Introduction
Osteoarthritis (OA) is a chronic disease that
affects the joint cartilage of middle-aged and elderly individuals,
which is associated with degradation of articular cartilage
(1). Articular cartilage is an
avascular tissue with limited regenerative ability, which is
composed of chondrocytes and extracellular matrix (ECM) components.
As the only cell type in articular cartilage, chondrocytes control
the balance between catabolism and anabolism, in order to maintain
appropriate functioning of the ECM (2,3).
During the process of OA, chondrocyte death is associated with
degradation of the ECM and calcification, thus suggesting a role
for cell death in the pathogenesis of OA. Therefore, enhancing
chondrocyte proliferation may be a potential method to inhibit the
development and progression of OA.
As a vital process for cell proliferation, the cell
cycle is composed of four key phases: G1, S (DNA
replication), G2 and M (mitosis). Between the S and M
phases, there are two gaps, G1 prior to the S phase and
G2 prior to the M phase (4). The activity of cyclin D1, and its
binding partners cyclin-dependent kinase (CDK)4/6, controls the
progression through every phase of the cell cycle and is implicated
in chondrocyte proliferation (5,6).
The normal function of cartilage is affected by numerous signaling
pathways, including the Wnt/β-catenin signaling pathway (7–9).
The Wnt/β-catenin signaling pathway serves an important role in the
regulation and control of cell proliferation and maintenance of
pheno-typic characteristics, and is important for the regulation of
cartilage function (10). Binding
of Wnt proteins to cell surface receptor complexes, which are
composed of Frizzled proteins and low-density lipoprotein
receptor-related proteins (LRP)5/6, results in the activation of
dishevelled, after which glycogen synthase kinase-3β (GSK-3β)
activity and β-catenin phosphorylation are inhibited.
Non-phosphorylated β-catenin accumulates in the cytoplasm and
translocates to the nucleus, where it binds to the transcription
factors T-cell factor (TCF) and lymphoid enhancer factor (LEF), in
order to regulate target gene expression (11,12). These factors accelerate cell cycle
progression by accommodating the expression of cyclin D1 (13). It has previously been reported
that the Wnt/β-catenin signaling pathway acts upstream of cyclin D1
(14). Therefore, the
Wnt/β-catenin pathway may have an important role in cell
proliferation via the regulation of cyclin D1.
Glucosamine (GlcN) is an amino-monosaccharide
synthesized from glucose, which is used for the biosynthesis of
glycoproteins and glycosaminoglycans (15). GlcN is a natural compound present
in the majority of human tissues, with the highest concentrations
detected in cartilage (16). GlcN
is widely used in the clinical treatment of OA, due to its
fundamental role in stimulating the metabolism of chondrocytes
(17,18). However, the molecular mechanisms
underlying the effects of GlcN on chondrocytes remain unclear.
Therefore, the present study aimed to clarify the mechanisms
underlying the effects of GlcN on chondrocytes.
Materials and methods
Preparation of GlcN
GlcN was obtained from Shanghai Aladdin Biochemical
Technology Co., Ltd. (Shanghai, China) and was dissolved in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS) (both from HyClone; GE Healthcare Life
Sciences, Logan, UT, USA). Subsequently, GlcN was filtered through
a 0.22-μm filter.
Isolation, culture and identification of
chondrocytes
Male, 4-week-old, Sprague-Dawley specific
pathogen-free rats (weight, 200–300 g) were purchased from Shanghai
SLAC Laboratory Animal Co., Ltd. (Shanghai, China). The care and
use of animals in the present study were conducted in strict
accordance with the Guide for the Care and Use of Laboratory
Animals of Fujian University of Traditional Chinese Medicine
(Fuzhou, China). The present study was approved by the Ethics
Committee of Fujian University of Traditional Chinese Medicine.
Sprague-Dawley rats were sacrificed using carbon dioxide (cage
size, 7×11×5 inches; flow rate, 1.3 l/min), according to the Guide
for the Care and Use of Laboratory Animals.
Chondrocytes were isolated from the knee cartilage
of Sprague-Dawley rats and were cultured as previously described
(19). Briefly, four
Sprague-Dawley rats were sacrificed, and their knees were stripped
and soaked in 75% ethanol for 15 min. The articular cartilage was
cut open in the bilateral knee joints, collected with a blade and
washed with PBS (HyClone; GE Healthcare Life Sciences) three times
under sterile conditions. The cartilage samples were cut into
1-mm3 sections and digested with 0.2% collagenase II
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 37°C in an
incubator containing 5% CO2. The supernatant fluid was
collected every 2 h, and was centrifuged at 110 × g for 5 min. The
isolated cells were resuspended in DMEM containing 10% FBS, and
were cultured in 50 ml culture flasks containing 4 ml DMEM
supplemented with 10% FBS at 37°C in a 5% CO2 incubator.
Primary chondrocytes were termed passage 0 (P0), and P2
chondrocytes at ~80% confluence were used in subsequent
experiments. P2 chondrocytes were identified by collagen II
immunohistochemistry (20).
Briefly, the cells were seeded onto a sterilized round coverslip
and placed in a 6-well plate (2×104 cells/well); the
cells were cultured at 37°C in a 5% CO2 incubator for 48
h. Subsequently, the cells were fixed with 4% paraformaldehyde
(Sigma-Aldrich; Merck KGaA) for 30 min at 4°C and blocked with 10%
bovine serum albumin (BSA) (Sigma-Aldrich; Merck KGaA) for 1 h. The
round coverslip was then incubated with a type II collagen rabbit
polyclonal antibody (dilution 1:500; BS1071; Bioworld Technology,
Inc., St. Louis Park, MN, USA) overnight at 4°C. The round
coverslip was then incubated with horseradish peroxidase
(HRP)-conjugated affinipure goat anti-rabbit immunoglobulin (Ig)G
(dilution: 1:200; ZB-2301) for 30 min, followed by incubation with
diaminobenzidine (ZLI-9018) (both from OriGene Technologies, Inc.,
Beijing, China) for 5 min at room temperature. The slides were
finally counterstained with hematoxylin (Sigma-Aldrich; Merck KGaA)
and were dehydrated. Images were captured under a light microscope
(BH2; Olympus Corporation, Tokyo, Japan). According to the MTT
assay, chondrocytes were treated with 0, 100, 200 and 300
μg/ml GlcN for 72 h. In order to further verify the mecha
nisms involved, chondrocytes were treated with 0.2 μg/ml
Dickkopf-1 (DKK-1; R&D Systems, Inc., Minneapolis, MN, USA) and
were treated with GlcN (200 μg/ml) in the presence or
absence of DKK-1 for 72 h at 37°C.
Evaluation of cell viability by MTT
assay
The P2 chondrocytes were plated onto a 96-well plate
at a density of 1.0×104/ml and were cultured for 24 h.
The cells were then treated with various concentrations of GlcN
(50, 100, 200, 300 and 600 μg/ml) for 24 or 72 h.
