Introduction
Natural pigments are known for possessing a wide
range of pharmacological and health-promoting properties, including
anti-bacterial, anti-viral, anticancer and antioxidant activities
(1,2). Increasing advances in medicinal
chemistry have played key roles in transforming a class of dietary
naturally produced pigments into potential medical therapeutics
(3). Biosynthetic pigments are
important natural pigments, not restricted by seasons, materials
and other conditions. Microorganisms can undergo continuous
fermentation to produce pigments to meet the increasing needs of
the community, and the majority of microbial pigments have
biological activities (4,5).
Although microbial pigments have been extensively
studied, fungal pigments remain less reported. Fungi are a common
and important species, and are widely distributed, easy to be
cultured, not requiring a rich amount of nutrients and having
potent antiviral properties. In the past, much attention was paid
to the hazards of Fusarium and to the toxins produced, but
not on its usefulness (1,2). We previously identified a new strain
Fusarium (Fusarium sp. JN158) in our laboratory. This
strain is capable of producing pigments. Following fermentation,
the colors of the crude pigments varied due to the different pH
levels. Under acidic conditions, the pigments were red in color,
while under alkaline conditions, the pigments were purple in color
and precipitated; they exhibited an antioxidant function and
exerted inhibitory effects on the proliferation of cancer cells
(6). The products were separated
by high-performance liquid chromatography with a diode-array
detector (HPLC-DAD) and exhibited 6 peaks (representing 6
compounds). The 6th peak representing the 6th compound was the
highest one and the distance between this peak with the surrounding
peaks was the greatest. This component was separated and purified,
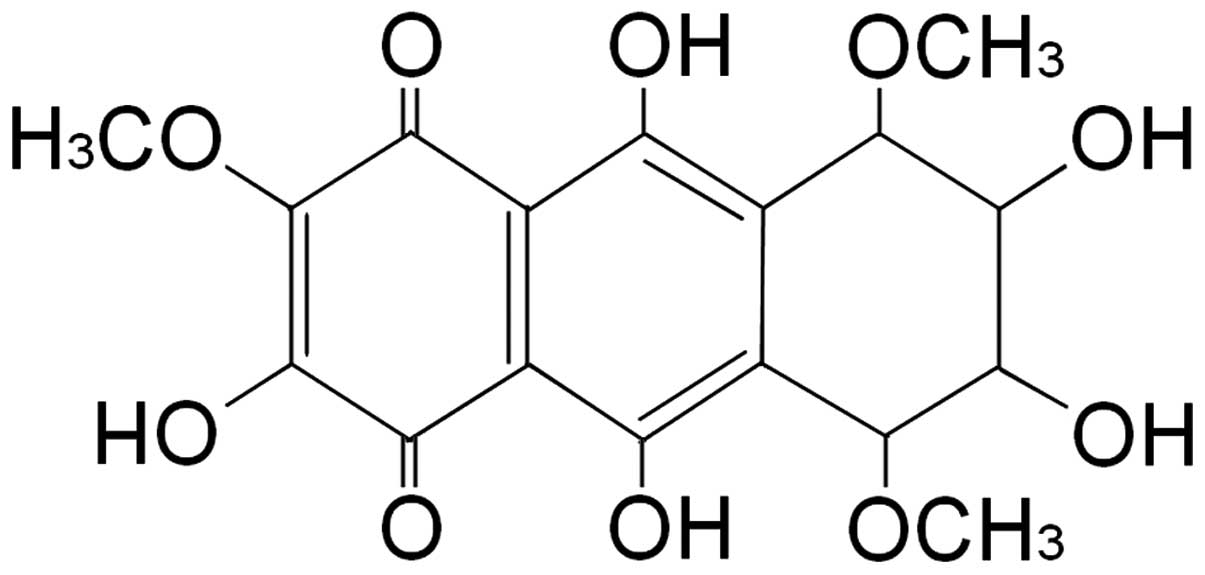
the purity of which was 98%. Based on the results of 1H
NMR and 13C NMR spectrum analysis, the compound was
determined to be a benzoquinone compound (one of the quinone
compounds) (7,8) (Fig.
1).
Pre-screening of fungal pigment at various
concentration using growth inhibition assays indicated that the
pigments from fungus exhibited anti-cancer effects, inhibiting
cancer cell proliferation (9).
Thus, led us to select compound VI for further investigations using
MCF-7 breast cancer cells. In the present study, we aimed to
examine the effects of compound VI on the proliferation of MCF-7
estrogen receptor (ER)+ cells and on MDA-MB-231
ER− cells, and to further elucidate the underlying
mechanisms. We demonstrated that ER is an important target for
therapeutic strategies aimed at controlling the proliferation of
hormone-dependent breast cancer cells. Of note, the activation of
nuclear factor-κB (NF-κB) may play a key role in ER+
tumors. The constitutive activation of NF-κB in breast tumors is
associated with more aggressive ER+ tumors. Recent data
have indicated that the activity of NF-κB is associated with ER
signaling in breast cancer cells (10–12).
The purpose of this study was to evaluate the
anti-proliferative activity in both MCF-7 and MDA-MB-231 human
breast cancer cell lines and the molecular mechanisms through which
compound VI inhibits ER signaling in androgen-dependent MCF-7
cells.
Materials and methods
Reagents
Monoclonal antibodies against vascular endothelial
growth factor (VEGF; sc-7269), caspase-3 (sc-7148), ERα (sc-73562),
progesterone receptor (PR; sc-130071), Bax (sc-20067), Bcl-2
(sc-130308), cyclin D1 (sc-70899) and NF-κB p65 (sc-372) were
provided by Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
RPMI-1640 medium (Gibco-Invitrogen, Grand Island, NY, USA) was
purchased from Shanghai Chemical Reagent Co. Ltd. (Shanghai,
China). The tetrazolium salt
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
was purchased from Sigma (Grand Island, NY, USA).
