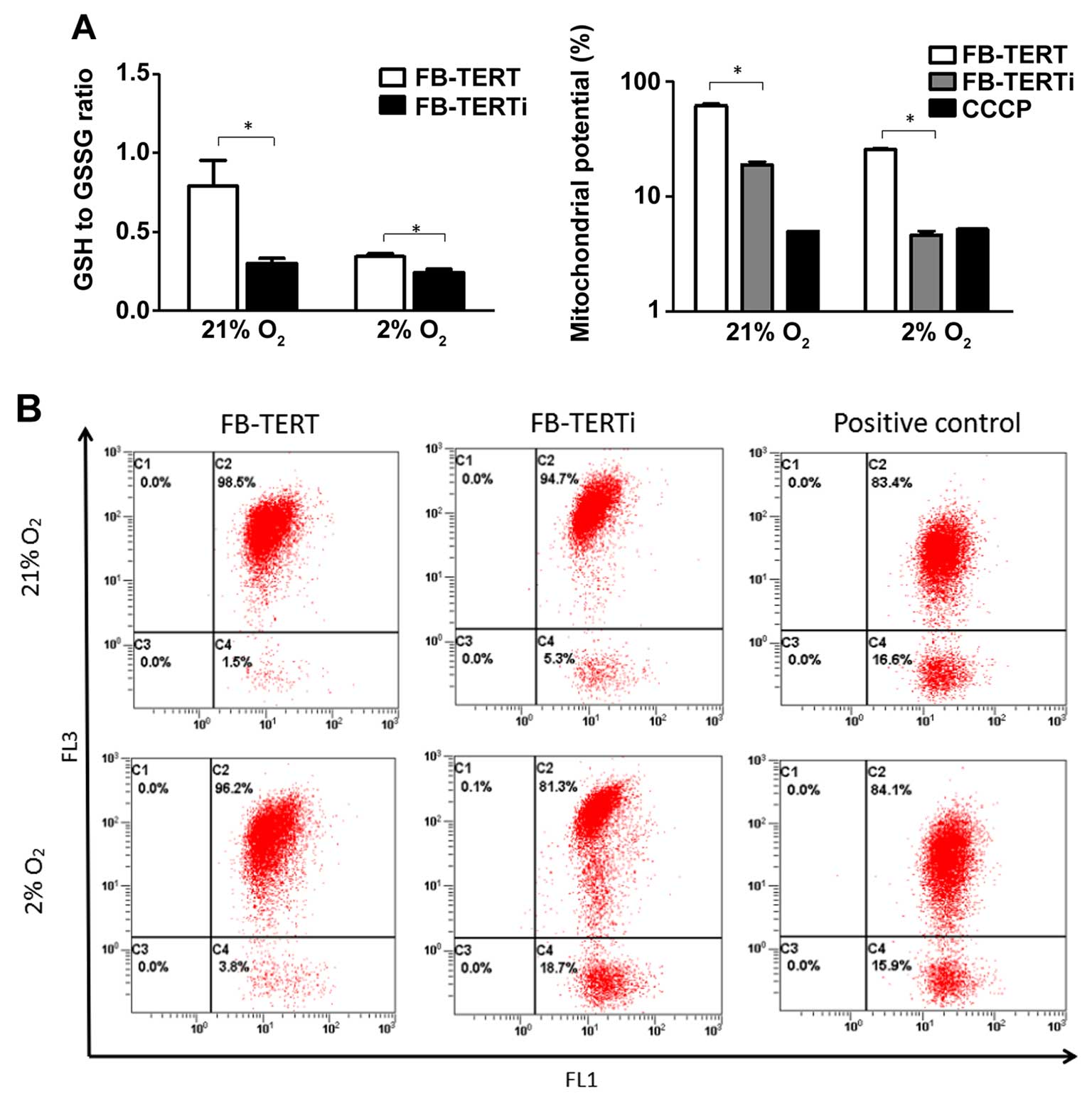
TERT-overexpressing fibroblasts exhibit
an enhanced GSH antioxidant defense capacity and a more stable ΔΨm
under hypoxic conditions
To determine whether TERT affects the cell
antioxidant defense capacity, the ratio of GSH/GSSG was assessed,
which alters cellular capacity in order to resist oxidative stress.
As such, fibroblasts overexpressing TERT or or the FB-TERT
fibroblasts transfected with hTERT siRNA were exposed to normoxic
or hypoxic conditions for 24 h. Exposure to hypoxia reduced the
ratio of GSH/GSSG, and the fibroblasts transfected with hTERT siRNA
maintained a lower ratio of GSH/GSSG (Fig. 6A), which provides evidence
indicating that TERT exerts protective effects on GSH metabolism
and against oxidative insults.