Subsequently, 100 μl MTT (1 mg/ml in PBS) was added to each
well and was incubated at 37°C for 4 h. The supernatant was then
removed and 150 μl dimethyl sulfoxide was added to dissolve
the formazan. The solution was agitated for 10 min and the optical
density was measured at 490 nm using an ELISA reader (model ELx800;
BioTek Instruments, Inc., Winooski, VT, USA).
Cell cycle analysis
Following treatment with GlcN, chondrocytes were
collected and chondrocyte density was adjusted to
1×106/ml. The process of staining was performed using a
cell cycle assay kit (BD Cycletest™ Plus DNA reagent kit; BD
Biosciences, Franklin Lakes, NJ, USA) according to the
manufacturer's protocol. Subsequently, the cells were analyzed
using fluorescence-activated cell sorting (FACSCalibur™; BD
Biosciences, San Diego, CA, USA). The percentage of cells in the
different phases of the cell cycle, including
G0/G1, S, G2 and M phases, was
calculated using ModFit software 4.0 (Verity Software House,
Topsham, ME, USA).
RNA extraction and reverse
transcription-polymerase chain reaction (RT-PCR) analysis
Following treatment with GlcN, total RNA was
extracted using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) according to the
manufacturer's protocol. Total RNA (1 μg) was reverse
transcribed into cDNA using a RevertAid First Strand cDNA Synthesis
kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The following primers were used for PCR to
determine the mRNA expression levels of CDK4, CDK6, cyclin D1,
Wnt-4, β-catenin, Frizzled-2 and GSK-3β: CDK4, forward 5′-GAA GAC
GAC TGG CCT CGA GA-3′, reverse, 5′-ACT GCG CTC CAG ATT CCT CC-3′;
CDK6, forward 5′-TTG TGA CAG ACA TCG ACG AG-3′, reverse 5′-GAC AGG
TGA GAA TGC AGG TT-3′; cyclin D1, forward 5′-AAT GCC AGA GGC GGA
TGA GA-3′, reverse 5′-GCT TGT GCG GTA GCA GGA GA-3′; Wnt-4, forward
5′-TCA GCC CAC AGG GTT TCC A-3′, reverse 5′-CGC TCG CCA GCA TGT CTT
T-3′; β-catenin, forward 5′-AAG GAA GCT TCC AGA CAT GC-3′, reverse
5′-AGC TTG CTC TCT TGA TTG CC-3′; Frizzled-2, forward 5′-TCG AGG
CCA ATT CGC AGT A-3′, reverse 5′-CAG GAA GGA TGT GCC GAT G-3′;
GSK-3β, forward 5′-AAA GTG CAT CGC TGG CTT A-3′, reverse 5′-GTC GAC
GGT TTG TTT CCA AT-3′; and β-actin, forward 5′-CAC CCG CGA GTA CAA
CCT TC-3′ and reverse 5′-CCC ATA CCC ACC ATC ACA CC-3′. The
thermocycling conditions (annealing temperature, annealing time and
number of cycles) were as follows: Wnt-4: 57°C, 45 sec, 35 cycles;
Frizzeld-2: 58°C, 45 sec, 35 cycles; Gsk-3β: 60°C, 30 sec, 28
cycles; β-catenin: 60°C, 45 sec, 35 cycles; Cyclin D1: 55°C, 30
sec, 35 cycles; CDK4: 60°C, 40 sec, 32 cycles; CDK6: 60°C, 30 sec,
30 cycles. The DNA bands were examined via gel electrophoresis
(1.5% agarose) using a gel documentation system (Model Gel Doc
2000; Bio-Rad Laboratories, Inc., Hercules, CA, USA). β-actin was
used as an internal control.
Western blot analysis
Following treatment with GlcN, proteins were
collected from the cells using radioimmunoprecipitation assay lysis
buffer supplemented with 1 mM phenylmethanesulfonyl fluoride (both
from Beyotime Institute of Biotechnology, Shanghai, China). Protein
concentration was quantified using the bicinchoninic acid assay
method. Proteins (20 μg) were separated by 12% SDS-PAGE and
were transferred onto polyvinylidene fluoride membranes (Thermo
Fisher Scientific, Inc.). After transfer, the membranes were
blocked in 5% skimmed milk for 1 h at room temperature and were
incubated overnight at 4°C with the following primary antibodies
(1:1,000): CDK4 (sc-260), CDK6 (sc-177), Wnt-4 (sc-5214),
Frizzled-2 (sc-68327), cyclin D1 (sc-718), β-actin (sc-47778)
(Santa Cruz Biotechnology, Inc., Dallas, TX, USA), GSK-3β (9315)
and β-catenin (9582) (Cell Signaling Technology, Inc., Beverly, MA,
USA). Membranes were then incubated with HRP-conjugated secondary
antibodies [anti-goat (dilution 1:500; ZDR-5105); anti-rabbit
(dilution 1:500; ZDR-5306); anti-mouse (dilution 1:1,000;
ZDR-5109), Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd.,
Beijing, China] at room tempera ture for 1 h. The immuno-complexes
were detected using a Bio-Rad ChemiDoc XRS+ imaging
system (Image Lab 3.0; Bio-Rad Laboratories, Inc.). β-actin was
used as an internal control.
Immunohistochemistry
Following treatment with GlcN, chondrocytes were
fixed with 4% paraformaldehyde for 15 min at room temperature and
were permeabilized with 1% Triton X-100 in PBS for 10 min at room
temperature. Subsequently, the chondrocytes, fixed with
paraformaldehyde and permeabilized with 1% Triton X-100 in PBS,
were washed three times and were blocked with 5% BSA in PBS for 1 h
at room temperature. After blocking, the chondrocytes were
incubated with rabbit anti-β-catenin antibody (dilution 1:200;
sc-7199; Santa Cruz Biotechnology, Inc.) overnight at 4°C. After
exposure to the primary antibody, the chondrocytes were washed and
incubated with the corresponding secondary antibody (DyLight 488
AffiniPure Goat Anti-Rabbit IgG) for 1 h at room temperature.
Finally, DAPI was used to stain cell nuclei at room temperature for
5 min. The signal was visualized and images were acquired using a
fluorescence microscope (LSM710; Carl Zeiss AG, Oberkochen,
Germany).
Statistical analysis
Data are presented as the means ± standard
deviation. The experiments were repeated 3 times. Data were
processed using SPSS software version 18.0 (SPSS, Inc., Chicago,
IL, USA) and were analyzed with Student's t-test or analysis of
variance. The multiple comparisons tests used were least
significant difference (LSD) and Dunnett tests. P<0.05 was
considered to indicate a statistically significant difference.
