Introduction
Esophageal carcinoma is the 6th frequent cause of
cancer-related mortality worldwide (1), and esophageal squamous cell
carcinoma (ESCC) accounts for ESCC) accounts for 90% of all
esophageal carcinoma cases in Asian countries (2). Although surgical techniques and
peri-operative management have progressed, the prognosis for
patients with ESCC remains poor (3). Thus, the identification of molecular
therapeutic targets which may be used in the treatment of ESCC is
one of the most promising avenues of research that lead to an
improvement in the survival of patients with this type of
refractory cancer. Some of the genetic alterations associated with
the development or progression of ESCC have been described
(4). However, few of these genes
have been demonstrated to be associated with the biological or
pathological characteristics of ESCC. Therefore, the identification
of novel genes associated with the progression of ESCC is
imperative.
The mammalian target of rapamycin (mTOR/p70S6 kinase
(p70S6K) signaling pathway is an important intracellular signal
transduction pathway, which is closely associated with cell
survival and cell proliferation in many types of cancer. Studies
have demonstrated that cell proliferation and cell cycle-related
proteins (such as cyclin B1 and p27) (4,5),
cell apoptosis-related proteins [such as cellular inhibitor of
apoptosis protein 1 (cIAP1) and caspase-3] (6,7)
and the key proteins [p70S6K and eukaryotic translation initiation
factor 4E-binding protein 1 (4EBP1)] (8). Studies have shown that the mTOR
pathway activates protein translation and regulates protein
synthesis, which is the central regulator of cell growth. mTOR
plays an important role in determining cell, tissue and organ size.
The mTOR pathway is a key pathway that regulates cell growth and
proliferation, which integrates signals from nutrient molecules,
energy states and growth factors to regulate a large number of life
processes, including autophagy (9–10),
ribose body biosynthesis (11,12) and metabolism (13). p70S6K is the main downstream
target of mTOR which make an important role in the translation
initiation, protein synthesis, cell cycle, cell migration in the
mTOR/p70S6K pathway (19). The
dysregulation of the pathway is associated with a variety of human
diseases, including cancer (14,15), diabetes (16) and leukemia (17,18). 4EBP1 is a downstream effector
molecule of mTOR, through which (by phosphorylation) mTOR regulates
cancer growth, proliferation and survival. cIAP1 is an inhibitor of
apoptosis protein that inhibits apoptosis mainly by inhibiting the
activity of caspase-3/7 and the role of nuclear factor (NF)-κB
(20). Caspase-3 is the most
important terminal cleavage enzyme in the process of apoptosis. It
is the terminal executive molecule downstream of the caspase
cascade, which is the intersection of the death receptor pathway
and mitochondrial pathway. Thus, caspase-3 is also known as death
protease (21). Furthermore, the
mTOR/p70S6K pathway plays an important role in the growth,
proliferation and anti-apoptosis of ESCC (22).
The anticancer activity of cardiac glycosides (the
main active ingredients in toad venom) has been a subject of
investigation since 1967. Emerging evidence now suggests that
cardiac glycosides are able to inhibit cell proliferation and
induce the apoptosis of various tumor cell lines. Bufalin, a major
bufadeinolide-like cardiac glycoside isolated from the skin and
parotid venom glands of the toad (23), has been shown to induce cancer
cell death through apoptotic pathways and has been investigted
experimentally in a number of types of cancer.
The aim of this study was to evaluate the anti-tumor
effects of bufalin by examining its effects on the mTOR/p70S6K
signaling pathway and on cell apoptosis in a mouse model of ESCC
created by the orthotopic transplanation of tumor cells. We
examined the effects of bufalin on the expression of
apoptosis-related proteins in order to determine the anti-tumor
mechanisms of action of bufalin on a molecular level.
Materials and methods
Cells, antibodies and reagents
The ESCC cell line, ECA109, was provided by the
Scientific Research Center of the Fourth Hospital of Hebei Medical
University (Hebei, China). A total of 36 Nu/Nu nude mice (half male
and half female, aged 4–6 weeks, weighed 18–22 g) purchased from
Vital River Laboratory Animal Technology Co., Ltd. (Beijing,
China). Our study was approved by the Ethics Committee of the
Fourth Hospital of Hebei Medical University. Bufalin was obtained
from Shanghai Yuanye Biotechnology Co., Ltd. (Shanghai, China) and
rapamycin was purchased from the Beyotime Institute of
Biotechnology (Shanghai, China). mTOR, p70S6K, 4EBP1, cIAP1,
caspase-3, Bad, Bcl-2, β-actin upstream and downstream primers were
all from Shanghai Sangon Biological Engineer ing Co., Ltd.,
(Shanghai, China). Rabbit anti-mTOR (1612-1), p70S6K (T2921),
p-p70S6K (ab2571), 4EBP1 (1557-1), p-4EBP1 (2250-1), cIAP1
(3302-1), Bad (1541-1), Bcl-2 (1017-1) and β-actin monoclonal
antibodies, and rat anti-active caspase-3 monoclonal antibody
(1476-1) were obtained from Epitomics (Burlingame, CA, USA). The
Alpha Innotech UV gel imaging system was from Alpha Innotech Co.,
(San Leandro, CA, USA) and the Odyssey two-color infrared laser
imaging system was from LI-COR Biosciences (Lincoln, NE, USA) The
inverted phase contrast microscope was from Nikon (Tokyo,
Japan).
Cell culture and tumor model
The ECA109 cells were seeded in 10% fetal bovine
serum (FBS) of RPMI-1640 medium and incubated at 37°C in a 5%
CO2 incubator. The medium was changed once every 2 to 3
days. A total of 36 nude mice were inoculated with ECA109 cancer
cells (each with 0.2 ml injected subcutaneously into the right side
of the axillary region) on the first day. The mice were provided
with adequate food and water and kept in an environment with a
temperature of 21±2°C and relative humidity of 30–70%. No ECA109
cancer cells were injected during feeding. The mice were allowed to
feed for 15 days and until the size of the tumors reached
approximately 1 cm3. The mice were then randomly divided
into 6 groups (n=6 per group) as follows: the model group
(saline-treated group), the low-dose bufalin group (0.5 mg/kg, BL
group), the medium-dose bufalin group (1 mg/kg, BM group), the
high-dose bufalin group (1.5 mg/kg, BH group), the rapamycin group
(RAPA group) and the combination group (administered bufalin and
rapamycin; BR group). The nude mice were injected for 30 days
intraperitoneally according to the dose in each group every day
apart from the combination group (once every other day). The mice
in the model group were injected with 0.2 ml saline once a day, and
those in the rapamycin group were injected with 50 μg/kg
rapamycin. The nude mice were sacrificed by cervical dislocation at
the end of the treatment period. The tumors were then removed and
approximately 0.5 cm3 of the tumor was cut and placed in
a refrigerator at −80°C for use in RT-PCR and western blot
analysis. The remaining parts of the tumors were fixed in formalin
for H&E staining, TUNEL and immunohistochemistry. The tumor
inhibition rate was calculated as follows: tumor inhibition (%) =
(mean tumor volume of model group - mean tumor volume of
intervention group)/mean tumor volume of model group ×100%.
Morphological analysis
The graded ethanol-dehydrated, paraffin-embedded,
hematoxylin and eosin (H&E)-stained, xylene-cleared tissue
sections were sliced at 4 μm. The H&E-stained sections
were observed under a microscope (Axio Scope.A1; Carl Zeiss
(Shanghai) Management Limited, Shanghai, China), and the groups
were divided according to the necrosis range, that is, focal
necrosis, a large area of necrosis, or a larger area of
necrosis.
TUNEL assay
The tumor tissues were fixed in 10% neutral
formalin, dehydrated, paraffin-embedded and sectioned prior to
TUNEL staining. The sections were then digestived with 20
μg/ml proteinase K (Beyotime Institute of Biotechnology) for
15 min. The apoptotic index in the tumor tissue was detected using
the ApopTag Peroxidase In Situ Apoptosis Detection kit (EMD
Millipore; Bilerica, MA, USA). The results of TUNEL staining were
observed under a microscope (Axio Scope.A1; Carl Zeiss (Shanghai)
Management Limited). The positive staining was observed in DAB
coloring liquid. A total of 10 high power field (HPF) were selected
to calculate the proportion of positive cells randomly and the
apoptotic index (AI) was the number of positive cells in 100
cells.
RT-PCR
Total RNA was extracted using TRIzol reagent (Sangon
Biotech (Shanghai) Co., Ltd., Shanghai, China) and the integrity of
RNA was determined using the Alpha Innotech Gel imaging system.
