Introduction
Glucocorticoids (GCs), also known as steroids, are
potent immunity regulators widely used in the treatment of
inflammatory diseases. However, despite their effectiveness, GC
usage is strictly controlled due to severe side-effects, such as
osteoporosis, diabetes, impaired immune systems and Cushing's
syndrome (1). Although numerous
alternative drugs have been developed, the use of GCs has not
decreased due to their effectiveness. GC long-term treatment has
been reported as a cause of secondary osteoporosis (2). Diseases that may be the cause of
secondary osteoporosis include malabsorption syndrome (3), hypogonadism (4) and Cushing's disease induced by the
long-term use of GCs, unlike primary osteoporosis, which is caused
by aging and menopause (5).
Factors that cause secondary osteoporosis include drugs, such as
anti-epileptic drugs (6) and
antidepressants (7); among
these, GCs are the most representative. The long-term use of GC
results in rapid bone loss, and patients who undergo long-term GC
treatment are at a higher a risk of bone fractures (8).
GC resistance is a condition that causes an increase
in GC levels in the body and eventually causes side-effects
(9). Therefore, controlling GC
resistance is a crucial step to overcoming the side-effects. The GC
receptor (GR) is known to bind GCs to regulate immune responses and
to control cytokines through NF-κB transcription (9,10). GC resistance can reduce the
affinity of the GR and may negatively affect its binding ability
(9). However, the exact
mechanisms involved remain unclear. Thus, further research on the
mechanisms of GC resistance is required in order to understand its
occurrence and prevent GC-induced osteoporosis (GIOP), as well as
the side-effects caused by GC resistance.
Lycii radicis cortex (LRC) has been used to
regulate high blood pressure, body temperature, pain and bone
disorders in East Asia (11,12). LRC has been shown to be involved
in bone metabolism; LRC has been shown to inhibit osteoclast
differentiation and to exert anti-osteoporotic effects in
ovariectomized rats (13). LRC
has also been shown to promote osteoblast differentiation (14). Kukoamine B, a component of LRC,
has been shown to exert anti-osteoporotic effects through the
control of osteoblasts and osteoclasts (15). However, its association with the
effects of GC-induced osteoporosis has not yet been studied. In
addition, the regulatory effects of LRC on GC-GR resistance have
not yet been demonstrated, at least to the best of our
knowledge.
The results demonstrated in the present study
confirmed that osteoporosis was not induced in the GOIP model
without allergic contact dermatitis (ACD) treatment. These results
differ from those of previously published studies demonstrating
that osteoporosis occurred in the group treated with only GC
(16,17). Therefore, the present study
evaluated the effects of LRC in animal models with underlying
disease. To the best of our knowledge, research methods to
elucidate the mechanisms responsible for the side-effects of GCs
have not been developed sufficiently to date. Therefore, the
present study aimed to establish the research methodology for the
evaluation of the effects of Korean drugs on bone loss induced by
GCs used for ACD. The present study investigated the regulatory
effects of LRC on the expression of GR in mouse macrophages, and in
GC-induced bone loss in vivo and in vitro. The
results suggest the potential for a new animal model of GIOP, and
demonstrate the potential use of LRC as an alternative treatment
for secondary osteoporosis.
Materials and methods
Reagents and antibodies
Dexamethasone (DEX;
C22H29FO5; molecular weight,
392.46 g/mol; cat. no. D4902), a representative GC,
lipopolysaccharide (LPS), scopoletin and 2,4-dinitrochlorobenzene
(DNCB) were obtained from Sigma-Aldrich (Merck KGaA). Institute of
Cancer Research (ICR) mice were supplied by the Animal Center of
NARA Biotech. Phosphorylated (p)-NF-κB (cat. no. 3033) and p-GR
antibodies (cat. no. 4161S) were purchased from Cell Signaling
Technology, Inc. Bone morphogenetic protein 2 (BMP-2, cat. no.
ab14933) and Runt-related transcription factor 2 (Runx2, cat. no.
ab76956) antibodies were purchased from Abcam. Actin (cat. no.
sc-8432) and Lamin B antibody (cat. no. sc-374015) was obtained
from Santa Cruz Biotechnology, Inc., calbindin-D28k antibody (cat.
no. PA1-931) was obtained from Thermo Fisher Scientific, Inc., and
HRP-conjugated IgG antibody (cat. no. 115-035-062) was obtained
from Jackson ImmunoResearch Laboratories, Inc. FBS,
penicillin/streptomycin and PBS were provided by Invitrogen; Thermo
Fisher Scientific, Inc. CellTiter 96® AQueous
Non-Radioactive Cell Proliferation assay was obtained from Promega
Corporation. Reverse transcription PCR (RT-PCR) primers were
purchased from GenoTech Corp.
Preparation of LRC
LRC was obtained from Kyung Hee University Medical
Center. LRC was boiled for 2 h with water at a 1:1 ratio. The
filtered extract was freeze-dried using a lyophilizer. The yield
was 6% (dried powder, 30 g). A voucher specimen (KH076) was
deposited at the Herbarium of the Department of Anatomy at Kyung
Hee University. The dried extract was dissolved in distilled
deionized water, filtered through a 0.22-µm syringe filter
system (RephiLe Bioscience) and stored at -20°C until use.
Establishment of the animal model
All animal experiments were approved by the Kyung
Hee University Animal Care and Use Committee [approval no.
KHUASP(SE)-17-082]. A total of 32 male ICR mice (10 weeks old;
weighing 35g) were bred in a controlled room (22±2°C temperature,
50±10% humidity, 12-h light/dark cycle).
The experimental conditions were as follows: i) Mice
in the normal condition treated with PBS and olive oil (9:1) only
(Fig. 1A, n=8); ii) the ACD +
topically applied (TA) GC condition, where DEX was TA following the
induction of ACD (Fig. 1B, n=8);
iii) the ACD + subcutaneously (SC) injected GC condition, where DEX
was SC injected following the induction of ACD (Fig. 1C, n=8); and iv) the GC (SC)
condition, where DEX was SC injected without the induction of ACD
(Fig. 1D, n=8).
 | Figure 1Establishment of an experimental
animal model of GIOP. (A) Mice in the normal condition treated with
PBS and olive oil (9:1) only. (B) ACD + GC (TA) group, where DEX
was topically treated following the induction of ACD. (C) ACD + GC
(SC) group, where DEX was injected for treatment under the skin
following the induction of ACD. (D) GC (SC) group, where DEX was
injected without the induction of ACD. DEX was subdivided into the
subcutaneous injection GC (SC) groups without causing underlying
diseases. ACD, allergic contact dermatitis; GC, glucocorticoid;
GIOP, GC-induced osteoporosis; TA, topically applied; SC,
subcutaneously injected; DEX, dexamethasone; DNCB,
2,4-dinitrochlorobenzene. |
For the induction of ACD, the backs of the mice were
challenged with 200 µl 1% DNCB for 3 days. After 4 days,
0.5% DNCB was applied to the backs of the mice 3 days per week for
8 weeks to induce ACD. The GC (SC) group was not challenged with
DNCB. In the ACD + GC (TA) group, 2 mg/kg DEX in 200 µl
PBS/olive oil (9:1) were applied to the dorsal skin of the mice 3
days per week for the induction of GC-related side-effects. In the
ACD + GC (SC) and GC (SC) groups, 2 mg/kg DEX in 1% ethanol in
saline were injected 3 days per week. The following
conditions/observations were considered humane end points: i)
Difficultly in ingesting food or water due to discomfort in
walking; ii) the mouse became unconscious or did not react to
external stimuli; iii) a weight loss ≥20% compared with other mice
of the same age; iv) severe infection, laceration and bleeding at
the stimulation site; and v) difficulty in maintaining a normal
posture due to weak energy. During the process of the animal
experiment, no mice died or exhibited signs which would lead to
humanitarian termination. At the end of the animal experiments, the
mice were sacrificed by cervical dislocation after collecting 2 ml
blood under deep anesthesia with 5% isoflurane diluted in
O2. The femur and dorsal skin tissues were collected,
and the muscles around the femur were neatly removed.
Design of experiments to evaluate the
effects of LRC on GIOP models
Following the establishment of the GIOP animal
model, to determine the effect of LRC on bone loss inhibition in
GIOP, a model of TA-induced osteoporosis under the ACD condition
was used in 32 new ICR mice (10 weeks old; weighing 35 g, for 56
days). The LRC low dose was considered to be 50 mg/ml, and the high
dose was considered to be 100 mg/ml. The mice were then divided
into 4 groups (n=8/group) as follows: The normal group without DNCB
and DEX stimulation and PBS/olive oil (9:1)-treated group (Nor),
DNCB-sensitized and DEX-treated group (DEX), LRC 50 mg/kg-treated
group (LRC_Low) and the LRC 100 mg/kg-treated group (LRC_High)
(Fig. 2).
Histological analysis and
immunohistochemistry (IHC)
The skin and femur tissues of the mice were embedded
in 10% neutral buffered formalin for 24 h. Micro-computed
tomography (CT) analysis was conducted according to Kim et
al (18). Briefly, the left
femurs were analyzed using a cone beam micro-CT system
(SkyScan1176; Skyscan; Kontich). The femurs were scanned from the
knee for 500 frames, and the images were visualized using Data
Viewer software (v.1.5.1.9; Skyscan; Kontich). Structural
parameters were determined using CT analyzer software (v1.15.4.0;
Skyscan; Kontich). Bone morphometric parameters, such as bone
mineral density (BMD, g/cm3), bone volume/total volume
(BV/TV, %), trabecular thickness (Tb.Th, mm), trabecular separation
(Tb.Sp, mm), trabecular number (Tb.N, 1/mm) and bone surface
density (BS/TV, 1/U) were measured.
Tissue staining was performed according to a
previous study (19). In brief,
paraffin-embedded tissue sections were prepared for IHC by
incubation with 0.3% H2O2 in methanol for 30
min. The tissues were then placed in proteinase K buffer
(Sigma-Aldrich; Merck KGaA) for 20 min. Tissues were incubated at
room temperature for 1 h in blocking solution (10% normal serum,
Gibco; Thermo Fisher Scientific, Inc.). The tissues were treated
anti-calbindin-D28k antibody (1:100) overnight at 4°C, and then
incubated at the room temperature for 1 h with peroxidase-labelled
IgG (1:100; rabbit; cat. no. BA-1000, Vector Laboratories, Inc.)
for 1 h. Tissue sections were then stained using the Vector ABC kit
(Vector Laboratories, Inc.) as recommended by the manufacturer, and
then incubated at room temperature for 7 min with diaminobenzidine
(DAB) tetrachloride mixed with NiCl2 + H2O
(Vector Laboratories, Inc.). Finally, the tissues were
counter-stained using hematoxylin. The femur tissues were
decalcified in EDTA-2Na for 3 weeks and then embedded in paraffin.