Results
Morphology and characteristics of
chondrocytes
Chondrocyte morphology has been described in
previous studies (21,22); the chondrocytes in the present
study exhibited a spherical, fusiform and slabstone shape, which is
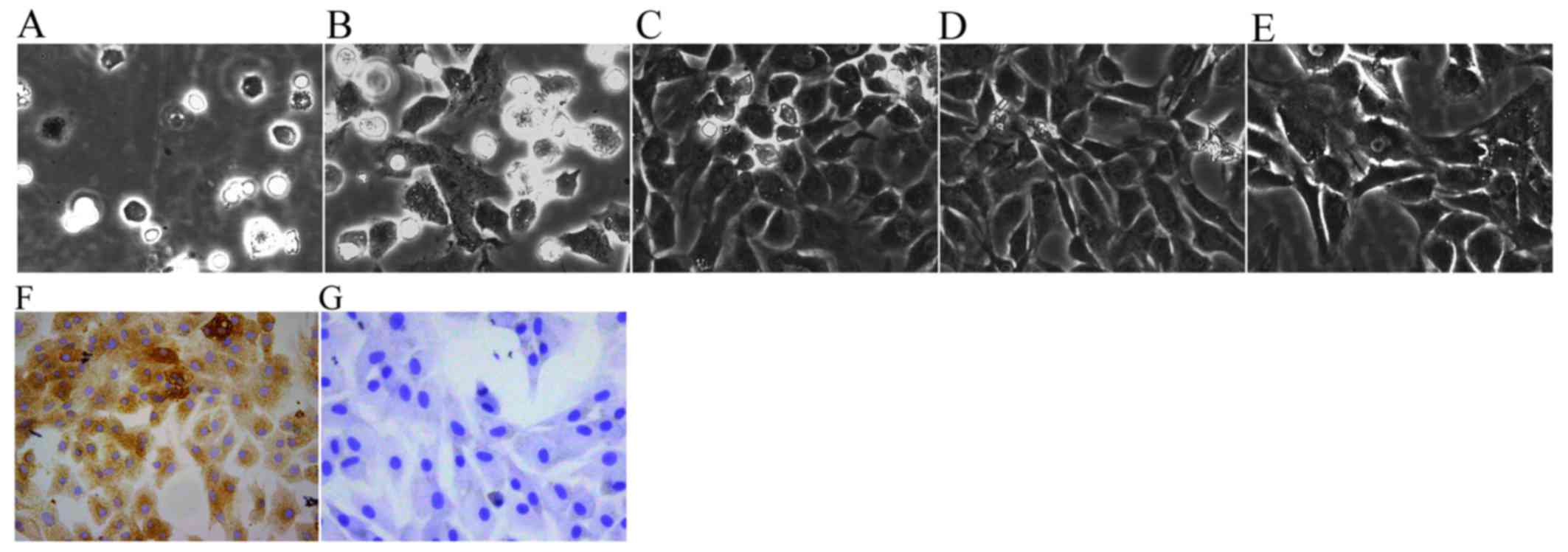
typical of chondrocytes (Fig. 1).
The newly isolated chondrocytes were small and round when first
suspended in DMEM. After being cultured for 2 h, the chondrocytes
gradually attached themselves to the culture flask (Fig. 1A). After being cultured for 3
days, the volumes of adherent cells became larger and some cells
began to elongate and form a fusiform shape (Fig. 1B). After 6 days of proliferation,
the cells coated the whole bottom of the culture flask (Fig. 1C). P1 and P2 chondrocytes spread
across the flask more rapidly and usually reached 80–90% density
within ~5 days (Fig. 1D and E).
The cytoplasm of P2 chondrocytes was stained brown, which indicated
that the P2 chondrocytes exhibited a more typical chondrocyte
morphology and contained large levels of collagen II (Fig. 1F) compared with in the negative
control cells (Fig. 1G).
Therefore, in view of the typical characteristics of P2 cells, P2
chondrocytes were used in subsequent experiments.
GlcN enhances chondrocyte viability and
promotes cell cycle progression
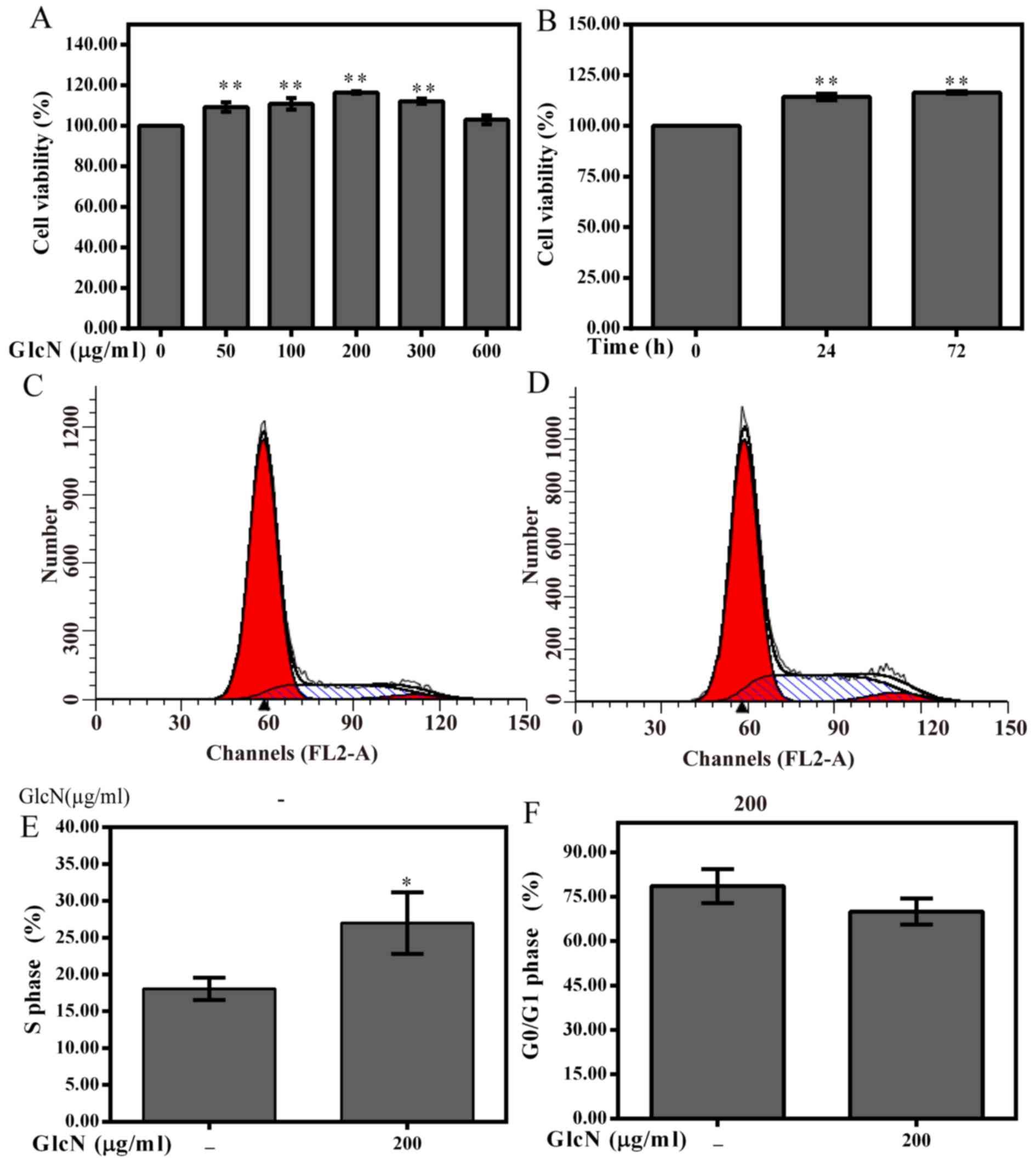
To investigate whether GlcN exerts effects on
chondrocyte viability, chondrocytes were treated with various
concentrations of GlcN (50, 100, 200, 300 and 600 μg/ml) for
72 h or with 100 μg/ml GlcN for the indicated time periods
(Fig. 2). GlcN treatment promoted
the viability of cells in a dose- and time-dependent manner
(Fig. 2A and B). According to the
MTT assay results, cell viability was markedly enhanced following
treatment with 200 μg/ml GlcN for 72 h; therefore, 0, 100,
200 and 300 μg/ml GlcN and 72 h were set as variables for
further experimentation.