Total RNA were reverse transcribed into cDNA. At the end of PCR
amplification, the PCR amplification products were confirmed and
analyzed. The primers for mTOR, p70S6K, 4EBP1, cAIP1, caspase-3
primers were verified by NCBI and β-actin was used as an internal
reference gene. The primer sequences were as follows: mTOR (318 bp)
forward, 5′-AGTGGACCAGTGGAAAC AGG-3′ and reverse,
5′-TTCAGCGATGTCTTGTGAGG-3′; p70S6K (188 bp) forward,
5′-TACTTCGGGTACTTGGTAA-3′ and reverse, 5′-GATGAAGGGATGCTTTACT-3′;
4EBP1 (156 bp) forward, 5′-ACCGGAAATTCCTGATGGAG-3′ and reverse,
5′-CCCGCTTATCTTCTGGGCTA-3′; cIAP (245 bp) forward,
5′-TCCCAGGTCCCTCGTATCAA-3′ and reverse, 5′-ATCCAGCATCAGGCCACAAC-3′;
caspase-3 (116 bp) forward, 5′-TCATTATTCAGGCCTGCCGTGGTA-3′ and
reverse, 5′-ATGCATACAAGAAGTCGGCCTCCA-3′. The reaction conditions
were as follows: for mTOR: 95°C for 5 min, 95°C for 30 sec, 60.1°C
for 30 sec, 72°C for 30 sec; 37 cycles, 72°C for 10 min; for
p70S6K: 95°C for 5 min, 95°C for 45 sec, 48°C for 45 sec, 72°C for
1 min, 37 cycles; 72°C for 5 min; for 4EBP1: 95°C for 5 min, 95°C
for 30 sec, 48°C for 30 sec, 72°C for 30 sec, 37 cycles; 72°C for
10 min; for cIAP1: 95°C for 5 min, 95°C for 30 sec, 60°C for 50
sec, 72°C for 30 min, 35 cycles; 72°C for 5 min; caspase-3 was 95°C
for 5 min, 95°C for 30 sec, 56°C for 45 sec, 72°C for 1 min, 35
cycles; 72°C for 10 min. The results were observed using the Alpha
Innotech UV gel imaging system and the grayscale value was then
calculated as follows: relative expression amount = grayscale value
of target gene/grayscale value of β-actin.
Western blot analysis
Approximately 30 mg fresh tumor tissue was ground
and place on ice. This was followed by the addition of 300
μl ice-cold lysis buffer and 2 μl PMSF. The samples
were then centrifuged at12,000 × g at 4°C for 15 min. The proteins
(50 μg) were denatured at 95°C in metal bath and then
subjected to SDS-PAGE and transferred onto PVDF membranes. The
membranes were then incubated with the primary antibodies, mTOR,
p70S6K, p-p70S6K, 4EBP1, p-4EBP1, cIAP1, active caspase-3, Bad and
Bcl-2 (dilution, 1:2,000), diluted in TBST overnight at 4°C in a
shaker. We then used an Odyssey two-color infrared laser imaging
system to capture and analyze the images after washing the
membranes 3 times with TTBS and the grayscale value was then
determined.
Immunohistochemistry
Immunohistochemistry was carried out using the
EnVision™ immunohistochemistry kit (Agilent Technologies, Santa
Clara, CA, USA) according to the manufacturer's instructions. This
was followed by the addition of antibodies (mTOR, p70S6K, p-p70S6K,
4EBP1, p-4EBP1, cIAP1, active caspase-3, Bcl-2 and Bad) to the
slices and incubation at room temperature for 150 min, paying
attention to ensure that the antibodies were in full contact with
the tissues. We determined the location of mTOR, p70S6K, p-p70S6K,
4EBP1, p-4EBP1, cIAP1, active caspase-3, Bcl-2 and Bad on the cell
membrane, nucleus or cytoplasm. The staining intensity was scored
as follows: no color corresponding to 0, canary yellow
corresponding to 1, pale brown corresponding to 2, tan
corresponding to 3; the percentage of positive cells was negative
corresponding to 0, positive cells <10% corresponding to 1,
positive cells between 11–50% corresponding to 2, positive cells
between 51–75% corresponding to 3, and positive cells >75%
corresponding to 4. The product of staining intensity and the
percentage of positive cells was negative (−) corresponding to 0,
weakly positive (+) corresponding to 1–4, moderate positive (++)
corresponding to 5–8 and strong positive (+++) corresponding to
9–12.
Statistical analysis
One-way analysis of variance (ANOVA) was used to
determine the differences between the 6 groups and multiple
comparisons between groups were made using the Student-Newman-Keuls
(SNK) test. The experimental data of the results of
immunohistochemistry were examined using the Kruskal-Wallis H test
and multiple comparisons between groups were made using the Nemenyi
test. The results were considered statistically significant at a
vaue of P<0.05.
Results
No obvious abnormalities were observed as regards
the dietary habits of the nude mice diet and the mental state of
the mice in all the groups (model, BL, BM, BH, RAPA and BR) during
the course of medication (P>0.05) (Table I). There were no significant
differences in the body weight of the mice in each group prior to
treatment (P>0.05), and even post-treatment (P>0.05)
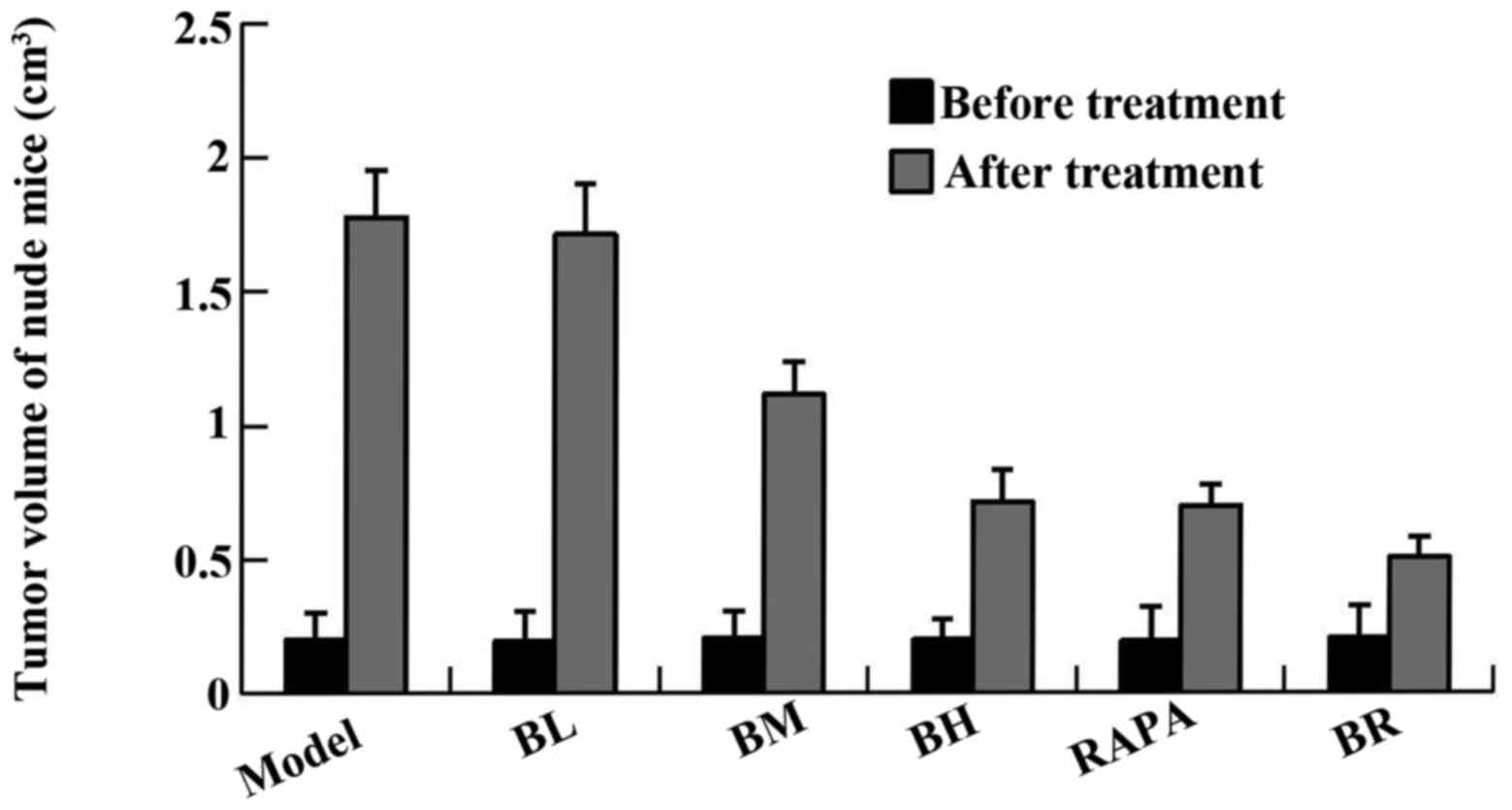
(Table II). The tumor size of
the mice in each group (model, BL, BM, BH, RAPA and BR) was
1.778±0.176, 1.714±0.188, 1.115±0.124, 0.713±0.122, 0.699±0.079 and
0.506±0.076 cm3, respectively and the tumor inhibition
rate was 4.1, 37.6, 60.1, 60.9 and 71.7%, respectively (Fig. 1, Table III). It proved that bufalin has
obvious antitumor effect.