The femur tissue was fixed using 10% neutral buffered formalin and
the tissues were embedded with paraffin. The embedded tissues were
cut into 5-µm-thick sections and stained with hematoxylin
and eosin (H&E, Sigma-Aldrich, Merck KGaA) at room temperature
for 10 and 10 min, respectively. The number of mast cells was
counted following toluidine blue staining (Sigma-Aldrich, Merck
KGaA) at room temperature for 2 min. All sections were examined
under a light microscope (DP73; Olympus Corporation).
ELISA
Serum was obtained by centrifuging the collected
blood at 14,310 x g for 20 min at 4°C. Measurement of the
expression of bone-related markers, such as receptor activation of
NF-κB ligand (RANKL), osteoprotegerin (OPG), sclerostin (SOST),
Dickkopf-1 (DKK1) and bone-derived fibroblast growth factor 23
(FGF23) in serum was performed by NEW Korea Industrial Co., Ltd.
and was detected using Multiplex ELISA.
Cell cytotoxicity assay
Murine RAW 264.7 cells (Korean Cell Line Bank;
Korean Cell Line Research Foundation) were cultured in DMEM
(Welgene, Inc.) containing 10% FBS and 1% penicillin/streptomycin.
Osteoblastic MC3T3-E1 cells were cultured in α-MEM containing 10%
FBS and 1% penicillin/streptomycin (Gibco; Thermo Fisher
Scientific, Inc.). The cell cultures were maintained at 37°C in a
humidified atmosphere of 5% CO2 in air. The cells were
seeded in a 96-well plate. The cells were subjected to experimental
methods according to a previous study (13). Cells were treated only with 10,
50 and 100 µg/ml LRC or with 1 µM DEX for 24 h. An
MTS solution was added at 20 µl per well. After 2 h, cell
viability was measured at 490 nm using an ELISA reader (Molecular
Devices, LLC). The optical density (OD) values of untreated cells
were normalized to 100%. MC3T3-E1 cells were seeded into 96-well
plates and incubated in a humidified incubator at 37°C overnight.
Following 48 h of culture, Cell Counting Kit-8 (CCK-8; Dojindo
Molecular Technologies, Inc.) assay was performed according to the
manufacturer's instructions. The medium in the 96-well plates was
exchanged for 100 µl 20% CCK-8 solution. After incubating
the cells for 1 h at 37°C, the absorbance at a wavelength of 450 nm
was measured with a microplate reader (Versamax; Molecular Devices,
LLC). The percentage of cell viability was calculated based on the
control wells. Lactate dehydrogenase (LDH) release was measured
using a LDH assay kit (Dojindo Molecular Technologies, Inc.)
according to the manufacturer's instructions.
Semi-quantitative RT-PCR
RT-PCR was conducted to a previous study (20). MC3T3-E1 cells were seeded at a
density of 1x106 cells/well in a 6-well plate for 24 h.
The cells were then treated with 10, 50 and 100 µg/ml LRC
and co-treated with DEX (1 µM) for either 2 or 4 days. The
medium was changed every 2 days. Total RNA was extracted using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.).
cDNA was prepared using SuperScript II reverse transcriptase
(Invitrogen; Thermo Fisher Scientific, Inc.) and cDNA was then
amplified in a PCR machine (C1000 Touch™ Thermal Cycler, Bio-Rad
Laboratories, Inc.) using Taq polymerase and each primer. GAPDH was
used as the reference gene. The list of primers used is presented
in Table I. These PCR reactants
was separated in an 2% agarose gel and visualized using NαBI
(Neoscience). mRNA expression was measured using ImageJ software
(v1.52a; National Institutes of Health).
 | Table IList of primers used for RT-PCR. |
Table I
List of primers used for RT-PCR.
| Marker name | Gene name | Accession no. | Species | Sequence |
|---|
| RUNX2 | Runx2 | NM_009820.5 | Mouse | F:
CCAACCGAGTCATTTAAGGCT |
| | | | R:
GCTCACGTCGCTCATCTTG |
| Osteocalcin
(OCN) | Bglap2 | NM_001032298.3 | Mouse | F: GCA ATA AGG TAG
TGA ACA GAC TCC |
| | | | R: GTT TGT AGG CGG
TCT TCA AGC |
| ALP | Alpl | NM_001287172.1 | Mouse | F:
CGGGACTGGTACTCGGATAA |
| | | | R:
TGAGATCCAGGCCATCTAGC |
| Osteonectin
(OSN) | Sparc | NM_001290817.1 | Mouse | F:
AAACATGGCAAGGTGTGTGA |
| | | | R:
TGCATGGTCCGATGTAGTC |
| Osteopontin
(OPN) | Spp1 | NM_001290377.1 | Mouse | F:
TCTGATGAGACCGTCACTGC |
| | | | R:
AGGTCCTCATCTGTGGCATC |
| Procol (COL1) | Col1a1 | NM_007742.4 | Mouse | F:
GCTCCTCTTAGGGGCCACT |
| | | | R:
CCACGTCTCACCATTGGGG |
| GAPDH | Gapdh | NM_008084.3 | Mouse | F:
ACTTTGTCAAGCTCATTTCC |
| | | | R:
TGCAGCGAACTTTATTGATG |
| Caspase-6 | Casp6 | NM_009811.4 | Mouse | F:
AGACAAGCTGGACAACGTGACC |
| | | | R:
CCAGGAGCCATTCACAGTTTCT |
| Caspase-9 | Casp9 | NM_001277932.1 | Mouse | F:
GGATCCATGCCCAGACCAGTGGACATTGG |
| | | | R:
GGTCTAGATTATGATGTTTTAAAGAAAAG |
| IAP-1 | Birc3 | NM_007464.3 | Mouse | F:
AAGTGTGTATGGACCGAGAG |
| | | | R:
GGACCATTAGTCTTGTTCAG |
| IAP-2 | Birc2 | NM_001291503.1 | Mouse | F:
TGTGTATGGACAGAGAGGTT |
| | | | R:
CAGCTTCTGATGTCCAACAA |
| X-IAP | Xiap | NM_001301641.1 | Mouse | F:
ATACGGAGGATGAGTCAAGT |
| | | | R:
GGTTGAACGTAATGACGGTG |
Alizarin Red S staining
Alizarin red S staining was used in the
mineralization assay of MC3T3-E1. Osteoblast differentiation and
Alizarin Red S staining were conducted as described in the study by
Kim et al (20). Briefly,
MC3T3-E1 cells were incubated in osteogenic differentiation medium
(α-MEM containing 10 mM β-glycerophosphate and 25 µg/ml
ascorbic acid) at a density of 5x105 cells/well in a
24-well plate for 24 h. MC3T3-E1 cells were treated with 10, 50 and
100 µg/ml LRC and co-treated with DEX (1 µM) for 4
weeks. The cells were fixed with 80% ice-cold ethanol for 1 h and
stained with 1 w/v (%) Alizarin Red S solution (Daejung Chemicals
& Metals Co., Ltd.) at the room temperature for 5 min. The
wells were destained with 500 µl 10% cetylpyridinium
chloride for 15 min. The OD value of the Alizarin Red S-stained
cells was measured at 570 nm using a spectrophotometer (Versamax;
Molecular Devices, LLC).
Western blot analysis
To confirm the effect of LRC on osteogenesis-related
markers, MC3T3-E1 cells were seeded at a density of
1x106 cells/well in a 6-well plate in a humidified
incubator at 37°C for 24 h. MC3T3-E1 cells were treated with 10, 50
and 100 µg/ml LRC and co-treated with DEX (1 µM) for
2 days. Whole protein lysate was extracted using RIPA buffer (50 mM
Tris-Cl, 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate and 0.1%
SDS with protease inhibitor and phosphatase inhibitor 2 and 3
cocktails). To confirm the inhibitory effect of LRC on GC-induced
resistance, RAW 264.7 cells were seeded at a density of
1x106 cells/well in a 6-well plate in a humidified
incubator at 37°C for 2 days. Subsequently, RAW 264.7 cells were
cultured with 10 and 100 µg/ml LRC or 0.1 and 1 µM
DEX + 1 µg/ml LPS for 2 or 24 h. The cells from each well
were lysed to extract nuclear proteins, which were purified using a
NE-PER Nuclear and Cytoplasmic Extraction reagent kit (Pierce;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. The extracted protein was quantified through
bicinchoninic acid assay. Western blot analysis was conducted
according to a previous study (13). Briefly summarized, the same
amount of protein (30 µg) was separated by protein size
through electrophoresis on 10% SDS-gel (100 V, 1.5 h), and then
transferred to nitrocellulose membranes (Whatman plc; Cytiva). To
prevent non-specific protein binding, the membrane was reacted with
5% skim milk in 0.05% TBST at the room temperature for 1 h, and
each primary antibody was attached at 4°C overnight. The dilution
concentration of each antibody was as follows: BMP-2, 1:500; Runx2,
1:1,000; actin, 1:500; p65, 1:1,000; p-GR (ser211), 1:500; p-GR
(ser226) and Lamin B, 1:500. The secondary antibodies (1:10,000;
source: Rabbit; cat. no. 111-035-045; 1:10,000; source: Mouse; cat.
no. 115-035-062; Jackson ImmunoResearch Laboratories, Inc.) were
then reacted at room temperature for 1 h and visualized using
enhanced chemiluminescence (Santa Cruz Biotechnology, Inc.).
Protein expression was measured using ImageJ software (v1.52a;
National Institutes of Health). The expression of each target band
was quantified using Actin and Lamin B.
Gas chromatography (GC)/mass spectrometry
(MS) analysis of LRC
The phytochemical analysis of LRC was performed with
an Agilent 6890 GC apparatus interfaced to an Agilent 5973 MS
system equipped with an electron ionization (EI) source and
autoinjector (Agilent Technologies, Inc.). GC/MS was conducted
according to a previous study (21). The GC system was built with a
DB-5 column (30.0 m x 0.25 mm x 0.25 µm). The temperature
varied from 80°C (5 min) to 200°C at 5°C/min, and the injection
volume was 2 µl. Injection was performed in the split mode
adjusted to 1:5. The carrier gas was helium at 1.0 ml/min. The
inlet, source and quadrupole temperatures were set at 290, 230 and
190°C, respectively. For MS detection, the EI mode with an
ionization energy of 70 eV was used, with a mass range at m/z
50-800. Agilent ChemStation software (Agilent Technologies, Inc.)
was used for data processing. The phytochemical compounds were
identified by mass fragmentation patterns compared via Wiley
Spectral Library search.