Cell lines
Pulmonary adenocarcinoma cells (A539), human gastric
carcinoma cells (MKN-45), hepatocellular carcinoma cells (HepG2),
human colon cancer cells (SW620), human umbilical vein endothelial
cells (HUVECs) and human breast cancer cells [MCF-7
(ER+), MDA-MB-231 (ER−)] were purchased from
Shanghai Institutes for Biological Sciences, Chinese Academy of
Sciences, Shanghai, China.
Cell viability and cell proliferation
assay
The cells were cultured in complete Dulbecco's
modified Eagle's medium (DMEM) medium containing 10% fetal bovine
serum (FBS). The cells were cultured in 24-well plates at the
concentration of 40,000 cells/ml. The pigment compound VI
(benzoquinone) was diluted in acid-ethanol (the control groups were
treated with an equal volume of acid-ethanol solution). The cells
were then incubated at 37°C in a humidified atmosphere of 5%
CO2 for 24 h. The cells were treated with 17β-estradiol
(E2, 0.1 µM; Sigma-Aldrich, Castle Hill, NSW,
Australia), fulvestrant (Fu, 0.13 µM; Sigma, Auckland, New
Zealand), compound VI (5, 10 and 15 µM), or the
corresponding control solutions. The cells were incubated at 37°C,
5% CO2 in a humidified incubator for 24 h. Assays were
initiated by the addition of 100 µl MTT solution (2 mg/ml)
to each well and incubating the cells for an additional 4 h at
37°C. Subsequently, the medium was removed and 1 ml
dimethylsulphoxide (DMSO) was added to each well. Finally, the
supernatants were transferred to 96-well plates in triplicate,
which were read at a wavelength of 490 nm with a Thermo Scientific
Multiskan® Spectrum spectrophotometer (Thermo Fisher
Scientific, Inc., Waltham, MA, USA).
The pigment inhibitory concentration
(IC50, defined as the drug concentration at which cell
growth was inhibited by 50%) was assessed from the dose-response
curves. The percentage viability of the pigment extract was
calculated using the formula: cell viability (%) = 100 −
[(ODpigment control) −
(ODpigment)/(ODpigment)] ×100.
Monitoring cell behavior in
real-time
Cell behavior was monitored in real-time using the
xCELLigence system E-Plate (ACEA Biosciences Inc., San Diego, CA,
USA). A total of 20,000 MCF-7 cells was seeded in each well and
fresh DMEM was added for a duration of 24 h. The DMEM was then
removed from these wells and medium containing compound IV at
various concentrations (VI IC25, compound VI
IC25; VI IC75, compound VI IC75)
was then added. The impedance value of each well was automatically
monitored by the xCELLigence system for a duration of 72 h and
expressed as a cell index (CI) value. The CI value was used to
represent the cell status based on the measured frequency-dependent
electrical impedance. The CI value was used as a global guide to
cellular behaviour including attachment, proliferation and
spreading.
Immunohistochemistry assay
The cells were seeded onto glass coverslips placed
in a 24-well plate (5×105 cells/well) in complete DMEM
medium containing 10% FBS and incubated overnight. The cells were
treated with VI IC50, Fu or the respective controls or
the vehicle. The VI IC50 Con used was an equal volume of
acid-ethanol solution. The fulvestrant control (Fu Con) used was an
equal volume of acetate solution. Complete medium only was used as
the vehicle. The cells were fixed using 4% paraformaldehyde, 0.15%
picric acid in 1X phosphate-buffered saline (PBS) for 20 min at
room temperature. The cells on glass coverslips were incubated with
the indicated primary antibodies at room temperature for 1 h. After
washing 3 times with PBS for 5 min, the slides were incubated for
15 min with the corresponding biotinylated secondary antibodies
(Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd., Beijing,
China), followed by 30 min of incubation with
streptavidin-horseradish peroxidase (DakoCytomation A/S, Glostrup,
Denmark). Color was developed with 3′-diaminobenzidine (DAB) for
3–7 min. Counterstaining was performed with hematoxylin for 5 min,
and the slides were coverslipped. To evaluate ERα and PR
expression, a semi-quantitative method was used to determine the
reaction intensity and percentage of positive cells (deep blue)
from 10 representative fields of the specimen at ×100 magnification
(Olympus DSX500, Olympus Corp., Tokyo, Japan).
Immunofluorescence staining and confocal
microscopy
This method was used to determine the expression of
NF-κB p65, caspase-3 in MCF-7 cells. The cells were seeded onto
glass cover-slips placed in a 24-well plate (5×105
cells/well). The groups and treatments were as described above. The
cells were treated with various concentrations of the pigment
compound VI. Following treatment, the cells were fixed with 4%
paraformaldehyde for 15 min at room temperature. The cells were
blocked with 10% normal goat serum (Gibco) prior to incubation with
rabbit anti-NF-κB p65, caspase-3 (Santa Cruz Biotechnology, Inc.)
or negative control rabbit IgG (Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd.) overnight at 4°C. After rinsing with PBS,
the cells were incubated with goat anti-rabbit Cy5 (Abcam,
Cambridge, MA, USA) and 1 µg/ml of Hoechst 33342 (Sigma) for
1 h at room temperature. Immunolabelling was visualized and imaged
using an Olympus DP71 fluorescence microscope (Olympus, Center
Valley, PA, USA).
Western blot analysis
The cells were washed twice with cold PBS and lysed
in ice-cold RIPA buffer (Sigma) supplemented with complete
mini-protease inhibitor cocktail tablet (Roche Diagnostics,
Mannheim, Germany). The cell lysates were centrifugated at 4°C,
12,000 rmp, for 30 min. We collected the supernatant and measured
the protein concentration using a Bradford kit (Thermo Fisher
Scientific, Inc.). Sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) electrophoresis was carried out using
standard protocols with 15 µg of total protein/well. The
primary antibodies used were as follows: mouse anti-human cyclin
D1, VEGF, NF-κB p65, Bax and Bcl-2 monoclonal antibodies (1:200).