As shown in Fig.
2C–F, the percentage of GlcN-treated chondrocytes in the
G0/G1 phase was reduced, whereas the
percentage of cells in S phase was increased compared with in the
untreated chondrocytes. Furthermore, the present study investigated
whether cell cycle progression was associated with an increase in
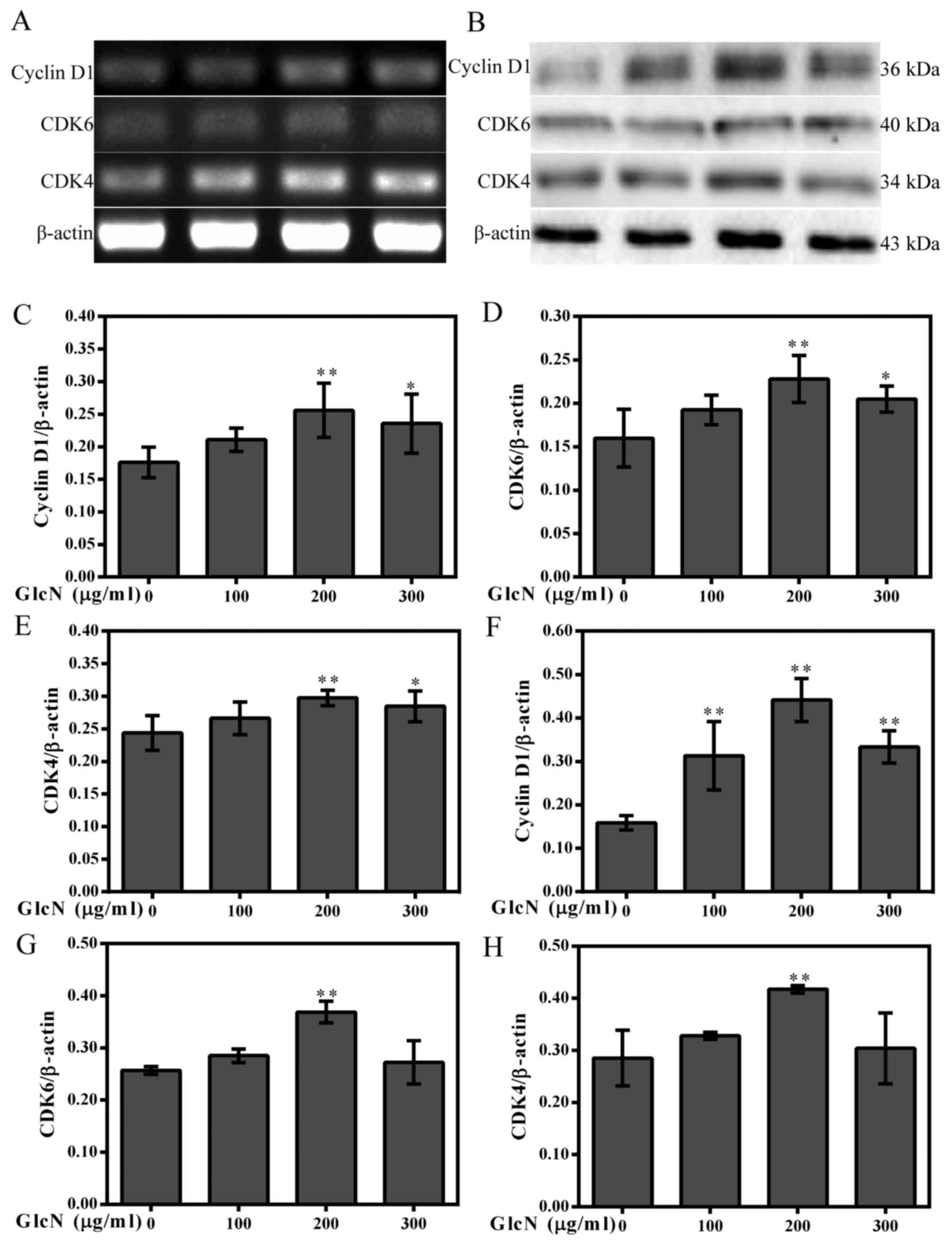
the protein expression levels of cyclins and CDKs. The results
indicated that GlcN upregulated the mRNA and protein expression
levels of cyclin D1, CDK4 and CDK6 (Fig. 3). These results indicated that
GlcN caused an increase of cell proliferation.