Statistical analysis
The data are presented as the mean ± SEM. All
experiments were repeated at least 3 times. All data were analyzed
using GraphPad Prism software (v5.01; GraphPad Software, Inc.).
One-way ANOVA was used to evaluate the treatment effect, a single
group was post-tested using Dunnett's, and multiple comparisons
between different groups were post-tested using Tukey's. P<0.05
was considered to indicate a statistically significant
difference.
Results
Evaluation of the established GIOP
models
The present study aimed to model osteoporosis
induction following the systemic injection or skin application of
GC following the development of a specific inflammatory disease,
namely ACD. Through the analysis of bone density through micro-CT
and histological examination, GIOP found to be induced by the DEX
dorsal skin application following the induction of ACD (Fig. 3A-C). In addition, as a result of
bone microstructure analysis, BMD was significantly decreased in
both the ACD + GC (TA) and ACD + GC (SC) groups, and the ACD + GC
(TA) group exhibited significant differences in Tb.Sp, Tb.Th and
BV/TV compared with those of the Nor group. However, the ACD + GC
(SC) group did not exhibit a significant difference in Tb.Sp or
BV/TV (Fig. 3D-G).
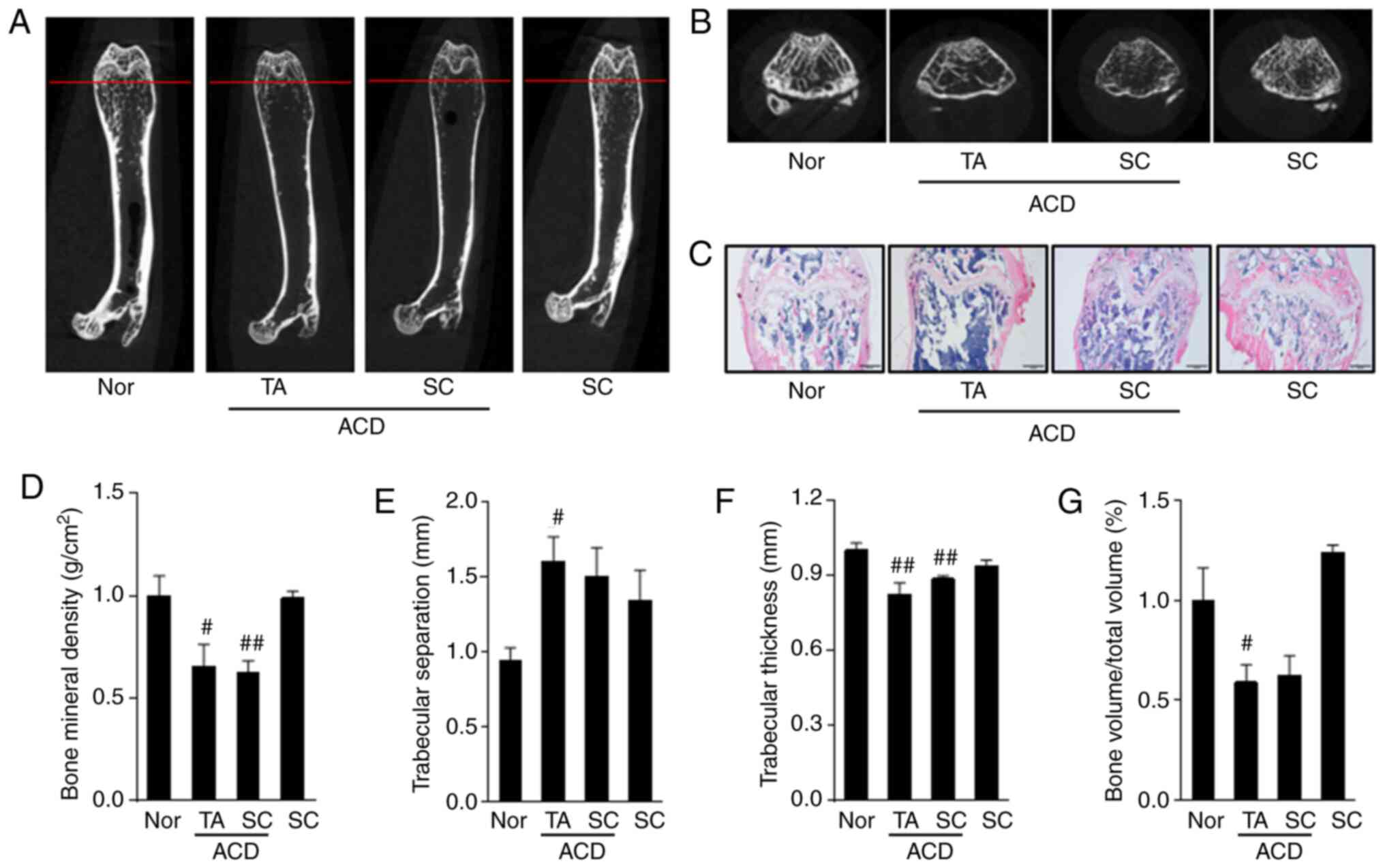 | Figure 3Bone structure analysis of mice femur
via micro-computed tomography. (A) Horizontal cross-section of a
mouse femur. (B) Vertical cross-section corresponding to the red
line shown in (A). (C) Paraffin-embedded section of the mouse
tissues stained with hematoxylin and eosin. (D) Bone mineral
density, (E) trabecular separation, (F) trabecular thickness and
(G) bone volume/total volume results for the different experimental
condition. In the ACD + GC (TA) model, GIO was induced by applying
DEX to the back of the animals following the induction of ACD with
DNCB. In the ACD + GC (SC) group, DNCB induced ACD and then DEX was
subcutaneously injected to induce GIOP. In the GC (SC) group, DEX
was injected subcutaneously without causing ACD. #P<0.05,
##P<0.01 vs. normal animals. Nor, normal group; ACD,
allergic contact dermatitis; GC, glucocorticoid; GIOP, GC-induced
osteoporosis; TA, topically applied; SC, subcutaneously injected;
DEX, dexamethasone; DNCB, 2,4-dinitrochlorobenzene. |
Expression of calbindin-D28k in the
duodenum and kidney of the animals in the experimental model
To evaluate the calcium absorption abilities of
digestive organs, the expression of a calcium binding protein
(calbindin-D28k) in the duodenum and kidney was evaluated (Fig. 4A and C). The expression of
calbindin-D28k was significantly decreased in the ACD + GC (TA)
group in both the duodenum and kidney. The expression of
calbindin-D28k was also inhibited in the duodenum in the ACD + GC
(SC) group. However, the expression of calbindin-D28k in the kidney
of the ACD + GC (SC) group appeared to be decreased compared to
that of the normal group; however, the difference was not
statistically significant (Fig. 4B
and D). On the whole, the results presented in Figs. 3 and 4 demonstrated that the induction of ACD
was a prerequisite for the occurrence of GIOP. However, GIOP did
not occur in animals without ACD. In addition, it was demonstrated
that the TA group exhibited osteoporosis more markedly than the SC
group, as reflected by bone microstructure or the expression of
calbindin-D28k in the duodenum. Finally, an animal model was
established by the TA of DEX under ACD inducing conditions.
Effect of LRC on bone loss in GIOP mouse
femur
The present study attempted to compare the effects
of LRC using the DEX administration method, which was applied to
the ACD + GC (TA) group. In both the micro-CT and histological
examination experiments, LRC exerted an inhibitory effect on
GC-induced osteoporosis (Fig. 5A and
B). The results of bone microstructure analysis revealed that
the LRC high group exhibited a significantly increased (BV/TV, Tb.N
and BS/TV) or decreased (Tb.Sp) compared with the GC group
(Fig. 5C-G). Tb.Th exhibited a
tendency to increase in the LRC low and LRC high groups compared
with the GC group, although the difference was not statistically
significant (Fig. 5H).