Following incubation with the primary antibodies at 4°C overnight,
the membranes were washed with PBST 3 times, and were then
incubated with HRP-labeled secondary antibody (1:2,000; Beijing
Zhongshan Golden Bridge Biotechnology Co., Ltd.) for 2 h at room
temperature. The blots were visualized using a chemiluminescent
detection kit (Thermo Fisher Scientific, Inc.). The gray ratio of
each band was measured using ImageJ software. Relative protein
expression was calculated using the following gray ratio: gray
ratio = gray of target band/β-actin gray.
Statistical analysis
SPSS 11.5 statistical software was used for data
processing, and data are the means and standard deviation (means ±
SD). Comparisons between 2 groups were made using the LSD test and
Hochberg statistical methods for statistical processing. The value
of α=0.05 was used as a significant test standard, with a value of
P<0.05 used to indicate a statistically significant
difference.
Results
Compound VI inhibits the proliferation of
MCF-7 cells
MTT assay was used to screen the isolated compound
VI against a panel of human cancer cell lines, including A549,
MCF-7, MKN-45, HepG2 and SW620 cells. As shown in Fig. 2, the isolated compound VI
displayed broad spectrum anti-proliferative activities in a
dose-dependent manner (Fig. 2A).
Compared with the corresponding controls, compound VI inhibited the
proliferation of different tumor cells in a dose-dependent manner
(Fig. 2A–C), while it did not
have any apparent effect on normal cells, such as HUVECs (Fig. 2D). We also found that compound VI
exerted a more potent inhibitory and significant effect on MCF-7
cell proliferation (Fig.
2A–C).
 | Figure 2Effects of compound VI on the
proliferation of A549, MCF-7, MKN-45, HepG2, SW620 cells or the
viability of MCF-7 and human umbilical vein endothelial cells
(HUVECs). (A–C) Cell proliferation was examined following treatment
with compound VI at 5, 10 and 15 µM for 24 h, respectively.
(D) Cell viability was examined following treatment with compound
VI at 5, 6, 7, 8, 9, 10, 11, 13 and 15 µM. Viability was
determined by MTT assay. OD, optical density; n=20 wells.
*p<0.05 vs. VI Con; **p<0.01 vs. VI
Con; #p<0.05 vs. fulvestrant (Fu). |
To further explore the correlation between the
concentration of compound VI and its inhibitory effect on MCF-7
cells, we employed 9 gradients of compound VI. The results revealed
that 7 µM of compound VI exerted a 25% inhibition, while 11
µM of compound VI exerted a 50% inhibition (Fig. 2D). Our findings further validated
that compound VI exerts an inhibitory effect on MCF-7 cell
proliferation in a concentration-dependent manner.
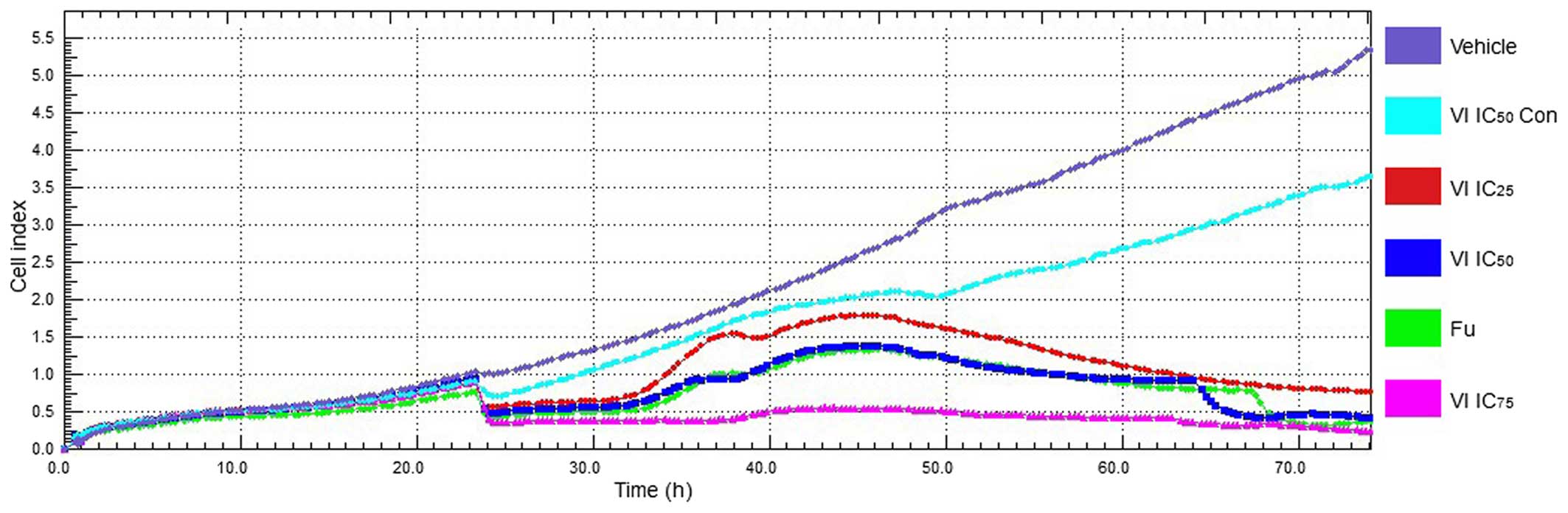
Determination of the behavior of MCF-7
cells treated with compound VI by real-time cell analysis
The xCELLigence system is a label-free cell-based
assay system, which integrates microelectronics and cell biology,
and is suitable for the uninterrupted monitoring of the biological
processes of living cells. The impedance measurements, which are
displayed as CI values, provide quantitative information about the
biological status of the cells including cell number, viability and
morphology (13). In this study,
we monitored the conditions of MCF-7 cells treated with compound VI
at various concentrations using the real-time cell-based assay
(RTCA) platform. The distinct patterns can be seen on the graph,
which can be attributed to cell adhesion and cell proliferation
(0–24 h) (Fig. 3).