Since the inhibitory effect of TERT on intracellular
ROS production and its protective antioxidant effects had been
established, we then assessed ΔΨm, which provides information about
mitochondrial membrane permeabilization. It has been shown that
hypoxic conditions lead to a decrease in ΔΨm. The decrease in ΔΨm
in the cells transfected with hTERT siRNA was greater than that
observed in the TERT-overexpressing cells (Fig. 6B), which provides further evidence
indicating that TERT expression endows cells with a survival
advantage against ROS production induced by hypoxia by inhibiting
mitochondrial permeabilization.
Discussion
Radiation-induced late injury to normal tissue
surrounding the tumor is a significant dose-limiting factor of
radiation therapy. The pathogenesis of radiation proctitis has not
yet been completed elucidated; it begins with initial
radiation-induced mucosal injury, which is followed by collagen
deposition in the ECM, tissue remodeling and fibrosis, and
subsequently, the tissue response to ischemia (20). Previous studies have demonstrated
that hypoxia following irradiation is an inevitable injury
mechanism that is related to changes in the levels of several
cytokines, such as HIF-1α, VEGF, TGF-β and CD31. The overexpression
of TGF-β induced by irradiation leads to the accumulation of
pathological amounts of the ECM in rectal tissues (4) and lung tissues (21,22). Hypoxia can induce oxidative
damage, mainly caused by ROS, which induces cell death through
mitochondrial dysfunction (23,24).
TERT is essential to the maintenance of telomerase,
which contributes to cell proliferation (12,13). Hypoxia not only transactivates
hTERT promoter activity, but also enhances endogenous hTERT
expression. The introduction of an anti-sense oligonucleotide for
HIF-1α diminishes hTERT expression during hypoxia, indicating that
the upregulation of hTERT by hypoxia is directly mediated through
HIF-1α (18). In the present
study, we demonstrate that increased TERT expression significantly
antagonizes the increase in cellular ROS in response to hypoxia.
GSH loss and oxidation are caused by the increased expression of
the rate-limiting enzyme of GSH synthesis, that is, GCLC. Studies
have proven that an early and sustained induction of GCLC occurs in
response to oxidative stress in hTERT-overexpressing cells
(19,25), which helps to antagonize a rapid
decline in cellular GSH levels, and maintaining a higher GSH/GSSG
level. Furthermore, the partial rescue from the decrease in ΔΨm in
the TERT-overexpressing group provided further evidence that TERT
endows cells with a survival advantage against ROS-mediated cell
toxic effects by inhibiting mitochondrial permeabilization. In
addition to antagonizing cellular ROS, the activation of autophagy
under hypoxic conditions was demonstraed in our study as an
underlying mechanism of the reducing effects of TERT on the
production of ROS.
Hypoxia is an inducer of autophagy, which is a
lysosom-mediated degradation process for non-essential or damaged
cellular constituents and serves to preserve the balance between
organelle biogenesis, protein synthesis and clearance (26). The association between autophagy
and cell death is complex, as autophagy can be involved in either
cell death or survival, depending on the cellular context (27,28). Previous studies have demonstrated
that autophagy can protect cells from radio- and chemotherapy in
breast, pharyngeal, cervical, lung and rectal tumors (29–31). Another study demonstrated that
hypoxia-induced autophagy is also involved in a HIF-1α-mediated
general mechanism of cell survival. BNIP3 and BNIP3L are downstream
targets of HIF-1α-mediated autophagy (24). The inhibition of autophagy by the
addition of chloroquine or the prevention of K63 ubiquitination
increases the formation of ROS (32). Our study demonstrates that
autophagy acts as a survival mechanism in cells exposed to hypoxia
through ROS reduction, which is in agreement with the findings of
these previous studies.
To investigate the mediator between TERT and
autophagy, we hypothesized that NF-κB may be a possible candidate.
The activation of the canonical NF-κB pathway is closely associated
with the regulation of autophagy (33,34). NF-κB, a transcription factor,
which plays an important role in the response to cell damage,
stress and inflammatory reactions, has been reported to have a
unique crosstalk with ROS (35).
The mammalian NF-κB family consists of 5 subunits: RelA/p65, RelB,
c-Rel, p50 and p52, and NF-κB is a heterodimer composed of p65 and
p50. Under normal conditions, NF-κB exists in the cytoplasm of the
cell in an inactive state through interactions with the inhibitor
of NF-κB. When exposed to specific stimuli, NF-κB is activated and
is rapidly transferred to inside the nucleus where it is
responsible for controlling transcription (36). Studies have also reported NF-κB
may be a target of hTERT. Indran et al demonstrated higher
levels of NF-κB p65 in hTERT-overexpressing cells, which also
exhibited a significantly higher level of manganese-dependent
superoxide dismutase (MnSOD) (19). In this study, we uncovered a
reduction of p-NF-κB p65 transfer into the nucleus with a decrease
in TERT expression, indicating an activating effect of TERT on
NF-κB, which is is accordance with these studies.