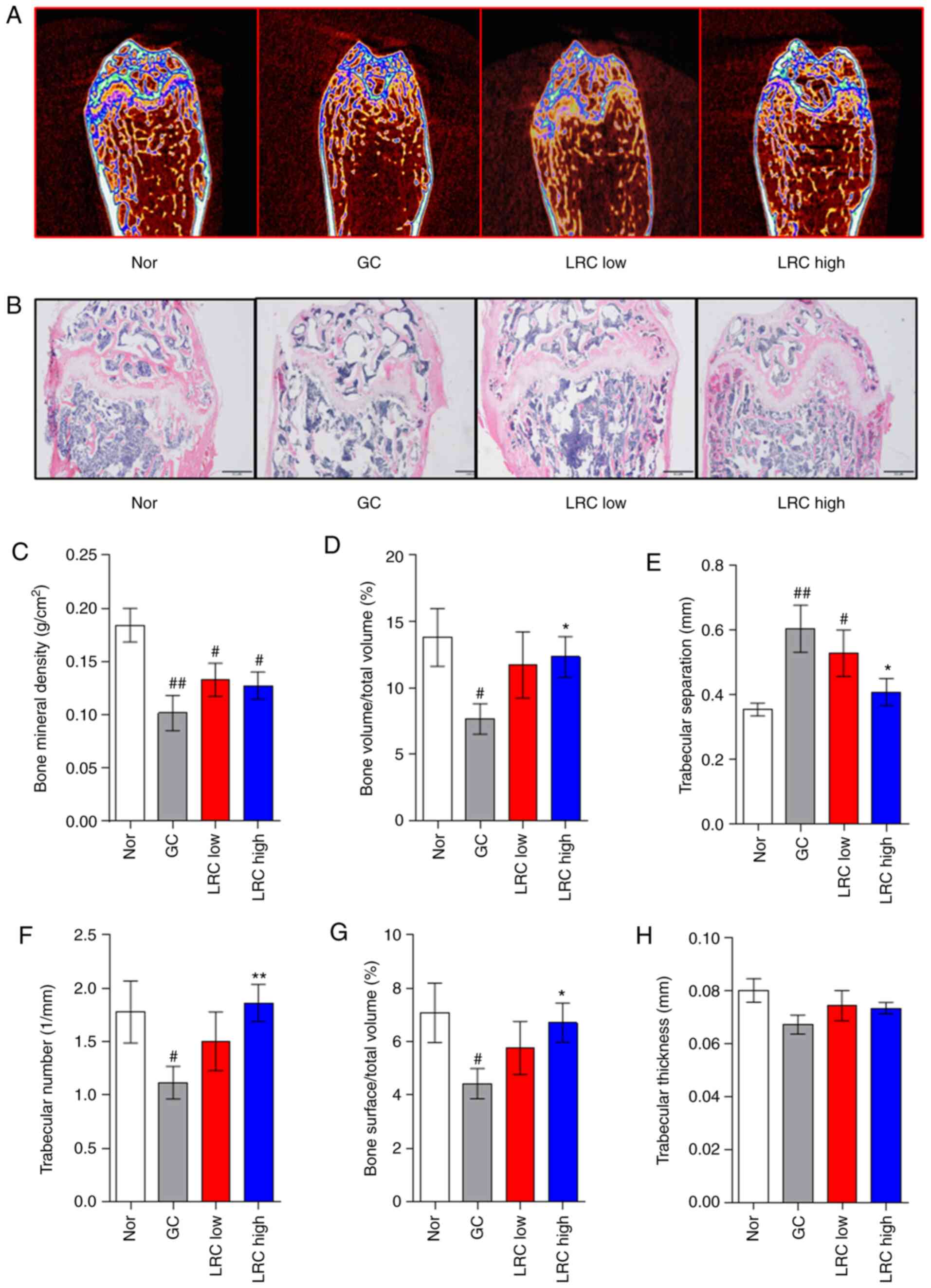 | Figure 5Effect of LRC on GIOP in an ACD mouse
model. DEX (2 mg/kg) was applied for 8 weeks following the
induction of ACD using 2,4-dinitrochlorobenzene on the skins of
mice. LRC extract (50 and 100 mg/kg) or distilled water were orally
administered to the mice. (A) After 8 weeks, the mouse femurs were
analyzed by micro-CT. (B) The femurs were sectioned and stained
with hematoxylin and eosin. The mouse bone parameters such as (C)
BMD, (D) BV/TV, (E) Tb.Sp, (F) Tb.N, (G) BS/TV and (H) Tb.Th were
analyzed by micro-CT. Data represent the means ± SEM of
experiments. #P<0.05, ##P<0.01 vs.
normal animals; *P<0.05, **P<0.01 vs.
GC animals. LRC, Lycii radicis cortex; ACD, allergic contact
dermatitis; GIOP, GC-induced osteoporosis; DEX, dexamethasone; CT,
computed tomography; GC, glucocorticoid; bone mineral density, BMD;
bone volume/total volume, BV/TV; trabecular separation, Tb.Sp;
trabecular number, Tb.N; trabecular thickness, Tb.Th; bone surface
density. |
Effect of LRC on calcium absorption in
duodenum and kidney tissues
The LRC high group exhibited significantly increased
duodenum calbindin-D28k levels (Fig.
6A and C). In the kidney tissue, calbindin-D28k expression was
upregulated by LRC, although the difference was not significant
(Fig. 6B and D).
Effect of LRC on DNCB-induced ACD
To investigate whether LRC maintains the
anti-inflammatory effects of DEX (22,23), the effects of LRC on immune cell
infiltration number and skin thickness were evaluated in an animal
model of ACD. The LRC high group exhibited a larger reduction in
the number of mast cells in DNCB-induced ACD skin tissues compared
with that of the GC group (Fig. 7A
and D). The LRC low group and LRC high group did not exhibit a
significant difference on the number of eosinophils compared with
the GC group (Fig. 7B and E).
The LRC high group experienced significantly increased epidermal
thickness compared with that of the GC (DEX-treated) group. These
results appear to be indicative of the protective effect of LRC
against skin atrophy, which is one of the side-effects of GC
(Fig. 7C and F). However, LRC
did not significantly affect the thickness of the dermis (Fig. 7G).
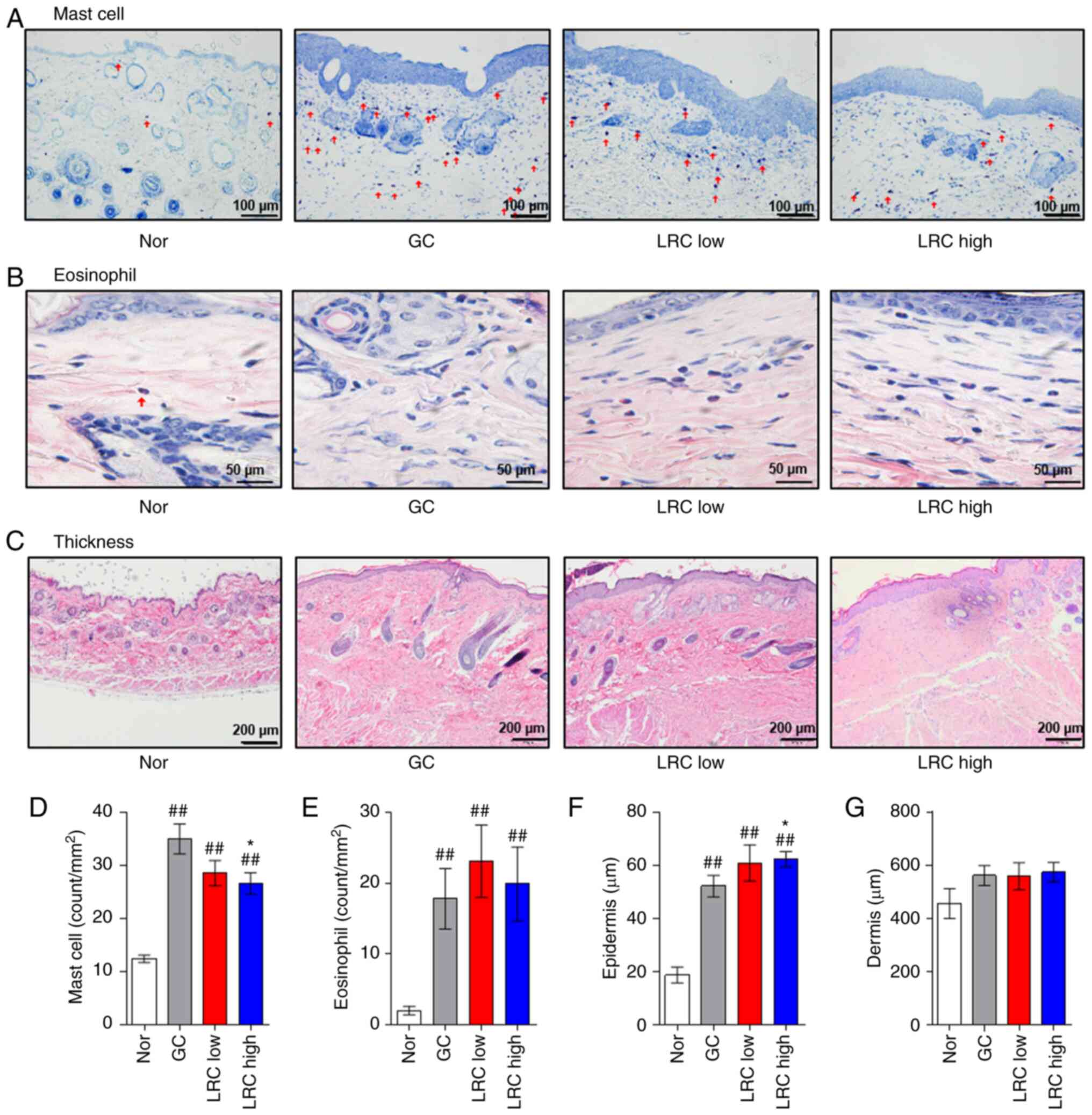 | Figure 7Effect of Lycii radicis cortex
on 2,4-dinitrochlorobenzene-induced allergic contact dermatitis
skin tissues. (A) Infiltration of mast cells was detected by
toluidine blue staining (magnification, x200; scale bar size, 100
µm). (B) Eosinophil infiltration was detected by H&E
staining (magnification, x400; scale bar size, 50 µm). (C)
The thickness of the epidermis and dermis was evaluated by H&E
staining of skin sections (magnification, x100; scale bar size, 200
µm). Measurement of the count of (D) mast cells and (E)
eosinophils. All measurements were performed in five fields in each
tissue. Measurement of the thickness of (F) epidermis and (G)
dermis. All measurements were performed in three fields in each
tissue. Data represent the mean ± SEM. ##P<0.01 vs.
normal animals; *P<0.05 vs. GC animals. H&E,
hematoxylin and eosin. LRC, Lycii Radicis Cortex; ACD, allergic
contact dermatitis; GC. glucocorticoids; GIOP, GC induced
osteoporosis; DEX, dexamethasone; H&E, hematoxylin and eosin
stain. |
Effect of LRC on osteoporosis-related
mediators in serum, and on body and organ weight
The results of ELISA revealed that GC significantly
increased the OPG and DKK1 levels, while the levels in the
LRC-treated groups were not affected compared with the GC group
(Table II). The GC group
exhibited significantly decreased body, liver, thymus and spleen
weight compared with the findings in the Nor group. LRC
significantly increased the testis weight compared with that of the
GC group. LRC increased liver weight compared with that of the GC
group, although not significantly (Table III).
| Table IIEffect of LRC on the
osteoporosis-related mediators in serum. |
Table II
Effect of LRC on the
osteoporosis-related mediators in serum.
| Group | No. of mice | RANKL (pg/ml) | OPG (ng/ml) | SOST (pg/ml) | DKK1 (ng/ml) | FGF23 (pg/ml) |
|---|
| Nor | 8 | 127.30±16.70 | 5.92±0.43 | 90.94±8.85 | 65.06±8.34 | 796.57±42.81 |
| GIOP | 8 | 101.27±29.39 | 10.96±1.16b | 257.24±84.09 | 99.97±7.28a |
1321.40±280.14b |
| LRC low | 8 | 41.82±6.01b | 9.26±1.08b |
133.97±16.18a | 77.16±7.28 |
1247.92±164.16a |
| LRC high | 8 | 46.82±8.53b | 9.66±1.10b | 150.52±32.90 |
102.13±13.36a |
1162.78±76.91b |
| Table IIIEffect of LRC on body and the organ
weight. |
Table III
Effect of LRC on body and the organ
weight.