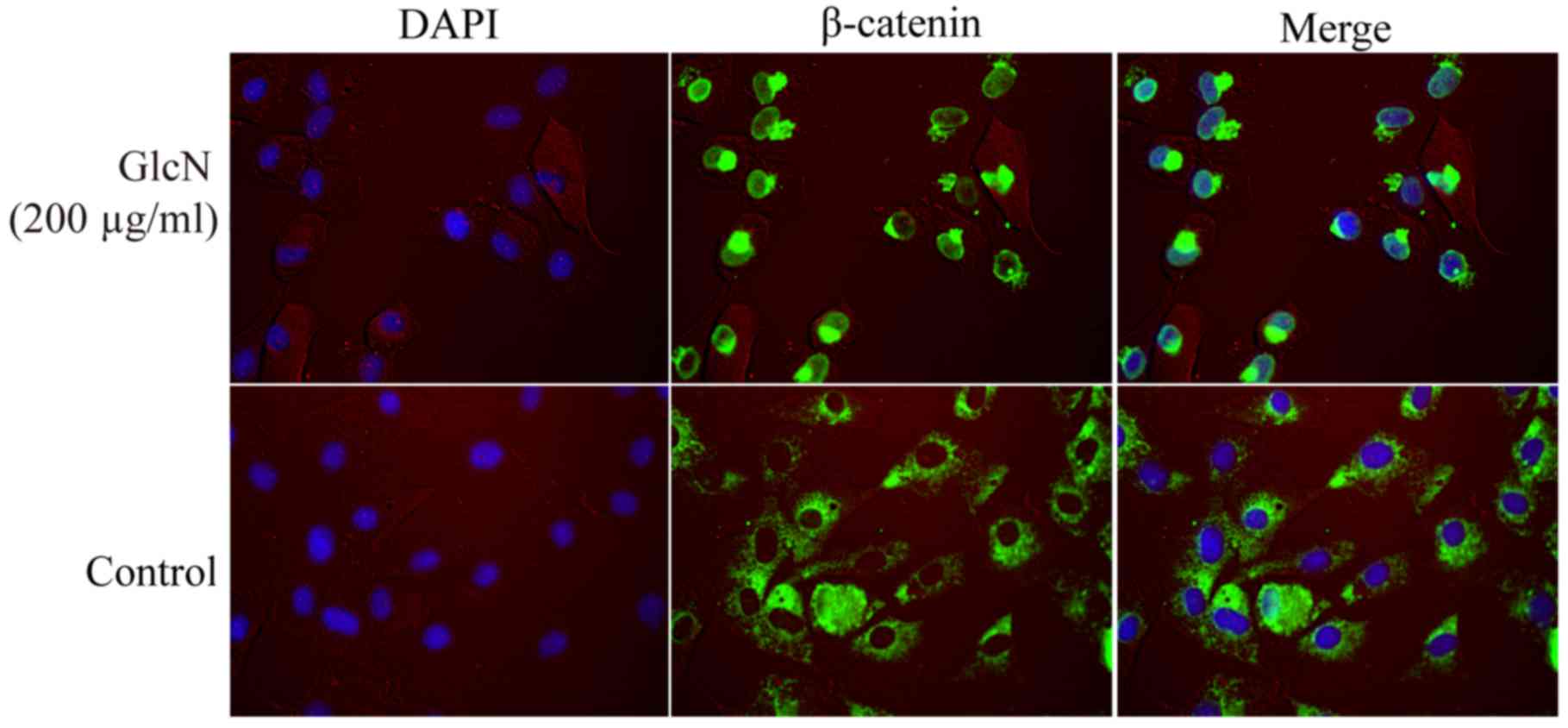
GlcN promotes the expression and nuclear
translocation of β-catenin
β-catenin is a well-known inducer of chondrocyte
proliferation and a cytoplasmic protein, which interacts with
TCF/LEF proteins to activate target genes, including cyclin D1
(23). The present study detected
β-catenin nuclear translocation and expression using
immunofluorescence, and western blotting and RT-PCR, respectively
(Figs. 4 and 5). The present results demonstrated that
GlcN may promote the mRNA and protein expression levels of
β-catenin (Fig. 5A, B, F and J).
The effects of GlcN on the nuclear translocation of β-catenin were
further confirmed by immunofluorescence staining.
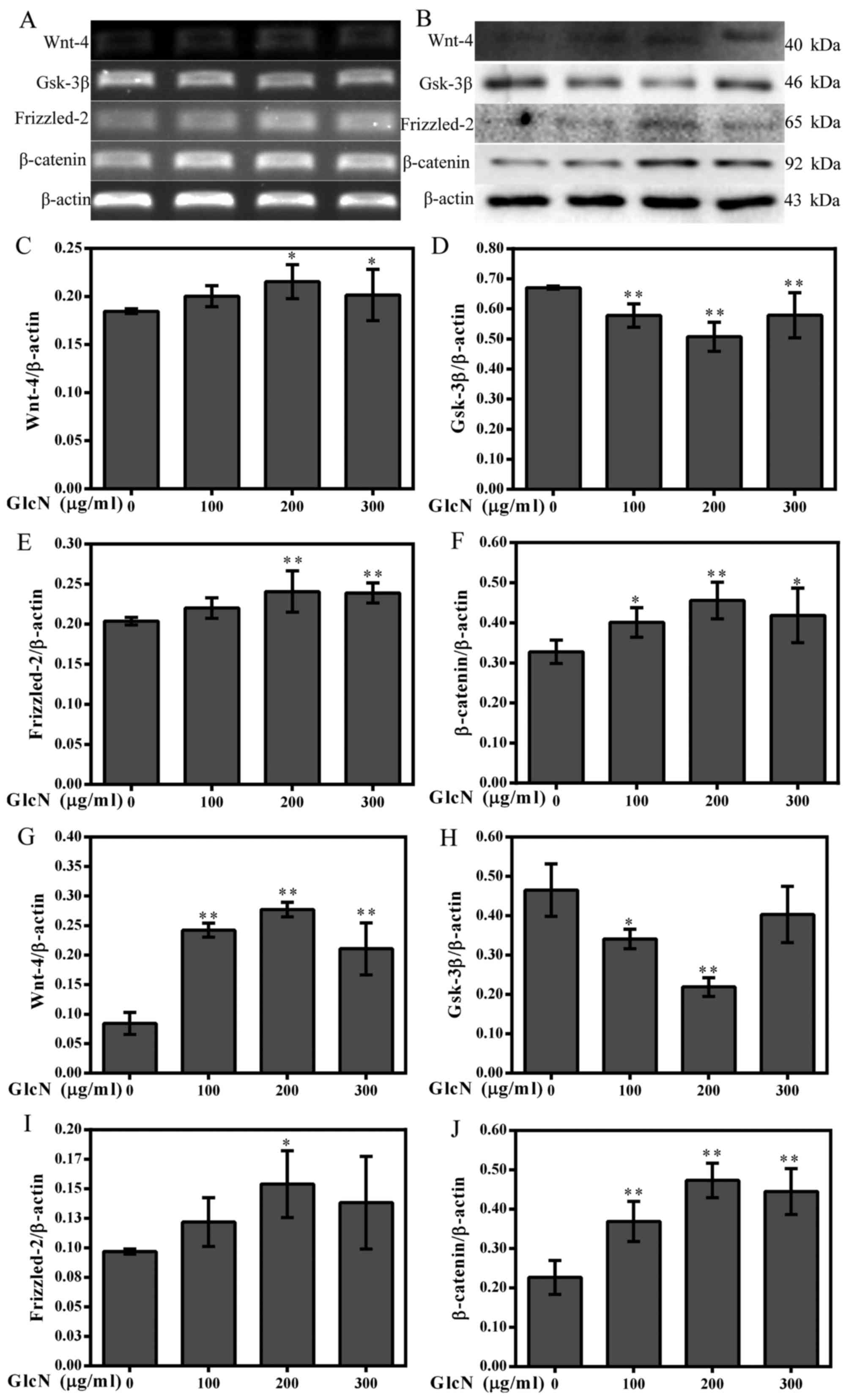 | Figure 5GlcN increases the expression of
Wnt-4, Frizzled-2 and β-catenin, and decreases the expression of
GSK-3β. (A) mRNA expression levels of Wnt-4, Frizzled-2, β-catenin
and GSK-3β were determined by reverse transcription-polymerase
chain reaction. (B) Western blot analysis was used to detect the
protein expression levels of Wnt-4, Frizzled-2, β-catenin and
GSK-3β. β-actin was used as an internal control. (C–F)
Semi-quantification of Wnt-4, Frizzled-2, β-catenin and GSK-3β mRNA
expression. (G-J) Semi-quantification of Wnt-4, Frizzled-2,
β-catenin and GSK-3β protein expression. *P<0.05,
**P<0.01 compared with untreated cells. GlcN,
glucosamine; GSK-3β, glycogen synthase kinase-3β. |
Immunofluorescence staining revealed that GlcN
markedly enhanced translocation of β-catenin into the nucleus
(Fig. 4). Following treatment
with GlcN, β-catenin staining was more intense and was localized in
the nucleus, whereas β-catenin was predominantly localized in the
cytoplasm in the untreated cell group. These results indicated that
GlcN may promote the expression and nuclear localization of
β-catenin.