Taken together, our present study demonstrated that
hypoxia and TERT expression increased in rectal fibrotic tissues
following irradiation. Further experiments revealed that hypoxia
induced cell apoptosis by promoting ROS production. TERT promoted
autophagy by activating p-NF-κB, which enabled the modulation of
intracellular ROS under hypoxic conditions by maintaining the
antioxidant status and mitochondrial function, and blunting
apoptotic signals. In the future, studies on the effects of hypoxia
and TERT in animal models are warranted. TERT may prove to be
target with which to enhance the anti-apoptotic ability of cells by
enhancing the cellular autophagy level, which may be beneficial for
the reduction of radiation-induced injury to normal tissue.
Acknowledgments
This study was supported by the Scientific Research
Foundation for the Returned Overseas Chinese Scholars (no. N130204)
from the China State Education Ministry, the National Natural
Science Foundation of China (nos. 81202148 and 31370838), the
Shanghai Pujiang Program (no. 13P1401600), and the Foundation of
Shanghai Committee of Science and Technology of China (no.
12DZ2260100).
References
|
1
|
Grodsky MB and Sidani SM: Radiation
proctopathy. Clin Colon Rectal Surg. 28:103–111. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Christofidou-Solomidou M, Pietrofesa RA,
Arguiri E, Schweitzer KS, Berdyshev EV, McCarthy M, Corbitt A,
Alwood JS, Yu Y and Globus RK: Space radiation-associated lung
injury in a murine model. Am J Physiol Lung Cell Mol Physiol.
308:L416–L428. 2015. View Article : Google Scholar :
|
|
3
|
Nordal RA, Nagy A, Pintilie M and Wong CS:
Hypoxia and hypoxia-inducible factor-1 target genes in central
nervous system radiation injury: a role for vascular endothelial
growth factor. Clin Cancer Res. 10:3342–3353. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu Y, Kudo K, Abe Y, Aoki M, Hu DL,
Kijima H and Nakane A: Hypoxia expression in radiation-induced late
rectal injury. J Radiat Res (Tokyo). 49:261–268. 2008. View Article : Google Scholar
|
|
5
|
Liu Y, Kudo K, Abe Y, Hu DL, Kijima H,
Nakane A and Ono K: Inhibition of transforming growth factor-beta,
hypoxia-inducible factor-1alpha and vascular endothelial growth
factor reduced late rectal injury induced by irradiation. J Radiat
Res (Tokyo). 50:233–239. 2009. View Article : Google Scholar
|
|
6
|
Distler JH, Jüngel A, Pileckyte M, Zwerina
J, Michel BA, Gay RE, Kowal-Bielecka O, Matucci-Cerinic M, Schett
G, Marti HH, et al: Hypoxia-induced increase in the production of
extracellular matrix proteins in systemic sclerosis. Arthritis
Rheum. 56:4203–4215. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Haroon ZA, Raleigh JA, Greenberg CS and
Dewhirst MW: Early wound healing exhibits cytokine surge without
evidence of hypoxia. Ann Surg. 231:137–147. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schmidt-Ullrich RK, Dent P, Grant S,
Mikkelsen RB and Valerie K: Signal transduction and cellular
radiation responses. Radiat Res. 153:245–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Spilsbury A, Miwa S, Attems J and Saretzki
G: The role of telomerase protein TERT in Alzheimer's disease and
in tau-related pathology in vitro. J Neurosci. 35:1659–1674. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Greider CW: Chromosome first aid. Cell.
67:645–647. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
De Boeck G, Forsyth RG, Praet M and
Hogendoorn PC: Telomere-associated proteins: cross-talk between
telomere maintenance and telomere-lengthening mechanisms. J Pathol.
217:327–344. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ahmed S, Passos JF, Birket MJ, Beckmann T,
Brings S, Peters H, Birch-Machin MA, von Zglinicki T and Saretzki
G: Telomerase does not counteract telomere shortening but protects
mitochondrial function under oxidative stress. J Cell Sci.
121:1046–1053. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Masutomi K, Possemato R, Wong JM, Currier
JL, Tothova Z, Manola JB, Ganesan S, Lansdorp PM, Collins K and
Hahn WC: The telomerase reverse transcriptase regulates chromatin
state and DNA damage responses. Proc Natl Acad Sci USA.
102:8222–8227. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jan HM, Wei MF, Peng CL, Lin SJ, Lai PS
and Shieh MJ: The use of polyethylenimine-DNA to topically deliver
hTERT to promote hair growth. Gene Ther. 19:86–93. 2012. View Article : Google Scholar
|
|
15
|
Qu Y, Duan Z, Zhao F, Wei D, Zhang J, Tang
B, Li J, Yang C and Mu D: Telomerase reverse transcriptase
upregulation attenuates astrocyte proliferation and promotes
neuronal survival in the hypoxic-ischemic rat brain. Stroke.