| Group | No. of mice | Body weight
(g) | Liver (g) | Testis (g) | Thymus (g) | Spleen (g) | Kidney (g) |
|---|
| Nor | 8 | 39.54±0.78 | 1.44±0.04 | 0.24±0.01 | 0.040±0.003 | 0.13±0.01 | 0.61±0.02 |
| GIOP | 8 | 32.64±0.74b | 1.25±0.04b | 0.20±0.02 | 0.014±0.001b | 0.10±0.01a | 0.58±0.02 |
| LRC low | 8 | 32.75±0.79b | 1.32±0.08a | 0.24±0.01c | 0.012±0.001b | 0.07±0.01b | 0.59±0.02 |
| LRC high | 8 | 31.65±0.77b | 1.31±0.08a | 0.24±0.01c | 0.013±0.002b | 0.08±0.01b | 0.57±0.02 |
Effect of LRC on GC-induced osteoblastic
MC3T3-E1 cell apoptosis
After confirming the ameliorating effects of LRC on
GIOP in in vivo experiments, in order to analyze the
mechanisms involved, the effects of LRC on the DEX-induced
apoptosis and differentiation of osteoblasts were verified in
MC3T3-E1 cells, which are known as osteoblast progenitor cells
(24). DEX exhibited toxicity to
the MC3T3-E1 cells; however, the cell survival rate was increased
by LRC in a concentration-dependent manner (Fig. 8A and B). In addition, the
expression of LDH in the cells which was increased by DEX was
inhibited through LRC treatment (Fig. 8C) The expression levels of
pro-apoptotic genes (caspase-6 and -9) were investigated by
semi-quantitative RT-PCR. DEX increased the mRNA levels of both
caspases, and DEX + LRC (100 µg/ml) decreased caspase-6 and
caspase-9 mRNA expression. DEX + LRC (50 µg/ml) and DEX +
LRC (100 µg/ml) significantly increased the expression of
X-linked inhibitor of apoptosis protein (XIAP), and DEX + LRC (100
µg/ml) increased the expression of inhibitor of apoptosis
protein (IAP)-1 and IAP-2 (Fig. 8D
and E).
LRC prevents the GC-induced suppression
of osteoblast differentiation via increasing BMP-2 and Runx2
protein expression
Alizarin Red S staining confirmed that DEX
attenuated the formation of mineralized matrix areas in the
MC3T3-E1 cells (Fig. 9A).
Treatment with DEX significantly reduced the mineralization levels
in MC3T3-E1 cells, and LRC at 100 µg/ml significantly
inhibited this decrease. In addition, the absorbance values
measured by extracting the dye revealed similar results (Fig. 9C). The protein expression levels
of BMP-2 and Runx2 were also measured (Fig. 9B). The results revealed that DEX
+ LRC (100 µg/ml) increased the expression levels of BMP-2
and Runx2, which were inhibited by DEX at the protein level
(Fig. 9D and E).
Effect of LRC on mRNA expression of
factors associated with the GC-induced suppression of osteoblast
differentiation of MC3T3-E1 cells
To evaluate the effects of LRC on the expression of
osteogenic differentiation-related gene markers, cells were treated
with DEX + LRC under osteogenic differentiation conditions for 4
days. Osteoblastic differentiation was measured by
semi-quantitative RT-PCR. The mRNA expression levels of Runx2,
alkaline phosphatase (ALP) and osteocalcin (OCN) were significantly
reduced by DEX treatment (Fig.
10A-D). However, there was no significant effect on the
expression of OSN or collagen type I α1 (Colla1) (Fig. 10A, F and G). As a result of
normalizing the expression of these genes to GAPDH, LRC
significantly increased the mRNA expression of osteoblastic
differentiation markers compared with that of the DEX-treated cells
(Fig. 10).
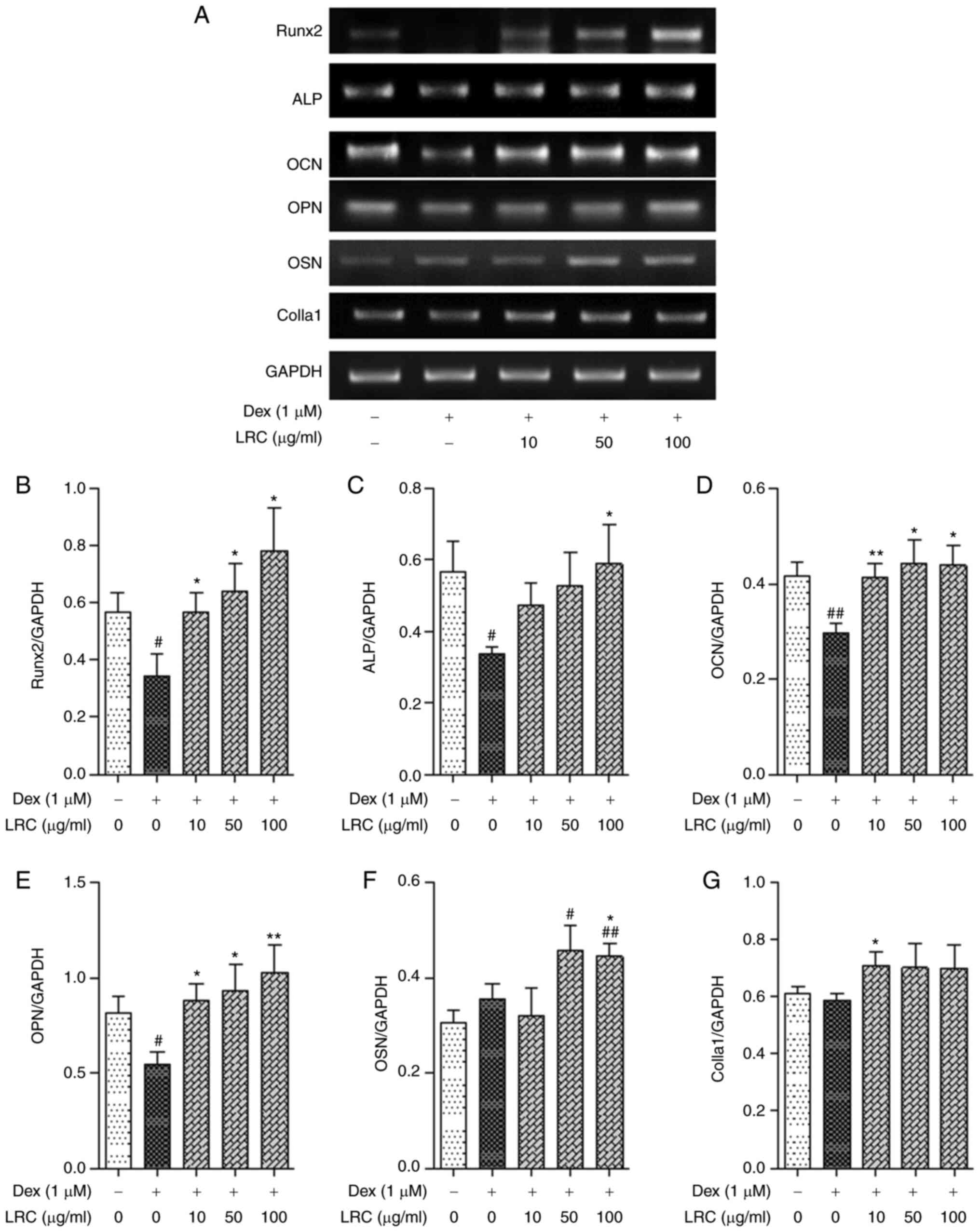 | Figure 10Effect of LRC on osteogenic factors
associated with osteoblast differentiation mRNA expression in
vitro. MC3T3-E1 cells were cultured in osteogenic
differentiation α-MEM containing DEX and LRC for 4 days. (A) The
expression of osteoblast-related genes was analyzed through
semi-quantitative RT-PCR. The expression of (B) Runx2, (C) ALP, (D)
OCN, (E) OPN, (F) OSN and (G) Colla1was normalized by GAPDH. Data
represent the means ± SEM of three independent experiments.
