Introduction
PANoptosis is a newly defined programmed cell death
(PCD) involving apoptosis, pyroptosis and necroptosis, presenting
an innovative, integrated view of cellular death mechanisms. This
paradigm underscores the intricate manner in which cells, at a
molecular level, orchestrate diverse signaling pathways to elicit a
comprehensive cell death response. PANoptosis has a pivotal role in
modulating immune responses and inflammatory processes, and in
determining cell fate (1). For
instance, it was recently shown that PANoptosis can integrate
disparate cell death pathways through the interplay of molecular
mechanisms, such as absent in melanoma 2, Pyrin and Z-DNA binding
protein 1 (ZBP1). This insight offers a novel perspective in
comprehending host defense mechanisms (2). Furthermore, the involvement of
PANoptosis in viral infections, particularly in cases of influenza
and herpes simplex virus type 1, has attracted considerable
scholarly interest, and its significant function in the mechanisms
of viral infection is coming to light (3). The occurrence of PANoptosis in
ischemic brain injuries highlights it as a potential therapeutic
target for managing central nervous system disorders (4). The discernible patterns of
PANoptosis have demonstrated their potential in predicting the
survival rates and responses to immune therapies in patients with
gastric cancer, thereby underscoring its profound impact on the
progression of this disease (5).
The role of PANoptosis in regulating the tumor microenvironment and
advancing cancer immunotherapy has been particularly highlighted,
providing novel avenues for cancer treatment research (6). Collectively, these findings not
only enhance our comprehension of the multifaceted nature of
PANoptosis but also pave the way for innovative approaches to
treating and preventing diseases.
Cardiovascular diseases (CVDs) represent a
significant global health concern, encompassing a spectrum of
conditions such as coronary artery disease, heart failure (HF) and
hypertension. Atherosclerosis (AS), myocardial infarction (MI), as
well as myocardial ischemia-reperfusion, are often accompanied by
cell death and acute/chronic inflammatory reactions. An increasing
number of studies have demonstrated that apoptosis, necroptosis or
pyroptosis participate in CVD progression and may thus have
promising therapeutic implications (7-9).
However, only one study has linked PANoptosis to CVDs, to the best
of our knowledge; Bi et al (10) found that FUN14 domain containing
1 (FUNDC1), a mitophagy receptor, protects against
doxorubicin-induced cardiomyocyte PANoptosis by stabilizing
mitochondrial DNA via interaction with Tu translation elongation
factor mitochondrial (TUFM). The present study revealed that it is
of notable significance to further study the role of PANoptosis in
CVDs. Little is understood regarding the function of PANoptosis in
CVDs. Therefore, there remains considerable room for research on
the specific mechanism of action of PANoptosis in CVDs, and thus,
PANoptosis may have a significant impact in this field.
In the present review, the role of PANoptosis in
CVDs is explored, focusing on its potential as a novel therapeutic
target. The definition and molecular mechanisms of apoptosis,
pyroptosis, necroptosis and PANoptosis are outlined, emphasizing
their pathophysiological impact on CVD progression. In addition,
the challenges and future directions in this emerging field are
discussed, aiming to provide a concise yet comprehensive overview
of the current body of knowledge and potential advancements in CVD
treatment.
Programmed cell death
Cell death, a meticulously regulated process, is
strongly influenced by the cellular environment and numerous
stimuli. PCD is executed through a suite of genetically encoded
mechanisms. PCD includes apoptosis, necroptosis and pyroptosis, and
it is an irreversible phenomenon (11). Broadly speaking, PCD pathways are
classified into lytic and non-lytic categories. The lytic forms,
such as pyroptosis, necroptosis and PANoptosis, are characterized
as 'violent' cell death types (12,13), as they are accompanied by a
violent inflammatory response. They entail the loss of membrane
potential and cellular swelling, leading to disruption of cellular
integrity and the consequent release of potent inflammatory
inducers. Conversely, non-lytic forms of PCD, characterized by
apoptosis, involve the systematic disintegration of dying cells
into smaller entities, termed apoptotic bodies. This process
ensures the containment of cellular contents, thereby averting
inflammatory responses (14). It
is particularly noteworthy that PANoptosis, a relatively newly
discovered form of cell death, uniquely combines aspects of various
PCD pathways including apoptosis, necroptosis and pyroptosis, and
is thus a unique, comprehensive form of cell death. Its
distinctiveness lies in the interplay of multiple signaling
pathways and molecular mechanisms, activated under a range of
conditions, leading to cell death. This versatile cell death
approach enhances cellular adaptability to stress and injury.
Research into PANoptosis is vital for understanding cellular
responses to death signals in inflammatory and infectious diseases
and cancer. Thus, studying PANoptosis may improve our understanding
of cell death and unveil novel therapeutic targets for the
management of these diseases. The present review examines the
intricate dynamics between various forms of cell death, apoptosis,
pyroptosis and necroptosis, and their consequential roles in CVDs.
The molecular mechanisms driving PANoptosis are methodically
outlined, emphasizing crucial components such as ZBP1,
receptor-interacting protein kinase 1 (RIPK1), RIPK3, mixed lineage
kinase domain-like pseudokinase (MLKL), TNF and toll-like receptor
(TLR) (Table I).
 | Table ICell death pathways: From programmed
cell death to PANoptosis. |
Table I
Cell death pathways: From programmed
cell death to PANoptosis.
| Disease | Death mode |
Target/intervention | Mechanism | (Refs.) |
|---|
|
Atherosclerosis | Apoptosis | Ox-LDL | Activates the
Fas/Fasl pathway | (26) |
| Hyperglycemia | Triggers
inflammatory cytokines and initiates a caspase cascade through
NF-κB pathway activation | (27) |
| Paeonol | The inhibitory
effect of Paeonol on apoptosis in VEC induced by high glucose/high
pressure is mediated through the regulation of the
SIRT1/FOXO3a/NF-κB pathway | (28) |
| Geniposide +
Notoginsenoside R1 | Activates
AMPK/mTOR/Nrf2 signaling pathway, inhibiting the NLRP3 inflammasome
and Bax/Bcl2/caspase-3 pathway | (29) |
| M-CSF | CSF1 deficiency
reduces macrophage proliferation and promotes macrophage apoptosis
(activated caspase-3) | (31) |
| Pyroptosis | Nicotine | Elevates caspase-1
expression | (52) |
| TMAO | Activates NF-κB and
MAPK signaling pathways, worsening endothelial dysfunction and
promoting AS plaque accumulation | (53) |
| High-fat diet +
TMAO | Increases caspase-1
and NLRP3 expression, accelerating AS progression | (54) |
| HDAC6 | Key role of
ROS/NF-κB/NLRP3 axis and mitochondria in macrophage pyroptosis | (56,57) |
| Eatp | Caspase-11/GSDMD
pyroptosis pathway | (58) |
| Irisin | Inhibition of
NLRP3-mediated pyroptotic cell death | (60) |
| Caspase-1 inhibitor
VX-765 | Significantly
reduces ox-LDL-induced endothelial cell pyroptosis | (61) |
| Necroptosis | RIP3 | RIP3 signaling
pathway | (77,78) |
| MLKL | MLKL directly
contributes to lesion development and necrotic core formation | (82) |
| Oxidative
stress | Exacerbating
oxidative stress, activates HIF-1α | (83) |
| Myocardial
infarction | Apoptosis | miR-182-5p | Suppressed Bcl-2
expression, and increased Bax, Bnip3, and caspase-3/7 activity
levels | (32) |
| IGF-1 | AKT/SFRP2/β-catenin
pathway | (33) |
| miR-338 | Inhibits
cardiomyocyte apoptosis in MI through the MAP3K2/JNK signaling
pathway | (34) |
| MEG3 (lncRNA) | Promotes apoptosis
(the expression of Caspase-3 and Bcl-2) | (35) |
| miR-146b | Mediates vascular
inflammation and apoptosis in MI, potentially through the
PI3K/Akt/NF-κB pathway | (36) |
| Gm18840 | Gm18840 drives
cardiomyocyte apoptosis | (37) |
| Pyroptosis | Calpain | NLRP3/ASC/caspase-1
axis | (62) |
| SIRT1 | Targeting oxidative
stress and NLRP3-mediated cell pyroptosis | (63) |
| Caspase-1 | Promotes
pyroptosis | (64,65) |
| Necroptosis | RIP3 | Via the
RIP1-RIP3-MLKL and RIP3-camkii pathways | (84,85) |
| TAK1 | Traf2 critically
regulates RIP1, RIP3 and MLKL necroptotic signaling with
TNFR1-associated death domain protein as an upstream regulator and
TAK1 as a downstream effector | (86) |
| TRAF2 | RIP1-mediated
necrosis | (87) |
| S-allyl cysteine
sulfoxide | Inhibits the
expression of RIP1, RIP3 and TRAF2 | (89) |
| miR-325-3p |
RIPK1/RIPK3/p-MLKL | (88) |
| Ad-HGF | Ad-HGF
significantly decreases the caspase 8 protein and activity levels,
which urges the cell to undergo necroptosis under hypoxia and block
of apoptosis | (90) |
| Heart failure | Apoptosis | miR-182 | miR-182 inhibits
cardiomyocyte apoptosis induced by non-ischemic HF via
downregulating PDCD4 and PACS2 | (38) |
| BYD | Ameliorates
myocardial apoptosis via the P38 MAPK-CRYAB pathway | (39) |
| Shenfu | Shenfu formula can
regulate the initiative factors Fas/Fas-L in the intrinsic pathway
and Bcl-2/Bax in the extrinsic apoptosis pathway to suppress
apoptosis | (40) |
| Pyroptosis | NLRP3 | Promotes collagen
synthesis and activates caspase-1, leading to IL-1β and IL-18
release, inducing pyroptosis | (66,67) |
| Caspase-1 | Regulates Ang
II-induced cardiac hypertrophy through IL-1β regulation | (68) |
| BMP-7 | BMP-7 attenuates
pyroptosis (caspase-1, IL-1β, IL-18 and gasdermin-D) | (69) |
| Necroptosis | Hsp90 | Hsp90 inhibitors
may limit RIP1-RIP3-MLKL pathway activation | (93) |
| CML | CML activates RIP3
through its receptor RAGE | (94) |
| RIP3 | RIP3-camkii | (95) |
| Combined use of
Necrostatin-1 (necroptosis inhibitor) and Z-VAD | RIP1-mediated
necrosis | (96) |
| Pulmonary
hypertension | Apoptosis | MFF | Drp1
activation | (41) |
| Notch1 signaling
pathway | Inhibits apoptosis
via Bcl-2 and survivin | (42) |
| PGE1 | HIF pathway | (43) |
| SOX2-OT
(lncRNA) | Modulates the
miR-455-3p/SUMO1 axis | (44) |
| miR-30d-5p | Regulates PASMC
toxicity and apoptosis, potentially through the Notch-3 signaling
pathway | (45) |
| Dec1-PPARγ
axis | Changes in the
Bax/Bcl-2 ratio and cleaved caspase 3 expression | (46) |
| BMPR2 | Loss of functional
BMPR2 signaling leads to increased PAEC apoptosis and cell | (47) |
| proliferation
through augmentation of TGF-β responses | |
| KCNA5 | Decreased apoptosis
(Annexin V-PI) | (48) |
| YM155 | Suppresses PASMC
proliferation and promotes PASMC apoptosis by inhibiting | (49) |
| survivin
expression | |
| Qiliqiangxin | Attenuates RV
myocardial apoptosis (Caspase-3, Bcl-2, BAX) | (50) |
| Pyroptosis | GLI1 | Orchestrates PASMC
pyroptosis by inducing ASC expression, targeting the ASC promoter
region | (70) |
| STAT1/PD-L1 | Induces caspase-1
expression | (72) |
| Caspase-1 | Catalyzes PASMC
proliferation via the caspase-1/IL-18/IL-6/STAT3 pathway | (73) |
| NLRC3 | Downregulates the
production of pro-inflammatory cytokines and induces pyroptosis by
inhibiting the inflammasome | (74) |
| miR-155 | Promotes
inflammation and induces PH through the c-Fos/NLRP3/caspase-1
pathway | (75) |
| Tannic acid | Ameliorates
MCT-induced PH through antioxidative properties by inhibiting the
NLRP3 inflammasome signaling pathway | (76) |
| Necroptosis | TLR & NLR
pathways | Activation of TLR
and NLR pathways is associated with the upregulation of DAMPs.
RIPK3-mediated necroptosis promotes DAMPs generation in MCT-induced
PH | (97) |
| RIP3 | Increased
pthr231/Ser232-RIP3 levels leading to various necrosis-like cell
deaths may contribute to PH pathomechanisms | (98) |
| Influenza
virus | PANoptosis | ZBP1 | ZBP1 recruits RIPK3
and caspase-8 to activate the ZBP1-NLRP3 inflammasome | (102) |
| COVID-19 | PANoptosis | ZBP1 | ZBP1 induced during
coronavirus infection limits the efficacy of IFN therapy by driving
inflammatory cell death and lethality | (103) |
| Microbial
infections, cancers, ALI/ARDS, ischemia-reperfusion and organic
failure | PANoptosis | ZBP1 | These PANoptosomes
further induce caspase-3/7 activation, GSDMD and GSDME cleavage,
and MLKL phosphorylation, resulting in membrane pore formation and
PANoptosis progression | (105-106) |
| Ischemic
stroke | PANoptosis | RIPK1 | RIPK1/RIPK3/MLKL
pathway | (107) |
| Psoriatic
Inflammation | PANoptosis | RIPK1 | RIPK1/RIPK3/MLKL
pathway | (108) |
| Cerebral ischemic
injury | PANoptosis | MLKL | RIPK1/RIPK3/MLKL
pathway | (114) |
| Depression | PANoptosis | MLKL | RIPK1/RIPK3/MLKL
pathway | (115) |
| COVID-19 | PANoptosis | TNF signaling
pathway | The JAK/STAT1/IRF1
axis activated by TNF-α and IFN-γ co-treatment induced iNOS for the
production of nitric oxide. Pharmacological inhibition and genetic
deletion of this pathway inhibited pyroptosis, apoptosis and
necroptosis in macrophages | (116) |
| The innate immune
system | PANoptosis | TLR signaling
pathway | The TLR signaling
pathway may indirectly modulate cell death mechanisms associated
with PANoptosis by activating NF-κb | (118) |
| Neurodegenerative
diseases | PANoptosis | TLR signaling
pathway | Although existing
studies did not establish a direct link between the TLR signaling
pathway and PANoptosis, they underscore the latent role of this
pathway in cellular death and inflammation | (119, 120) |
| Inflammation and
ischemic stroke | PANoptosis | Intracellular
stress pathways | ER stress-autophagy
axis | (121, 122) |
| Myocardial
injury | PANoptosis | Mitochondrial
signaling pathways | FUNDC1 stabilizes
mitochondrial DNA by binding to TUFM, thereby protecting
cardiomyocytes from DOX-induced PANoptosis | (10) |
| Pancreatic ductal
adenocarcinoma | PANoptosis | Mitochondrial
signaling pathways | SEP may enter
cancer cells effectively, then damage nuclear DNA, boost
mitochondrial superoxide anion radicals and affect various
signaling pathways related to redox homeostasis and tumor
metabolism | (124) |
| Sepsis and HLH | PANoptosis | Mitochondrial
signaling pathways | PANoptotic
stimulation induces RET and ROS in mitochondria, while 1-methoxy
PMS and dimethyl fumarate can inhibit PANoptosis by suppressing
RET-mediated oxidation of mitochondrial DNA | (125) |
| Ischemic stroke and
CI/RI | PANoptosis | Mitochondrial
signaling pathways | Exposure to stimuli
during CI/RI can lead to the initiation of the apical sensors, such
as ZBP1, which then induces the activation of proteins involved in
pyroptosis, apoptosis and necroptosis to form the ZBP1-PANoptosome
and mediate PANoptosis | (126) |
Apoptosis, the prototypical PCD pathway, can be
initiated via intrinsic and extrinsic mechanisms. When a
mitochondrion is compromised, the mitochondrial outer membrane
exhibits increased permeability, facilitating the release of
various molecules, including cytochrome C. Cytoplasmic cytochrome C
is sensed by the apoptotic protease activating factor-1, leading to
the formation of the apoptosome with initiator caspase-9. This
mature caspase-9 subsequently activates the downstream effector
caspase-3. In addition, endoplasmic reticulum (ER) stress induces
endogenous apoptosis by activating caspase-12 and caspase-3 in turn
by releasing Ca2+ in the ER cavity. Extrinsic apoptosis
is induced by death receptors such as Fas and TNF-α receptors, with
pro-apoptotic molecules such as Fas-associated protein with death
domain and caspase-8 identified downstream. Consequently, caspase-9
and caspase-8 are initiator caspases for the intrinsic and
extrinsic apoptosis pathways, respectively. These pathways converge
on activating the same effector enzymes, caspase-3 and caspase-7,
culminating in apoptosis (11)
(Fig. 1).