After having determined the optimal conditions to
study the behavior of the MCF-7 cells (data not shown), we then
determined whether compound VI affects the behavior of MCF-7 cells.
In order to accomplish this, the MCF-7 cells were seeded in DMEM
containing various concentrations of compound VI. Cell behavior was
monitored using RTCA over a period of 72 h. The results revealed
that the CI value of MCF-7 cells was robust and did not exhibit any
detectable difference within DMEM medium or without compound VI
treatment from 0 to 24 h (Fig.
3). However, cell proliferation was significantly compromised
with compound VI treatment from 24 to 72 h (Fig. 3). As shown in Fig. 3, the cells in the vehicle-treated
group continuously grew more rapidly (dark purple line); however,
cells in the treated group grew at a slower rate during the period
of 24–48 h. The CI value of the treatment groups was the hightest
at 46 h. After this time point, the CI values of the treated cells
gradually decreased [VI IC25 (red line), VI
IC50 (dark blue line), Fu (green line) and VI
IC75 (light purple line)], while the cells in the VI
IC50 control group grew more rapidly [Fig. 3, cyan (light blue) line]. Taken
together, this observation suggests that compound VI exerts an
inhibitory effect on MCF-7 cell proliferation, and these inhibitory
effects are evident when cells are treated for a period >24
h.
Compound VI treatment decreases the
expression of ERα and PR in MCF-7 cells
ER signaling is the most attractive target for the
clinical therapy of ER+ breast cancer. Within the
anti-proliferative environment, there is compelling evidence to
indicate that, in estrogen-sensitive human breast cancer cell
lines, such as MCF-7, treatment with certain agents disrupts
estrogen-responsive gene expression and inhibits estrogen-dependent
proliferation. PR expression is traditionally used as a clinical
indicator of ER function (that is, PR is an ER target gene)
(14).
In this study, to determine whether the compound
affects the expression of ERα and PR in MCF-7 cells, we employed
immunohistochemisty staining assay. Compound VI was added at the
IC50 concentration to the MCF-7 cells for 24 h. The
number of MCF-7 cells whose cytoplasm was stained deep blue
represented the cells expressing ERα and PR. Immunohistochemical
analysis demonstrated that the expression of ERα and PR was
markedly inhibited in the cells treated with compound VI at the
IC50 concentration (compare Fig. 4A or B, VI IC50 to
Fig. 4, VI IC50
Con).
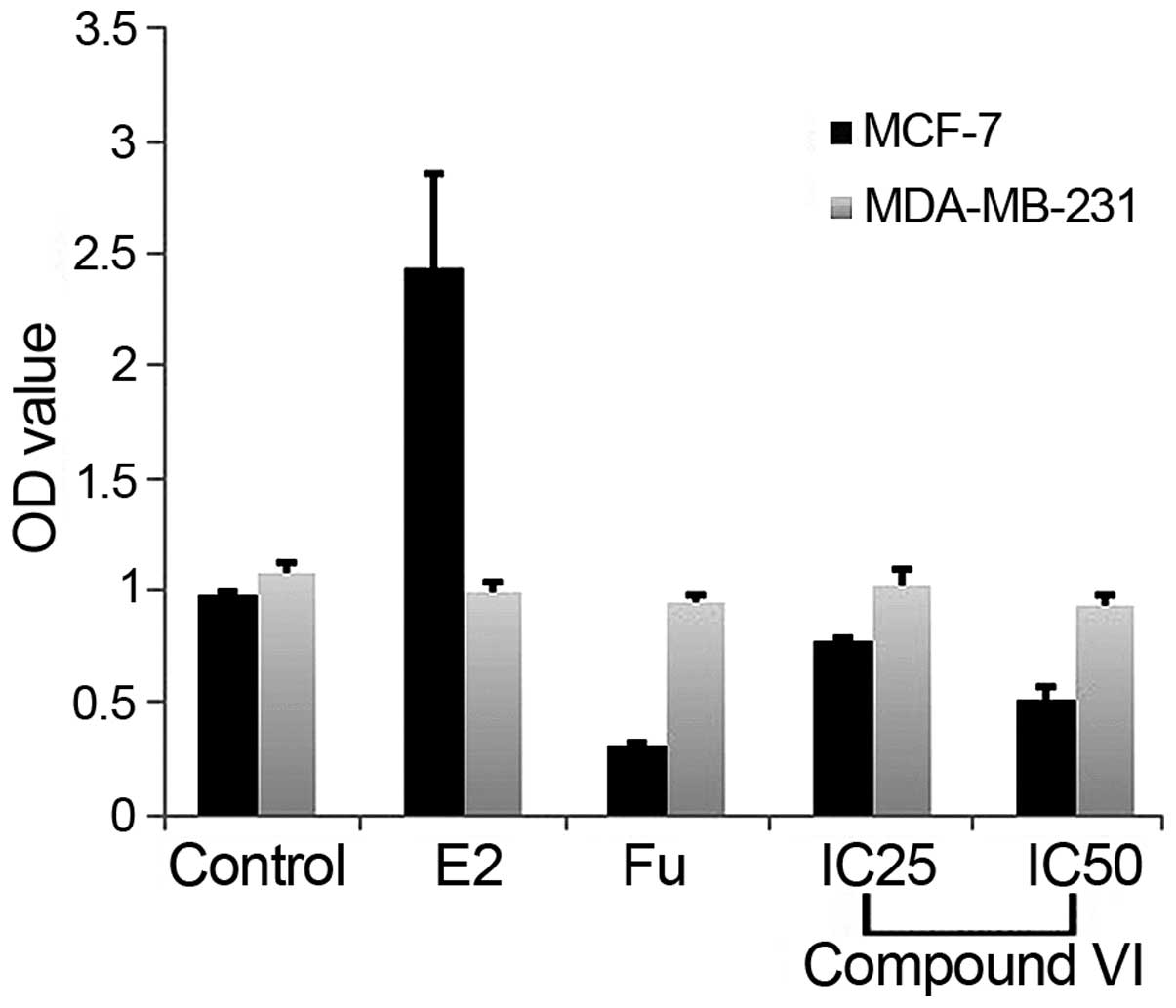
Compound VI differentially regulates the
viability of ER+ and ER− breast cancer
cells
The effects of the benzoquinone compound on cell
viability were evaluated by MTT assay in ER+ MCF-7 and
ER− MDA-MB-231 cells (Fig.