#P<0.05, ##P<0.01 vs. untreated cells;
*P<0.05, **P<0.01 vs. DEX-treated
cells. LRC, Lycii radicis cortex; DEX, dexamethasone;
RT-PCR, reverse transcription-PCR; Runx2, runt-related
transcription factor 2; ALP, alkaline phosphatase; OCN,
osteocalcin; OPN, osteopontin; OSN, osteonectin; Colla1, collagen
type I α1. |
Effect of LRC on GC-induced resistance in
macrophages
Macrophages play an important role in immunity and
immune responses (25). In the
present study, LRC or DEX + LRC did not exert any toxicity on RAW
264.7 macrophage cells (Fig. 11A
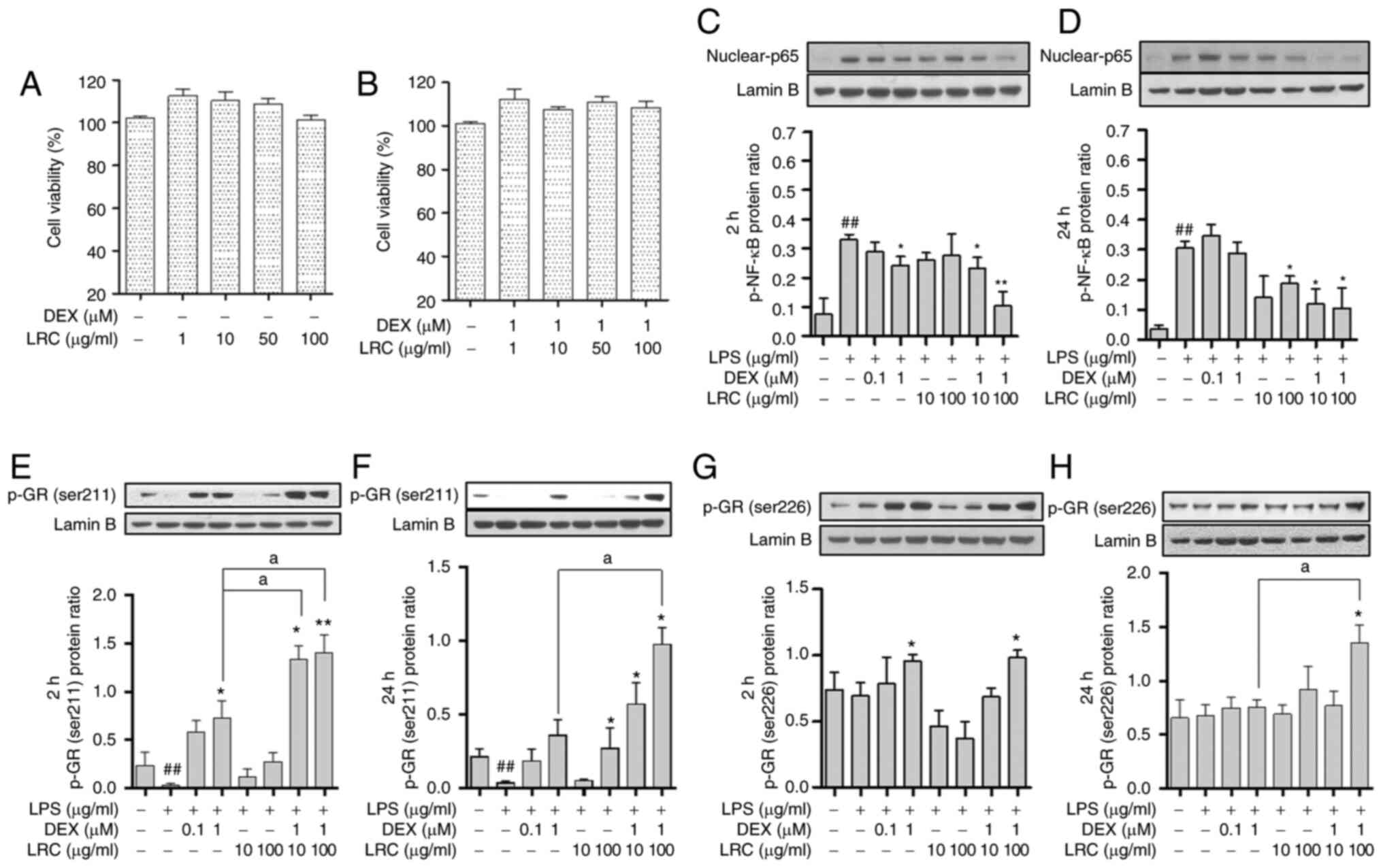
and B). DEX significantly inhibited the p-NF-κB levels at 2 h,
while DEX decreased p-NF-κB expression at 24 h, although the
difference was not significant. When LRC was co-treated with DEX,
it inhibited p-NF-κB at both 2 and 24 h (Fig. 11C and D). GC resistance is
considered the main cause of the GC-induced side-effects (26). In the present study, treatment
with DEX increased the expression of p-GR at 2 h (Fig. 11E and G). LRC also increased the
levels of p-GR. However, at 24 h (long-term), the expression of
p-GR decreased at <2 h due to GC resistance. DEX + LRC prevented
this inhibition of p-GR expression in the long-term (Fig. 11F and H). p-NF kB was also
identified as an inflammatory mediator that was associated with the
inhibitory effect of DEX + LRC on p-GR resistance.
Identification of LRC through GC/MS
analysis
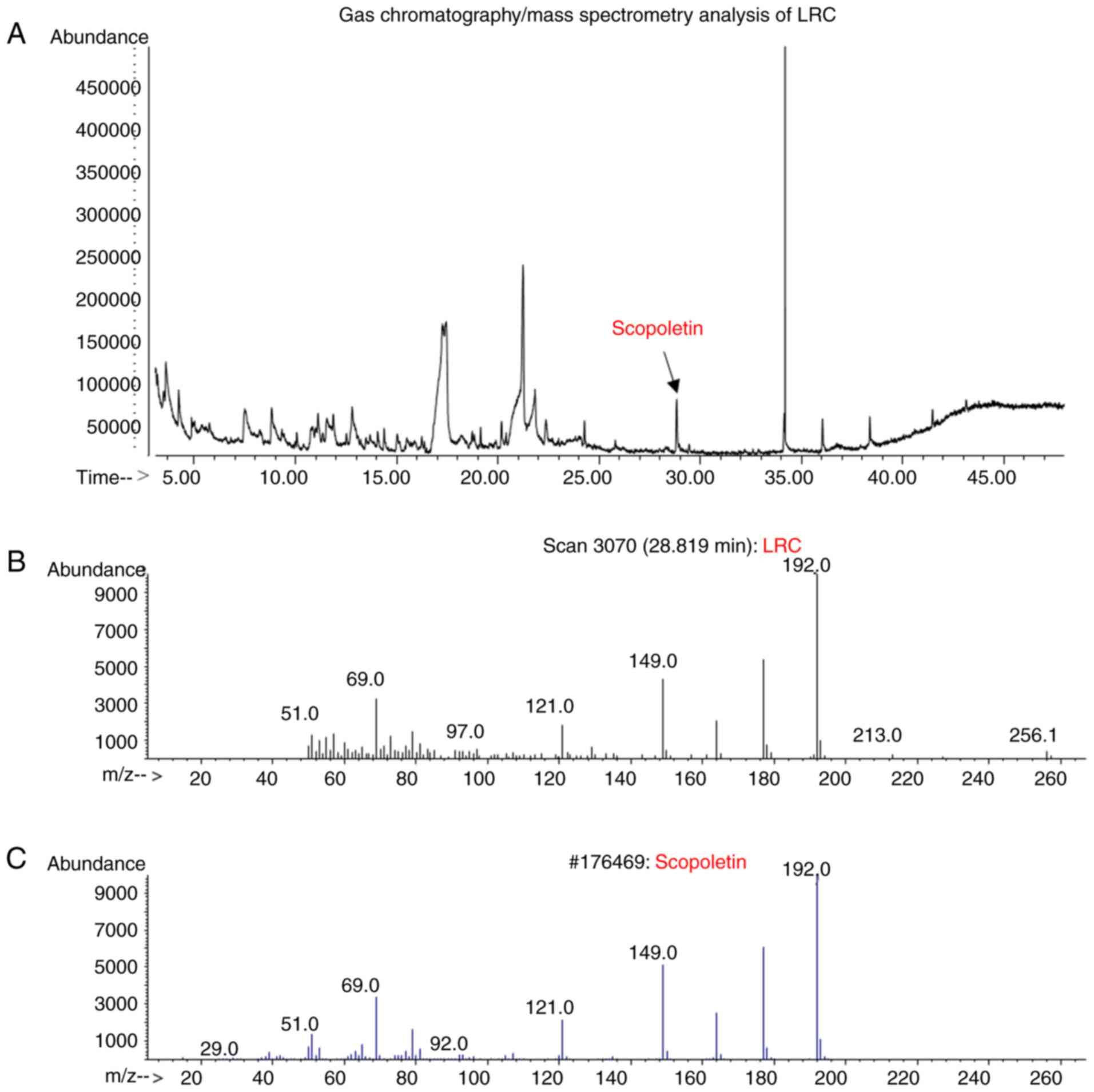
Scopoletin has been reported to be a constituent
compound of LRC (27). In the
present study, GC/MS analysis revealed that scopoletin was detected
in the LRC water extract, and the results were compared with
scopoletin standard to confirm that the peaks matched (Fig. 12).
Discussion
Inflammatory diseases, including allergic
dermatitis, are treated with GC prescriptions (28). Despite their side-effects, GCs
are prescribed as a treatment for the majority of allergic
dermatitis cases as GC topical treatment is effective and has a low
cost (29). Although patients
with ACD are being treated with GC under strict guidelines, there
is still a high risk of GIOP (30,31). Despite extensive research on
GIOP, previous models were induced without the underlying disease,
which may not reflect the actual situation of the patient.
Therefore, the present study compared GC-treated mice with ACD and
GC-treated mice without ACD. In the present study, a GIOP model was
established in the case of ACD being the underlying disease as a
precondition of secondary osteoporosis for depicting the patients'
condition. The results revealed that GIOP did not occur in the
absence of the underlying disease, ACD. This differs from
previously published reports stating that DEX decreased BMD in mice
(32,33). However, other studies have
reported that DEX increases the bone progenitor ability of bone
progenitor cells in vitro (34,35). The difference between these
results may be due to a variety of reasons, including the
concentration of DEX used in the study or the duration of the
experiment. The present study found that LRC had an effect on GIOP
and also proved to enhance immunity against DEX application, when
administered orally, as in the treatment of clinical patients.
Overall, the results of the present study suggest that the use of
LRC alleviates the side-effects of GCs.
Osteoporosis is a typical side-effect of GCs, where
the bone quality decreases due to changes in bone density and
microstructure (36). The
qualitative increase in cancellous bone structures such as Tb.Sp,
Tb.N and BS/TV along with the increase in bone mass through BMD and
BV/TV is important. In the present study, LRC significantly
suppressed the loss of bone density induced by DEX, and led to a
qualitative improvement of various bone microstructures. These
results suggest that LRC can attenuate GIOP. Calcium is the most
important mineral for bone formation and for maintaining the
skeleton. Calbindin-D28k, a calcium-binding protein, is known to be
an important factor for calcium metabolism and absorption of
calcium by metabolic organs such as the duodenum and kidney
(37,38). It has been demonstrated
calbindin-D28k is produced by osteoblastic cells, and protects
osteoblasts from apoptosis (39). Therefore, calbindin-D28k is
expected to play an important role in the mechanism leading to
secondary osteoporosis. In a previous study by the authors, the
expression of calbindin-D28k was observed in the duodenum and
kidneys of mice (40). In the
present study, IHC analysis of calbindin-D28k expression in kidney
and duodenal tissue revealed that LRC increased the
calcium-absorbing ability which was decreased by DEX. These results
indicate that LRC is involved in the protein-mediated mechanism of
calcium-absorption associated with the inhibition of osteoblast
death.
In addition, the present study aimed to verify
whether LRC interferes with the anti-inflammatory effects of DEX in
an ACD model. LRC was used along with DEX to demonstrate that LRC
complemented the effects of DEX and decreased its side-effects
in vivo and in vitro. Mast cells can release
inflammatory mediators and cytokines for participating in
inflammatory reactions such as flushing, itching and swelling
(41). In previous animal
experiments, mast cells were more significantly reduced by DEX +
LRC than by DEX only, and the eosinophil infiltration number was
maintained by treatment with LRC + DEX compared with that of DEX
treatment alone. Epidermis and dermis atrophy are side-effects of
GCs (42). In the present study,
LRC significantly reduced dermis atrophy. LRC usage combined with
DEX did not interfere with the anti-inflammatory effects of DEX.
This suggests that the experimental combination of existing drugs
and herbal medicines may be able to show synergy and may have a
valuable potential.
Long-term DEX use has been reported to lead to the
up-regulation of caspases-3, -6 and -9, which in turn leads to the
apoptosis of osteoblasts (43).