GlcN increases Wnt-4 and Frizzled-2
expression, and decreases GSK-3β expression
β-catenin is a downstream intracellular signaling
molecule of the Wnt/β-catenin signaling pathway (24). The Wnt/β-catenin signaling pathway
inhibits phosphorylation of β-catenin, thus resulting in its
stabilization, cytoplasmic accumulation and subsequent nuclear
translocation. A previous study demonstrated that the Wnt/β-catenin
signaling pathway has an important function in controlling a wide
range of developmental processes, including cell proliferation
(24). Therefore, to detect
whether the effects of GlcN on chondrocyte proliferation are
associated with the Wnt/β-catenin signaling pathway, the mRNA and
protein expression levels of relevant factors in the Wnt/β-catenin
signaling pathway were detected. The results indicated that the
mRNA and protein expression levels of Wnt-4 and Frizzled-2 were
increased in the GlcN-treated chondrocytes compared with in
untreated cells (Fig. 4A–C, E, G and
I), whereas the mRNA and protein expression levels of GSK-3β
were reduced compared with in untreated cells (Fig. 4D and H). These results indicated
that the effects of GlcN on chondrocyte proliferation may be
associated with the Wnt/β-catenin signaling pathway.
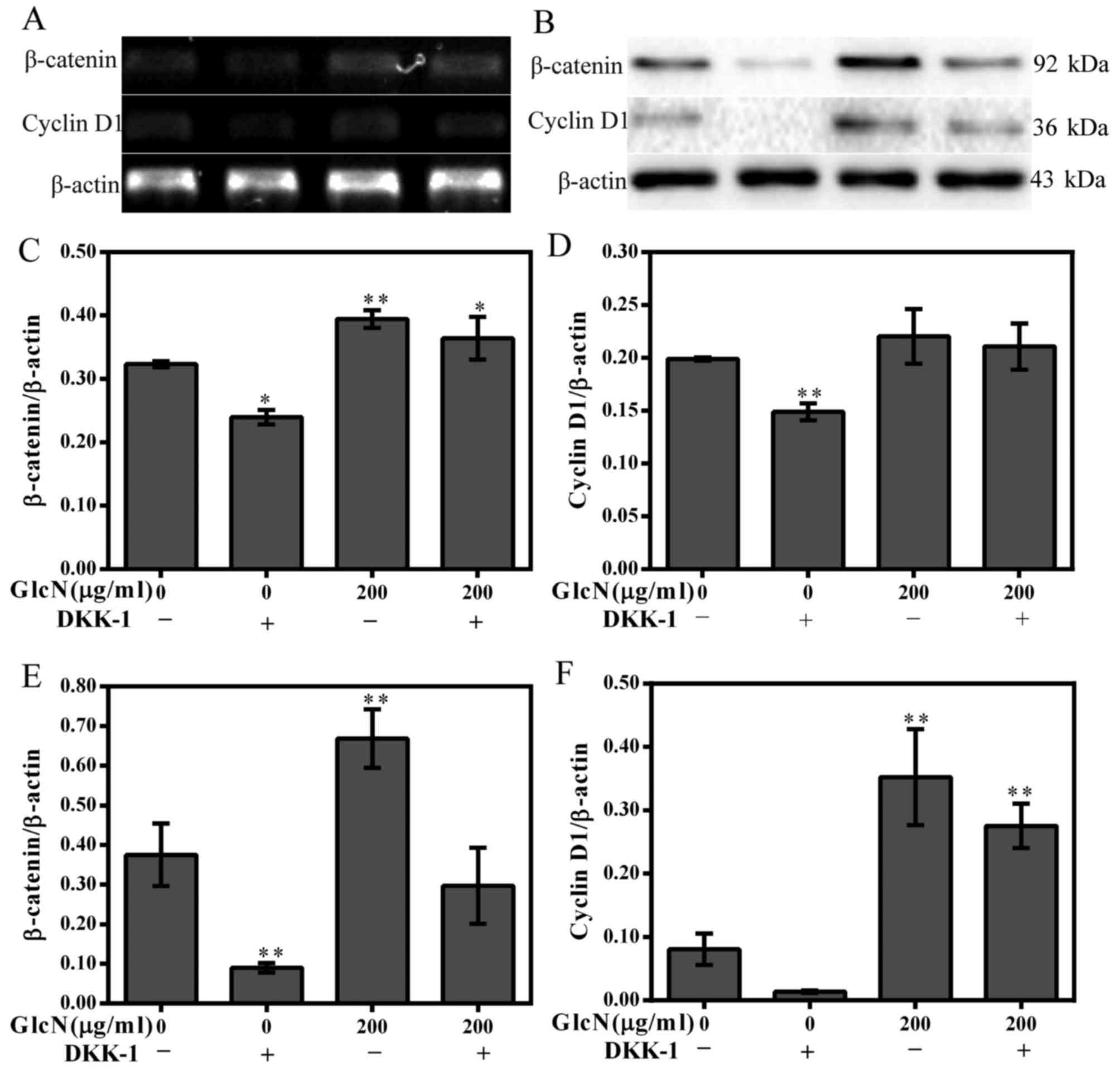
Cyclin D1 and β-catenin expression is
partly decreased following inhibition of the Wnt/β-catenin
signaling pathway
With regards to the aforementioned results of the
present study, it is unclear whether the Wnt/β-catenin signaling
pathway serves a crucial role in the regulation of chondrocyte
proliferation. Therefore, DKK-1 was used to block the Wnt/β-catenin
signaling pathway. The results revealed that the expression levels
of cyclin D1 and β-catenin were partly inhibited by DKK-1 (Fig. 6). These results further
strengthened the evidence that GlcN may participate in the
regulation of chondrocyte proliferation via the Wnt/β-catenin
signaling pathway.