 | Table IThe mental state and dietary habits
of the nude mice during the course of medication. |
Table I
The mental state and dietary habits
of the nude mice during the course of medication.
| Groups | Mental state | Dietary habits
|
|---|
| Fodder consumption
(g) | Post-treatment |
|---|
| Model | Good | 3.2±0.3 | 4.5±0.1 |
| Low-dose
bufalin | Good | 3.4±0.3 | 4.4±0.3 |
| Medium-dose
bufalin | Good | 3.5±0.2 | 4.6±0.3 |
| High-dose
bufalin | Good | 3.8±0.2 | 4.9±0.2 |
| Rapamycin | Good | 3.4±0.3 | 4.6±0.5 |
| Combination | Good | 3.6±0.2 | 4.6±0.1 |
| Table IIAverage weight of nude mice in the
transplanted model before and after treatment. |
Table II
Average weight of nude mice in the
transplanted model before and after treatment.
| Groups | Body weight (g)
|
|---|
| Pre-treatment | Post-treatment |
|---|
| Model | 20.9±2.4 | 27.9±3.0 |
| Low-dose
bufalin | 20.8±1.5 | 28.2±1.9 |
| Medium-dose
bufalin | 20.8±2.1 | 27.9±2.9 |
| High-dose
bufalin | 20.9±2.7 | 28.1±2.8 |
| Rapamycin | 20.9±1.9 | 27.9±2.0 |
| Combination | 20.8±2.1 | 27.8±2.6 |
| Table IIIAverage tumor volume and inhibiting
rate of the transplanted mode before and after treatment. |
Table III
Average tumor volume and inhibiting
rate of the transplanted mode before and after treatment.
| Groups | Tumor volume
(cm3)
| Tumor inhibition
(%) |
|---|
| Pre-treatment | Post-treatment |
|---|
| Model | 0.199±0.103 | 1.778±0.176 | – |
| Low-dose
bufalin | 0.194±0.111 | 1.714±0.188 | 3.6 |
| Medium-dose
bufalin | 0.206±0.099 | 1.115±0.124a | 37.3 |
| High-dose
bufalin | 0.197±0.079 | 0.713±0.122a | 59.9 |
| Rapamycin | 0.192±0.126 | 0.699±0.079a | 60.7 |
| Combination | 0.203±0.121 | 0.506±0.076a | 71.5 |
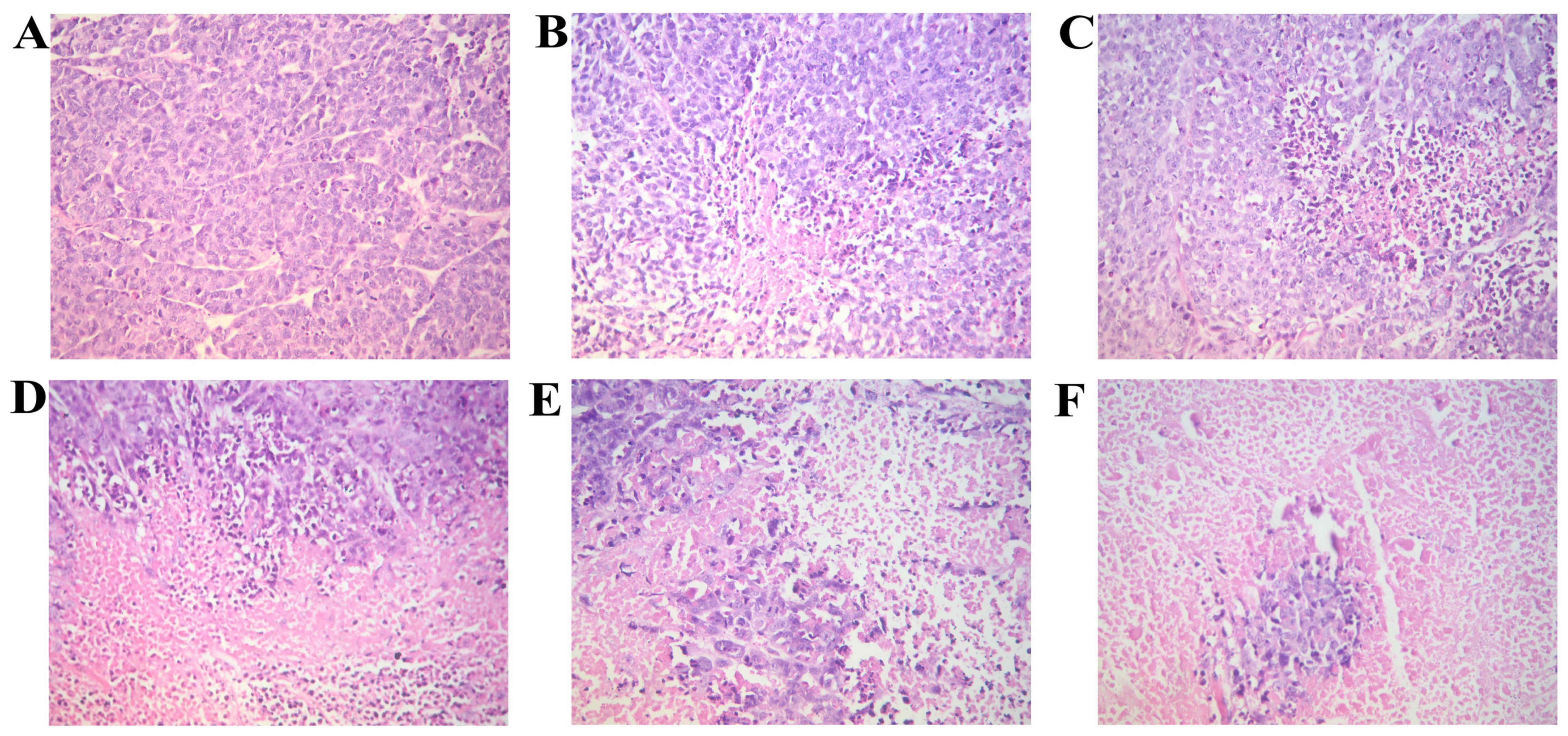
Morphology of cells in tumor tissue
We observed the morphological changes in the tumor
tissue isolated from the nude in each group mice with
orthotopically transplanted tumors. The hematoxylin and eosin
(H&E)-stained sections were observed under a microscope. Tumor
necrosis was evident in all the groups, apart from the model group
in which necrosis was hardly evident. The tissues from the BL group
exhibited the most extensive focal necrosis; the tisues from the BM
group exhibited a large area of necrosis; the tissues from the BH,
RAPA and BR groups exhibited large areas of necrosis and cancer
cell clumps were scattered in the necrotic tissues (Fig. 2).
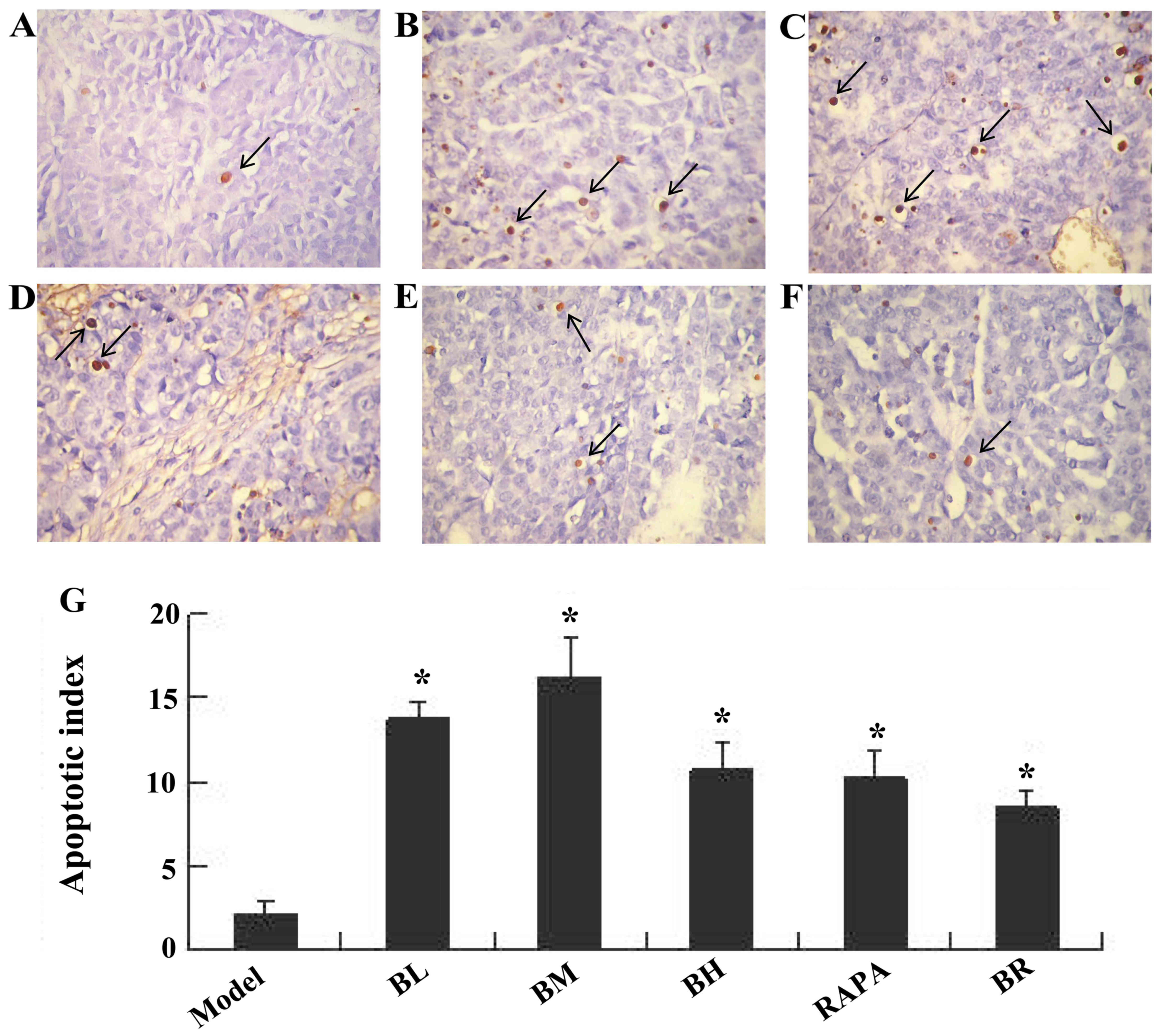
Bufalin promotes cell apoptosis in ESCC
tumor tissue, as shown by TUNEL assay
Apoptotic nuclei were stained brownish yellow. The
apoptotic indexes of each treatment groups (BL, BM, BH, RAPA and
BR) were significantly higher (13.67, 16.17, 10.83, 10.33 and 8.5%)
than those of the model group (2.17%); those of the BM group were
the highest (Fig. 3).