Prevention of osteoblast apoptosis is an important mechanism in the
pathogenesis of secondary osteoporosis. In the present study, LRC
inhibited caspase-6 and -9 expression. Caspase-6 is an initiator of
the caspase pathway, which is involved in mitochondrial damage
(44). The activation of
caspase-9 is inhibited by members of the IAP family, including XIAP
(45). Caspase-6, an effector
caspase that is activated by caspase-9 in the apoptosome, can also
be blocked by XIAP (46). The
relative expression level of XIAP is one of the main factors
regulating mitochondrial apoptosis downstream processes (47). According to previous studies, DEX
increases the protein levels of caspase-6, caspase-9 and MC3T3-E1,
suggesting that DEX is involved in apoptosis (16,44). In the present study, LRC
decreased the levels of caspase-6 and -9 that were induced by DEX.
Furthermore, LRC increased the levels of XIAP and IAPs. These
results indicate that LRC inhibits osteoblast apoptosis induced by
DEX through a caspase-mediated mechanism.
The reduction in bone formation due to the death of
osteoblasts causes an imbalance in bone metabolism with
osteoclasts, resulting in osteoporosis. Therefore, it is important
to determine the expression of bone-forming factors, such as BMP-2,
Runx2, ALP, OCN, osteopontin (OPN), osteonectin (OSN) and Colla1,
along with apoptotic markers (48). Initially, osteoblasts actively
proliferate to produce type I collagen extracellular matrix
(49). The expression of ALP is
enhanced immediately following the proliferation period, and
mineralization begins, which results in an increased expression of
OCN and OPN (49). The
expression of the OCN and ALP genes is known to be stimulated by
BMP-2 (50,51). Runx2 is a transcription factor,
and its activity is increased by BMP-2 signaling (51). Runx2 binds to osteogenic genes to
regulate the expression of Colla1, OPN and OCN (a late bone
differentiation marker) (52).
In the present study, LRC improved the osteoblast mineralization
inhibited by DEX, and it also increased the expression of Runx2,
BMP-2, ALP, OCN, OPN, OSN and Colla1, which are involved in
osteoblast mineralization and differentiation. These results
demonstrate that LRC increases the differentiation of osteoblasts
inhibited by DEX through a Runx2- and BMP-2-mediated mechanism.
This confirms the effect of LRC, and the osteoblast differentiation
effect of LRC was demonstrated through the indicators of the exact
mechanism (16). The results of
osteoblast differentiation and death in vitro were
consistent with the results of the animal experiments, which
suggests that combined treatment with DEX and LRC inhibited
osteoporosis in animals through the inhibition of osteoblast
apoptosis and differentiation.
GC resistance is the cause of GC-associated
side-effects, since GC resistance leads to an increase in the
length of GC treatment and in the dose of GCs (9). GR is known to be the main factor of
the GC resistance mechanism (9,10). To investigate the effects of LRC
on GC resistance, the authors performed GR Ser-211 and GR Ser-226
protein expression experiments in macrophages, which are innate
immune cells that play a central role in the inflammatory response
(26). The sensitivity of GR is
closely associated with GC resistance. GC binds to ligands to
activate receptors, and the complex is translocated from the
cytoplasm to the nucleus (53,54). GR regulates gene expression by
interacting with transcription factors and NF-κB, and the majority
of these mechanisms have anti-inflammatory actions (55). Furthermore, negative GR elements
(GREs), across the site of transcription, can regulate the
functions of bone (mediated by OCN), skin (via keratins) and
inflammation (56). A reduction
in the receptor's affinity to the ligand is mainly responsible for
GC resistance (57). The reason
for this reduced affinity is suspected to be: i) Delayed or failed
translocation of the GC-GR complex to the nucleus; and ii)
decreased transcriptional activity due to decreased GRE binding
(57). The phosphorylation sites
of GR are Ser203, Ser211 and Ser226, where the numbers refer to the
numbering scheme of the N-terminal transcriptional regulatory
domain of human GR. Previous studies have reported that p-GR Ser211
and Ser226 (but not Ser203) appear to contribute to gene
transcriptional responses to GCs (58,59). The results of the present
demonstrated that LRC increased the expression of p-GR transcribed
in the nucleus via the GC response. In addition, it was observed
that LRC increased and maintained the expression p-GR, which was
decreased by GC resistance caused by the CG long-term response.
These results suggest that LRC exerts a positive effect on GC
resistance. Therefore, LRC supports subsequent regulatory immune
responses. The increased p-GR expression effect of LRC inhibits GC
resistance, which is the cause of GC side-effects; however, it is
unclear whether this result directly affects secondary
osteoporosis. Therefore, further studies are required to compare
the anti-inflammatory effects of DEX at a high dose without LRC
with those of DEX at a low dose with LRC.
The present study has the following limitations:
There was no experimental group treated only with DNCB. Thus, the
question of whether osteoporosis can be induced by treatment with
DNCB alone may be reconsidered. To eliminate this concern, it is
necessary to demonstrate in future studies whether osteoporosis is
induced by treatment with DNCB alone for a long period of time. In
addition, the present study considered that atrophy of the skin
occurred due to long-term DEX treatment. Previous studies have
indicated that when GC is applied to mice for 18 days (60), skin atrophy similar to that of
humans is found. Since the present study applied DEX for a much
longer period (namely 8 weeks), it can be considered that the skin
exhibited side-effects. In the future, these claims should be
verified by comparing DNCB-only treatment and DEX treatment
groups.
In conclusion, the present study focused on the
possibility that herbal medicines could overcome the side-effects
of GCs and may become a complete replacement for GC treatment. The
present study identified the method that was most suitable for
secondary osteoporosis animal models. The results of the present
study are meaningful in establishing and demonstrating the value of
the combination of herbal medicine and GC. The present results
revealed that LRC inhibited osteoblast apoptosis induced by DEX and
increased the levels of osteoblast differentiation-related genes.
LRC also significantly reduced GIOP as a side-effect of DEX, while
maintaining its anti-inflammatory effects. This suggests that LRC
has the potential to be used and developed as a drug to relieve the
side-effects of GCs and to maximize their effectiveness in
combination with DEX (Fig.
13).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YS and JHK conceptualized the study. BL performed
all the experiments. MK, EYK, HJP and HSJ contributed to the
statistical analysis. SH, MK, HJP and HSJ assisted with the
interpretation of the results. BL drafted the manuscript. BL and
JHK confirm the authenticity of all the raw data. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Kyung
Hee University Animal Care and Use Committee
[KHUASP(SE)-17-082].
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
References
|
1
|
Frenkel B, White W and Tuckermann J:
Glucocorticoid-induced osteoporosis. Adv Exp Med Biol. 872:179–215.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Henneicke H, Gasparini SJ,
Brennan-Speranza TC, Zhou H and Seibel MJ: Glucocorticoids and
bone: Local effects and systemic implications. Trends Endocrinol
Metab. 25:197–211. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mirza F and Canalis E: Management of
endocrine disease: Secondary osteoporosis: pathophysiology and
management. Eur J Endocrinol. 173:R131–151. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tuck S and Francis R: Testosterone, bone
and osteoporosis. Front Horm Res. 37:123–132. 2009. View Article : Google Scholar
|
|
5
|
Briot K and Roux C: Glucocorticoid-induced
osteoporosis. RMD Open. 1:e0000142015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shen C, Chen F, Zhang Y, Guo Y and Ding M:
Association between use of antiepileptic drugs and fracture risk: A
systematic review and meta-analysis. Bone. 64:246–253. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moura C, Bernatsky S, Abrahamowicz M,
Papaioannou A, Bessette L, Adachi J, Goltzman D, Prior J, Kreiger
N, Towheed T, et al: Antidepressant use and 10-year incident
fracture risk: The population-based Canadian Multicentre
Osteoporosis Study (CaMoS). Osteoporos Int. 25:1473–1481. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Van Staa TP, Leufkens HG, Abenhaim L,
Zhang B and Cooper C: Use of oral corticosteroids and risk of
fractures. J Bone Miner Res. 15:993–1000. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Barnes PJ and Adcock IM: Glucocorticoid
resistance in inflammatory diseases. Lancet. 373:1905–1917. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schaaf MJ and Cidlowski JA: Molecular
mechanisms of glucocorticoid action and resistance. J Steroid
Biochem Mol Biol. 83:37–48. 2002. View Article : Google Scholar
|
|
11
|
Xie LW, Atanasov AG, Guo DA, Malainer C,
Zhang JX, Zehl M, Guan SH, Heiss EH, Urban E, Dirsch VM and Kopp B:
Activity-guided isolation of NF-κB inhibitors and PPARγ agonists
from the root bark of Lycium chinense Miller. J Ethnopharmacol.
152:470–477. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao D, Li Q, Liu Z, Li Y, Liu Z, Fan Y, Li
K, Han Z and Li J: Hypoglycemic effects and mechanisms of action of
Cortex Lycii Radicis on alloxan-induced diabetic mice. Yakugaku
Zasshi. 127:1715–1721. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim JH, Kim EY, Lee B, Min JH, Song DU,
Lim JM, Eom JW, Yeom M, Jung HS and Sohn Y: The effects of Lycii
Radicis Cortex on RANKL-induced osteoclast differentiation and
activation in RAW 264.7 cells. Int J Mol Med. 37:649–658. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Park E, Jin HS, Cho DY, Kim J, Kim MC,
Choi CW, Jin Y, Lee JW, Park JH, Chung YS, et al: The effect of
Lycii Radicis Cortex extract on bone formation in vitro and in
vivo. Molecules. 19:19594–19609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park E, Kim J, Kim MC, Yeo S, Kim J, Park
S, Jo M, Choi CW, Jin HS, Lee SW, et al: Anti-osteoporotic effects
of kukoamine B Isolated from Lycii radicis cortex extract on
osteoblast and osteoclast cells and ovariectomized osteoporosis
model mice. Int J Mol Sci. 20:27842019. View Article : Google Scholar :
|
|
16
|
Kim J, Lee H, Kang KS, Chun KH and Hwang
GS: Protective effect of Korean Red Ginseng against
glucocorticoid-induced osteoporosis in vitro and in vivo. J Ginseng
Res. 39:46–53. 2015. View Article : Google Scholar
|
|
17
|
Ma Y, Yang H and Huang J: Icariin
ameliorates dexamethasone-induced bone deterioration in an
experimental mouse model via activation of microRNA186 inhibition
of cathepsin K. Mol Med Rep. 17:1633–1641. 2018.
|
|
18
|
Kim EY, Kim JH, Kim M, Park JH, Sohn Y and
Jung HS: Abeliophyllum distichum Nakai alleviates postmenopausal
osteoporosis in ovariectomized rats and prevents RANKL-induced
osteoclastogenesis in vitro. J Ethnopharmacol. 257:1128282020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim M, Kim HS, Kim JH, Kim EY, Lee B, Lee
SY, Jun JY, Kim MB, Sohn Y and Jung HS: Chaenomelis fructus
inhibits osteoclast differentiation by suppressing NFATc1
expression and prevents ovariectomy-induced osteoporosis. BMC
Complement Med Ther. 20:352020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim JH, Kim M, Jung HS and Sohn Y:
Leonurus sibiricus L. ethanol extract promotes osteoblast
differentiation and inhibits osteoclast formation. Int J Mol Med.