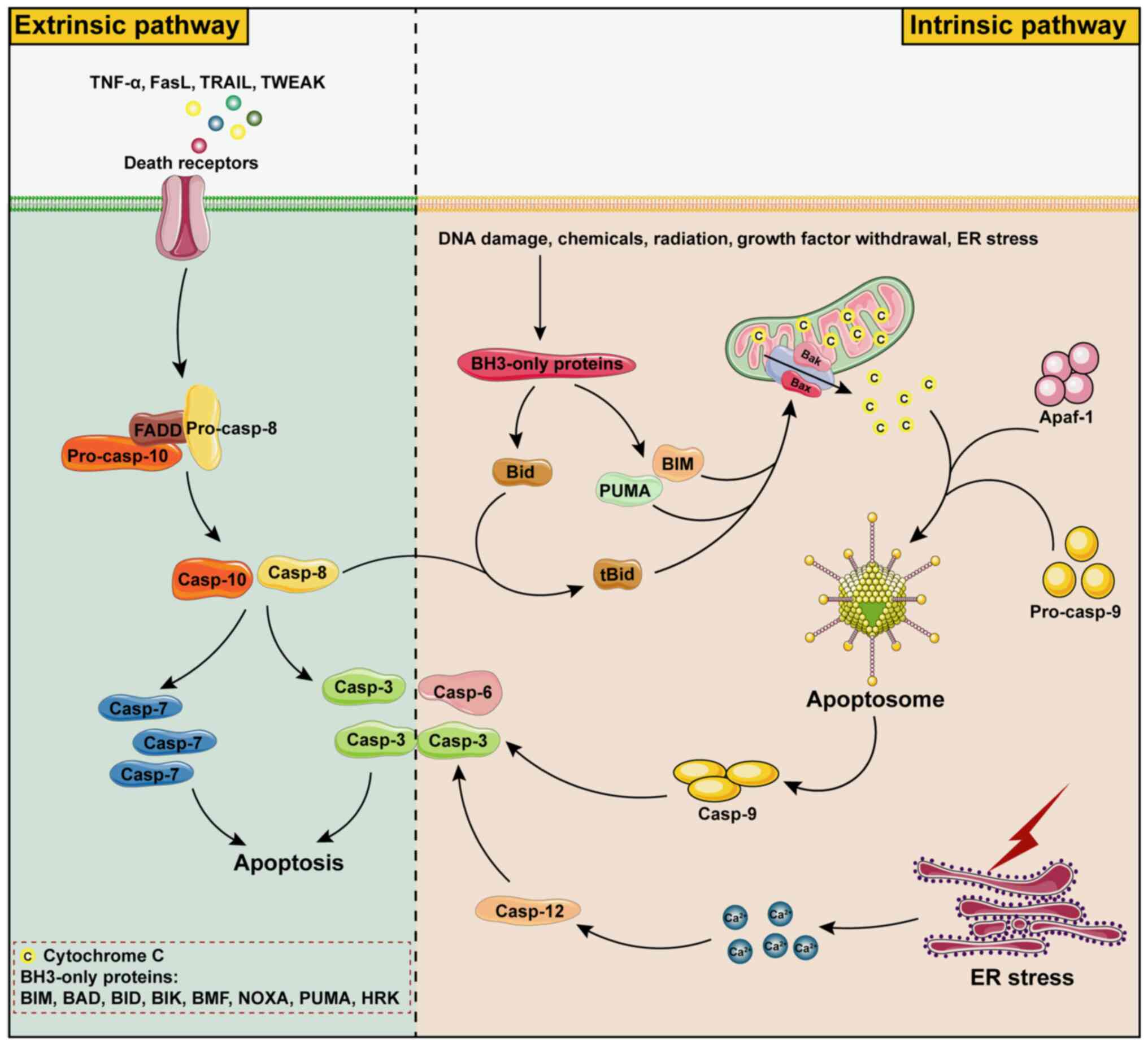 | Figure 1Mechanisms of apoptosis. Apoptosis
can be caused by endogenous and exogenous factors. Exogenous
apoptosis is mainly triggered by death receptors and then FADD
binds to pro-caspase-8 or pro-caspase-10 and induces apoptosis by
activating caspase-8/10 and then caspase-3/7 sequentially.
Endogenous apoptosis is divided into the mitochondrial pathway and
the ER pathway, which is mainly caused by activation of
pro-apoptotic proteins BH3-only proteins by factors such as DNA
damage, chemical stimuli, radiation, growth factor withdrawal and
ER stress, etc. BH3-only proteins mainly include BAD, BID, BIK,
BIM, BMF, HRK, NOXA and PUMA, of which BIM, PUMA and tBID are
potent apoptosis initiators, which form channels in mitochondria by
shearing BAX and BAK proteins. Cytochrome C in mitochondria is
released into the cytoplasm through BAX/BAK channels and binds to
APAF1 and pro-caspase-9 to form the apoptosome, which in turn
causes cascade activation of caspase-9 and caspase-3/6 to induce
apoptosis. In addition, apoptosis can also be induced by promoting
Ca2+ release from the ER after ER stress, which leads to
caspase-12-dependent activation of caspase-3. BH3, BCL-2 homologous
region 3; BAD, BCL-2 associated agonist of cell death; BID,
BH3-interacting domain death agonist; BIK, BCL-2 interacting
killer; BIM, BCL-2 interacting mediator of cell death; BMF, BCL-2
modifying factor; HRK, Harakiri; NOXA,
phorbol-12-myristate-13-acetate induced protein 1; PUMA, p53
upregulated modulator of apoptosis; APAF1, apoptotic peptidase
activator 1; ER, endoplasmic reticulum; Casp, caspase; TNF, tumor
necrosis factor; TLR, Toll-like receptor; FADD, Fas-associated
death domain; FASL, factor-related apoptosis ligand; TRAIL, tumor
necrosis factor-related apoptosis-inducing ligand; TWEAK, tumor
necrosis factor-like weak inducer of apoptosis. |
Pyroptosis, a caspase-1-dependent form of PCD, is
activated by the Nod-like receptor (NLR) family pyrin domain
containing 3 (NLRP3) inflammasome, comprising a sensor protein, the
adaptor protein apoptosis-associated speck-like protein containing
CARD (ASC) and caspase-1. Caspase-1 activation cleaves its
substrates, inducing the maturation of inflammatory cytokines
pro-IL-1β and pro-IL-18. The understanding of pyroptosis has
evolved with the discovery of various inflammasomes that activate
different inflammatory caspases (caspase-1, -4, -5 and -11)
(15). Recent studies have
broadened this understanding by identifying additional mechanisms
that lead to pyroptosis. These include the non-canonical
inflammasome pathway, which involves caspase-1-independent cleavage
of the protein gasdermin D (GSDMD), as well as the cleavage of
GSDMD by caspase-8. In addition, the cleavage of GSDMA by
Streptococcal pyrogenic exotoxin B (SpeB) protease, the cleavage of
GSDMB by caspase-3/6/7 or Granzyme A, the cleavage of GSDMC by
caspase-8 and the cleavage of GSDME by caspase-3, have been
recognized, expanding the definition of pyroptosis beyond the
traditional caspase-1-dependent pathway (16-18) (Fig. 2).
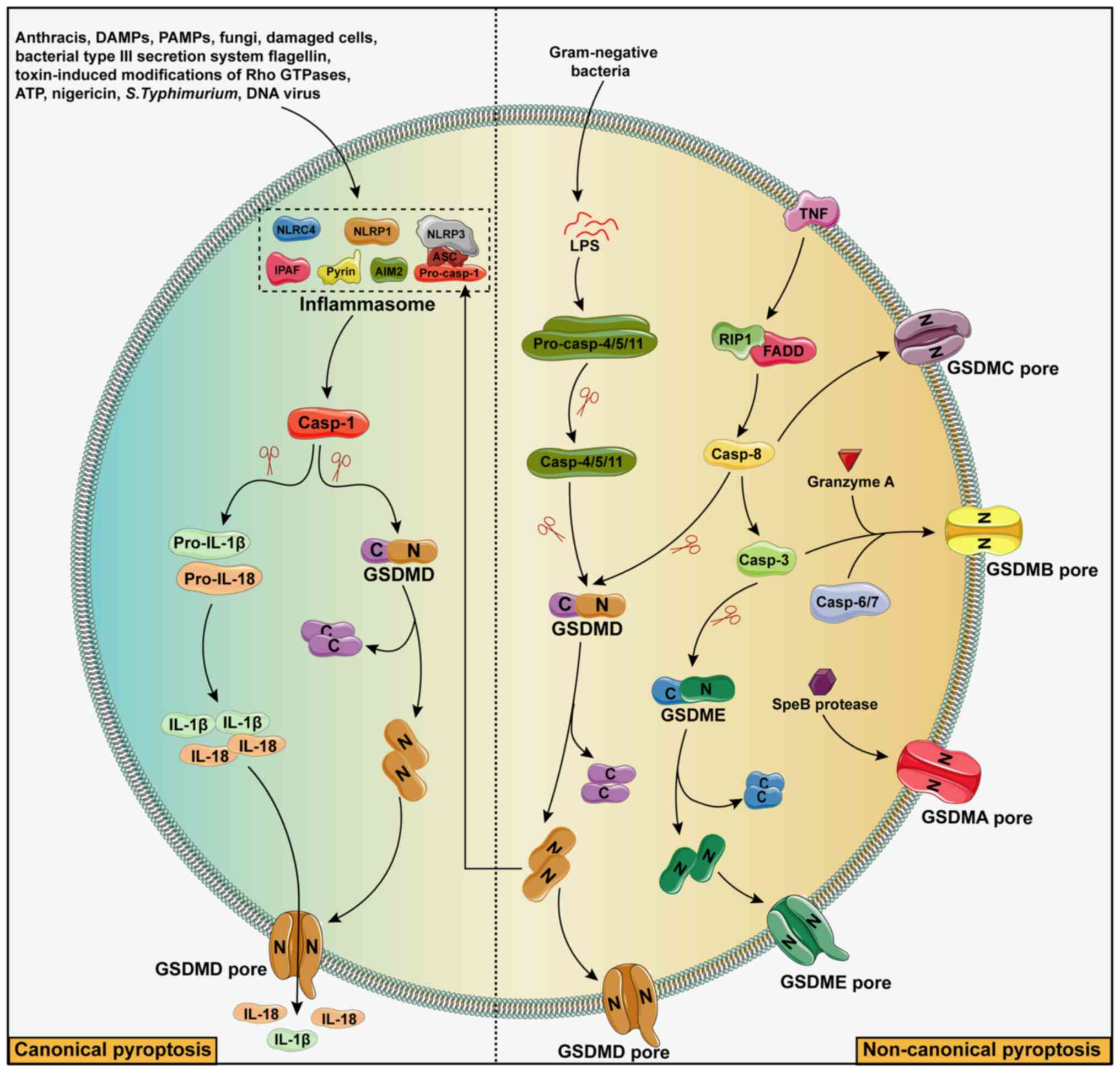 | Figure 2Mechanisms of pyroptosis. Depending
on the type of stressor, pyroptosis can be induced by canonical or
non-canonical pathways. Stressors that activate the classical
pathway include Anthracis, PAMPs, DAMPs, fungi, damaged cells,
bacterial type III secretion system flagellin, toxin-induced
modifications of Rho GTPases, ATP, nigericin, S.
Typhimurium, double-stranded DNA viruses, etc., and the
stressors that activate the non-canonical pathway are mainly
gram-negative bacteria. Inflammasomes that activate the canonical
pathway mainly include six types, namely NLRP1-inflammasome,
NLRP3-inflammasome, NLRC4-inflammasome, IPAF-inflammasome,
AIM2-inflammasome and pyrin-inflammasome. After the assembly of the
inflammasome, caspase-1 can be activated. The activated caspase-1
can shear the GSDMD to obtain N-terminal GSDMD with active domain
peptide on the one hand, inducing cell membrane perforation, then
cell swelling and rupture, and releasing the contents to cause
inflammation. On the other hand, activated caspase-1 cuts the
precursors of IL-1β and IL-18 to form active IL-1β and IL-18, which
are released outside the cell to recruit inflammatory cells and
further amplify the inflammatory response. For the non-canonical
pathway of cellular pyroptosis, signals such as gram-negative
bacteria, granzyme A and SpeB protease induce cell membrane
perforation and ultimately cellular pyroptosis by activating
caspase-3/4/5/6/7/8/11 through the formation of active N-terminal
peptides such as GSDMA, GSDMB, GSDMC, GSDMD and GSDME. Casp,
caspase; PAMP, pathogen-associated molecular pattern; DAMP,
damage-associated molecular pattern; AIM2, absent in melanoma 2;
NLRP1, nucleotide-binding domain-containing, leucine-rich
repeat-containing and pyrin domain-containing protein 1; IPAF,
ICE-protease activating factor; ASC, adapter protein
apoptosis-associated speck-like protein containing a caspase
recruitment domain; NLRC4, NLR-family CARD-containing protein 4;
GSDM, gasdermin; LPS, lipopolysaccharide; RIP1,
receptor-interacting serine/threonine protein kinase 1; pro-IL-1β,
pro-interleukin-1β; pro-IL-18, pro-interleukin-18; ATP, adenosine
triphosphate. |
Necroptosis, an adjustable form of cell necrosis
that operates independently of caspases (non-caspase-dependent),
shares several characteristics with both apoptosis and necrosis.
This process is predominantly characterized by notable cell
swelling and the compromise of cellular membrane integrity, which
may precipitate an inflammatory response (19). Necroptosis is initiated by death
receptors and is triggered by ligands attached to these receptors.
Crucial molecules in the necroptosis pathway, particularly those
participating in the tumor necrosis factor (TNF)-α/TNF
receptor-induced pathway, include RIP1, RIP3 and MLKL. This
RIP1-RIP3-MLKL cascade is recognized as the classical pathway of
necroptosis. Various drugs and compounds effectively inhibit this
programmed necrosis by targeting key necroptotic molecules such as
RIP1, RIP3 and MLKL (20).
Elucidating the molecular mechanisms underlying cardiomyocyte
necroptosis and its implications in the pathophysiology of CVDs
holds substantial clinical significance. In-depth research into
necroptosis may reveal novel interventional strategies and
therapeutic targets, thereby potentially lowering CVD-associated
mortality rates and improving the prognosis of patients (Fig. 3).