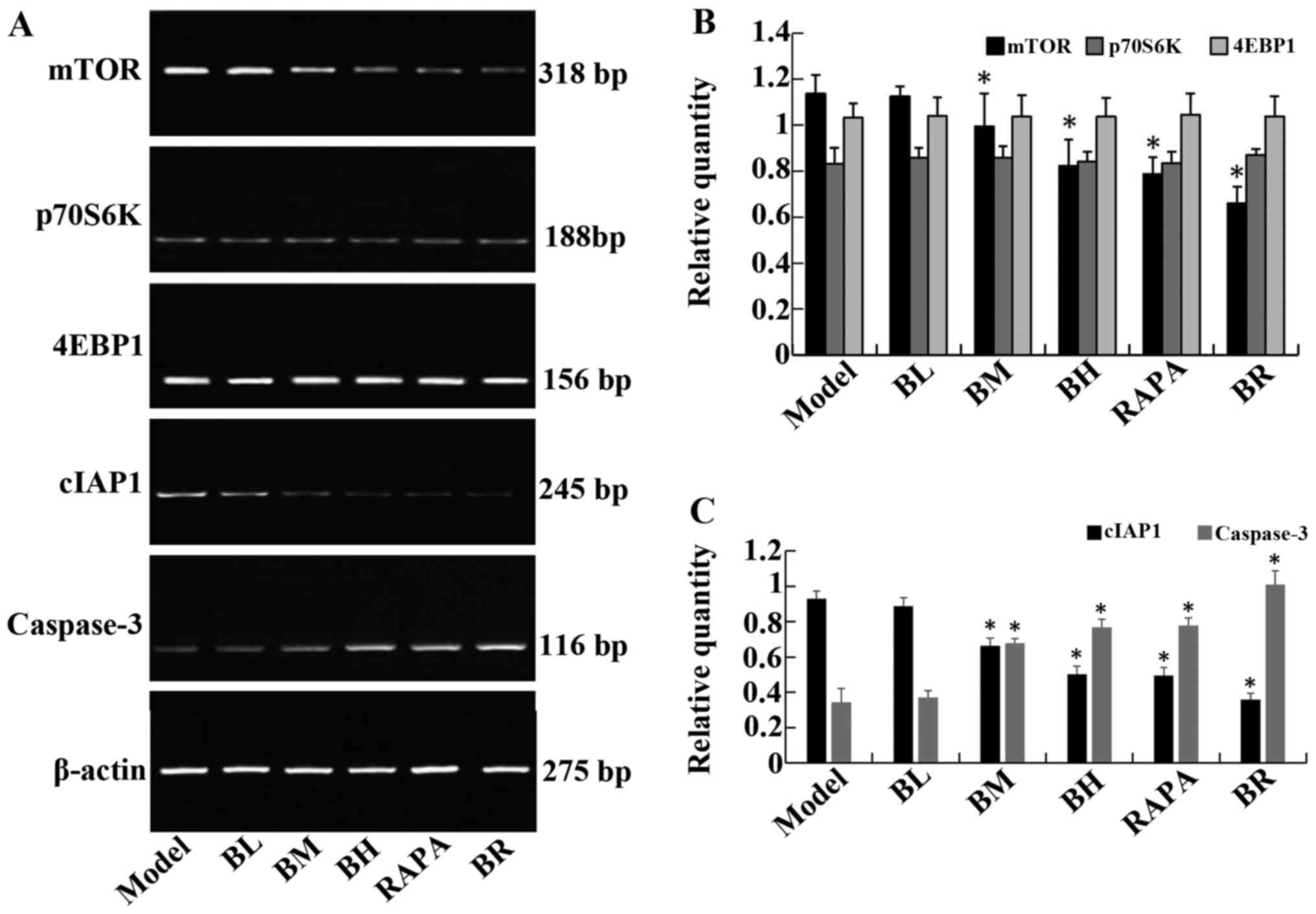
Bufalin affects the expression of
apoptosis-related genes
The mRNA expression levels of mTOR and cIAP1
gradually decreased in the treatment groups (BL, BM, BH, RAPA and
BR) compared to the model group, whereas the mRNA expression levels
of caspase-3 gradually increased; significant differences were
observed (P<0.05). It should be noted that no significant
differences were observed between the model and BL groups
(P>0.05), and between the BH and RAPA groups (P>0.05). The
mRNA expression levels of p70S6K and 4EBP1 did not differ
significantly between the groups (P>0.05; model, BL, BM, BH,
RAPA and BR) (Fig. 4).
Bufalin increases the expression of
apoptotic proteins, and decreases that of anti-apoptotic
proteins
The protein expression levels of mTOR, p-p70S6K,
p-4EBP1, cIAP1 and Bcl-2 gradually decreased in the treatment
groups (BL, BM, BH, RAPA and BR) compared to the model group.
However, the protein levels of caspase-3 and Bad gradually
increased, with significant differences between the each groups
(P<0.05). Again, it should be noted that no significant
differences were observed between the model and BL groups
(P>0.05), and between the BH and RAPA groups (P>0.05). No
statistically significant differences were observed in the protein
expression levels of p70S6K and 4EBP1 (P>0.05) between the
(model, BL, BM, BH, RAPA and BR) (Fig. 5).