1A). Compared to the controls, compound VI exerted an overall
decrease in the OD value in a dose-dependent manner. The compound,
however, had no effect on MDA-MB-231 cells, suggesting that the
anti-proliferative effects of compound VI are dependent on the
presence of ERα (Fig. 5).
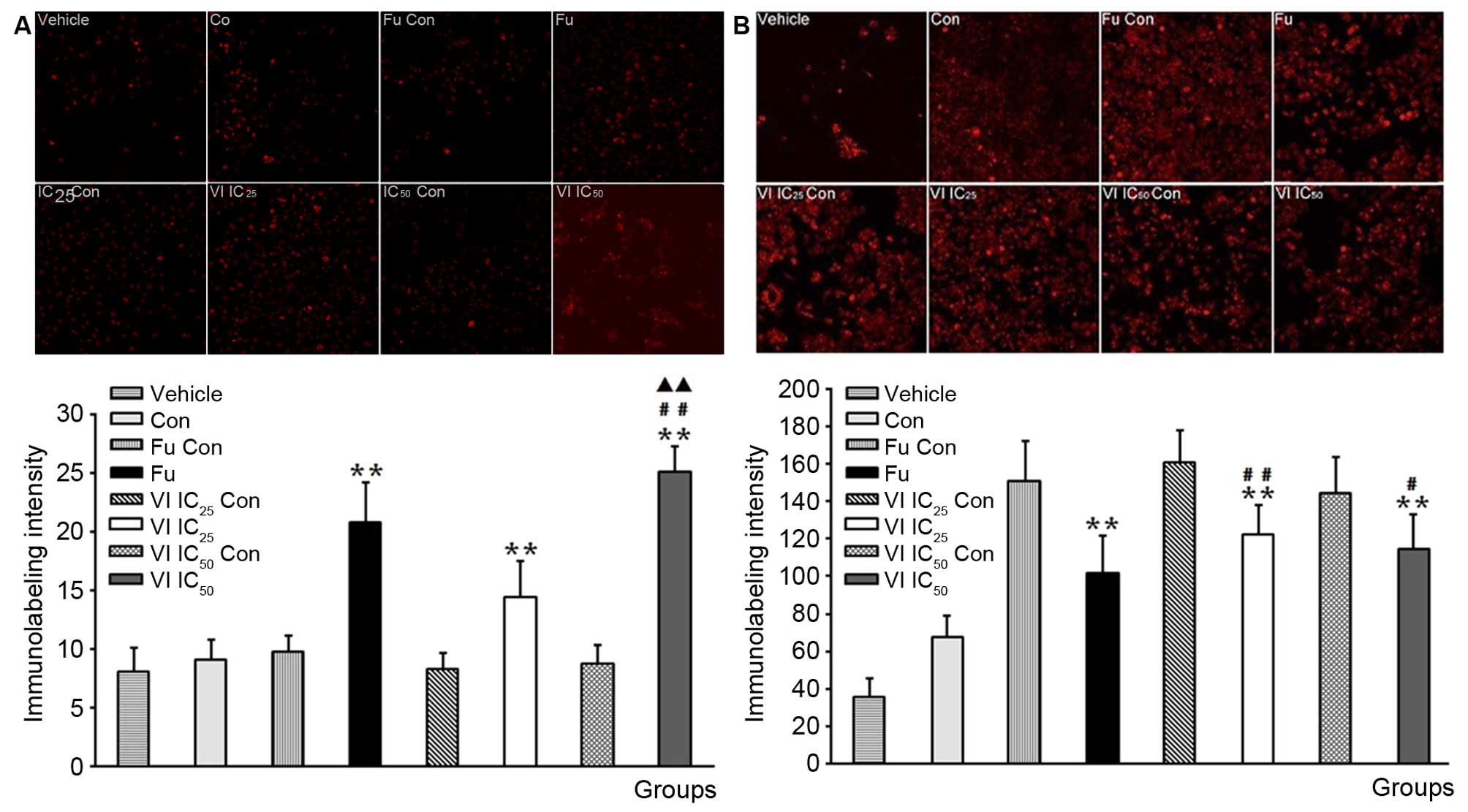
Caspase-3 and NF-κB immunofluorescence
staining
Of note, the activation of the pro-inflammatory
transcription factor, NF-κB, may play a key role in ER+
tumors, while caspases-3 plays an essential role in cell apoptosis
and has been termed an 'executioner' protein for its role in cell
apoptosis and may be used as a biomaker to predict tumor response
to treatment (15). Quinone has
been shown to induce the apoptosis of tumor cells by suppressing
NF-κB, activating the Akt pathway and suppressing tumor
angiogenesis (16). Thus, in this
study, we wished to determine whether compound VI affects the
expression of NF-κB and caspases-3. The results of
immunofluorescence staining revealed that caspase-3 was exclusively
localized in the cytoplasm of MCF-7 cells (Fig. 6A, vehicle) while NF-κB was
observed in the nuclei of MCF-7 cells (Fig. 6B, vehicle). We further explored
the changes in the expression of NF-κB and caspase-3 following
treatment of the cells with compound VI (VI IC25 and VI
IC50) for 24 h. We observed that the expression of NF-κB
was decreased (Fig. 6B), whereas
that of caspase-3 was increased in the MCF-7 cells following
treatment with compound VI (Fig.