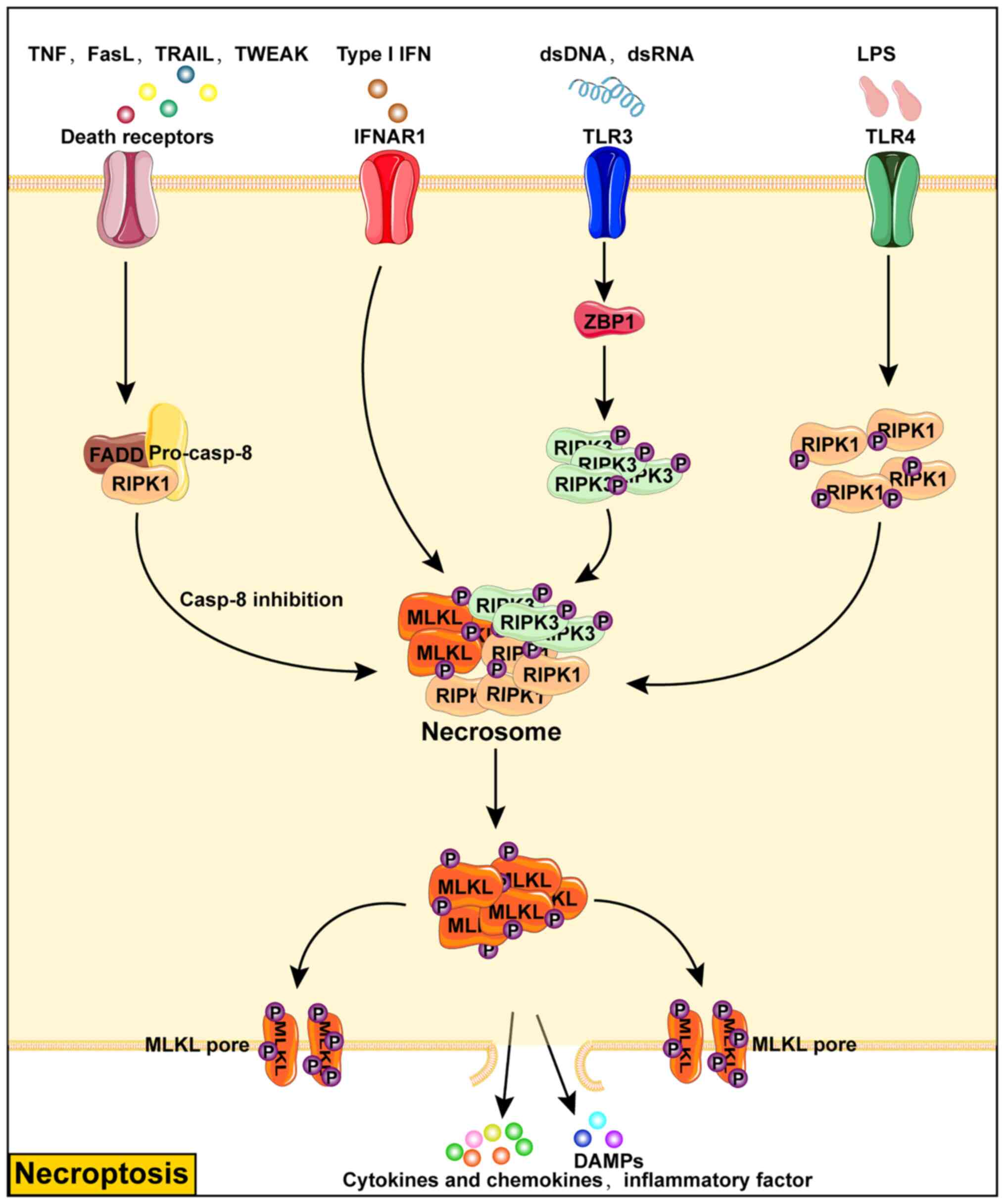 | Figure 3Mechanisms of necroptosis.
Necroptosis is a mode of programmed cell death that is not
dependent on caspase activity and is caused by phosphorylated MLKL
punching holes in the cell membrane causing the release of
intracellular associated antigens, DAMP and ultimately cell lysis
and death. Necrotic apoptosis can be triggered by the stimulation
of death receptors, IFNAR1, TLR3, TLR4 and other receptors. In
particular, upon death receptor stimulation, necroptosis can be
induced only if caspase-8 activity is inhibited, which promotes the
assembly of necrosome through the binding of FADD to RIPK1 and
further leads to the phosphorylation and activation of the
necroptosis executor MLKL. The ligand type I IFN promotes the
assembly of necrosome by activating IFNAR1 to cause necroptosis.
Endogenous or exogenous dsDNA and dsRNA activate ZBP1 by TLR3
receptor and cause phosphorylation of RIPK3 to promote necrosome
assembly. By contrast, LPS mediates the assembly of necrosome
through TLR4 receptor, which in turn promotes the phosphorylation
of RIPK1. IFN, type I interferon; IFNAR1, IFN receptor; RIPK1,
receptor-interacting serine/threonine protein kinase 1; RIPK3,
receptor-interacting protein kinase 3; TLR, Toll-like receptor;
LPS, lipopolysaccharide; DAMP, damage-associated molecular pattern;
MLKL, mixed lineage kinase domain-like protein; ZBP1, Z-DNA binding
protein 1; dsDNA, double-stranded DNA; dsRNA, double-stranded
RNA. |
PCD is integral to organismal development,
maintaining homeostasis, and a critical component of innate immune
responses. However, over-activation of PCD pathways can be
deleterious, contributing to acute injuries and chronic
degenerative diseases (19). Its
involvement is noted in conditions such as acute spinal cord
injuries (21) and chronic
degenerative diseases such as AS and vascular calcification
(22). Despite the diversity of
PCD types, the CVD-associated signaling pathways function as a
coordinated system with highly interconnected pathways capable of
mutual compensation (23).
Apoptosis, pyroptosis and necroptosis represent distinct PCD
pathways, intricately interwoven in determining the cell fate and
governing physiological processes, thus establishing a dynamic
equilibrium system. These three pathways are activated in various
cell death mechanisms in response to various external and internal
cues. Apoptosis is the primary mechanism for maintaining tissue
homeostasis and eliminating damaged cells, while pyroptosis is
closely associated with immune regulation and the orchestration of
inflammatory responses. By contrast, necroptosis signifies a
non-apoptotic and non-pyroptotic mode of cell death, which can
replace apoptosis or pyroptosis in specific circumstances. Despite
their unique characteristics, these pathways interact, adapting to
cellular demands in a range of environmental contexts, thereby
conferring upon the biological organism the versatility and
adaptability required for determining cell fate.
Apoptosis and its relationship with
CVDs
Cardiomyocytes, characterized as terminally
differentiated cells, have traditionally been understood to
exclusively succumb to necrosis. This long-held perspective,
however, has been challenged by emerging debates over the potential
role of apoptosis in these cells. Significantly, apoptosis is not
merely associated with cardiac pathologies; it is a pivotal process
in the broader spectrum of an organism's vital functions. Essential
for preserving both the physiological integrity and structural
morphology of the body, apoptosis must be carefully balanced.
Deviations, manifesting as excessive or deficient apoptotic
activity, can result in a spectrum of morphological and functional
aberrations within the organism. Research has highlighted the role
of cardiomyocyte apoptosis in a range of CVDs, including AS, MI, HF
and pulmonary hypertension (PH), underscoring the importance of
apoptosis in both the pathogenesis and progression of CVDs.
Role of apoptosis in AS
In the early stages of AS development, the apoptosis
of vascular endothelial cells (VECs) induced by oxidized
low-density lipoprotein (ox-LDL) leads to endothelial dysfunction,
a critical factor in the formation of AS. This results in the
accumulation of VECs, vascular smooth muscle cells (VSMCs),
macrophages and their derivative foam cells within the vessel, and
thus the formation of atherosclerotic plaques (24,25). Research has shown that ox-LDL
induces VEC apoptosis by activating the Fas/FasL pathway (26). At the same time, hyperglycemic
conditions trigger the production of inflammatory cytokines and
initiate a caspase cascade through the activation of the NF-κB
pathway, exacerbating VEC apoptosis (27). In addition, the inhibitory effect
of Paeonol on apoptosis in VEC induced by high glucose/high
pressure is mediated through the regulation of the
SIRT1/FOXO3a/NF-κB pathway (28). The combination of geniposide and
notoginsenoside R1 has been shown to effectively inhibit
inflammation and apoptosis in AS. This effect is mediated through
the activation of the adenosine 5'-monophosphate activated protein
kinase/mechanistic target of rapamycin (mTOR)/nuclear factor
erythroid 2-related factor 2 signaling pathway, which subsequently
inhibits the NLRP3 inflammasome and the Bax/Bcl2/caspase-3 pathway,
which are pivotal in regulating apoptosis within atherosclerotic
plaques (29).
VSMC apoptosis is crucial in the formation and
progression of atherosclerotic plaques, particularly during the
later stages, where ongoing apoptosis can thin the fibrous cap,
increasing plaque instability (30). Furthermore, the role of
macrophage colony-stimulating factor (CSF) in regulating macrophage
proliferation and apoptosis has been highlighted. Local production
of CSF1 by smooth muscle cells and endothelial cells, rather than
circulating CSF1, is identified as the primary driver of macrophage
expansion in atherosclerotic lesions. This finding underscores the
significance of local cellular interactions in modulating apoptosis
within the plaque microenvironment (31).
Apoptosis and MI
MI, a leading cause of mortality worldwide, is
primarily characterized by the death of cardiac tissue due to
ischemia. A critical aspect of MI pathophysiology is apoptosis, or
PCD, which has a pivotal role in the progression of cardiac damage
and the subsequent remodeling of the myocardium. Understanding the
mechanisms and implications of apoptosis in MI is crucial for
developing targeted therapeutic strategies. The present review
elucidates the current understanding of apoptosis in MI,
referencing key research findings.
Apoptosis in cardiac cells post-MI is a double-edged
sword. While it eliminates damaged cells, excessive apoptosis can
lead to the detrimental loss of functional myocardium, contributing
to HF. Recent studies have highlighted various molecular pathways
and factors involved in regulating apoptosis during MI. For
instance, the role of microRNA (miR)-182-5p in apoptosis regulation
during MI has been explored, offering insight into potential
therapeutic targets (32).
In addition, the insulin-like growth factor-1
mediated enhancement of bone marrow stem cell viability and
anti-apoptotic effects in MI through the secreted frizzled-related
protein 2 pathway has been demonstrated, suggesting a novel
approach to stem cell therapy in cardiac repair (33). Furthermore, the role of miR-338
in inhibiting cardiomyocyte apoptosis in MI through the MAP3K2/JNK
signaling pathway has been identified, highlighting the therapeutic
potential of miRNA-based interventions (34).
Long non-coding (lnc)RNA Maternally expressed gene 3
has been found to regulate cardiomyocyte apoptosis post-MI,
suggesting a novel avenue for gene therapy (35). Furthermore, miR-146b has been
shown to mediate vascular inflammation and apoptosis in patients
who have experienced MI, potentially via the PI3K/Akt/NF-κB
signaling pathway (36).
Finally, the novel non-coding transcriptional regulator Gm18840 has
been identified as a promoter of myocardial cell apoptosis in MI,
highlighting novel potential targets for therapeutic intervention
(37).
Thus, apoptosis has a multifaceted role in the
pathogenesis of MI. Understanding the mechanisms underlying
apoptosis in MI can provide insight into potential therapeutic
strategies for this debilitating disease.
Apoptosis and HF
HF is a complex clinical syndrome characterized by a
reduced cardiac output, originating from alterations in myocardial
energy metabolism, excitation-contraction coupling and cardiac
structural changes stemming from various etiologies. This decline
in cardiac output triggers several compensatory mechanisms, both
intrinsically within the heart and systemically. These include an
increased heart rate, enhanced myocardial tension, augmented
myocardial contraction and significant ventricular remodeling.
However, prolonged activation of these compensatory responses
eventually leads to progressive ventricular remodeling,
exacerbating HF.
Apoptosis of cardiac cells contributes to the
deterioration of cardiac function in HF. Various molecular pathways
and external stimuli influence the regulation of apoptosis in
cardiomyocytes. For instance, miR-182 has been identified as a
regulator that inhibits cardiomyocyte apoptosis induced by
non-ischemic HF, highlighting the therapeutic potential of
targeting specific miRNAs in the treatment of HF (38). In addition, oxidative
stress-induced myocardial apoptosis is ameliorated by Baiyangdian
via the P38 MAPK-αB crystallin signaling pathway, highlighting a
novel target for the management of acute MI (39). The Shenfu formula has been found
to reduce cardiomyocyte apoptosis in a rat model of HF, suggesting
the potential of Traditional Chinese Medicines in managing HF
(40).
Apoptosis and PH
PH is increasingly being recognized as a progressive
and debilitating condition, characterized by the constrictive
remodeling of pulmonary arteries, such as the abnormal
proliferation of pulmonary arterial smooth muscle cells (PASMCs)
and apoptosis of endothelial cells (PAECs). This pathological
transformation exacerbates pulmonary vascular resistance,
precipitating right HF and ultimately mortality. The pathogenesis
of PH is complex, encompassing a multifaceted interplay of genetic,
molecular and environmental factors. Of note, the dysregulation of
apoptosis has been identified as a pivotal factor in the
progression of PH. Recent research has illuminated the molecular
underpinnings of dysregulation of apoptosis in PH. The
mitochondrial fission factor has emerged as a crucial element, with
its midzone fission process being increased in PH-affected PASMCs.
This increase results in increased cellular proliferation and
reduced apoptosis, signifying a potential target for therapeutic
intervention (41). In addition,
the Notch1 signaling pathway has been shown to modulate endothelial
proliferation and apoptosis in PH, highlighting Notch inhibitors as
a prospective therapeutic avenue (42).
The synergy between apoptosis and other cellular
processes, including inflammation, migration and metabolic
reprogramming, in PH has also attracted attention. Prostaglandin
E1, for example, has been shown to influence mesenchymal stem cell
properties via the hypoxia-inducible factor (HIF) pathway, thereby
impacting apoptosis in PH (43).
Furthermore, the long non-coding (lnc)RNA SOX2-OT, can positively
promote the transcription of SOX2 gene (one of the major regulators
of pluripotency), has been highlighted as a potential diagnostic
biomarker for PH, affecting PASMC proliferation, migration,
anti-apoptotic activities and inflammation (44). miRNAs have garnered recognition
for their pivotal role in regulating apoptosis in PH. Specifically,
the miR-30 family, and more notably miR-30d-5p, were identified as
regulators of PASMC toxicity and apoptosis, potentially via the
Notch-3 signaling pathway. This regulatory role is significant, as
evidenced in both blood samples from patients with PH and animal
models, in contrast to control groups (45). The differentiated embryo
chondrocyte expressed gene 1-peroxisome proliferative-activated
receptor-γ axis has also been delineated as a critical element in
hypoxia-modulated signaling, integral to the imbalance between
proliferation and apoptosis in PAECs. This finding underscores the
complexity of the cellular milieu in PH and the nuanced equilibrium
required for vascular homeostasis (46).
Genetic influences are also of paramount importance
in the dysregulation of apoptosis observed in PH. Mutations in bone
morphogenetic protein receptor 2, have been associated with
metabolic aberrations and dysregulated signaling pathways,
influencing apoptosis and contributing to PH pathogenesis (47). Furthermore, newly identified
loss-of-function variants in potassium voltage-gated channel
subfamily A member 5 in patients with PH suggest a potential
causative or contributing role through altered channel
functionality (48). Emerging
therapeutic strategies targeting apoptosis have shown potential in
PH management. For instance, The survivin inhibitor YM155 has
demonstrated efficacy in suppressing PASMC proliferation and
promoting apoptosis by inhibiting survivin expression, thereby
mitigating pulmonary vascular remodeling in PH (49). Similarly, Qiliqiangxin has been
shown to inhibit PH-induced right ventricular remodeling by
decreasing mitochondrial-associated apoptotic pathways and
improving metabolic reprogramming (50).
In summary, the role of apoptosis in PH is
multifaceted and critical to its pathogenesis. Delving into the
mechanisms of apoptosis in PH may highlight novel therapeutic
targets for the management of PH. The current research landscape
presents exciting opportunities for further investigation and the
development of targeted interventions.
Pyroptosis and its relationship with
CVDs
Pyroptosis represents a distinct, pro-inflammatory
paradigm of PCD, characterized by an intricate amalgamation of
morphological changes reminiscent of both apoptosis and necrosis.
Central to this process is the formation of pores within the
cytoplasmic membrane, precipitating acute cellular swelling,
rupture and subsequent discharge of pro-inflammatory agents and
intracellular components. Crucially, pyroptosis exerts a profound
influence on the onset and progression of a range of pathologies,
including AS, MI, HF and PH. This influence is primarily exerted
through the modulation of inflammatory responses and cell death.
Furthermore, the molecular entities associated with pyroptosis have
a significant role in dictating the CVD trajectory and clinical
outcomes, underscoring their potential as therapeutic targets.
Pyroptosis and AS
Pyroptosis, a pro-inflammatory form of cell death,
has a pivotal role in the pathogenesis of AS. Pyroptosis
significantly exacerbates the instability of atherosclerotic
plaques, leading to plaque rupture and thrombus formation. This
process is a crucial trigger for acute cardiovascular events,
primarily through the extensive release of pro-inflammatory
cytokines (51).
Endothelial cells are at the forefront of AS
development and progression, serving as a critical determinant in
its pathogenesis. Several risk factors, such as hyperlipidemia,
hyperglycemia, hypertension, smoking and inflammation, precipitate
endothelial cell pyroptosis via the activation of the caspase-1
pathway. This activation not only triggers the release of
pro-inflammatory cytokines but also compromises the structural
integrity of the cell membrane. A study by Wu et al
(52) underscored the
aggravating effect of nicotine on arterial atherosclerotic lesions
in apolipoprotein E (ApoE)−/− mice, concomitantly
elevating caspase-1 expression. Furthermore, trimethylamine N-oxide
(TMAO), a byproduct of the phosphatidylcholine metabolism from the
gut microbiota, exacerbates endothelial dysfunction by interacting
with the NF-κB and MAPK signaling pathways, thus promoting the
accrual of AS plaques (53).