42:3542–3550. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tao X, Ming-Kun Y, Wei-Bin S, Hai-Long G,
Rui K and Lai-Yong T: Role of telomerase reverse transcriptase in
glial scar formation after spinal cord injury in rats. Neurochem
Res. 38:1914–1920. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Osanai M, Tamaki T, Yonekawa M, Kawamura A
and Sawada N: Transient increase in telomerase activity of
proliferating fibroblasts and endothelial cells in granulation
tissue of the human skin. Wound Repair Regen. 10:59–66. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nishi H, Nakada T, Kyo S, Inoue M, Shay JW
and Isaka K: Hypoxia-inducible factor 1 mediates upregulation of
telomerase (hTERT). Mol Cell Biol. 24:6076–6083. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Indran IR, Hande MP and Pervaiz S: hTERT
overexpression alleviates intracellular ROS production, improves
mitochondrial function, and inhibits ROS-mediated apoptosis in
cancer cells. Cancer Res. 71:266–276. 2011. View Article : Google Scholar
|
|
20
|
Okunieff P, Cornelison T, Mester M, Liu W,
Ding I, Chen Y, Zhang H, Williams JP and Finkelstein J: Mechanism
and modification of gastrointestinal soft tissue response to
radiation: role of growth factors. Int J Radiat Oncol Biol Phys.
62:273–278. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang S, Zhang M, Chen C, Cao Y, Tian Y,
Guo Y, Zhang B, Wang X, Zhang L, et al: Triptolide mitigates
radiation-induced pulmonary fibrosis. Radiat Res. 184:509–517.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Martin M, Lefaix J and Delanian S:
TGF-beta1 and radiation fibrosis: a master switch and a specific
therapeutic target? Int J Radiat Oncol Biol Phys. 47:277–290. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Murphy MP: How mitochondria produce
reactive oxygen species. Biochem J. 417:1–13. 2009. View Article : Google Scholar
|
|
24
|
Lee J, Giordano S and Zhang J: Autophagy,
mitochondria and oxidative stress: cross-talk and redox signalling.
Biochem J. 441:523–540. 2012. View Article : Google Scholar :
|
|
25
|
Meister A and Anderson ME: Glutathione.
Annu Rev Biochem. 52:711–760. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun Y, Xing X, Liu Q, Wang Z, Xin Y, Zhang
P, Hu C and Liu Y: Hypoxia-induced autophagy reduces
radiosensitivity by the HIF-1α/miR-210/Bcl-2 pathway in colon
cancer cells. Int J Oncol. 46:750–756. 2015.
|
|
27
|
Kroemer G and Jäättelä M: Lysosomes and
autophagy in cell death control. Nat Rev Cancer. 5:886–897. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Baehrecke EH: Autophagy: dual roles in
life and death? Nat Rev Mol Cell Biol. 6:505–510. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He WS, Dai XF, Jin M, Liu CW and Rent JH:
Hypoxia-induced autophagy confers resistance of breast cancer cells
to ionizing radiation. Oncol Res. 20:251–258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Apel A, Herr I, Schwarz H, Rodemann HP and
Mayer A: Blocked autophagy sensitizes resistant carcinoma cells to
radiation therapy. Cancer Res. 68:1485–1494. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sannigrahi MK, Singh V, Sharma R, Panda NK
and Khullar M: Role of autophagy in head and neck cancer and
therapeutic resistance. Oral Dis. 21:283–291. 2015. View Article : Google Scholar
|
|
32
|
Rouschop KM, Ramaekers CH, Schaaf MB,
Keulers TG, Savelkouls KG, Lambin P, Koritzinsky M and Wouters BG:
Autophagy is required during cycling hypoxia to lower production of
reactive oxygen species. Radiother Oncol. 92:411–416. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zeng M, Wei X, Wu Z, Li W, Li B, Zhen Y,
Chen J, Wang P and Fei Y: NF-κB-mediated induction of autophagy in
cardiac ischemia/reperfusion injury. Biochem Biophys Res Commun.
436:180–185. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Y, Wu Y, Wu D, Tashiro S, Onodera S
and Ikejima T: NF-kappab facilitates oridonin-induced apoptosis and
autophagy in HT1080 cells through a p53-mediated pathway. Arch
Biochem Biophys. 489:25–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bubici C, Papa S, Dean K and Franzoso G:
Mutual cross-talk between reactive oxygen species and nuclear
factor-kappa B: molecular basis and biological significance.
Oncogene. 25:6731–6748. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sun Z, Yin Z, Liu C and Tian J: The
changes in the expression of NF-KB in a degenerative human
intervertebral disc model. Cell Biochem Biophys. 72:115–122. 2015.
View Article : Google Scholar
|