44:913–926. 2019.PubMed/NCBI
|
|
21
|
Konappa N, Udayashankar AC, Krishnamurthy
S, Pradeep CK, Chowdappa S and Jogaiah S: GC-MS analysis of
phytoconstituents from Amomum nilgiricum and molecular docking
interactions of bioactive serverogenin acetate with target
proteins. Sci Rep. 10:164382020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang EY, Chen AY and Zhu BT: Mechanism of
dinitrochlorobenzene-induced dermatitis in mice: Role of specific
antibodies in pathogenesis. PLoS One. 4:e77032009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Park G, Kim HG, Lim S, Lee W, Sim Y and Oh
MS: Coriander alleviates 2,4-dinitrochlorobenzene-induced contact
dermatitis-like skin lesions in mice. J Med Food. 17:862–868. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hiura K, Sumitani K, Kawata T, Higashino
K, Okawa M, Sato T, Hakeda Y and Kumegawa M: Mouse osteoblastic
cells (MC3T3-E1) at different stages of differentiation have
opposite effects on osteoclastic cell formation. Endocrinology.
128:1630–1637. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Elhelu MA: The role of macrophages in
immunology. J Natl Med Assoc. 75:314–317. 1983.PubMed/NCBI
|
|
26
|
van Rossum EF and Lamberts SW:
Glucocorticoid resistance syndrome: A diagnostic and therapeutic
approach. Best Pract Res Clin Endocrinol Metab. 20:611–626. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Potterat O: Goji (Lycium barbarum and L.
Chinense): Phytochemistry, pharmacology and safety in the
perspective of traditional uses and recent popularity. Planta Med.
76:7–19. 2010. View Article : Google Scholar
|
|
28
|
Moghadam-Kia S and Werth VP: Prevention
and treatment of systemic glucocorticoid side effects. Int J
Dermatol. 49:239–248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lagos BR and Maibach HI: Frequency of
application of topical corticosteroids: An overview. Br J Dermatol.
139:763–766. 1998. View Article : Google Scholar
|
|
30
|
Haeck I, van Velsen S, de Bruin-Weller M
and Bruijnzeel-Koomen C: Bone mineral density in patients with
atopic dermatitis. Chem Immunol Allergy. 96:96–99. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Garg NK and Silverberg JI: Eczema is
associated with osteoporosis and fractures in adults: A US
population-based study. J Allergy Clin Immunol. 135:1085–1087 e2.
2015. View Article : Google Scholar
|
|
32
|
Zhang J, Song J and Shao J: Icariin
attenuates glucocorticoid-induced bone deteriorations, hypocalcemia
and hypercalciuria in mice. Int J Clin Exp Med. 8:7306–7314.
2015.PubMed/NCBI
|
|
33
|
Fan S, Gao X, Chen P and Li X: Myricetin
ameliorates glucocorticoid-induced osteoporosis through the ERK
signaling pathway. Life Sci. 207:205–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shalhoub V, Conlon D, Tassinari M, Quinn
C, Partridge N, Stein GS and Lian JB: Glucocorticoids promote
development of the osteoblast phenotype by selectively modulating
expression of cell growth and differentiation associated genes. J
Cell Biochem. 50:425–440. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Igarashi M, Kamiya N, Hasegawa M, Kasuya
T, Takahashi T and Takagi M: Inductive effects of dexamethasone on
the gene expression of Cbfa1, Osterix and bone matrix proteins
during differentiation of cultured primary rat osteoblasts. J Mol
Histol. 35:3–10. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Parkinson IH and Fazzalari NL:
Interrelationships between structural parameters of cancellous bone
reveal accelerated structural change at low bone volume. J Bone
Miner Res. 18:2200–2205. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kutuzova GD, Akhter S, Christakos S,
Vanhooke J, Kimmel-Jehan C and Deluca HF: Calbindin D(9k) knockout
mice are indistinguishable from wild-type mice in phenotype and
serum calcium level. Proc Natl Acad Sci USA. 103:12377–12381. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Christakos S, Gabrielides C and Rhoten WB:
Vitamin D-dependent calcium binding proteins: Chemistry,
distribution, functional considerations, and molecular biology.
Endocr Rev. 10:3–26. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bellido T, Huening M, Raval-Pandya M,
Manolagas SC and Christakos S: Calbindin-D28k is expressed in
osteoblastic cells and suppresses their apoptosis by inhibiting
caspase-3 activity. J Biol Chem. 275:26328–26332. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim MH, Lee GS, Jung EM, Choi KC, Oh GT
and Jeung EB: Dexamethasone differentially regulates renal and
duodenal calcium-processing genes in calbindin-D9k and -D28k
knockout mice. Exp Physiol. 94:138–151. 2009. View Article : Google Scholar
|
|
41
|
Church MK and Levi-Schaffer F: The human
mast cell. J Allergy Clin Immunol. 99:155–160. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schoepe S, Schacke H, May E and Asadullah
K: Glucocorticoid therapy-induced skin atrophy. Exp Dermatol.
15:406–420. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ji YZ, Geng L, Zhou HB, Wei HC and Chen
HD: Chinese herbal medicine Yougui Pill reduces exogenous
glucocorticoid-induced apoptosis in anterior pituitary cells.
Neural Regen Res. 11:1962–1968. 2016. View Article : Google Scholar
|
|
44
|
Chua CC, Chua BH, Chen Z, Landy C and
Hamdy RC: Dexamethasone induces caspase activation in murine
osteoblastic MC3T3-E1 cells. Biochim Biophys Acta. 1642:79–85.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li P, Nijhawan D, Budihardjo I,
Srinivasula SM, Ahmad M, Alnemri ES and Wang X: Cytochrome c and
dATP-dependent formation of Apaf-1/caspase-9 complex initiates an
apoptotic protease cascade. Cell. 91:479–489. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fischer U and Schulze-Osthoff K: New
approaches and therapeutics targeting apoptosis in disease.
Pharmacol Rev. 57:187–215. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Trapp T, Korhonen L, Besselmann M,
Martinez R, Mercer EA and Lindholm D: Transgenic mice
overexpressing XIAP in neurons show better outcome after transient
cerebral ischemia. Mol Cell Neurosci. 23:302–313. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Riggs BL and Melton LJ III: The prevention
and treatment of osteoporosis. N Engl J Med. 327:620–627. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Owen TA, Aronow M, Shalhoub V, Barone LM,
Wilming L, Tassinari MS, Kennedy MB, Pockwinse S, Lian JB and Stein
GS: Progressive development of the rat osteoblast phenotype in
vitro: Reciprocal relationships in expression of genes associated
with osteoblast proliferation and differentiation during formation
of the bone extracellular matrix. J Cell Physiol. 143:420–430.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Huang W, Rudkin GH, Carlsen B, Ishida K,
Ghasri P, Anvar B, Yamaguchi DT and Miller TA: Overexpression of
BMP-2 modulates morphology, growth, and gene expression in
osteoblastic cells. Exp Cell Res. 274:226–234. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lee KS, Kim HJ, Li QL, Chi XZ, Ueta C,
Komori T, Wozney JM, Kim EG, Choi JY, Ryoo HM and Bae SC: Runx2 is
a common target of transforming growth factor beta1 and bone
morphogenetic protein 2, and cooperation between Runx2 and Smad5
induces osteoblast-specific gene expression in the pluripotent
mesenchymal precursor cell line C2C12. Mol Cell Biol. 20:8783–8792.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gu H, Boonanantanasarn K, Kang M, Kim I,
Woo KM, Ryoo HM and Baek JH: Morinda citrifolia leaf extract
enhances osteogenic differentiation through activation of
Wnt/beta-catenin signaling. J Med Food. 21:57–69. 2018. View Article : Google Scholar
|
|
53
|
Schaaf MJ and Cidlowski JA: Molecular
determinants of glucocorticoid receptor mobility in living cells:
The importance of ligand affinity. Mol Cell Biol. 23:1922–1934.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Maltese P, Canestrari E, Palma L, Ruzzo A,
Corini F, Menotta M, Andreoni F, Latiano A, Annese V and Magnani M:
High resolution melting (HRM) analysis for the detection of
ER22/23EK, BclI, and N363S polymorphisms of the glucocorticoid
receptor gene. J Steroid Biochem Mol Biol. 113:269–274. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
De Bosscher K, Vanden Berghe W and
Haegeman G: The interplay between the glucocorticoid receptor and
nuclear factor-kappaB or activator protein-1: Molecular mechanisms
for gene repression. Endocr Rev. 24:488–522. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Dostert A and Heinzel T: Negative
glucocorticoid receptor response elements and their role in
glucocorticoid action. Curr Pharm Des. 10:2807–2816. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Charmandari E, Kino T, Ichijo T and
Chrousos GP: Generalized glucocorticoid resistance: Clinical
aspects, molecular mechanisms, and implications of a rare genetic
disorder. J Clin Endocrinol Metab. 93:1563–1572. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Blind RD and Garabedian MJ: Differential
recruitment of glucocorticoid receptor phospho-isoforms to
glucocorticoid-induced genes. J Steroid Biochem Mol Biol.
109:150–157. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wang Z, Chen W, Kono E, Dang T and
Garabedian MJ: Modulation of glucocorticoid receptor
phosphorylation and transcriptional activity by a
C-terminal-associated protein phosphatase. Mol Endocrinol.
21:625–634. 2007. View Article : Google Scholar
|
|
60
|
Woodbury R and Kligman AM: The hairless
mouse model for assaying the atrophogenicity of topical
corticosteroids. Acta Derm Venereol. 72:403–406. 1992.PubMed/NCBI
|