Under a high-fat diet, ApoE−/− mice exhibit a pronounced
oxidative stress response in vascular endothelial cells attributed
to TMAO, which also upregulates caspase-1 and NLRP3 expression,
thereby promoting AS progression (54).
Macrophage-derived foam cells significantly
contribute to the instability of atherosclerotic plaques (55). The ROS/NF-κB/NLRP3 axis and
mitochondria have crucial roles in macrophage pyroptosis during AS
(56,57). The non-classical caspase-11/GSDMD
pyroptosis pathway has also been associated with AS. In addition,
the observed reduction in the area of atherosclerotic plaques in
ApoE and GSDME double-knockout mice under a high-fat diet
underscores the significance of GSDME within macrophages (58).
VSMCs are also pivotal in AS development. VSMC
apoptosis not only reduces the content of extracellular matrix
(ECM), rendering the fibrous cap susceptible to rupture, but it may
also intensify inflammation within the vascular wall, thereby
elevating the risk of plaque instability. It has been shown that
ox-LDL accumulation is intricately associated with VSMC apoptosis.
Intriguingly, while low concentrations of ox-LDL promote a
phenotypic shift in these cells from contractile to synthetic,
facilitating the secretion of inflammatory cytokines; higher
concentrations of ox-LDL are implicated in directly inducing
apoptosis (59). Subsequent
studies have further delineated the profound impact of ox-LDL,
revealing its capacity to transform VSMCs into foam cells and
increase the expression of NLRP3, ASC, caspase-1 and GSDMD. This
underscores the mechanism by which ox-LDL, via the activation of
the caspase-1-dependent pathway, promotes VSMC apoptosis and the
subsequent release of IL-1β and IL-18. Such events contribute to
the diminution of the ECM, the attenuation of the fibrous cap, the
amplification of local inflammatory responses and the promotion of
AS (60). The caspase-1
inhibitor VX-765 significantly reduces ox-LDL-induced endothelial
cell pyroptosis, thereby slowing the progression of AS (61).
Pyroptosis and MI
The key complication of MI therapy is myocardial
ischemia/reperfusion injury (MI/RI). A growing body of evidence
indicates a strong association between cell pyroptosis and MI/RI.
The crucial role of NLRP3 and caspase-1-mediated cell pyroptosis in
MI/RI is highlighted below.
The silencing of calpain can mitigate myocardial
dysfunction caused by MI/RI, primarily through the inhibition of
the NLRP3/ASC/caspase-1 axis (62). In addition, targeting oxidative
stress and NLRP3-mediated cell pyroptosis by miR-29a-targeted
silencing of sirtuin 1 has also been demonstrated to alleviate
MI/RI (63). Further research
has established that mice with caspase-1 overexpression show an
increased MI area (64).
Conversely, VX-765, a selective caspase-1 inhibitor, effectively
reduces MI in MI/RI injury models (65).
Pyroptosis and HF
HF represents the terminal stage in the progression
of CVDs, characterized by a complex pathology that encompasses
myocardial fibrosis, hypertrophy and excessive inflammation, among
which pyroptosis has a pivotal role. This phenomenon profoundly
impacts cardiac structure and function and offers novel
perspectives for therapeutic strategies against HF.
Myocardial fibrosis, the thickening of the
myocardial interstitium due to excessive deposition of ECM
proteins, is primarily driven by activated fibroblasts, the main
producers of ECM proteins. Zhang et al (66) discovered that the activation of
the NLRP3 inflammasome in fibroblasts promotes the synthesis of
collagen and activates caspase-1, leading to the release of IL-1β
and IL-18, thereby inducing pyroptosis. Furthermore, IL-1β and
IL-18 can exacerbate HF progression by activating the release of
TNF-α and promoting the efflux of Ca2+ from the
sarcoplasmic reticulum. Myocardial hypertrophy is crucial for
maintaining cardiac ejection function. However, pathological
myocardial hypertrophy induced by disease can precipitate HF. Zeng
et al (67) reported that
in patients with dilated cardiomyopathy, NLRP3 inflammasome
activation is increased, accompanied by myocardial cell pyroptosis,
and this is inversely correlated with cardiac ejection function.
These findings underscore the role of the NLRP3 inflammasome,
primarily through IL-1β, in promoting structural cardiomyopathic
changes associated with myocardial dysfunction. This is further
validated by the fact that caspase-1 modulates Ang II-induced
cardiac hypertrophy via the regulation of IL-1β (68). Conversely, the anti-inflammatory
role of bone morphogenetic protein-7 partially alleviates
myocardial hypertrophy and fibrosis, highlighting its potential
therapeutic value in managing HF (69).
Pyroptosis and PH
PH is a CVD closely associated with structural
abnormalities and functional dysregulation of the pulmonary
vasculature. The progression of this disease is influenced by
genetic and environmental factors, involving numerous vasoactive
molecules and signaling pathways. Damage to pulmonary arterial
endothelial and smooth muscle cells leads to abnormal
vasoconstriction, remodeling, thrombosis, inflammatory responses
and aberrant proliferation and apoptosis of vessels, thereby
triggering PH.
Under hypoxic conditions, the relationship between
cell pyroptosis and the formation of PH has become increasingly
apparent. Research indicates that the activation of Gli zinc finger
transcription factor 1 (GLI1) promotes cell pyroptosis in PASMCs
under hypoxia. GLI1 orchestrates PASMC pyroptosis by inducing ASC
expression, targeting the ASC promoter region. Inhibition of GLI1
reverses in vivo PH symptoms and pyroptosis (70). Furthermore, a study by Cero et
al (71) found that mice
lacking ASC exhibited significantly reduced symptoms of PH and
right ventricular remodeling under hypoxic conditions, suggesting
that the activation of inflammasomes has a crucial role in the
development of PH. Further investigation revealed that signal
transducer and activator of transcription (STAT1) facilitated PD-L1
upregulation in hypoxia-induced PH models, initiating
caspase-1-dependent pyroptosis in PASMCs, accelerating pulmonary
vascular fibrosis and ultimately leading to PH (72). Udjus et al (73) employed a caspase-1 knockout mouse
model and caspase-1 inhibitors to demonstrate that in
hypoxia-induced PH models, caspase-1 increased PASMC proliferation
via the caspase-1/IL-18/IL-6/STAT3 pathway. Additional evidence
from Zha et al (74)
showed a significant reduction of NLRC3 in patients with PH,
proposing it as a potential diagnostic biomarker and prognostic
indicator. Chai et al (75) found that increased expression of
miR-155, which promotes inflammation and induces PH through the
c-Fos/NLRP3/caspase-1 pathway, was observed in monocrotaline
(MCT)-induced pulmonary arterial hypertension (PAH) mouse models.
Inhibition of c-Fos and NLRP3 mitigated the inflammatory impact of
miR-155 in these models. Tang et al (76) demonstrated that tannic acid
ameliorated MCT-induced PAH via its antioxidative properties
through the inhibition of the NLRP3 inflammasome signaling pathway
in a rat model.
Necroptosis and its relationship with
CVDs
The preceding chapters highlighted that PCD, an
inherently genetic and systematic form of active cell death,
consists of processes such as apoptosis, pyroptosis and
necroptosis. Of note, necroptosis has a pivotal role in the
etiology of a spectrum of CVDs, including but not limited to AS,
MI, HF and PH. In recent years, there has been a notable surge in
research focusing on the intricate signaling pathways and molecular
mechanisms that underpin necroptosis. These studies have provided
profound insight into the complex interplay of factors governing
this form of cell death, offering novel perspectives on potential
therapeutic strategies.
Necroptosis and AS
Necroptosis has a pivotal role in the progression of
AS, with its hallmark proteins RIP3 and MLKL being empirically
demonstrated in human carotid atherosclerotic plaque. Studies have
shown that, while knockout of RIP3 in an ApoE−/− mouse
model had minimal impact on early-stage AS, it significantly
alleviated late-stage atherosclerotic lesions, revealing the
connection between necroptosis and plaque stability (77). Furthermore, elevated plasma
levels of RIP3 are positively associated with the severity of
coronary artery disease, suggesting its potential as an indicator
for assessing the severity of coronary artery disease (78).
The death of macrophages, particularly necroptosis
driven by ox-LDL, is crucial for forming the necrotic core. This
process releases inflammatory cytokines such as IL-6 and IL-1α,
triggering further inflammatory responses (79-81). Evidence indicates that the
absence of MLKL can reduce macrophage necroptosis and the necrotic
core within plaques, highlighting the pivotal role of MLKL in the
development of AS (82). In
addition, exacerbating oxidative stress promotes this process,
activating HIF-1α and intensifying macrophage necroptosis through
enhanced ROS production (83).
Necroptosis and MI
MI, the most severe manifestation of acute coronary
syndrome, has seen a significant improvement in patient survival
rates through interventional treatments. However, efficiently
addressing cardiac remodeling and dysfunction resulting from the
loss of a substantial number of myocardial cells remains a
significant challenge in its treatment.
The increase in RIP3 protein expression is closely
associated with MI. Mice lacking RIP3, following ligation of the
left anterior descending coronary artery, exhibit improved cardiac
function and reduced myocardial hypertrophy, highlighting the
central role of necroptosis in MI and the subsequent cardiac
remodeling (84). Research by
Yang et al (85) showed
that both MLKL and Ca2+/calmodulin-dependent protein
kinase II (CaMKII) were involved in the pathogenesis of chronic
chest pain associated with myocardial ischemia induced by RIP3,
positing that MI/RI could be co-mediated by the RIP1-RIP3-MLKL and
RIP3-CaMKII pathways. TGF-β-activated kinase 1 and TNF
receptor-associated factor 2 have been shown to counteract
necroptosis and regulate cardiac remodeling, offering novel
potential targets for treating ventricular remodeling and HF
(86,87). Studies have shown that S-allyl
cysteine sulfoxide, found in garlic, and miR-325-3p can alleviate
cardiac damage post-MI by inhibiting necroptosis (88,89). In addition, recombinant
adenovirus hepatocyte growth factor improved cardiac remodeling by
promoting autophagy and necroptosis and inhibiting apoptosis
(90), presenting a novel avenue
for intervention in cardiac remodeling post-MI via the regulation
of cell death processes.
Necroptosis and HF
Necroptosis has been identified as a critical factor
in the progression of HF and has a substantial role in ventricular
remodeling. Early research focused on necrosis and apoptosis as the
primary mechanisms of MI/RI, while recent findings have highlighted
the critical role of necroptosis in cell death triggered by MI/RI
(91). The groundbreaking study
by Szobi et al (92)
revealed that in samples from patients with HF, pivotal necroptosis
mediators such as RIP1, RIP3 and MLKL, alongside their
phosphorylated alternatives, were notably upregulated compared to
those in healthy individuals. This is accompanied by a marked
decrease in the expression of activated caspase-8, suggesting the
potential suppression of apoptosis in the milieu of HF, thereby
paving the way for necroptosis.
In addition, the post-TAC elevation in RIP1, RIP3,
MLKL, and their phosphorylated counterparts can be effectively
reversed by applying inhibitors targeting the molecular chaperone
heat shock protein (HSP). These inhibitors also significantly
reduce TNF-α-induced necroptosis by disrupting the RIP1-RIP3-MLKL
interaction. This finding posits that inhibition of HSP90 could
yield therapeutic benefits by reducing the activation of the
RIP1-RIP3-MLKL pathway in compromised cardiac tissue (93). Research has also revealed that
carboxymethyllysine, a late glycation end product derived from
neutrophils, activates RIP3 through its receptor RAGE, leading to
necroptotic death of myocardial cells (94). Furthermore, RIP3-activated CaMKII
promotes cell death, highlighting novel therapeutic targets for
treating cardiac injury and HF (95). The combined use of Nec-1 and a
caspase inhibitor, zVAD-FMK, demonstrated the occurrence of
multiple forms of cell death during MI/RI, providing novel
therapeutic avenues for the recovery of cardiac function (96).
Necroptosis and PH
Xiao et al (97) demonstrated that inflammation and
immunity are critical factors in developing pulmonary vascular
remodeling and PH. They discovered the enrichment of TLR and NLR
pathways and NF-κB-mediated inflammatory and immune profiling in
MCT-induced PAH. Furthermore, they found that the activation of TLR
and NLR pathways was associated with the upregulation of
damage-associated molecular patterns (DAMPs) and that
RIPK3-mediated necroptosis contributed to DAMP generation in
MCT-induced PAH.
A study by Jarabicová et al (98) investigated the role of cell loss
due to necroptosis and its association with pyroptosis in organ
damage during PH. They found increased levels of
pThr231/Ser232-RIP3, leading to various necrosis-like cell death
types, which may have contributed to the pathological mechanisms
occurring during PH. In addition, plasma RIP3 may serve as a novel
diagnostic and prognostic marker for cardiac injury in PH.
PANoptosis
PANoptosis, an acronym embodying pyroptosis,
apoptosis and necroptosis, is a unique PCD paradigm within innate
immunity elicited by innate immune triggers. This complex process
involves the PANoptosome, a sophisticated multi-protein assembly
adept at amalgamating elements from various PCD modalities. The
biological ramifications of PANoptosis are characterized by an
interplay and regulatory overlap among pyroptosis, apoptosis and
necroptosis, surpassing the specific limitations of each pathway
(Fig. 4).
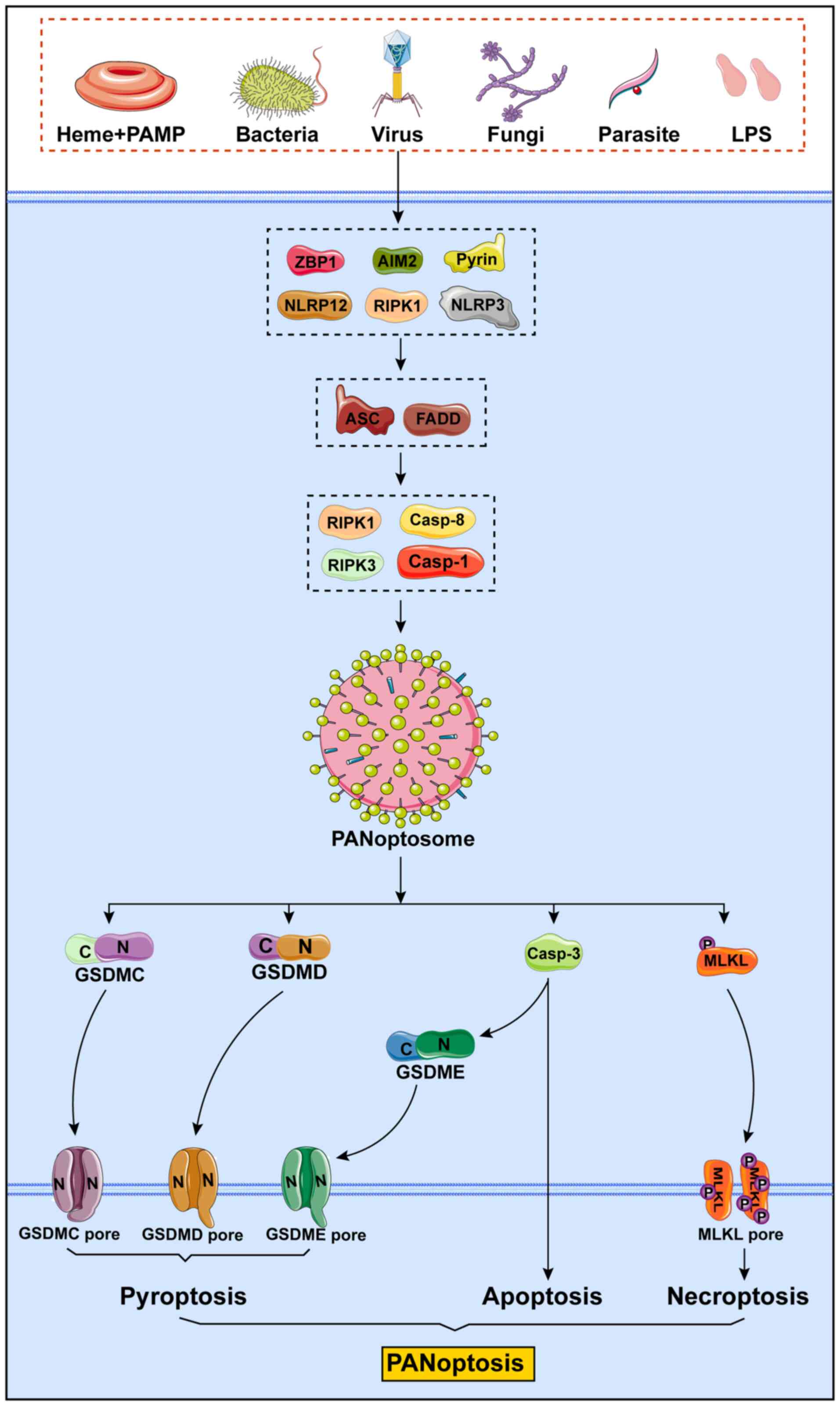 | Figure 4Mechanisms of PANoptosis. PANoptosis
is an innate immune, lytic and inflammatory cell death pathway
driven by caspases and RIPKs and regulated by the PANoptosome,
which possesses the characteristics of apoptosis, pyroptosis and
necroptosis. Different triggering factors can induce the formation
of different PANoptosomes to cause PANoptosis. The main sensors
that have been recognized to date are ZBP1, AIM2, pyrin, NLRP12,
RIPK1 and NLRP3. These sensors bind to the adapters ASC or FADD,
which then form the PANoptosome under the action of catalytic
effectors, such as RIPK1, caspase-8, RIPK3 and caspase-1, thereby
causing PANoptosis of the cells. PAMP, pathogen-associated
molecular pattern; LPS, lipopolysaccharide; ZBP1, Z-DNA binding
protein 1; AIM2, absent in melanoma 2; NLRP12, nucleotide-binding
leucine-rich repeat-containing receptor 12; NLRP3, the
nucleotide-binding domain leucine-rich repeat and pyrin domain
containing receptor 3; RIPK1, receptor-interacting serine/threonine
protein kinase 1; RIPK3, receptor-interacting serine/threonine
protein kinase 3; ASC, adapter protein apoptosis-associated
speck-like protein containing a caspase recruitment domain; GSDM,
gasdermin; MLKL, mixed lineage kinase domain-like protein. |
The PANoptosome, central to this process, is not
merely a conglomerate of inflammasome components, including the
NLRP3 inflammasome and the ASC, but also a nexus for apoptotic and
necroptotic molecules such as caspase-8, caspase-3, RIPK1, RIPK3
and MLKL (99). The activation
of PANoptosis underscores a crucial insight: The singular
inhibition of any one programmed necrotic pathway, such as impeding
the NLRP3 inflammasome or curtailing the production of GSDMD, is
insufficient to abrogate cell death by PANoptosis (100).