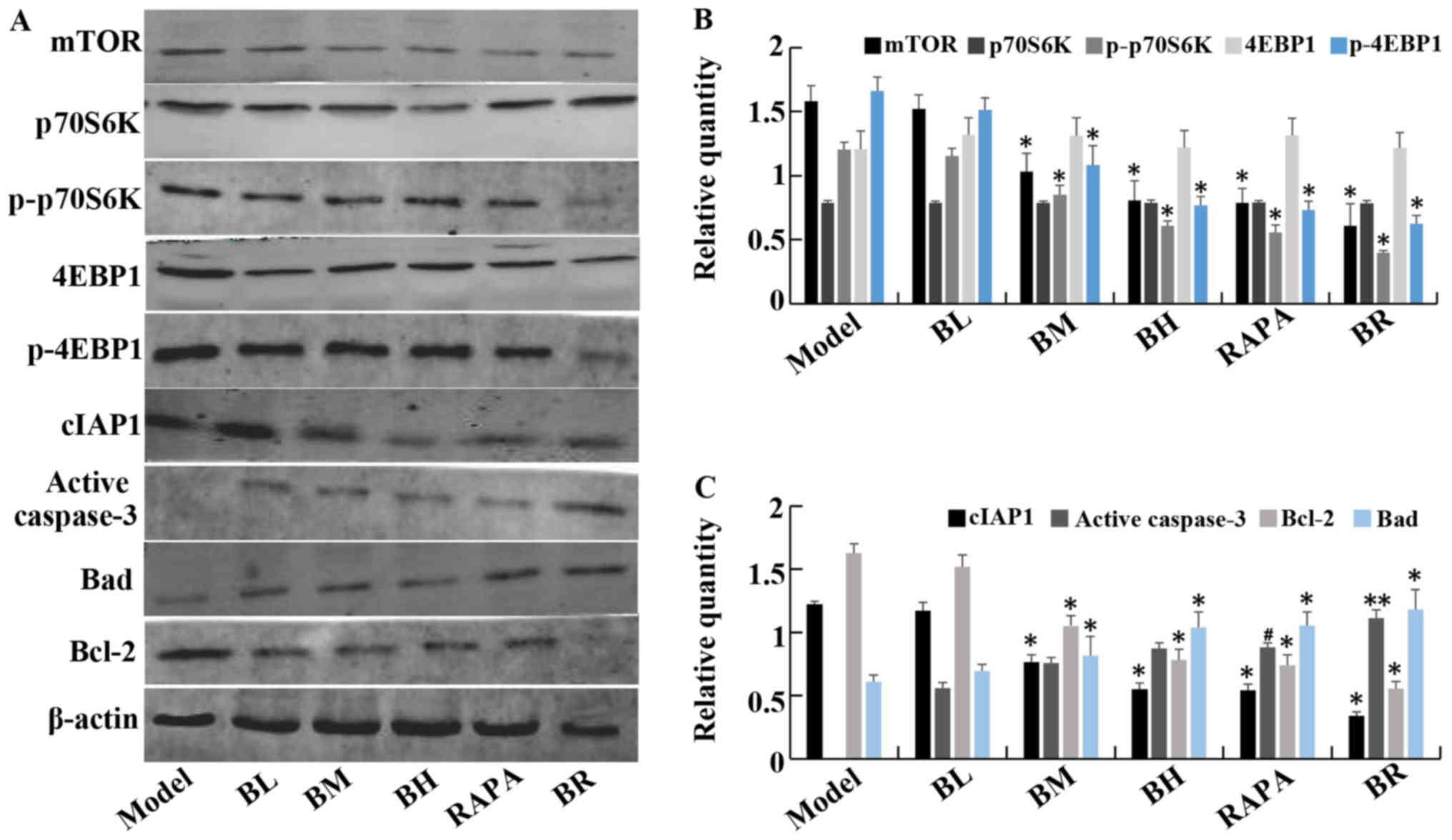 | Figure 5The expression of mTOR, P70S6K,
p-P70S6K, 4EBP1, p-4EBP1, cIAP1, active caspase-3, Bad, Bcl-2
protein in orthotopic transplanted tumor of human esophageal
squamous cell carcinoma (ESCC) in nude mice. (A) Western blotting
of mTOR, P70S6K, p-P70S6K, 4EBP1, p-4EBP1, cIAP1, active caspase-3,
Bad and Bcl-2 protein levels in each groups. (B) The relative
expression of mTOR, P70S6K, p-P70S6K, 4EBP1, p-4EBP1 in each
groups. (C) The relative expression of cIAP1, active caspase-3, Bad
and Bcl-2 in each groups. **P<0.05 vs. model;
#P<0.05 vs. medium-dose bufalin;
*P<0.05 vs. rapamycin and high-dose bufalin. |
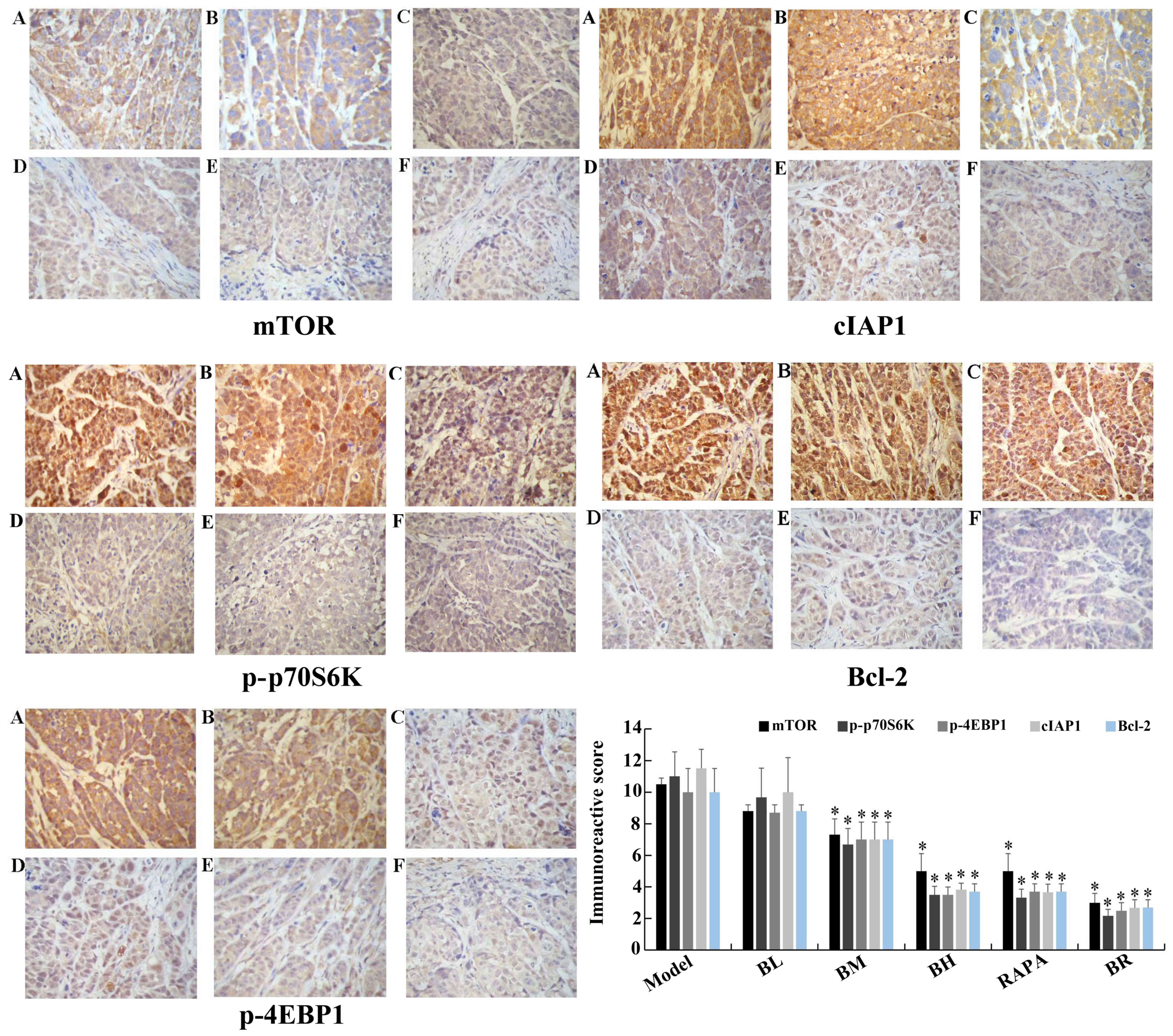
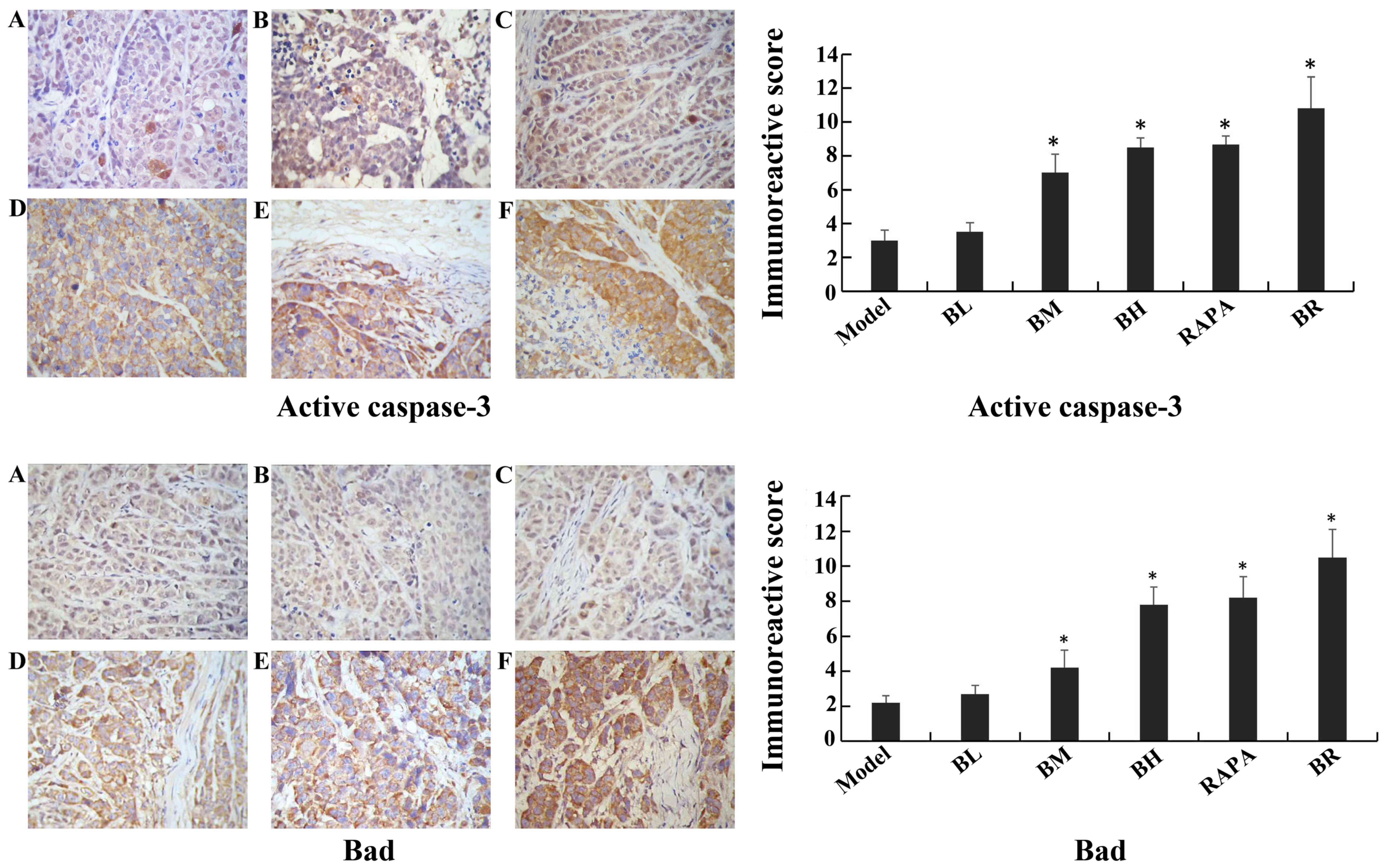
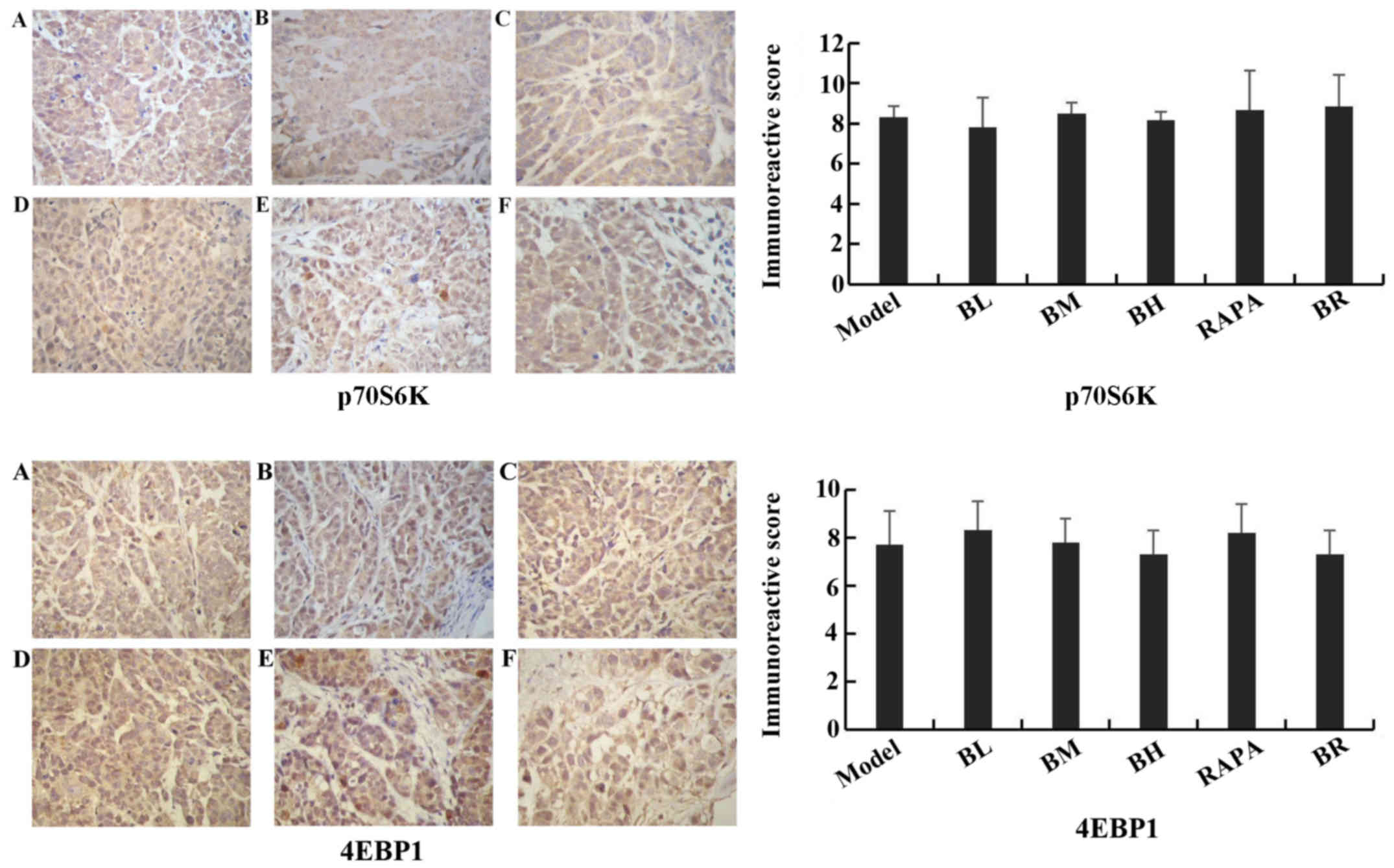
Immunohistochemistry assay
The immunoreactive scores for mTOR, p-p70S6K,
p-4EBP1, cIAP1 and Bcl-2 gradually decreased in each treatment
group (BL, BM, BH, RAPA and BR) (Fig.
6), whereas those for active caspase-3 and Bad gradually
increased compared to the model group (Fig. 7), with significant differences
observed (P<0.05). However, no significant differences were
observed between the model and BL groups (P>0.05), and between
the BH and RAPA groups (P>0.05). The immunoreactive scores for
p70S6K, 4EBP1 did not differ significantly between the groups
(model, BL, BM, BH, RAPA and BR (P>0.05) (Fig. 8).
Discussion
Bufalin is one of the main components extracted from
toad venom and is used in traditional Chinese medicine (TCM)
(23). It is known to exert
anti-tumor effects. Bufalin has been shown to exert anti-tumor,
diuretic, cardiac, blood pressure-promoting, anesthetic,
anti-inflammatory, anti-radiation and antitussive effects and has
been shown to inhibit the Na+/K+-ATP enzyme
(23). Krenn and Kopp suggested
that the most effective composition of bufalin had digoxin-like
immunoreactive effects (23).
Bufalin is an effective antitumor agent and has been applied in the
therapy of diverse malignant tumors (24). Studies have shown that cardiac
glycoside drugs inhibit the proliferation of tumor cells and induce
apoptosis in gastric cancer (25), prostate cancer (26), liver cancer (27), endometrial cancer (28), ovarian cancer (28) and colon cancer (29). Lee et al found that bufalin
inhibited tumor growth by inhibiting endothelial cell proliferation