6A). To further evaluate NF-κB expression and cleaved caspase-3
activation, their protein levels were examined in the cells by
western blot analysis (Fig. 7).
Treatment with compound VI inhibited NF-κB expression and increased
the cleavage of caspase-3.
 | Figure 7Relative expression levels of active
caspase-3 (cleaved caspase-3), nuclear factor-κB (NF-κB) p65, Bax,
Bcl-2, cyclin D1, vascular endothelial growth factor (VEGF) in
MCF-7 cells. The cells were treated with compound VI
(IC25 and IC50) for 24 h. (A) The western
blot bands of Bax, Bcl-2, cleaved caspase-3, cyclin D1, NF-κB p65,
VEGF. (B) Densitometric analysis was performed to quantify the
expression levels of the detected proteins. Data represent the
means ± SD of 7 independent experiments. **p<0.01 vs.
VI IC25 Con or IC50 Con;
#p<0.05 or ##p<0.01 vs. fulvestrant
(Fu); ▲p<0.05 or ▲p<0.01 vs. VI
IC25. |
Effect of compound VI on Bax, Bcl-2,
NF-κB p65, caspase-3, VEGF and cyclin D1 expression
To further explore the molecular mechanisms
responsible for the inhibitory effects of compound VI on MCF-7 cell
proliferation, we examined the protein expression of Bax, Bcl-2,
NF-κB p65, caspase-3, VEGF and cyclin D1 by western blot analysis.
The results revealed that following treatment with compound VI at
IC25 and IC50 for 24 h, the levels of Bax and
cleaved caspase-3 markedly increased compared to those of control
group (p<0.01; Fig. 7). By
contrast, the levels of Bcl-2, NF-κB, cyclin D1 and VEGF in the
cells treated with compound VI at IC50 for 24 h were
significantly lower than those of the control group (p<0.01;
Fig. 7). However, the levels of
cleaved caspase-3 and Bax in the cells treated with compound VI at
IC50 were markedly higher than those of the control
group (p<0.01; Fig. 7B).
Discussion
Breast cancer is one of the most common malignancies
affecting women worldwide and the second leading cause of
cancer-related mortality in women (17). Surgical interventions and
chemotherapy are able to significantly reduce mortality; however,
both of these interventions have several limitations such as
potential toxicity, severe side-effects and resistance to
anti-estrogen compounds (18).
Thus, new approaches to combat this neoplasm are required, also
taking into account the new compounds that they may be used to
prevent cancer.
Many of the natural products/secondary metabolites
with interesting biological properties are considered as
potentially important sources of novel bioactive anticancer
compounds and are capable of producing chemically diverse compounds
without any side-effects or with less side-efffects for a variety
of clinical applications (19).
Among the natural substances, quinone pigments (found in bacteria,
fungi, animals and plants) have attracted attention in recent years
due to both their importance in vital biochemical processes and
their several known biological activities, such as antitumor,
anti-bacterial and anti-viral effects (20,21). In the present study, the new
pigments, produced by Fusarium sp. JN158, were shown to
exert anti-proliferative effects on cancer cells (Fig. 2); however, the biological
functions of the pigment compound are largely unknown.
Pigment compounds have been shown to have potent
anti-proliferative activity against human colon cancer Caco-2 cells
and ER+ human breast cancer cell lines (18). In this study, The pigment compound
VI inhibited the proliferation of different cancer cell lines in a
concentration-dependent manner, but the MCF-7 (ER+) cell
line was found to be more sensitive to compound VI. Compound VI did
not affect the proliferation of the triple-negative (MDA-MB-231)
human breast cancer cell line, following incubatoin for 24 h, as
shown by MTT assay. The reduction in the percentage of MCF-7 cell
viability revealed that the bioactive compound VI exerted
anti-proliferation effects at various concentrations. The results
were in accordance with those of the xCELLigence RTCA platform
assay; the CI value of the MCF-7 cells gradually decreased after 24
h of treatment with compound VI. Of note, compound VI significantly
suppressed ERα and PR expression in MCF-7 cells. ERα plays a key
role in the development of estrogen-dominant breast cancer. The
majority of ER+ primary tumors depend on estrogen
signaling for their growth and survival (22). ERα and ERβ, members of the nuclear
receptor superfamily, are structurally similar with slight
differences in their ligand binding domains. Thus, the receptors
can be modulated by ligands that are structurally similar to the
endogenous ligand 17β-estradiol (E2) (23). We hypothesized that treatment with
compound VI may inhibit estrogen signaling in MCF-7 cells to block
cell proliferation. Indeed, compound VI inhibited the
proliferation/or induced the apoptosis of MCF-7 cells by
suppressing ERα expression and estrogen signaling.