In essence, PANoptosis epitomizes an inflammatory
PCD pathway governed by specific activators and regulated under the
supervision of the PANoptosome (Fig.
2), which also suggests a molecular framework. This pathway
encapsulates the quintessential characteristics of pyroptosis,
apoptosis and necroptosis. However, its complexity and multifaceted
nature defy reductionist explanations limited to any singular PCD
mechanism.
Potential molecular mechanisms of
PANoptosis
Recent advancements in cellular biology have
delineated three distinct classes of molecules integral to the
composition of the (101)
PANoptosome: i) Sensor molecules, exemplified by ZBP1, adept at
identifying DAMPs and pathogen-associated molecular patterns
(PAMPs); ii) structural molecules, encompassing caspase-8, NLRP3
inflammasome, caspase-1 and RIPK3; and iii) effector molecules,
including caspase-3, GSDMD and MLKL, among others. ZBP1, which
exhibits robust innate immune sensing capabilities, has been
recognized as a pivotal signal initiator in innate immune responses
and PANoptosis (102). The role
of ZBP1-mediated PANoptosis is twofold; it facilitates the
eradication of invasive pathogens and uncontrolled tumor cells, yet
anomalously, it can also induce excessive inflammatory responses in
a range of contexts, infectious and non-infectious alike (103).
Of note, during PANoptosis, there is a concurrent
activation of caspase-1, caspase-8 and caspase-3, inducing cell
death through the intricate interplay within the caspase system.
Caspase-8, initially identified as a harbinger of apoptosis, has
emerged as a seminal regulatory node bridging apoptotic and
necrotic pathways. It serves as a precursor in NLRP3 inflammasome
activation, thereby inducing pyroptosis (104). Hence, caspase-8 assumes a
central role in PANoptosis, orchestrating the delicate balance
between apoptosis, necroptosis and pyroptosis, and ultimately
influencing the cell's fate by activating specific death
signals.
The advent of PANoptosis and the concept of the
PANoptosome represent a paradigm shift in our comprehension of
innate immune responses and PCD mechanisms. This conceptual
framework unravels the extensive molecular crosstalk inherent to
various innate immune pathways. Identifying novel triggers and
upstream targets is imperative for improving our understanding of
this distinct form of cell death, potentially paving the way for
groundbreaking therapeutic interventions.
ZBP1
ZBP1-mediated PANoptosis has a critical role in
eliminating invasive pathogens and uncontrolled tumor cells.
However, aberrant cell death mediated by ZBP1 can also induce
excessive inflammatory responses in both infectious and
non-infectious environments (102,103). In addition, during the process
of PANoptosis, caspase-1, caspase-8 and caspase-3 are concurrently
activated, relying on additional interconnections within the
caspase systems to promote cell death (102). Notably, caspase-8, an initiator
of apoptosis, was one of the earliest discovered intermediaries in
both the apoptotic and necrotic pathways. Caspase-8 also acts
upstream of NLRP3 inflammasome activation, facilitating the
occurrence of pyroptosis (105). Therefore, as a central
regulatory factor in PANoptosis, caspase-8 influences the interplay
between apoptosis, necroptosis and pyroptosis. It determines the
type of cell death through the activation of death signaling
molecules, playing a key role in the context of PANoptosis
(106).
RIPK1
RIPK1 was identified as a pivotal serine/threonine
kinase, instrumental in regulating cell survival, apoptosis and
necrosis. Within PANoptosis, RIPK1 engages in critical interactions
with proteins such as RIPK3 and MLKL, thereby modulating an array
of cell death pathways, encompassing apoptosis, necrosis and
pyroptosis (107,108). A notable instance of the
regulatory role of RIPK1 was observed in cerebral ischemic injury,
where it governs cell death via the RIPK3/MLKL pathway. A study
investigated the effect of polymyxin B, a polypeptide antibiotic,
on necroptosis in a rat model of stroke (109). The results indicated that
polymyxin B could enhance the endosomal sorting complex required
for transport III machinery and suppress the RIPK1/RIPK3/MLKL
pathway, leading to reduced necroptosis and reduced brain injury in
the rat model of stroke. This effect was also observed in
hypoxia-treated HT22 cells, suggesting the potential of polymyxin B
in treating necroptosis-related brain damage.
Furthermore, the significance of RIPK1 extends to
its involvement in skin inflammation and keratinocyte necrosis. A
study by Duan et al (110) highlighted the critical role of
necroptosis in psoriasis, a common autoimmune and chronic
inflammatory skin disorder. They found that RIPK1 and MLKL were
significantly upregulated in human psoriatic lesions, and the
increased tendency of necroptosis in imiquimod-induced psoriasiform
skin in mice highlighted the involvement of necroptosis in the
pathogenesis of psoriasis. Of note, the study demonstrated that
inhibiting necroptosis using the inhibitor of RIPK1
R-7-Cl-O-Necrostatin-1 (Nec-1s) and MLKL-inhibitor necrosulfonamide
could effectively block inflammatory responses and reduce the
production of key inflammatory factors in both cell and mouse
psoriasis models.
RIPK3
RIPK3, functioning synergistically with RIPK1,
drives cells towards necroptosis. The activation of RIPK3 catalyzes
the phosphorylation of MLKL (111), culminating in the disruption of
the cell membrane and subsequent cell death. Central to promoting
PANoptosis, RIPK3 has been found to induce necroptotic death in
tumor cells via the RIPK1/MLKL pathway. The study by Alaaeldin
et al (112) was
primarily centered on the impact of a novel ciprofloxacin
derivative on RIPK3 and its implications for PANoptosis in cancer
therapy. The findings revealed that the ciprofloxacin derivative
effectively bound to and inhibited topoisomerases I and II,
significantly reducing the proliferation, migration and colony
formation of the cancer cells. Crucially, the study highlighted the
derivative's ability to increase the proportion of apoptotic cells
and activate the necro-apoptotic pathway, primarily by stimulating
RIPK3, RIPK1 and MLKL proteins. This research provides valuable
insight into the role of RIPK3 in PANoptosis and underscores the
potential of targeting this pathway as a novel approach to cancer
treatment. The ability of the ciprofloxacin derivative to modulate
RIPK3 and induce PANoptosis opens novel avenues for the development
of anticancer therapies, particularly for HepG2 and A549 cancer
types, and contributes to the broader understanding of the
molecular mechanisms underlying cancer cell death. Furthermore,
recent research identified the modulatory role of RIPK3 in cell
death and inflammation, particularly via its interactions with the
RIPK1 and MLKL pathways. It underscores the pivotal function of
RIPK3 in initiating and regulating necroptotic processes, as well
as its diverse and complex activation in response to various
cellular stressors (113).
MLKL
MLKL is a critical effector in pyroptosis and
PANoptosis. Phosphorylated by RIPK3, MLKL triggers the rupture of
the cell membrane, leading to cell death. The activation of MLKL is
closely associated with pathological processes in various disease
models. For instance, in cerebral ischemic injury, MLKL is a
crucial regulator of cell death, operating through the RIPK1/RIPK3
pathway. Zhang et al (114) investigated the role of MLKL in
PANoptosis, particularly in ischemic brain injury following stroke,
focusing on the effects of ligustroflavone, a compound with
anti-inflammatory properties. They showed that ligustroflavone
reduced brain injury and decreased the levels of
necroptosis-associated proteins, including MLKL, in a rat model of
ischemic stroke. However, the study noted an inconsistency in the
regulation of RIPK1 levels, suggesting a need for further
exploration of a ligustroflavone selective mechanism on
necroptosis-related proteins. While the study identified potential
targets of ligustroflavone, including RIPK1, RIPK3 and MLKL using
the Molecular Operating Environment program, the unaltered levels
of RIPK1 call for a more detailed investigation into its
interaction with MLKL in PANoptosis. The study highlighted the
potential of ligustroflavone as a therapeutic agent for ischemic
brain injury. Thus, there is a need for a deeper understanding of
the molecular mechanisms concerning MLKL in PANoptosis.
The role of MLKL extends to the pharmacodynamics of
antidepressants, where it regulates the necroptotic death of
neuronal cells via the RIPK1-RIPK3-MLKL pathway. Yan et al
(115) focused on the
antidepressant effects of Xiaoyaosan, in particular examining its
influence on MLKL-mediated necroptosis in the context of
depression. They revealed that Xiaoyaosan impacted the MLKL pathway
in a mouse model of depression. This suggested that the therapeutic
effect of Xiaoyaosan on depression may involve modulating MLKL and
the related necroptotic processes, offering a novel perspective for
treating depression by targeting specific aspects of
PANoptosis.
TNF signaling pathway and PANoptosis
TNF-α, by interacting with TNF receptors on the cell
surface, triggers a complex cascade of downstream signals. These
signals lead to various cell fates, including apoptosis, necrosis
or other forms of cell death, covering the range of PANoptosis. The
activation of RIPK1 and RIPK3 is central to this signaling cascade.
These kinases have a key role in determining cellular outcomes,
decisively influencing a cell's fate toward survival, apoptosis or
necrosis. The complex interactions within the TNF signaling
pathway, especially between RIPK1 and RIPK3, underscore the
complexity of cellular responses to pathological stimuli.
Karki et al (116) discovered that in COVID-19, the
combined production of TNF-α and IFN-γ by innate immune cells
induced PANoptosis. They also found that deleting individual
mediators of pyroptosis, apoptosis or necroptosis was insufficient
to prevent cell death. Ma et al (117) explored the role of lncRNA SPRY4
intronic transcript 1 (SPRY4-IT1) in hepatocellular carcinoma
(HCC), focusing on its role in the TNF signaling pathway and
potential implications for PANoptosis. Although the study offered
valuable insight into the role of SPRY4-IT1 in HCC and its link to
the TNF signaling pathway, it raised questions regarding the direct
link between SPRY4-IT1 and PANoptosis. Further research is required
to explore how SPRY4-IT1, through its interaction with the TNF
signaling pathway, may influence PANoptosis in HCC, thereby
contributing to tumor progression and metastasis.
TLR signaling pathway in PANoptosis
The TLR signaling pathway, a cornerstone in
immunological research, has garnered extensive attention for its
critical role in recognizing PAMPs and DAMPs. This pathway,
activating adaptor proteins, such as myeloid differentiation factor
88 and Toll-interleukin 1 receptor domain-containing adapter
inducing IFN-β, leads to the subsequent activation of NF-κB and
other transcription factors, orchestrating a cascade of
inflammatory responses and cell death (118). While the implications of the
TLR signaling pathway in conventional cell death modalities such as
apoptosis and necrosis have been thoroughly investigated, its
direct contribution to PANoptosis remains undetermined. Considering
the regulatory influence of the TLR signaling pathway over the
traditional cell death modes, it is plausible to hypothesize its
potential involvement in PANoptosis. Of note, the TLR signaling
pathway may indirectly modulate cell death mechanisms associated
with PANoptosis by activating NF-κB.
Contemporary research endeavors have predominantly
concentrated on delineating the role of the TLR signaling pathway
within specific pathological contexts, such as neurodegenerative
diseases, cancer and inflammatory disorders. Recent studies
demonstrated the therapeutic potential of polyphenolic compounds in
neurodegenerative diseases via modulation of the TLR signaling
pathway, indicating the regulatory effects of probiotic-derived
metabolites on the TLR signaling pathway in inflammatory responses
(119,120). Although these studies do not
establish a direct link between the TLR signaling pathway and
PANoptosis, they underscore the latent role of this pathway in
cellular death and inflammation, laying a foundational framework
for future investigations into its role in PANoptosis.
In summary, the lack of direct evidence linking the
TLR signaling pathway to the regulation of PANoptosis, juxtaposed
with its pivotal role in immune responses and cell death, paves the
way for future research. Future studies may unravel the potential
of this pathway in PANoptosis, an exploration that promises to
yield novel insight into the pathogenesis and therapeutic
approaches for diseases associated with PANoptosis. This emerging
field of study stands at the forefront of expanding our
understanding of complex cellular death mechanisms and their
implications for human health and disease.
Intracellular stress pathways in
PANoptosis
In the study of PANoptosis, the roles of
intracellular stress pathways, particularly ER stress and
autophagy, have not been directly confirmed to have a clear
association with PANoptosis. However, the roles of these pathways
in responding to prolonged stress and maintaining cellular
homeostasis may imply their potential impact on PANoptosis. ER
stress is a cellular response to abnormal protein folding or
accumulation, typically activated through the protein kinase
RNA-like endoplasmic reticulum kinase (PERK) and IRE1α pathways.
Autophagy, on the other hand, is an intracellular degradation
process that maintains cellular homeostasis by digesting and
recycling cellular components.
Although current research has not directly linked ER
stress and autophagy with the activation of PANoptosis, their roles
in responding to stress and regulating cell death are undeniable.
For example, it has been suggested that the interaction between ER
stress and autophagy can contribute to the development of novel
therapeutic strategies to alleviate cellular stress and
inflammation associated with various pathologies (121). In addition, the regulation of
the ER stress-autophagy axis may be a potential strategy for
treating neurodegeneration and neurological deficits following
ischemic stroke (122). While
these studies have not directly demonstrated the role of ER stress
and autophagy in PANoptosis, they reveal the significance of these
pathways in cellular stress responses and the regulation of
cellular fate.
Mitochondrial signaling pathways in
PANoptosis
In the intricate landscape of PANoptosis regulation,
mitochondrial signaling pathways emerge as central players,
particularly in orchestrating cell death processes. Mitochondria,
quintessential for energy metabolism, also serve as pivotal
determinants of cellular fate. Dysfunctions in these organelles can
pivotally steer cells toward PANoptosis. Emerging research has
elucidated a profound association between alterations in
mitochondrial function and the initiation of PANoptosis. A notable
example is the increased permeability of the mitochondrial outer
membrane, culminating in the release of cytochrome C (123). This event, pivotal in
apoptosis, may also be a critical trigger of PANoptosis.
Furthermore, the generation of ROS by mitochondria is implicated in
amplifying cell death pathways, including PANoptosis. Consequently,
nuanced shifts in mitochondrial functionality are increasingly
recognized as determinants in the decision between survival and
PANoptosis.