Discussion
Functional alterations to chondrocytes have an
important role in cartilage damage, and the proliferation of
chondrocytes is required for the maintenance of cellular volume
(25). Numerous studies have
indicated that an association exists between cartilage degradation
and chondrocyte apoptosis (26,27). Therefore, promotion of chondrocyte
proliferation is essential and may be a potential therapeutic
strategy for the treatment of OA. In the present study, the results
demonstrated that GlcN promoted chondrocyte proliferation via the
Wnt/β-catenin signaling pathway. Firstly, the results indicated
that GlcN treatment promoted chondrocyte proliferation via the
promotion of cell cycle progression. The results of an MTT assay
revealed that GlcN promoted chondrocyte viability in a dose- and
time-dependent manner. Furthermore, the results of a flow
cytometric analysis demonstrated that the percentage of
chondrocytes in G0/G1 phase was markedly
decreased, whereas the percentage of chondrocytes in S phase was
significantly increased in cells treated with GlcN compared with in
untreated cells. The cell cycle is responsible for cell
proliferation; however, there are two restriction points,
G1/S and G2/S, within the cell cycle. The
G1/S restriction point begins during the initial
synthesis of DNA and the G2/S restriction point starts
at the beginning of mitosis. The G1/S restriction point
is more important since it determines which cell cycle phases the
cells pass through (6). The
present results indicated that GlcN treatment promoted chondrocyte
proliferation via the promotion of cell cycle progression.
To further determine the mechanism underlying the
effects of GlcN on chondrocyte proliferation, the protein
expression levels of cyclins and CDKs were detected. Cell cycle
progression is predominantly regulated via CDKs and cyclins, which
are two basic protein families of the cell cycle control system.
Various cyclin/CDKs exert their functions in different phases of
the cell cycle. In the cell cycle, cyclin D1 is the active
regulatory factor for the procession of G1/S phase, and
is important to cell proliferation (28). Cyclin D1 forms a complex with
CDKs, which proceed through the G1/S phase transition by
translocating to the nucleus and inducing cell signaling (29). It has been reported that cyclin
D/CDK4-6 and cyclin E/CDK2 allow progression in G1 and
elicit the G1/S transition (30). The present results suggested that
GlcN treatment may promote the progression of chondrocytes from
G1 to S phase by regulating cyclin D1, CDK4 and CDK6
expression. It has also been reported that GlcN increases cell
migration, cell cycle regulatory protein expression (cyclin D1,
CDK4, cyclin E and CDK2), and the percentage of S phase cells in
mouse embryonic stem cells (31).
In addition, GlcN and chondroitin sulfate association increases
tibial epiphyseal growth plate proliferation and bone formation in
ovariectomized rats (22).
It has previously been indicated that the expression
of cyclin D1 is directly regulated by numerous transcription
factors, including β-catenin (21,32). Through interaction with TCF/LEF
and coactivators, β-catenin is able to activate the downstream
target genes of cyclin D1, thus resulting in cell proliferation
(33). As the key effector of the
Wnt/β-catenin signaling pathway, β-catenin is responsible for
transducing the signal to the nucleus and initiating transcription
of Wnt-specific genes, which are responsible for the control of
cell fate decisions in several cells and tissues (34). Our previous study also indicated
that the Wnt/β-catenin signaling pathway mediates rat chondrocyte
proliferation (21). In the
present study, the results suggested that GlcN may increase the
expression levels of Wnt-4, Frizzled-2 and β-catenin, whereas it
may decrease GSK-3β expression. In addition, GlcN significantly
promoted the translocation of β-catenin into the nucleus, thus
suggesting that GlcN promotes chondrocyte proliferation via the
Wnt/β-catenin signaling pathway.
To confirm whether GlcN enhances chondrocyte
proliferation via the Wnt/β-catenin signaling pathway, DKK-1 was
used to inhibit the Wnt/β-catenin signaling pathway. DKK-1 binds to
LRP5/6 on target cells, thus resulting in the inhibition of
Wnt/β-catenin signaling (35,36). In the Wnt/β-catenin signaling
pathway, GSK-3β is thought to be phosphorylated in the absence of
Wnt signaling, consequently inducing the degrada tion of β-catenin
and decreasing cyclin D1 expression. The present results suggested
that the expression levels of β-catenin and cyclin D1 were partly
inhibited by DKK-1, further indicating that GlcN may promote
chondrocyte proliferation through the Wnt/β-catenin signaling
pathway. These results indicated GlcN may participate in the
regulation of chondrocyte proliferation via the Wnt/β-catenin
signaling pathway. In addition, it also demonstrated that
chondrocyte proliferation was not completely dependent on the
Wnt/β-catenin signaling pathway.
In conclusion, the present results demonstrated that
GlcN promotes the proliferation of chondrocytes via the
Wnt/β-catenin signaling pathway. However, it is undeniable that
other signal pathways may also be involved in this regulation.
Therefore, further studies are required to verify this conclusion
using cell and animal models.
Acknowledgments
The authors appreciate the research platform of
Fujian Key Laboratory of Rehabilitation Technology, Fuzhou, Fujian,
China.
References
|
1
|
Hada S, Kaneko H, Sadatsuki R, Liu L,
Futami I, Kinoshita M, Yusup A, Saita Y, Takazawa Y, Ikeda H, et
al: The degeneration and destruction of femoral articular cartilage
shows a greater degree of deterioration than that of the tibial and
patellar articular cartilage in early stage knee osteoarthritis: A
cross-sectional study. Osteoarthritis Cartilage. 22:1583–1589.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moo EK, Han SK, Federico S, Sibole SC,
Jinha A, Abu Osman NA, Pingguan-Murphy B and Herzog W:
Extracellular matrix integrity affects the mechanical behaviour of
in-situ chondrocytes under compression. J Biomech. 47:1004–1013.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gao Y, Liu S, Huang J, Guo W, Chen J,
Zhang L, Zhao B, Peng J, Wang A, Wang Y, et al: The ECM-cell
interaction of cartilage extracellular matrix on chondrocytes.
BioMed Res Int. 2014:6484592014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Onumah OE, Jules GE, Zhao Y, Zhou L, Yang
H and Guo Z: Overexpression of catalase delays G0/G1- to S-phase
transition during cell cycle progression in mouse aortic
endothelial cells. Free Radic Biol Med. 46:1658–1667. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lamb R, Lehn S, Rogerson L, Clarke RB and
Landberg G: Cell cycle regulators cyclin D1 and CDK4/6 have
estrogen receptor-dependent divergent functions in breast cancer
migration and stem cell-like activity. Cell Cycle. 12:2384–2394.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li X, Chen J, Liang W, Li H, Liu F, Weng
X, Lin P, Chen W, Zheng C, Xu H, et al: Bushen Zhuangjin Decoction
promotes chondrocyte proliferation by stimulating cell cycle
progression. Exp Ther Med. 9:839–844. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin AC, Seeto BL, Bartoszko JM, Khoury MA,
Whetstone H, Ho L, Hsu C, Ali SA and Alman BA: Modulating hedgehog
signaling can attenuate the severity of osteoarthritis. Nat Med.
15:1421–1425. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Appleton CT, Usmani SE, Mort JS and Beier
F: Rho/ROCK and MEK/ERK activation by transforming growth
factor-alpha induces articular cartilage degradation. Lab Invest.