and angiogenesis (30). Xia et
al found that the anti-tumor effects of bufalin were associated
with the induction of tumor cell apoptosis (31). Yamada et al found that the
anti-tumor effects of bufalin were associated with the induction of
tumor cell differentiation (32).
Currently, the exact mechanisms responsble for the anti-tumor
effects of bufalin are not clear. Xu et al indicated that
bufalin is an active compound of the traditional Chinese medicine,
Chansu, which exhibits significant anti-tumor activities in many
solid tumors and leukemia cell lines (33). Qiu et al also demonstrated
that bufalin inbibits the proliferation and invasion of
hepatocellular carcinoma cells (34). Our study demonstrated that bufalin
exerts anti-tumor effects. However, further research is required to
fully determine the effects of bufalin in ESCC.
A previous study suggested that bufalin decreased
the phosphorylation levels of extracellular signal-regulated kinase
(ERK) in vitro, while the levels of non-phosphorylation ERK
were not affected. It was suggested that the anti-tumor effects of
bufalin were associated with the activation of ERK (35).
Numerous studies have confirmed that the mTOR/p70S6K
signaling pathway plays a central role in cell survival, growth and
proliferation, and is significantly activated in a variety of
tumors, such as breast cancer, prostate cancer and cervical cancer,
and has become a novel target in cancer therapy (36–41). mTOR is a member of the PIKK
superfamily and is an untypical serine/threonine protein kinase. It
is highly conserved evolutionary (42). mTOR regulates protein synthesis,
cell growth, and proliferation and apoptosis (43). p70S6K is an important key factor
downstream of the mTOR pathway. It is a substrate which activates
mTOR directly and promotes protein synthesis. p70S6K is
phosphorylated into p-p70S6K through mTOR and the translational
activity of p-p70S6K is approximately 100-fold greater than that of
p70S6K (44). The activation
process of p70S6K is very complex, and it requires at least 7
serine/threonine phosphorylation sites (45). These can be divided into two
regions, one for Thr389, Thr229 and Ser404; phosphorylation at
Thr389 could be directly activated protein kinase. The second is
responsible for the regulation of kinase activity at the C-terminus
(Ser411, Ser424, Ser418 and Thr421) (45).