Shi et al (124) explored the roles of
mitochondrial ROS (mtROS) and reverse electron transport (RET) in
the induction of PANoptosis and showed that anti-RET reagents such
as 1-methoxy PMS (a stable electron transfer mediator in CCK-8
kits) and dimethyl fumarate can effectively inhibit PANoptosis by
blocking mtROS production. In a related context, Yuan et al
(125) used a multi specific
platinum complex SEP, which was constructed by conjugating a
quinone derivative seratrodast to a prodrug of cisplatin, as the
first metal complex capable of inducing PANoptosis in KRAS-mutant
pancreatic ductal adenocarcinoma cells, offering a novel strategy
to overcome apoptotic resistance. In doxorubicin (DOX)-induced
myocardial injury, FUNDC1 stabilizes mitochondrial DNA by binding
to TUFM, thereby protecting cardiomyocytes from DOX-induced
PANoptosis (10). Finally, She
et al (126) also
emphasized the significance of mitochondrial dysfunction in
ischemic stroke and cerebral ischemia/RI. This links them to
PANoptosis and other types of PCD, suggesting neuroprotective
agents targeting mitochondrial dysfunction as promising treatments
for related brain injuries.
These studies highlight the pivotal role of
mitochondrial signaling in PANoptosis, underscoring the
significance of mitochondrial dysfunction and enhanced outer
membrane permeability in cell death processes. However, they also
highlight the gaps in our understanding of mitochondrial
involvement in PANoptosis. Present research predominantly
concentrates on the impact of the mitochondria on traditional cell
death pathways; however, there is a pressing need for more
comprehensive studies on their specific roles within
PANoptosis.
These insights show the multifaceted roles of
mitochondrial signaling in cell death and inflammatory responses
and pave the way for novel explorations into the regulation of
PANoptosis. Future research endeavors should focus on elucidating
the direct interplay between functional shifts of mitochondria and
PANoptosis and its consequent effects on cell fate. Unraveling
these connections holds substantial promise for developing
innovative therapeutic strategies aimed at targeting PANoptosis,
and would thus mark a significant stride in cellular and molecular
biology.
Comparison of PANoptosis with other forms of
cell death
PANoptosis, as an encompassing cell death mechanism,
uniquely bridges and diverges from conventional modalities such as
apoptosis, necroptosis and pyroptosis in molecular mechanisms.
Apoptosis, a caspase-dependent PCD, is characterized by distinctive
nuclear condensation and DNA fragmentation. Necroptosis is usually
'violent' and is characterized by phosphorylated MLKL creating
holes in the cell membrane, leading to membrane rupture and a
subsequent inflammatory response. Pyroptosis is another type of PCD
that involves major members of the caspase family, such as caspases
1, 3, 4, 5, 8 and 11, and is mediated by cellular membrane pores
via the GSDM family of proteins, which ultimately leads to membrane
disruption and the release of cellular components. PANoptosis
amalgamates these varied mechanisms, yielding a multifaceted and
intricate cellular fate-determination process.
Within the PANoptosis pathway, apoptotic proteins,
notably caspases, assume pivotal roles. Caspase-8, for example,
extends beyond its traditional role in the execution of apoptosis,
potentially catalyzing PANoptosis via interplay with molecules from
other cell death pathways. Pioneering research by Zheng et
al (102) delineates the
integral role of ZBP1 in innate immune responses against viral RNA
and endogenous nucleic acids. During influenza virus infection,
ZBP1-mediated sensing of Z-RNA induces PANoptosis, a synchronized
cell death pathway orchestrated by the PANoptosome complex. This
pathway includes the assembly of the ZBP1-NLRP3 inflammasome,
activated upon ZBP1 recruitment of RIPK3 and caspase-8 in response
to Z-RNA. This assembly is further modulated by various host
factors, including those involved in the type I interferon
signaling pathway, caspase-6 and influenza viral proteins such as
M2, NS1 and PB1-F2. In addition, the necrotic attributes in
PANoptosis are underpinned by its integration of key molecules from
a range of cell death pathways, such as RIPK1 and RIPK3, with MLKL
playing a crucial role as a necroptosis executor (127).
Autophagy is a type II cell death process in which
autophagosomes are formed under the action of autophagy-related
genes and degrade aging or damaged organelles and protein
aggregates by binding with lysosomes to meet the metabolic needs of
cells and renewing certain organelles. Autophagy is involved in the
development of tumors, autoimmune, renal, neurological,
cardiovascular, cerebrovascular, metabolic and several other
diseases (128-130). Since autophagy is a multistep
process, it can be studied using autophagy inducers and inhibitors
specific to the different stages of this process. Classical
autophagy inhibitors include not only PI3K inhibitors that inhibit
the initiation phase of autophagy such as 3-methyladenine, LY294002
and wortmannin, but also drugs that act on the activity of
lysosomal enzymes in the terminal phase of autophagy, such as
leupeptin, pepstatin A, E-64d, bafilomycin A1, concanamycin A,
concanamycin B, methyl- or propylamine, chloroquine and neutral
red. Autophagy inducers include rapamycin, Torin1, PP242,
KU-0063794, PI-103 and NVP-BEZ235, which inhibit mTOR activity
(131). In contrast to
PANoptosis, autophagy is not dependent on caspase enzymes and RIPKs
but occurs solely based on the actions of lysosomes.
ER stress is caused by the accumulation of misfolded
proteins within the ER and activates the unfolded protein response
(UPR). ER stress is present in a wide range of diseases, such as
cancer, neurodegenerative diseases, metabolic disorders, hepatic
and renal diseases and CVDs (132-134). There are three primary ER
stress sensors, including inositol-requiring enzyme 1 (IRE1),
activating transcription factor 6 and PERK, which are not only
involved in the modulation of ER function but also regulate the
rate of protein synthesis. Certain reagents that stabilize protein
folding intermediates, the drug ISRIB that regulates the
phosphorylation state of the α-subunit of eukaryotic translational
initiation factor 2, PERK inhibitor GSK2606414 and IRE1 inhibitor
MKC8866, can all be used to study ER stress (135). The most important difference
between ER stress and PANoptosis is that in vivo
accumulation of protein aggregates induces the UPR, which leads to
ER stress, and further ER stress can initiate apoptosis. In this
regard, ER stress and PANoptosis are related.
Anoikis is a form of apoptosis observed in tissue
cells (except blood cells), and is a form of PCD triggered by the
loss of cell adhesion to the ECM or inappropriate cell adhesion,
with the aim of removing misplaced or dislodged cells under
physiological or pathological conditions. It has an important role
in organismal development, tissue homeostasis and tumor metastasis.
Anoikis is closely associated with several types of cancer, liver
disease, cardiomyopathy and other diseases (136-138). Of note, anoikis is commonly
activated during tumorigenesis, which is one of the body's
self-protective means of preventing tumor metastasis. In metastatic
cancer, cancer cells acquire anoikis resistance through autocrine
or paracrine activation of survival signals, alterations of
integrin expression patterns and inhibition of death pathways, to
regain the ability to attach and spread, which ultimately leads to
invasion and metastasis (139-141). From an anti-anoikis
perspective, normalizing ECM remodeling by targeting matrix-related
enzymes (matrix metalloproteinases, lysyl oxidase and lysyl
oxidase-like proteins) or ECM-associated cell-surface receptors
(integrin, discoidin domain receptors and CD44), and the use of
vitamin D may have significant therapeutic anti-tumor potential
(138,142-144). Anoikis uses caspases as key
signaling molecules and primarily involves the PI3K/Akt, integrin,
MAPK, Wnt/β-catenin, TGF-β, Smad and NF-κB pathways, and is thus
both distinct and related to PANoptosis.
Conclusions and future perspectives
PANoptosis has emerged as a novel and distinct form
of PCD, integrating elements from apoptosis, pyroptosis and
necroptosis. This convergence is epitomized in the PANoptosome
complex, a sophisticated entity that coordinates the synchronous
activation of these diverse cell death pathways, thereby enriching
the contemporary understanding of cellular demise. In delineating
the specific pathways of apoptosis, pyroptosis and necroptosis,
PANoptosis stands out for its amalgamation of these mechanisms. It
presents a dynamic interplay involving caspases, GSDMs, RIPKs and
other molecular actors, highlighting a complex network of
interactions. This understanding highlights novel avenues for
therapeutic interventions, particularly in diseases characterized
by immune responses and inflammation.
However, the current understanding of PANoptosis in
the context of CVDs is still limited and much remains to be
explored regarding the molecular intricacies and clinical
applications. Therefore, the promise of PANoptosis in CVDs is
substantial. It offers a comprehensive framework for understanding
complex pathologies such as MI and PH, where multiple forms of cell
death coexist. Importantly, PANoptosis underscores the inflammatory
aspects of CVDs, such as AS, providing potential anti-inflammatory
strategies. The PANoptosis pathway involves apoptotic proteins,
inflammasomes and necroptotic elements, hinting at a complex
regulatory network that could be pivotal in cardiovascular
pathologies. For example, the pro-inflammatory aspects of
PANoptosis, as observed in the recruitment of RIPK3 and caspase-8,
may contribute to the characteristic inflammatory milieu observed
during atherosclerotic plaque formation or in the response to
infarction in myocardial tissues. Similarly, the involvement of the
type I interferon signaling pathway components within the
PANoptosis mechanism may highlight a role in the modulation of
immune responses in HF or PH. Future research should focus on
delineating the roles of PANoptosis in these states, its dynamic
interplay with traditional cell death pathways, and its regulatory
mechanisms. Understanding PANoptosis in CVDs could open new avenues
for therapeutic interventions. As an emerging field, PANoptosis has
the potential to significantly impact biomedical research, offering
deeper insight into the complexities of cell death and novel
therapeutic prospects for various diseases, including those
affecting the cardiovascular system.
In conclusion, PANoptosis represents a significant
advance in our understanding of cell death, it is essential to
acknowledge certain limitations within this review. The interplay
between PANoptosis and other forms of cell death, such as
ferroptosis and autophagy, is currently unclear. Recent findings
(145) suggest that the
ferroptosis inhibitor liproxstatin-1 may have a protective role
against steatosis and steatohepatitis in metabolic
dysfunction-associated fatty liver disease mice by potentially
regulating PANoptosis, implying a plausible link between
ferroptosis and PANoptosis. Considering these recent discoveries,
further studies could investigate the potential connections between
PANoptosis and these additional mechanisms, elucidating whether
they intersect or operate independently in various disease
contexts. Consequently, advancing our comprehension of cell death
mechanisms, including the interplay between different forms, stands
as a crucial direction for future research in the field. Its
potential to revolutionize diagnostics, inspire innovative
therapeutic strategies and tailor treatments to individual needs
positions it as a pivotal area for future exploration.
Availability of data and materials
Not applicable.
Authors' contributions
WL and XG conceived and designed the review. XG
wrote the manuscript. XG, SL and CM assisted with the drawings. MC,
SL, CM, YH and WL reviewed and edited the manuscript. All authors
have read and approved the manuscript. Data authentication is not
applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
All the figures in the manuscript were drawn in
Adobe Illustrator 2020.
Funding
The study was supported by the National Natural Science
Foundation of China (grant no. 81700269), Key Platform of
Department of Education of Guangdong Province (grant no.
2021LSYS007), Discipline Construction Project of Guangdong Medical
University (grant no. 4SG21233G), Zhanjiang Science and Technology
Development Special Funding Competitive Allocation Project (grant
nos. 2022E05011, 2022A01196 and 2021A05158) and the Project of
Administration of Traditional Chinese Medicine of Guangdong
Province of China (grant no. 20221208).
References
|
1
|
Sun X, Yang Y, Meng X, Li J, Liu X and Liu
H: PANoptosis: Mechanisms, biology, and role in disease. Immunol
Rev. 321:246–262. 2024. View Article : Google Scholar
|
|
2
|
Lee S, Karki R, Wang Y, Nguyen LN,
Kalathur RC and Kanneganti T-D: AIM2 forms a complex with pyrin and
ZBP1 to drive PANoptosis and host defence. Nature. 597:415–419.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gullett JM, Tweedell RE and Kanneganti TD:
It's all in the PAN: Crosstalk, plasticity, redundancies, switches,
and interconnectedness encompassed by PANoptosis underlying the
totality of cell death-associated biological effects. Cells.
11:14952022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yan WT, Yang YD, Hu XM, Ning WY, Liao LS,
Lu S, Zhao WJ, Zhang Q and Xiong K: Do pyroptosis, apoptosis, and
necroptosis (PANoptosis) exist in cerebral ischemia? Evidence from
cell and rodent studies. Neural Regen Res. 17:1761–1768. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pan H, Pan J, Li P and Gao J:
Characterization of PANoptosis patterns predicts survival and
immunotherapy response in gastric cancer. Clin Immunol.
238:1090192022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pandian N and Kanneganti TD: PANoptosis: A
unique innate immune inflammatory cell death modality. J Immunol.
209:1625–1633. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Piamsiri C, Maneechote C, Siri-Angkul N,
Chattipakorn SC and Chattipakorn N: Targeting necroptosis as
therapeutic potential in chronic myocardial infarction. J Biomed
Sci. 28:252021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang H, Yu W, Wang Y, Wu R, Dai Y, Deng Y,
Wang S, Yuan J and Tan R: p53 contributes to cardiovascular
diseases via mitochondria dysfunction: A new paradigm. Free Radic
Biol Med. 208:846–858. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Toldo S and Abbate A: The role of the
NLRP3 inflammasome and pyroptosis in cardiovascular diseases. Nat
Rev Cardiol. 21:219–237. 2024. View Article : Google Scholar
|
|
10
|
Bi Y, Xu H, Wang X, Zhu H, Ge J, Ren J and
Zhang Y: FUNDC1 protects against doxorubicin-induced cardiomyocyte
PANoptosis through stabilizing mtDNA via interaction with TUFM.
Cell Death Dis. 13:10202022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y and Kanneganti TD: From pyroptosis,
apoptosis and necroptosis to PANoptosis: A mechanistic compendium
of programmed cell death pathways. Comput Struct Biotechnol J.
19:4641–4657. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen W, Gullett JM, Tweedell RE and
Kanneganti TD: Innate immune inflammatory cell death: PANoptosis
and PANoptosomes in host defense and disease. Eur J Immunol.
53:e22502352023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ji X, Huang X, Li C, Guan N, Pan T, Dong J
and Li L: Effect of tumor-associated macrophages on the pyroptosis
of breast cancer tumor cells. Cell Commun Signal. 21:1972023.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nagata S and Tanaka M: Programmed cell
death and the immune system. Nat Rev Immunol. 17:333–340. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Man SM, Karki R and Kanneganti TD:
Molecular mechanisms and functions of pyroptosis, inflammatory
caspases and inflammasomes in infectious diseases. Immunol Rev.
277:61–75. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu Y, Zhang J, Yu S, Li Y, Zhu J, Zhang K
and Zhang R: Cell pyroptosis in health and inflammatory diseases.
Cell Death Discov. 8:1912022. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu X, Xia S, Zhang Z, Wu H and Lieberman
J: Channelling inflammation: Gasdermins in physiology and disease.
Nat Rev Drug Discov. 20:384–405. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feng Y, Li M, Yangzhong X, Zhang X, Zu A,
Hou Y, Li L and Sun S: Pyroptosis in inflammation-related
respiratory disease. J Physiol Biochem. 78:721–737. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu H and Sun A: Programmed necrosis in
heart disease: Molecular mechanisms and clinical implications. J
Mol Cell Cardiol. 116:125–134. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mishra PK, Adameova A, Hill JA, Baines CP,
Kang PM, Downey JM, Narula J, Takahashi M, Abbate A, Piristine HC,
et al: Guidelines for evaluating myocardial cell death. Am J
Physiol Heart Circ Physiol. 317:H891–H922. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shi Z, Yuan S, Shi L, Li J, Ning G, Kong X
and Feng S: Programmed cell death in spinal cord injury
pathogenesis and therapy. Cell Proliferation. 54:e129922021.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li M, Wang ZW, Fang LJ, Cheng SQ, Wang X
and Liu NF: Programmed cell death in atherosclerosis and vascular
calcification. Cell Death Dis. 13:4672022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi C, Cao P, Wang Y, Zhang Q, Zhang D,
Wang Y, Wang L and Gong Z: PANoptosis: A cell death characterized
by pyroptosis, apoptosis, and necroptosis. J Inflamm Res.
16:1523–1532. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei Q, Ren H, Zhang J, Yao W, Zhao B and
Miao J: An inhibitor of Grp94 inhibits OxLDL-Induced autophagy and
apoptosis in VECs and stabilized atherosclerotic plaques. Front
Cardiovasc Med. 8:7575912021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo J, Li J, Zhang J, Guo X, Liu H, Li P,
Zhang Y, Lin C and Fan Z: LncRNA PVT1 knockdown alleviated
ox-LDL-induced vascular endothelial cell injury and atherosclerosis
by miR-153-3p/GRB2 axis via ERK/p38 pathway. Nutr Metab Cardiovasc
Dis. 31:3508–3521. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang W, Tang W, Shan E, Zhang L, Chen S,
Yu C and Gao Y: MiR-130a-5p contributed to the progression of
endothelial cell injury by regulating FAS. Eur J Histochem.