90:20–30. 2010. View Article : Google Scholar
|
|
9
|
Yuasa T, Otani T, Koike T, Iwamoto M and
Enomoto-Iwamoto M: Wnt/beta-catenin signaling stimulates matrix
catabolic genes and activity in articular chondrocytes: Its
possible role in joint degeneration. Lab Invest. 88:264–274. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yasuhara R, Ohta Y, Yuasa T, Kondo N,
Hoang T, Addya S, Fortina P, Pacifici M, Iwamoto M and
Enomoto-Iwamoto M: Roles of β-catenin signaling in phenotypic
expression and proliferation of articular cartilage superficial
zone cells. Lab Invest. 91:1739–1752. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rosenbluh J, Wang X and Hahn WC: Genomic
insights into WNT/β-catenin signaling. Trends Pharmacol Sci.
35:103–109. 2014. View Article : Google Scholar
|
|
12
|
Wang Y, Li YP, Paulson C, Shao JZ, Zhang
X, Wu M and Chen W: Wnt and the Wnt signaling pathway in bone
development and disease. Front Biosci (Landmark Ed). 19:379–407.
2014. View Article : Google Scholar
|
|
13
|
Long F, Schipani E, Asahara H, Kronenberg
H and Montminy M: The CREB family of activators is required for
endochondral bone development. Development. 128:541–550.
2001.PubMed/NCBI
|
|
14
|
Miclea RL, Karperien M, Bosch CA, van der
Horst G, van der Valk MA, Kobayashi T, Kronenberg HM, Rawadi G,
Akçakaya P, Löwik CW, et al: Adenomatous polyposis coli-mediated
control of beta-catenin is essential for both chondrogenic and
osteogenic differentiation of skeletal precursors. BMC Dev Biol.
9:262009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nagaoka I, Igarashi M and Sakamoto K:
Biological activities of glucosamine and its related substances.
Adv Food Nutr Res. 65:337–352. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Reginster JY, Neuprez A, Lecart MP, Sarlet
N and Bruyere O: Role of glucosamine in the treatment for
osteoarthritis. Rheumatol Int. 32:2959–2967. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Salazar J, Bello L, Chávez M, Añez R,
Rojas J and Bermúdez V: Glucosamine for osteoarthritis: Biological
effects, clinical efficacy, and safety on glucose metabolism.
Arthritis (Egypt). 2014:4324632014.
|
|
18
|
Henrotin Y and Lambert C: Chondroitin and
glucosamine in the management of osteoarthritis: An update. Curr
Rheumatol Rep. 15:3612013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li H, Li X, Liu G, Chen J, Weng X, Liu F,
Xu H, Liu X and Ye H: Bauhinia championi (Benth.) Benth.
polysaccharides upregulate Wnt/β-catenin signaling in chondrocytes.
Int J Mol Med. 32:1329–1336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin P, Weng X, Liu F, Ma Y, Chen H, Shao
X, Zheng W, Liu X, Ye H and Li X: Bushen Zhuangjin decoction
inhibits TM-induced chondrocyte apoptosis mediated by endoplasmic
reticulum stress. Int J Mol Med. 36:1519–1528. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Weng X, Lin P, Liu F, Chen J, Li H, Huang
L, Zhen C, Xu H, Liu X, Ye H, et al: Achyranthes bidentata
polysaccharides activate the Wnt/β-catenin signaling pathway to
promote chondrocyte proliferation. Int J Mol Med. 34:1045–1050.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu F, Li X, Cai L, Li H, Chen J, Wong X,
Xu H, Zheng C, Liu X and Ye H: Achyranthes bidentata
polysaccharides induce chondrocyte proliferation via the promotion
of the G1/S cell cycle transition. Mol Med Rep. 7:935–940. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shtutman M, Zhurinsky J, Simcha I,
Albanese C, D'Amico M, Pestell R and Ben-Ze'ev A: The cyclin D1
gene is a target of the beta-catenin/LEF-1 pathway. Proc Natl Acad
Sci USA. 96:5522–5527. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huelsken J and Birchmeier W: New aspects
of Wnt signaling pathways in higher vertebrates. Curr Opin Genet
Dev. 11:547–553. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chan BY, Fuller ES, Russell AK, Smith SM,
Smith MM, Jackson MT, Cake MA, Read RA, Bateman JF, Sambrook PN, et
al: Increased chondrocyte sclerostin may protect against cartilage
degradation in osteoarthritis. Osteoarthritis Cartilage.
19:874–885. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim HT, Lo MY and Pillarisetty R:
Chondrocyte apoptosis following intraarticular fracture in humans.
Osteoarthritis Cartilage. 10:747–749. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
D'Lima DD, Hashimoto S, Chen PC, Colwell
CW Jr and Lotz MK: Human chondrocyte apoptosis in response to
mechanical injury. Osteoarthritis Cartilage. 9:712–719. 2001.
View Article : Google Scholar
|
|
28
|
Löwenheim H, Reichl J, Winter H, Hahn H,
Simon C, Gültig K, Müller A, Zenner HP, Zimmermann U and Knipper M:
In vitro expansion of human nasoseptal chondrocytes reveals
distinct expression profiles of G1 cell cycle inhibitors for
replicative, quiescent, and senescent culture stages. Tissue Eng.
11:64–75. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sherr CJ: D-type cyclins. Trends Biochem
Sci. 20:187–190. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ghiani C and Gallo V: Inhibition of cyclin
E-cyclin-dependent kinase 2 complex formation and activity is
associated with cell cycle arrest and withdrawal in oligodendrocyte
progenitor cells. J Neurosci. 21:1274–1282. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jeon JH, Suh HN, Kim MO and Han HJ:
Glucosamine-induced reduction of integrin β4 and plectin complex
stimulates migration and proliferation in mouse embryonic stem
cells. Stem Cells Dev. 22:2975–2989. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yano F, Kugimiya F, Ohba S, Ikeda T,
Chikuda H, Ogasawara T, Ogata N, Takato T, Nakamura K, Kawaguchi H,
et al: The canonical Wnt signaling pathway promotes chondrocyte
differentiation in a Sox9-dependent manner. Biochem Biophys Res
Commun. 333:1300–1308. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kikuchi A: Regulation of beta-catenin
signaling in the Wnt pathway. Biochem Biophys Res Commun.
268:243–248. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Valenta T, Hausmann G and Basler K: The
many faces and functions of β-catenin. EMBO J. 31:2714–2736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ye S, Wang J, Yang S, Xu W, Xie M, Han K,
Zhang B and Wu Z: Specific inhibitory protein Dkk-1 blocking
Wnt/β-catenin signaling pathway improve protectives effect on the
extracellular matrix. J Huazhong Univ Sci Technolog Med Sci.
31:657–662. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gifre L, Ruiz-Gaspà S, Monegal A, Nomdedeu
B, Filella X, Guañabens N and Peris P: Effect of glucocorticoid
treatment on Wnt signalling antagonists (sclerostin and Dkk-1) and
their relationship with bone turnover. Bone. 57:272–276. 2013.
View Article : Google Scholar : PubMed/NCBI
|