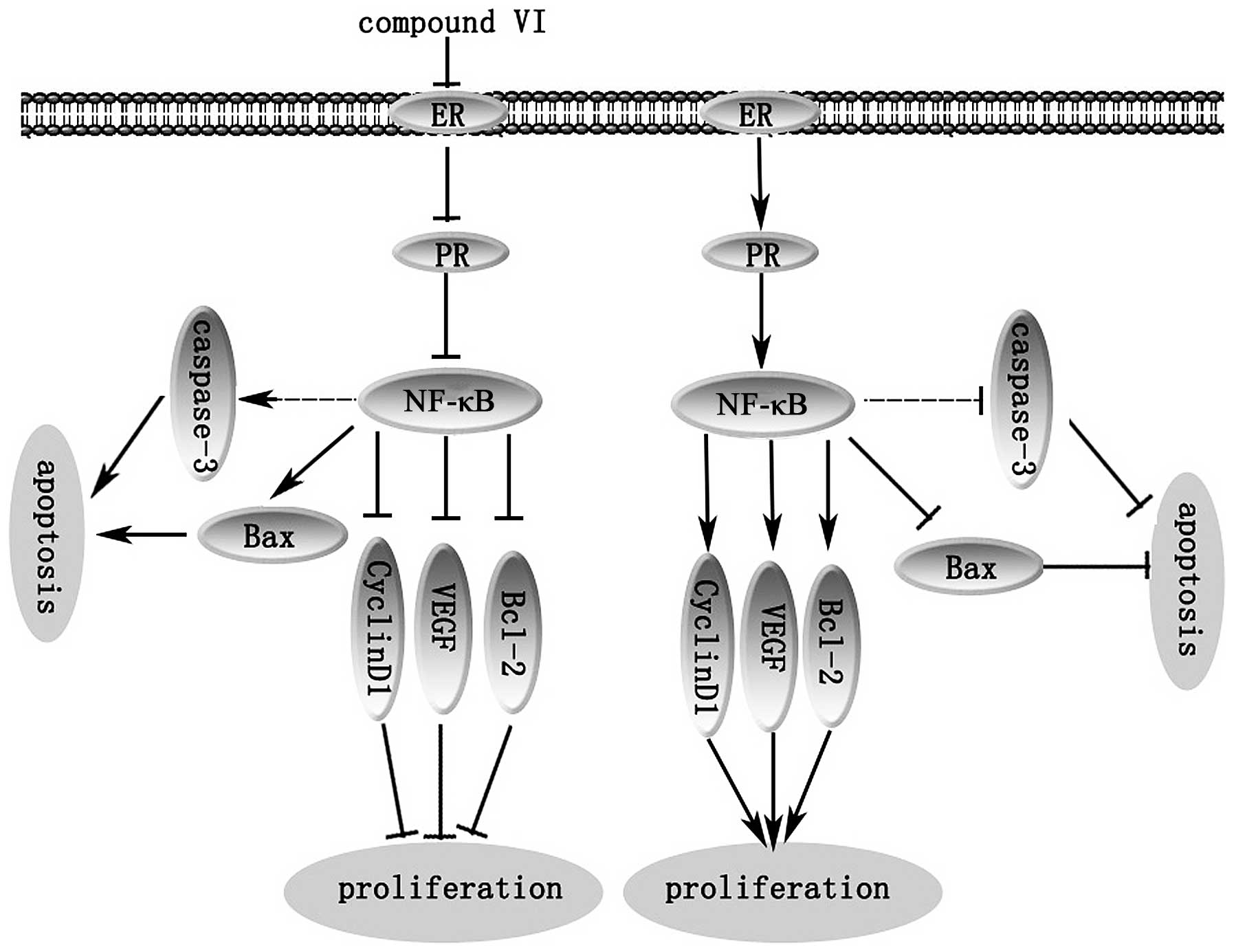
ER and PR are highly expressed in MCF-7 cells, and
significantly correlate with NF-κB expression in breast cancer. The
constitutive activation of the NF-κB pathway has been reported to
be involved in cellular proliferation, the inhibition of apoptosis
and the promotion of angiogenesis, and serves as an important
component for resistance to therapy in breast cancer (24–27). NF-κB can then upregulate the
transcription of its target genes. It is noteworthy to mention that
the reduced expression of NF-κB suppresses the expression of
downstream target genes involved in tumor cell proliferation.
Therefore, it was speculated that compound VI may block the
expression of NF-κB through the inactivation of NF-κB in breast
cancer cells, and may inhibit the expression of downstream target
genes, including cyclin D1, VEGF and Bcl-2 (24–27). It seemed that the inhibitory
effect on NF-κB accounted for the anti-proliferative effects of
compound VI. Furthermore, the perturbation of the protein
expression levels of Bax and Bcl-2 is an important factor to
determine the susceptibility of tumor cells to anticancer agents
(24–27). Therefore, the anticancer pigment
compound with the ability to suppress NF-κB translocation may be
effective in inducing the apoptosis of cancer cells. Thus, compound
VI suppresses the proliferation of MCF-7 cells by suppressing NF-κB
(Fig. 8).
In the present study, we identified the
effectiveness of pigment compound VI as an anti-proliferative agent
in MCF-7 cells, expanding its potential therapeutic utility for the
treatment of human cancers. This pigment compound may be
particularly advantageous when considering that severe side-effects
are often a limiting factor for the use of chemotherapeutic drugs
in the treatment of breast cancer.
Acknowledgments
This study was supported by the Chinese National
Natural Science Fundation (grant no. 81160531), the Jiangxi Natural
Science Fundation (grant no. 20161BAB205223), the Jiangxi
Department of Education (grant no. GJJ10528).
References
|
1
|
Mata-Gómez LC, Montañez JC, Méndez-Zavala
A and Aguilar CN: Biotechnological production of carotenoids by
yeasts: An overview. Microb Cell Fact. 13:122014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jackson H, Braun CL and Ernst H: The
chemistry of novel xanthophyll carotenoids. Am J Cardiol.
101:50D–57D. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kang B, Zhang X, Wu Z, Wang Z and Park S:
Production of citrinin-free Monascus pigments by submerged culture
at low pH. Enzyme Microb Technol. 55:50–57. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jomova K and Valko M: Health protective
effects of carotenoids and their interactions with other biological
antioxidants. Eur J Med Chem. 70:102–110. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hu W, Gao Q, Hamada MS, Dawood DH, Zheng
J, Chen Y and Ma Z: Potential of pseudomonas chlororaphis subsp.
aurantiaca strain Pcho10 as a biocontrol agent against Fusarium
graminearum. Phytopathology. 104:1289–1297. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zeng XL, Cai YJ, Liao XR, Zeng XK, Li ZH
and Zhang DB: Extraction and physico-chemical properties of
pigments from liquid-cultured mycelia of Fusarium sp. B158. Food
Science. 33:131–135. 2012.In Chinese.
|
|
7
|
Reigan P, David S, Guo WC and Ross D: A
mechanistic and structural analysis of the inhibition of the 90-kDa
heat shock protein by the benzoquinone and hydroquinone ansamycins.