66:33422022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sayed S, Faruq O, Preya UH and Kim JT:
Cathepsin S knockdown suppresses endothelial inflammation,
angiogenesis, and complement protein activity under hyperglycemic
conditions in vitro by inhibiting NF-κB signaling. Int J Mol Sci.
24:54282023. View Article : Google Scholar
|
|
28
|
Tang H, Li K, Zhang S, Lan H, Liang L,
Huang C and Li T: Inhibitory effect of paeonol on apoptosis,
oxidative stress, and inflammatory response in human umbilical vein
endothelial cells induced by high glucose and palmitic acid induced
through regulating SIRT1/FOXO3a/NF-κB pathway. J Interferon
Cytokine Res. 41:111–124. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu X, Xu Y, Cheng S, Zhou X, Zhou F, He
P, Hu F, Zhang L, Chen Y and Jia Y: Geniposide combined with
Notoginsenoside R1 attenuates inflammation and apoptosis in
atherosclerosis via the AMPK/mTOR/Nrf2 signaling pathway. Front
Pharmacol. 12:6873942021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Du M, Wang C, Yang L, Liu B, Zheng Z, Yang
L, Zhang F, Peng J, Huang D and Huang K: The role of long noncoding
RNA Nron in atherosclerosis development and plaque stability.
iScience. 25:1039782022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sinha SK, Miikeda A, Fouladian Z,
Mehrabian M, Edillor C, Shih D, Zhou Z, Paul MK, Charugundla S,
Davis RC, et al: Local M-CSF (Macrophage Colony-stimulating factor)
expression regulates macrophage proliferation and apoptosis in
atherosclerosis. Arterioscler Thromb Vasc Biol. 41:220–233. 2021.
View Article : Google Scholar
|
|
32
|
Niu N, Miao H and Ren H: Effect of
miR-182-5p on apoptosis in myocardial infarction. Heliyon.
9:e215242023. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin M, Liu X, Zheng H, Huang X, Wu Y,
Huang A, Zhu H, Hu Y, Mai W and Huang Y: IGF-1 enhances BMSC
viability, migration, and anti-apoptosis in myocardial infarction
via secreted frizzled-related protein 2 pathway. Stem Cell Res
Ther. 11:222020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fu DL, Jiang H, Li CY, Gao T, Liu MR and
Li HW: MicroRNA-338 in MSCs-derived exosomes inhibits cardiomyocyte
apoptosis in myocardial infarction. Eur Rev Med Pharmacol Sci.
24:10107–10117. 2020.PubMed/NCBI
|
|
35
|
Wu H, Zhao ZA, Liu J, Hao K, Yu Y, Han X,
Li J, Wang Y, Lei W, Dong N, et al: Long noncoding RNA Meg3
regulates cardiomyocyte apoptosis in myocardial infarction. Gene
Ther. 25:511–523. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao L, Yang XR and Han X: MicroRNA-146b
induces the PI3K/Akt/NF-κB signaling pathway to reduce vascular
inflammation and apoptosis in myocardial infarction by targeting
PTEN. Exp Ther Med. 17:1171–1181. 2019.PubMed/NCBI
|
|
37
|
Luo C, Xiong S, Huang Y, Deng M, Zhang J,
Chen J, Yang R and Ke X: The Novel Non-coding transcriptional
regulator Gm18840 drives cardiomyocyte apoptosis in myocardial
infarction post ischemia/reperfusion. Front Cell Dev Biol.
9:6159502021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhou F, Fu WD and Chen L: MiRNA-182
regulates the cardiomyocyte apoptosis in heart failure. Eur Rev Med
Pharmacol Sci. 23:4917–4923. 2019.PubMed/NCBI
|
|
39
|
Zhang Y, Li C, Meng H, Guo D, Zhang Q, Lu
W, Wang Q, Wang Y and Tu P: BYD Ameliorates oxidative
stress-induced myocardial apoptosis in heart failure post-acute
myocardial infarction via the P38 MAPK-CRYAB signaling pathway.
Front Physiol. 9:5052018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yan X, Wu H, Ren J, Liu Y, Wang S, Yang J,
Qin S and Wu D: Shenfu Formula reduces cardiomyocyte apoptosis in
heart failure rats by regulating microRNAs. J Ethnopharmacol.
227:105–112. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Colpman P, Dasgupta A and Archer SL: The
role of mitochondrial dynamics and mitotic fission in regulating
the cell cycle in cancer and pulmonary arterial hypertension:
Implications for dynamin-related protein 1 and mitofusin2 in
hyperproliferative diseases. Cells. 12:18972023. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dabral S, Tian X, Kojonazarov B, Savai R,
Ghofrani HA, Weissmann N, Florio M, Sun J, Jonigk D, Maegel L, et
al: Notch1 signalling regulates endothelial proliferation and
apoptosis in pulmonary arterial hypertension. Eur Respir J.
48:1137–1149. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang DT, Tuo L, Bai X, Bing WD, Qu QX,
Zhao X, Song GM, Bi YW and Sun WY: Prostaglandin E1 reduces
apoptosis and improves the homing of mesenchymal stem cells in
pulmonary arterial hypertension by regulating hypoxia-inducible
factor 1 alpha. Stem Cell Res Ther. 13:3162022. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jiang Y, Hei B, Hao W, Lin S, Wang Y, Liu
X, Meng X and Guan Z: Clinical value of lncRNA SOX2-OT in pulmonary
arterial hypertension and its role in pulmonary artery smooth
muscle cell proliferation, migration, apoptosis, and inflammatory.
Heart Lung. 55:16–23. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hu F, Liu H, Wang C, Li H and Qiao L:
Expression of the microRNA-30 family in pulmonary arterial
hypertension and the role of microRNA-30d-5p in the regulation of
pulmonary arterial smooth muscle cell toxicity and apoptosis. Exp
Ther Med. 23:1082022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li X, Liu C, Qi W, Meng Q, Zhao H, Teng Z,
Xu R, Wu X, Zhu F, Qin Y, et al: Endothelial Dec1-PPARγ axis
impairs proliferation and apoptosis homeostasis under hypoxia in
pulmonary arterial hypertension. Front Cell Dev Biol. 9:7571682021.
View Article : Google Scholar
|
|
47
|
Cuthbertson I, Morrell NW and Caruso P:
BMPR2 mutation and metabolic reprogramming in pulmonary arterial
hypertension. Circ Res. 132:109–126. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vera-Zambrano A, Lago-Docampo M, Gallego
N, Franco-Gonzalez JF, Morales-Cano D, Cruz-Utrilla A,
Villegas-Esguevillas M, Fernández-Malavé E, Escribano-Subías P,
Tenorio-Castaño JA, et al: Novel Loss-of-function KCNA5 variants in
pulmonary arterial hypertension. Am J Respir Cell Mol Biol.
69:147–158. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ye B, Peng X, Su D, Liu D, Huang Y, Huang
Y and Pang Y: Effects of YM155 on the proliferation and apoptosis
of pulmonary artery smooth muscle cells in a rat model of high
pulmonary blood flow-induced pulmonary arterial hypertension. Clin
Exp Hypertens. 44:470–479. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lu Y, Wu J, Sun Y, Xin L, Jiang Z, Lin H,
Zhao M and Cui X: Qiliqiangxin prevents right ventricular
remodeling by inhibiting apoptosis and improving metabolism
reprogramming with pulmonary arterial hypertension. Am J Transl
Res. 12:5655–5669. 2020.PubMed/NCBI
|
|
51
|
Xu YJ, Zheng L, Hu YW and Wang Q:
Pyroptosis and its relationship to atherosclerosis. Clin Chim Acta.
476:28–37. 2018. View Article : Google Scholar
|
|
52
|
Wu X, Zhang H, Qi W, Zhang Y, Li J, Li Z,
Lin Y, Bai X, Liu X, Chen X, et al: Nicotine promotes
atherosclerosis via ROS-NLRP3-mediated endothelial cell pyroptosis.
Cell Death Dis. 9:1712018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Din AU, Hassan A, Zhu Y, Yin T, Gregersen
H and Wang G: Amelioration of TMAO through probiotics and its
potential role in atherosclerosis. Appl Microbiol Biotechnol.
103:9217–9228. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu P, Chen J, Chen J, Tao J, Wu S, Xu G,
Wang Z, Wei D and Yin W: Trimethylamine N-oxide promotes
apoE-/-mice atherosclerosis by inducing vascular endothelial cell
pyroptosis via the SDHB/ROS pathway. J Cell Physiol. 235:6582–6591.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Moore KJ, Sheedy FJ and Fisher EA:
Macrophages in atherosclerosis: A dynamic balance. Nat Rev Immunol.
13:709–721. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Peng X, Chen H, Li Y, Huang D, Huang B and
Sun D: Effects of NIX-mediated mitophagy on ox-LDL-induced
macrophage pyroptosis in atherosclerosis. Cell Biol Int.
44:1481–1490. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xu S, Chen H, Ni H and Dai Q: Targeting
HDAC6 attenuates nicotine-induced macrophage pyroptosis via
NF-κB/NLRP3 pathway. Atherosclerosis. 317:1–9. 2021. View Article : Google Scholar
|
|
58
|
Wu L, Xie W, Li Y, Ni Q, Timashev P, Lyu
M, Xia L, Zhang Y, Liu L, Yuan Y, et al: Biomimetic nanocarriers
guide extracellular ATP homeostasis to remodel energy metabolism
for activating innate and adaptive immunity system. Adv Sci
(Weinh). 9:e21053762022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kiyan Y, Tkachuk S, Hilfiker-Kleiner D,
Haller H, Fuhrman B and Dumler I: oxLDL induces inflammatory
responses in vascular smooth muscle cells via urokinase receptor
association with CD36 and TLR4. J Mol Cell Cardiol. 66:72–82. 2014.
View Article : Google Scholar
|
|
60
|
Pang Q, Wang P, Pan Y, Dong X, Zhou T,
Song X and Zhang A: Irisin protects against vascular calcification
by activating autophagy and inhibiting NLRP3-mediated vascular
smooth muscle cell pyroptosis in chronic kidney disease. Cell Death
Dis. 13:2832022. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Li Y, Niu X, Xu H, Li Q, Meng L, He M,
Zhang J and Zhang Z and Zhang Z: VX-765 attenuates atherosclerosis
in ApoE deficient mice by modulating VSMCs pyroptosis. Exp Cell
Res. 389:1118472020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yue RC, Lu SZ, Luo Y, Wang T, Liang H,
Zeng J, Liu J and Hu HX: Calpain silencing alleviates myocardial
ischemia-reperfusion injury through the NLRP3/ASC/Caspase-1 axis in
mice. Life Sci. 233:1166312019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ding S, Liu D, Wang L, Wang G and Zhu Y:
Inhibiting MicroRNA-29a protects myocardial ischemia-reperfusion
injury by targeting SIRT1 and suppressing oxidative stress and
NLRP3-mediated pyroptosis pathway. J Pharmacol Exp Ther.
372:128–135. 2020. View Article : Google Scholar
|
|
64
|
Rauf A, Shah M, Yellon DM and Davidson SM:
Role of Caspase 1 in ischemia/reperfusion injury of the myocardium.
J Cardiovasc Pharmacol. 74:194–200. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Do Carmo H, Arjun S, Petrucci O, Yellon DM
and Davidson SM: The Caspase 1 inhibitor VX-765 protects the
isolated rat heart via the RISK pathway. Cardiovasc Drugs Ther.
32:165–168. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang X, Fu Y, Li H, Shen L, Chang Q, Pan
L, Hong S and Yin X: H3 relaxin inhibits the collagen synthesis via
ROS- and P2X7R-mediated NLRP3 inflammasome activation in cardiac
fibroblasts under high glucose. J Cell Mol Med. 22:1816–1825. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zeng C, Duan F, Hu J, Luo B, Huang B, Lou
X, Sun X, Li H, Zhang X, Yin S and Tan H: NLRP3
inflammasome-mediated pyroptosis contributes to the pathogenesis of
non-ischemic dilated cardiomyopathy. Redox Biol. 34:1015232020.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Bai Y, Sun X, Chu Q, Li A, Qin Y, Li Y,
Yue E, Wang H, Li G, Zahra SM, et al: Caspase-1 regulate
AngII-induced cardiomyocyte hypertrophy via upregulation of IL-1β.
Biosci Rep. 38:BSR201714382018. View Article : Google Scholar
|
|
69
|
Aluganti Narasimhulu C and Singla DK:
Amelioration of diabetes-induced inflammation mediated pyroptosis,
sarcopenia, and adverse muscle remodelling by bone morphogenetic
protein-7. J Cachexia Sarcopenia Muscle. 12:403–420. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
He S, Ma C, Zhang L, Bai J, Wang X, Zheng
X, Zhang J, Xin W, Li Y, Jiang Y, et al: GLI1-mediated pulmonary
artery smooth muscle cell pyroptosis contributes to hypoxia-induced
pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol.
318:L472–L482. 2020. View Article : Google Scholar
|
|
71
|
Cero FT, Hillestad V, Sjaastad I, Yndestad
A, Aukrust P, Ranheim T, Lunde IG, Olsen MB, Lien E, Zhang L, et
al: Absence of the inflammasome adaptor ASC reduces hypoxia-induced
pulmonary hypertension in mice. Am J Physiol Lung Cell Mol Physiol.
309:L378–L387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhang M, Xin W, Yu Y, Yang X, Ma C, Zhang
H, Liu Y, Zhao X, Guan X, Wang X and Zhu D: Programmed death-ligand
1 triggers PASMCs pyroptosis and pulmonary vascular fibrosis in
pulmonary hypertension. J Mol Cell Cardiol. 138:23–33. 2020.
View Article : Google Scholar
|
|
73
|
Udjus C, Cero FT, Halvorsen B, Behmen D,
Carlson CR, Bendiksen BA, Espe EKS, Sjaastad I, Løberg EM, Yndestad
A, et al: Caspase-1 induces smooth muscle cell growth in
hypoxia-induced pulmonary hypertension. Am J Physiol Lung Cell Mol
Physiol. 316:L999–L1012. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zha LH, Zhou J, Li TZ, Luo H, He JN, Zhao
L and Yu ZX: NLRC3: A novel noninvasive biomarker for pulmonary
hypertension diagnosis. Aging Dis. 9:843–851. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Chai SD, Li ZK, Liu R, Liu T, Dong MF,
Tang PZ, Wang JT and Ma SJ: The role of miRNA-155 in
monocrotaline-induced pulmonary arterial hypertension through
c-Fos/NLRP3/caspase-1. Mol Cell Toxicol. 16:311–320. 2020.
View Article : Google Scholar
|
|
76
|
Tang B, Chen G, Liang M, Yao J and Wu Z:
Ellagic acid prevents monocrotaline-induced pulmonary artery
hypertension via inhibiting NLRP3 inflammasome activation in rats.
Int J Cardiol. 180:134–141. 2015. View Article : Google Scholar
|
|
77
|
Lin J, Li H, Yang M, Ren J, Huang Z, Han
F, Huang J, Ma J, Zhang D, Zhang Z, et al: A role of RIP3-mediated
macrophage necrosis in atherosclerosis development. Cell Rep.
3:200–210. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Guo N, Zhou H, Zhang Q, Fu Y, Jia Q, Gan
X, Wang Y, He S, Li C, Tao Z, et al: Exploration and bioinformatic
prediction for profile of mRNA bound to circular RNA
BTBD7_hsa_circ_0000563 in coronary artery disease. BMC Cardiovasc
Disord. 24:712024. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Meng L, Jin W, Wang Y, Huang H, Li J and
Zhang C: RIP3-dependent necrosis induced inflammation exacerbates
atherosclerosis. Biochem Biophys Res Commun. 473:497–502. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Karunakaran D, Geoffrion M, Wei L, Gan W,
Richards L, Shangari P, DeKemp EM, Beanlands RA, Perisic L,
Maegdefessel L, et al: Targeting macrophage necroptosis for
therapeutic and diagnostic interventions in atherosclerosis. Sci
Adv. 2:e16002242016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Meng L, Jin W and Wang X: RIP3-mediated
necrotic cell death accelerates systematic inflammation and
mortality. Proc Natl Acad Sci USA. 112:11007–11012. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Rasheed A, Robichaud S, Nguyen MA,
Geoffrion M, Wyatt H, Cottee ML, Dennison T, Pietrangelo A, Lee R,
Lagace TA, et al: Loss of MLKL (Mixed Lineage Kinase Domain-Like
Protein) decreases necrotic core but increases macrophage lipid
accumulation in atherosclerosis. Arterioscler Thromb Vasc Biol.