4EBP1 is a negative regulator of protein translation
translation in mammalian cells. When cells are exposed to external
stimuli, 4EBP1 is inactivated by activated mTOR which causes
dissociation from eIF-4E (46).
In the presence of mTOR inhibitors, 4EBP1 is dephosphorylated and
combines with eIF-4E, inhibiting the initiation of translation. In
our study, we measured the tumor volume in nude mice following
treatment with low- or high-dose bufalin and bufalin in combination
with rapamycin. Compared with the model group, tumor growth was
decreased in the BM, BH, RAPA and BR groups, whereas tumor growth
was not significantly altered in the BL group. This suggests that
bufalin inhibits tumor growth at medium and high doses, whereas it
has no significant inhibitory effect at lower doses. The results
from western blot analysis and immunohistochemistry indicated that
the expression of p-p70S6K and p-4EBP1 decreased following
treatment with bufalin and/or rapamycin. The most significant
decrease was observed in the BR (combination) group. Of note, the
expression of p70S6K and 4EBP1 exhibited no significant change. We
thus inferred that bufalin inhibited the phosphorylation and
activity of p70S6K and 4EBP1 by inhibiting the mTOR/p70S6K pathway.
Bufalin and rapamycin may thus inhibit the mTOR/p70S6K pathway
synergistically. Therefore, it can be concluded that the anti-tumor
effects of bufalin are mediated via the mTOR/p70S6K pathway and via
the inhibition of the activity of p70S6K and 4EBP1. The results of
RT-PCR demonstrated that the mRNA expression levels of p70S6K,
4EBP1 were not significantly altered in each group. Bufalin
inhibited the phosphorylation of p70S6K, which further confirmed
our above conclusions.
Recently, it has been shown that the occurrence of
malignant tumor is associated with the dysregulation of apoptosis.
Inhibitor of apoptosis proteins (IAPs) are a type of highly
conserved proteins which regulate the activity of caspase-3/-7/-9
and immune signals to inhibit the process of apoptosis. cIAP1
inhibits apoptosis mainly by suppressing the activity of
caspase-3/-7 and NF-κB (20).
cIAP1 plays a pivotal role in a large number of malignancies. In
China, the incidence of esophageal cancer is high and reports on
cIAP1 in esophageal cancer are limited. Xu et al suggested
that a high expression of cIAP1 in ESCC was associated with the
chemosensitivity of esophageal cancer (47). Zhang et al found that the
overexpression of XIAP in esophageal cancer and the downregulation
of XIAP was associated with RNA interference and could enhance the
chemotherapeutic sensitivity of esophageal cancer (48). In addition, caspase proteases
exist in the cytoplasm and can cleave peptide bonds in aspartic
acid residue of the target protein specifically (48). Caspase-3 is the most critical
terminal cleavage enzyme during apoptosis, which is the terminal
executive molecule downstream of caspase cascade. Caspase-3 is the
intersection of the death receptor pathway and mitochondrial
pathway. Thus, caspase-3 is also known as the death protease
(21). Normally, caspase-3 exists
in the cell cytoplasm as zymogen form (pro-caspase) When the
zymogen is stimulated by an apoptotic signal, pro-caspase can be
activated to translate into active-caspase-3, which is involved in
the process of apoptosis.
The Bcl-2 family plays a critical role in the
process of apoptosis, as it consists of anti-apoptotic proteins
(Bcl-2, Bcl-xL, Bcl-w and ced-9) and pro-apoptotic proteins (Bad,
Bax, Bak and Bcl-xS). Bcl-2 is anti-apoptotic and can inhibit cell
apoptosis, prolonging the life of cells, whereas Bad promotes cell
apoptosis (49). In this study,
apoptosis was detected by TUNEL assay. We found that the apoptotic
index was highest in the BL and BM groups, it was decreased in the
BH group. It was thus suggested that bufalin induces apoptosis
mainly at low- and medium-doses, while it can kill tumor cells
directly at high doses, leading to large areas of necrosis. In the
RAPA and BR groups, large areas of necrosis were observed, but the
apoptotic index was lower than that of the BL and BM groups. The
results of RT-PCR, western blot analysis and immunohistochemistry
revealed that the mRNA and protein expression of cIAP1 gradually
decreased in each group, suggesting that bufalin exerted a
dose-dependent effect. That is to say, with the increasing
concentration of bufalin, the expression of cIAP1 decreased, being
lowest in the BR group. Simultaneously, the expression of caspase-3
mRNA and active caspase-3 protein increased in each group. With the
inreasing concentrations of bufalin, the expression of Bad
increased, whereas that of Bcl-2 decreased. Thus, bufalin exerted
pro-apoptotic effects by regulating the expression levels of Bcl-2
and Bad. There was a certain dose-dependent effect. We speculated
that bufalin and rapamycin induced apoptosis synergistically. The
anti-tumor effects of bufalin may be associated with the
down-regulation of cIAP1 and the upregulation of of caspase-3 in
esophageal cancer cells. As cIAP1 can inhibit the activation of
caspase-3, with the decrease in the expression of cIAP1, the
caspase-3 was activated in the esophageal cancer cells. In this
way, caspase-3 is activated and can promote the initiation of the
apoptotic process in esophageal cancer cells.
In conclusion, our study demonstrates tht bufalin
exerts palpable anti-tumor effects on orthotopically transplanted
ESCC tumors in nude mice. Bufalin decreased the levels of
phosphorylated p70S6K and 4EBP1, whereas the levels of p70S6K and
4EBP1 were not affected. We thus suggest that bufalin inhibits
tumor growth by suppressing the activation of p70S6K and 4EBP1.
Bufalin also decreased the expression of cIAP1 and Bcl-2, and it
upregulated active caspase-3 and Bad simultaneously. Thus, the
induction of tumor cell apoptosis may be one of the critical
antitumor mechanisms of bufalin.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China. The authors would like to thank the
research specialist staff of Laboratory Animal Center at the Fourth
Hospital of Hebei Medical University. Finally, we would also like
to thank the professors and technical staff of the Department of
Pathology in the Fourth Hospital of Hebei Medical University.
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
2
|
Rustgi AK and El-Serag HB: Esophageal
carcinoma. N Engl J Med. 371:2499–2509. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siewert JR and Ott K: Are squamous and
adenocarcinomas of the esophagus the same disease? Semin Radiat
Oncol. 17:38–44. 2007. View Article : Google Scholar
|
|
4
|
Ou Y, Ma L, Ma L, Huang Z, Zhou W, Zhao C,
Zhang B, Song Y, Yu C and Zhan Q: Overexpression of cyclin B1
antagonizes chemotherapeutic-induced apoptosis through PTEN/Akt
pathway in human esophageal squamous cell carcinoma cells. Cancer
Biol Ther. 14:45–55. 2013. View Article : Google Scholar :
|
|
5
|
Tong Q, Zhang W, Jin S, Li S and Chen Z:
The relationship between p27(kip1) expression and the change of
radiosensitivity of esophageal carcinoma cells. Scand J
Gastroenterol. 46:173–176. 2011. View Article : Google Scholar
|
|
6
|
Imoto I, Yang ZQ, Pimkhaokham A, Tsuda H,
Shimada Y, Imamura M, Ohki M and Inazawa J: Identification of cIAP1
as a candidate target gene within an amplicon at 11q22 in
esophageal squamous cell carcinomas. Cancer Res. 61:6629–6634.
2001.PubMed/NCBI
|
|
7
|
Dhar R, Persaud SD, Mireles JR and Basu A:
Proteolytic cleavage of p70 ribosomal S6 kinase by caspase-3 during
DNA damage-induced apoptosis. Biochemistry. 48:1474–1480. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fingar DC, Richardson CJ, Tee AR, Cheatham
L, Tsou C and Blenis J: mTOR controls cell cycle progression
through its cell growth effectors S6K1 and 4E-BP1/eukaryotic
translation initiation factor 4E. Mol Cell Biol. 24:200–216. 2004.
View Article : Google Scholar :
|
|
9
|
Neufeld TP: TOR-dependent control of
autophagy: biting the hand that feeds. Curr Opin Cell Biol.
22:157–168. 2010. View Article : Google Scholar :
|
|
10
|
Ganley IG, Lam du H, Wang J, Ding X, Chen
S and Jiang X: ULK1.ATG13.FIP200 complex mediates mTOR signaling
and is essential for autophagy. J Biol Chem. 284:12297–1305. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zinzalla V, Stracka D, Oppliger W and Hall
MN: Activation of mTORC2 by association with the ribosome. Cell.
144:757–768. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oh WJ, Wu CC, Kim SJ, Facchinetti V,
Julien LA, Finlan M, Roux PP, Su B and Jacinto E: mTORC2 can
associate with ribosomes to promote cotranslational phosphorylation
and stability of nascent Akt polypeptide. EMBO J. 29:3939–3951.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cornu M and Hall MN: mTORC1 and mTORC2 in
energy homeostasis. The Enzymes: Structure, Function and Regulation
of TOR Complexes from Yeasts to Mammals. Tamanoi F and Hall MN:
Academic Press; Amsterdam: pp. 263–278. 2010, View Article : Google Scholar
|
|
14
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Um SH, Frigerio F, Watanabe M, Picard F,
Joaquin M, Sticker M, Fumagalli S, Allegrini PR, Kozma SC, Auwerx J
and Thomas G: Absence of S6K1 protects against age- and
diet-induced obesity while enhancing insulin sensitivity. Nature.