Mol Pharmacol. 79:823–832. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pedrini N, Ortiz-Urquiza A, Huarte-Bonnet
C, Fan Y, Juárez MP and Keyhani NO: Tenebrionid secretions and a
fungal benzoquinone oxidoreductase form competing components of an
arms race between a host and pathogen. Proc Natl Acad Sci USA.
112:231–240. 2015. View Article : Google Scholar
|
|
9
|
Gaascht F, Dicato M and Diederich M: Venus
flytrap (Dionaea muscipula Solander ex Ellis) contains powerful
compounds that prevent and cure cancer. Front Oncol. 3:202–210.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Boopalan T, Arumugam A, Parada J,
Saltzstein E and Lakshmanaswamy R: Receptor activator for nuclear
factor-κB ligand signaling promotes progesterone-mediated
estrogen-induced mammary carcinogenesis. Cancer Sci. 106:25–33.
2015. View Article : Google Scholar :
|
|
11
|
King AE, Collins F, Klonisch T, Sallenave
JM, Critchley HO and Saunders PT: An additive interaction between
the NFkappaB and estrogen receptor signalling pathways in human
endometrial epithelial cells. Hum Reprod. 25:510–518. 2010.
View Article : Google Scholar
|
|
12
|
Ashcroft KJ, Syed F and Bayat A:
Site-specific keloid fibroblasts alter the behaviour of normal skin
and normal scar fibroblasts through paracrine signalling. PLoS One.
8:e756002013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim JH, Jekarl DW, Kim M, Oh EJ, Kim Y,
Park IY and Shin JC: Effects of ECM protein mimetics on adhesion
and proliferation of chorion derived mesenchymal stem cells. Int J
Med Sci. 11:298–308. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Marconett CN, Sundar SN, Poindexter KM,
Stueve TR, Bjeldanes LF and Firestone GL: Indole-3-Carbinol
triggers aryl hydrocarbon receptor-dependent estrogen receptor
(ER)α protein degradation in breast cancer cells disrupting an
ERα-GATA3 transcriptional cross-regulatory loop. Mol Biol Cell.
21:1166–1177. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mezzanotte L, An N, Mol IM, Löwik CW and
Kaijzel EL: A new multicolor bioluminescence imaging platform to
investigate NF-κB activity and apoptosis in human breast cancer
cells. PLoS One. 9:e855502014. View Article : Google Scholar
|
|
16
|
Tozawa K, Sagawa M and Kizaki M: Quinone
methide tripterine, celastrol, induces apoptosis in human myeloma
cells via NF-κB pathway. Int J Oncol. 39:1117–1122. 2011.PubMed/NCBI
|
|
17
|
Kruk J: Overweight, obesity, oxidative
stress and the risk of breast cancer. Asian Pac J Cancer Prev.
15:9579–9586. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schiavano GF, De Santi M, Brandi G,
Fanelli M, Bucchini A, Giamperi L and Giomaro G: Inhibition of
breast cancer cell proliferation and in vitro tumorigenesis by a
new red apple cultivar. PLoS One. 10:e01358402015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ahmadian M and Samah AA: Application of
health behavior theories to breast cancer screening among Asian
women. Asian Pac J Cancer Prev. 14:4005–4013. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gordaliza M: Synthetic strategies to
terpene quinones/hydroquinones. Mar Drugs. 10:358–402. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bertanha CS, Januário AH, Alvarenga TA,
Pimenta LP, Silva ML, Cunha WR and Pauletti PM: Quinone and
hydroquinone metabolites from the ascidians of the genus Aplidium.
Mar Drugs. 12:3608–3633. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Magnani L, Ballantyne EB, Zhang X and
Lupien M: PBX1 genomic pioneer function drives ERα signaling
underlying progression in breast cancer. PLoS Genet.
7:e10023682011. View Article : Google Scholar
|
|
23
|
Shanle EK and Xu W: Endocrine disrupting
chemicals targeting estrogen receptor signaling: Identification and
mechanisms of action. Chem Res Toxicol. 24:6–19. 2011. View Article : Google Scholar :
|
|
24
|
Condon JC, Hardy DB, Kovaric K and
Mendelson CR: Up-regulation of the progesterone receptor (PR)-C
isoform in laboring myometrium by activation of nuclear
factor-kappaB may contribute to the onset of labor through
inhibition of PR function. Mol Endocrinol. 20:764–775. 2006.
View Article : Google Scholar
|
|
25
|
Dai XL, Zhou SL, Qiu J, Liu YF and Hua H:
Correlated expression of Fas, NF-kappaB, and VEGF-C in infiltrating
ductal carcinoma of the breast. Eur J Gynaecol Oncol. 33:633–639.
2012.
|
|
26
|
Reuter S, Prasad S, Phromnoi K, Ravindran
J, Sung B, Yadav VR, Kannappan R, Chaturvedi MM and Aggarwal BB:
Thiocolchicoside exhibits anticancer effects through
down-regulation of NF-κB pathway and its regulated gene products
linked to inflammation and cancer. Cancer Prev Res (Phila).
3:1462–1472. 2010. View Article : Google Scholar
|
|
27
|
Miller Susanne C, Huang RL and Sakamuru S:
Identification of known drugs that act as inhibitors of NF-κB
signaling and their mechanism of action. Biochem Pharmacol.
9:1272–1280. 2010. View Article : Google Scholar
|