431:200–205. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ly C, Arechiga AF, Melo JV, Walsh CM and
Ong ST: Bcr-Abl kinase modulates the translation regulators
ribosomal protein S6 and 4E-BP1 in chronic myelogenous leukemia
cells via the mammalian target of rapamycin. Cancer Res.
63:5716–5722. 2003.PubMed/NCBI
|
|
18
|
Kharas MG, Deane JA, Wong S, O'Bosky KR,
Rosenberg N, Witte ON and Fruman DA: Phosphoinositide 3-kinase
signaling is essential for ABL oncogene-mediated transformation of
B-lineage cells. Blood. 103:4268–4275. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y and Li X: The research progress of
p70S6K1 and 4E-BPs of the AKt/mTOR signal pathway in promoting the
occurrence of tumor. J Modern Oncol. 24:847–850. 2016.
|
|
20
|
Xu Y, Liu F, Zhou LP and Zhao XH: c-IAP1
expression and tumor chemosensitivity in esophageal squamous cell
carcinoma. World Chin J Digestol. 19:1138–1144. 2011.In
Chinese.
|
|
21
|
Wang Q, Huang Y, Ni Y, Wang H and Hou Y:
siRNA targeting midkine inhibits gastric cancer cells growth and
induces apoptosis involved caspase-3,8,9 activation and
mitochondrial depolarization. J Biomed Sci. 14:783–795. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu MY, Hou GQ, Zhang Y, Bei WJ and Yan
AH: Effects of mTOR siRNA on mTOR/p70S6K signaling pathway in
esophageal squamous cell carcinoma cells and the growth of
transplanted tumor in nude mice. Zhonghua Zhong Liu Za Zhi.
33:334–339. 2011.In Chinese. PubMed/NCBI
|
|
23
|
Krenn L and Kopp B: Bufadienolides from
animal and plant sources. Phytochemistry. 48:1–29. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yin PH, Liu X, Qiu YY, Cai JF, Qin JM, Zhu
HR and Li Q: Anti-tumor activity and apoptosis-regulation
mechanisms of bufalin in various cancers: new hope for cancer
patients. Asian Pac J Cancer Prev. 13:5339–5343. 2012. View Article : Google Scholar
|
|
25
|
Li D, Qu X, Hou K, Zhang Y, Dong Q, Teng
Y, Zhang J and Liu Y: PI3K/Akt is involved in bufalin-induced
apoptosis in gastric cancer cells. Anticancer Drugs. 20:59–64.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhai XF, Fang FF, Liu Q, Meng YB, Guo YY
and Chen Z: miR-181a contributes to bufalin-induced apoptosis in
PC-3 prostate cancer cells. BMC Complement Altern Med. 13:3252013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qi F, Inagaki Y, Gao B, Cui X, Xu H,
Kokudo N, Li A and Tang W: Bufalin and cinobufagin induce apoptosis
of human hepato-cellular carcinoma cells via Fas- and
mitochondria-mediated pathways. Cancer Sci. 102:951–958. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Takai N, Ueda T, Nishida M, Nasu K and
Narahara H: Bufalin induces growth inhibition, cell cycle arrest
and apoptosis in human endometrial and ovarian cancer cells. Int J
Mol Med. 21:637–643. 2008.PubMed/NCBI
|
|
29
|
Xie CM, Chan WY, Yu S, Zhao J and Cheng
CH: Bufalin induces autophagy-mediated cell death in human colon
cancer cells through reactive oxygen species generation and JNK
activation. Free Radic Biol Med. 51:1365–1375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee DY, Yasuda M, Yamamoto T, Yoshida T
and Kuroiwa Y: Bufalin inhibits endothelial cell proliferation and
angiogenesis in vitro. Life Sci. 60:127–134. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xia J, Jiang M and Jiang Y: The research
progress of the anti-tumor effect of toad venom and its effective
components in vitro. Modern Oncol. 16:1441–1444. 2008.In
Chinese.
|
|
32
|
Yamada K, Hino K, Tomoyasu S, Honma Y and
Tsuruoka N: Enhancement by bufalin of retinoic acid-induced
differentiation of acute promyelocytic leukemia cells in primary
culture. Leuk Res. 22:589–595. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu Y, Chen M, Jin XF, Qian C, Xu XM and
Zhang X: Research progress of in vitro and in vivo anti-tumor
effects and formulation of bufalin. Zhongguo Zhong Yao Za Zhi.
39:2829–2833. 2014.In Chinese. PubMed/NCBI
|
|
34
|
Qiu DZ, Zhang ZJ, Wu WZ and Yang YK:
Bufalin, a component in Chansu, inhibits proliferation and invasion
of hepatocellular carcinoma cells. BMC Complement Altern Med.
13:1852013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ding Y, Wang XL and Deng HY: Mechanism of
bufalin affecting the proliferation and migration of human
esophageal carcinoma cells through inhibiting the activity of
Raf/MEK/ERK pathway. Chinese General Practice. 212015.In
Chinese.
|
|
36
|
Easley CA IV, Ben-Yehudah A, Redinger CJ,
Oliver SL, Varum ST, Eisinger VM, Carlisle DL, Donovan PJ and
Schatten GP: mTOR-mediated activation of p70S6K induces
differentiation of pluripotent human embryonic stem cells. Cell
Reprogram. 12:263–273. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Smolewski P: Recent developments in
targeting the mammalian target of rapamycin (mTOR) kinase pathway.
Anticancer Drugs. 17:487–494. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang YJ, Dai Q, Sun DF, Xiong H, Tian XQ,
Gao FH, Xu MH, Chen GQ, Han ZG and Fang JY: mTOR signaling pathway
is a target for the treatment of colorectal cancer. Ann Surg Oncol.
16:2617–2628. 2009.In Chinese. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li G, Shan C, Liu L, Zhou T, Zhou J, Hu X,
Chen Y, Cui H and Gao N: Tanshinone IIA inhibits HIF-1α and VEGF
expression in breast cancer cells via
mTOR/p70S6K/RPS6/4EBP1signaling pathway. PLoS One.
10:e01174402015.In Chinese. View Article : Google Scholar
|
|
40
|
Saraswati S, Kumar S and Alhaider AA:
α-santalol inhibits the angiogenesis and growth of human prostate
tumor growth by targeting vascular endothelial growth factor
receptor 2-mediated AKT/mTOR/P70S6K signaling pathway. Mol Cancer.
12:1472013. View Article : Google Scholar
|
|
41
|
Shin JM, Jeong YJ, Cho HJ, Park KK, Chung
IK, Lee IK, Kwak JY, Chang HW, Kim CH, Moon SK, et al: Melittin
suppresses HIF-1α/VEGF expression through inhibition of ERK and
mTOR/p70S6K pathway in human cervical carcinoma cells. PLoS One.
8:e693802013.In Chinese. View Article : Google Scholar
|
|
42
|
Zheng J: mTOR signal pathway and tumor.
Life Sciences. 18:261–265. 2006.
|
|
43
|
Pan Z, Zhang L and Jiang J: The research
progress of mTOR. Chin J Cell Biol. 28:395–398. 2006.
|
|
44
|
Jefferies HB, Fumagalli S, Dennis PB,
Reinhard C, Pearson RB and Thomas G: Rapamycin suppresses 5′TOP
mRNA translation through inhibition of p70s6k. EMBO J.
16:3693–3704. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cheng HD, Zhao QN, Song WD, Sarengaowa and
Liu F: The current research of the component p-P70S6K of mTOR
signal pathway and colon cancer. Journal of Disease Monitor and
Control. 6:78–79. 2012.
|
|
46
|
Asimomytis A, Karanikou M, Rodolakis A,
Vaiopoulou A, Tsetsa P, Creatsas G, Stefos T, Antsaklis A,
Patsouris E and Rassidakis GZ: mTOR downstream effectors, 4EBP1 and
eIF4E, are overexpressed and associated with HPV status in
precancerous lesions and carcinomas of the uterine cervix. Oncol
Lett. 12:3234–3240. 2016.PubMed/NCBI
|
|
47
|
Xu Y and Zhao XH: IAP family and
IAP-targeted therapy. Life Sciences. 22:161–168. 2010.
|
|
48
|
Zhang S, Ding F, Luo A, Chen A, Yu Z, Ren
S, Liu Z and Zhang L: XIAP is highly expressed in esophageal cancer
and its downregulation by RNAi sensitizes esophageal carcinoma cell
lines to chemotherapeutics. Cancer Biol Ther. 6:973–980. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cong YM, Liu P, Jin DP, et al: The
relationship of Bcl-2 family with cell apoptosis. J Chin Med Res.
8:125–127. 2008.In Chinese.
|