40:1155–1167. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Akhtar S, Hartmann P, Karshovska E,
Rinderknecht FA, Subramanian P, Gremse F, Grommes J, Jacobs M,
Kiessling F, Weber C, et al: Endothelial hypoxia-inducible
Factor-1α promotes atherosclerosis and monocyte recruitment by
upregulating MicroRNA-19a. Hypertension. 66:1220–1226. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Luedde M, Lutz M, Carter N, Sosna J,
Jacoby C, Vucur M, Gautheron J, Roderburg C, Borg N, Reisinger F,
et al: RIP3, a kinase promoting necroptotic cell death, mediates
adverse remodelling after myocardial infarction. Cardiovasc Res.
103:206–216. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Yang Z, Li C, Wang Y, Yang J, Yin Y, Liu
M, Shi Z, Mu N, Yu L and Ma H: Melatonin attenuates chronic pain
related myocardial ischemic susceptibility through inhibiting
RIP3-MLKL/CaMKII dependent necroptosis. J Mol Cell Cardiol.
125:185–194. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Guo X, Yin H, Li L, Chen Y, Li J, Doan J,
Steinmetz R and Liu Q: Cardioprotective role of tumor necrosis
factor receptor-associated factor 2 by suppressing apoptosis and
necroptosis. Circulation. 136:729–742. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Li L, Chen Y, Doan J, Murray J, Molkentin
JD and Liu Q: Transforming growth factor β-activated kinase 1
signaling pathway critically regulates myocardial survival and
remodeling. Circulation. 130:2162–2172. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhang DY, Wang BJ, Ma M, Yu K, Zhang Q and
Zhang XW: MicroRNA-325-3p protects the heart after myocardial
infarction by inhibiting RIPK3 and programmed necrosis in mice. BMC
Mol Biol. 20:172019. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Yue LJ, Zhu XY, Li RS, Chang HJ, Gong B,
Tian CC, Liu C, Xue YX, Zhou Q, Xu TS and Wang DJ: S-allyl-cysteine
sulfoxide (alliin) alleviates myocardial infarction by modulating
cardiomyocyte necroptosis and autophagy. Int J Mol Med.
44:1943–1951. 2019.PubMed/NCBI
|
|
90
|
Liu J, Wu P, Wang Y, Du Y, A N, Liu S,
Zhang Y, Zhou N, Xu Z and Yang Z: Ad-HGF improves the cardiac
remodeling of rat following myocardial infarction by upregulating
autophagy and necroptosis and inhibiting apoptosis. Am J Transl
Res. 8:4605–4627. 2016.PubMed/NCBI
|
|
91
|
Škėmienė K, Jablonskienė G, Liobikas J and
Borutaitė V: Protecting the heart against
ischemia/reperfusion-induced necrosis and apoptosis: The effect of
anthocyanins. Medicina (Kaunas). 49:84–88. 2013.PubMed/NCBI
|
|
92
|
Szobi A, Gonçalvesová E, Varga ZV, Leszek
P, Kuśmierczyk M, Hulman M, Kyselovič J, Ferdinandy P and Adameová
A: Analysis of necroptotic proteins in failing human hearts. J
Transl Med. 15:862017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Marunouchi T, Nishiumi C, Iinuma S, Yano E
and Tanonaka K: Effects of Hsp90 inhibitor on the RIP1-RIP3-MLKL
pathway during the development of heart failure in mice. Eur J
Pharmacol. 898:1739872021. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Yang J, Zhang F, Shi H, Gao Y, Dong Z, Ma
L, Sun X, Li X, Chang S, Wang Z, et al: Neutrophil-derived advanced
glycation end products-Nε-(carboxymethyl) lysine promotes
RIP3-mediated myocardial necroptosis via RAGE and exacerbates
myocardial ischemia/reperfusion injury. FASEB J. 33:14410–14422.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Zhang T, Zhang Y, Cui M, Jin L, Wang Y, Lv
F, Liu Y, Zheng W, Shang H, Zhang J, et al: CaMKII is a RIP3
substrate mediating ischemia- and oxidative stress-induced
myocardial necroptosis. Nat Med. 22:175–182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Koshinuma S, Miyamae M, Kaneda K, Kotani J
and Figueredo VM: Combination of necroptosis and apoptosis
inhibition enhances cardioprotection against myocardial
ischemia-reperfusion injury. J Anesth. 28:235–241. 2014. View Article : Google Scholar
|
|
97
|
Xiao G, Zhuang W, Wang T, Lian G, Luo L,
Ye C, Wang H and Xie L: Transcriptomic analysis identifies
Toll-like and Nod-like pathways and necroptosis in pulmonary
arterial hypertension. J Cell Mol Med. 24:11409–11421. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Jarabicová I, Horváth C, Veľasová E, Bies
Piváčková L, Vetešková J, Klimas J, Křenek P and Adameová A:
Analysis of necroptosis and its association with pyroptosis in
organ damage in experimental pulmonary arterial hypertension. J
Cell Mol Med. 26:2633–2645. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Tweedell RE and Kanneganti TD: Advances in
inflammasome research: Recent breakthroughs and future hurdles.
Trends Mol Med. 26:969–971. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Zhao P, Yao R, Du X and Yao Y: Research
progress on the role of PANoptosis in human diseases. Zhonghua Yi
Xue Za Zhi. 102:2549–2554. 2022.
|
|
101
|
Malireddi RKS, Kesavardhana S and
Kanneganti TD: ZBP1 and TAK1: Master Regulators of NLRP3
Inflammasome/Pyroptosis, Apoptosis, and Necroptosis (PAN-optosis).
Front Cell Infect Microbiol. 9:4062019. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Zheng M and Kanneganti TD: The regulation
of the ZBP1-NLRP3 inflammasome and its implications in pyroptosis,
apoptosis, and necroptosis (PANoptosis). Immunol Rev. 297:26–38.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Karki R, Lee S, Mall R, Pandian N, Wang Y,
Sharma BR, Malireddi RS, Yang D, Trifkovic S, Steele JA, et al:
ZBP1-dependent inflammatory cell death, PANoptosis, and cytokine
storm disrupt IFN therapeutic efficacy during coronavirus
infection. Sci Immunol. 7:eabo62942022. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Oberst A, Dillon CP, Weinlich R, McCormick
LL, Fitzgerald P, Pop C, Hakem R, Salvesen GS and Green DR:
Catalytic activity of the caspase-8-FLIP(L) complex inhibits
RIPK3-dependent necrosis. Nature. 471:363–367. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Gurung P, Anand PK, Malireddi RK, Vande
Walle L, Van Opdenbosch N, Dillon CP, Weinlich R, Green DR,
Lamkanfi M and Kanneganti TD: FADD and caspase-8 mediate priming
and activation of the canonical and noncanonical Nlrp3
inflammasomes. J Immunol. 192:1835–1846. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wang L, Zhu Y, Zhang L, Guo L, Wang X, Pan
Z, Jiang X, Wu F and He G: Mechanisms of PANoptosis and relevant
small-molecule compounds for fighting diseases. Cell Death Dis.
14:8512023. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Liu X, Tang AL, Cheng J, Gao N, Zhang G
and Xiao C: RIPK1 in the inflammatory response and sepsis: Recent
advances, drug discovery and beyond. Front Immunol. 14:11141032023.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Qi Z, Zhu L, Wang K and Wang N:
PANoptosis: Emerging mechanisms and disease implications. Life Sci.
333:1221582023. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Tian J, Zhang YY, Peng YW, Liu B, Zhang
XJ, Hu ZY, Hu CP, Luo XJ and Peng J: Polymyxin B reduces brain
injury in ischemic stroke rat through a mechanism involving
targeting ESCRT-III Machinery and RIPK1/RIPK3/MLKL pathway. J
Cardiovasc Transl Res. 15:1129–1142. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Duan X, Liu X, Liu N, Huang Y, Jin Z,
Zhang S, Ming Z and Chen H: Inhibition of keratinocyte necroptosis
mediated by RIPK1/RIPK3/MLKL provides a protective effect against
psoriatic inflammation. Cell Death Dis. 11:1342020. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Meng Y, Davies KA, Fitzgibbon C, Young SN,
Garnish SE, Horne CR, Luo C, Garnier JM, Liang LY, Cowan AD, et al:
Human RIPK3 maintains MLKL in an inactive conformation prior to
cell death by necroptosis. Nat Commun. 12:67832021. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Alaaeldin R, Abdel-Rahman IM, Ali FEM,
Bekhit AA, Elhamadany EY, Zhao QL, Cui ZG and Fathy M: Dual
Topoisomerase I/II Inhibition-Induced Apoptosis and Necro-Apoptosis
in cancer cells by a novel ciprofloxacin derivative via
RIPK1/RIPK3/MLKL activation. Molecules. 27:79932022. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Morgan MJ and Kim YS: Roles of RIPK3 in
necroptosis, cell signaling, and disease. Exp Mol Med.
54:1695–1704. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Zhang YY, Liu WN, Li YQ, Zhang XJ, Yang J,
Luo XJ and Peng J: Ligustroflavone reduces necroptosis in rat brain
after ischemic stroke through targeting RIPK1/RIPK3/MLKL pathway.
Naunyn Schmiedebergs Arch Pharmacol. 392:1085–1095. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Yan ZY, Jiao HY, Chen JB, Zhang KW, Wang
XH, Jiang YM, Liu YY, Xue Z, Ma QY, Li XJ and Chen JX:
Antidepressant mechanism of traditional Chinese medicine formula
Xiaoyaosan in CUMS-induced depressed mouse model via
RIPK1-RIPK3-MLKL mediated necroptosis based on Network Pharmacology
Analysis. Front Pharmacol. 12:7735622021. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Karki R, Sharma BR, Tuladhar S, Williams
EP, Zalduondo L, Samir P, Zheng M, Sundaram B, Banoth B, Malireddi
RKS, et al: Synergism of TNF-α and IFN-γ triggers inflammatory cell
death, tissue damage, and mortality in SARS-CoV-2 infection and
cytokine shock syndromes. bioRxiv [Preprint] 2020.10.29.361048.
2020.
|
|
117
|
Ma W, Chen X, Wu X, Li J, Mei C, Jing W,
Teng L, Tu H, Jiang X, Wang G, et al: Long noncoding RNA SPRY4-IT1
promotes proliferation and metastasis of hepatocellular carcinoma
via mediating TNF signaling pathway. J Cell Physiol. 235:7849–7862.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Kawasaki T and Kawai T: Toll-Like receptor
signaling pathways. Front Immunol. 5:4612014. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Damoogh S, Vosough M, Hadifar S, Rasoli M,
Gorjipour A, Falsafi S and Behrouzi A: Evaluation of E. coli
Nissle1917 derived metabolites in modulating key mediator genes of
the TLR signaling pathway. BMC Res Notes. 14:1562021. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Azam S, Jakaria M, Kim IS, Kim J, Haque ME
and Choi DK: Regulation of Toll-like receptor (TLR) signaling
pathway by polyphenols in the treatment of Age-linked
neurodegenerative diseases: Focus on TLR4 signaling. Front Immunol.
10:10002019. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Chipurupalli S, Samavedam U and Robinson
N: Crosstalk between ER stress, autophagy and inflammation. Front
Med (Lausanne). 8:7583112021. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Zhu Y, Yu J, Gong J, Shen J, Ye D, Cheng
D, Xie Z, Zeng J, Xu K, Shen J, et al: PTP1B inhibitor alleviates
deleterious microglial activation and neuronal injury after
ischemic stroke by modulating the ER stress-autophagy axis via PERK
signaling in microglia. Aging (Albany NY). 13:3405–3427. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Tait SWG and Green DR: Mitochondria and
cell death: Outer membrane permeabilization and beyond. Nat Rev Mol
Cell Biol. 11:621–632. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Shi FL, Li Q, Xu R, Yuan LS, Chen Y, Shi
ZJ, Li YP, Zhou ZY, Xu LH, Zha QB, et al: Blocking reverse electron
transfer-mediated mitochondrial DNA oxidation rescues cells from
PANoptosis. Acta Pharmacol Sin. 45:594–608. 2024. View Article : Google Scholar
|
|
125
|
Yuan X, Zhang S, Zhong X, Yuan H, Song D,
Wang X, Yu H and Guo Z: The induction of PANoptosis in KRAS-mutant
pancreatic ductal adenocarcinoma cells by a multispecific platinum
complex. SCC. 1978–1984
|
|
126
|
She R, Liu D, Liao J, Wang G, Ge J and Mei
Z: Mitochondrial dysfunctions induce PANoptosis and ferroptosis in
cerebral ischemia/reperfusion injury: From pathology to therapeutic
potential. Front Cell Neurosci. 17:11916292023. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Briard B, Malireddi RKS and Kanneganti TD:
Role of inflammasomes/pyroptosis and PANoptosis during fungal
infection. PLoS Pathog. 17:e10093582021. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Klionsky DJ, Petroni G, Amaravadi RK,
Baehrecke EH, Ballabio A, Boya P, Bravo-San Pedro JM, Cadwell K,
Cecconi F, Choi AMK, et al: Autophagy in major human diseases. EMBO
J. 40:e1088632021. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Debnath J, Gammoh N and Ryan KM: Autophagy
and autophagy-related pathways in cancer. Nat Rev Mol Cell Biol.
24:560–575. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
González-Rodríguez P, Klionsky DJ and
Joseph B: Autophagy regulation by RNA alternative splicing and
implications in human diseases. Nat Commun. 13:27352022. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Klionsky DJ, Abdel-Aziz AK, Abdelfatah S,
Abdellatif A, Abdoli A, Abel S, Abeliovich H, Abildgaard MH,
Princely Abudu Y, Acevedo-Arozena A, et al: Guidelines for the use
and interpretation of assays for monitoring autophagy (4th
edition)1. Autophagy. 17:1–382. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Ajoolabady A, Kaplowitz N, Lebeaupin C,
Kroemer G, Kaufman RJ, Malhi H and Ren J: Endoplasmic reticulum
stress in liver diseases. Hepatology. 77:619–639. 2023. View Article : Google Scholar
|
|
133
|
Ren J, Bi Y, Sowers JR, Hetz C and Zhang
Y: Endoplasmic reticulum stress and unfolded protein response in
cardiovascular diseases. Nat Rev Cardiol. 18:499–521. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Chen X and Cubillos-Ruiz JR: Endoplasmic
reticulum stress signals in the tumour and its microenvironment.
Nat Rev Cancer. 21:71–88. 2021. View Article : Google Scholar :
|
|
135
|
Marciniak SJ, Chambers JE and Ron D:
Pharmacological targeting of endoplasmic reticulum stress in
disease. Nat Rev Drug Discov. 21:115–140. 2022. View Article : Google Scholar
|
|
136
|
Hu C, Wu Z and Li L: Pre-treatments
enhance the therapeutic effects of mesenchymal stem cells in liver
diseases. J Cell Mol Med. 24:40–49. 2020. View Article : Google Scholar
|
|
137
|
Rafiq K, Hanscom M, Valerie K, Steinberg
SF and Sabri A: Novel mode for neutrophil protease cathepsin
G-mediated signaling: Membrane shedding of epidermal growth factor
is required for cardiomyocyte anoikis. Circ Res. 102:32–41. 2008.
View Article : Google Scholar
|
|
138
|
Yuan Z, Li Y, Zhang S, Wang X, Dou H, Yu
X, Zhang Z, Yang S and Xiao M: Extracellular matrix remodeling in
tumor progression and immune escape: from mechanisms to treatments.
Mol Cancer. 22:482023. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Weems A D, Welf ES, Dr Iscoll MK , Zhou
FY, Mazloom-Farsibaf H, Chang BJ, Murali VS, Gihana GM, Weiss BG,
Chi J, et al: Blebs promote cell survival by assembling oncogenic
signalling hubs. Nature. 615:517–525. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Nakad A, Jonard P, Geubel A, Warzee P,
Coppens JP, Dehennin JP and Dive C: CA 19-9 in neoplasms.
Comparison with CEA. Acta Gastroenterol Belg. 50:36–44. 1987.In
French. PubMed/NCBI
|
|
141
|
Taylor MJ: Clinical cryobiology of
tissues: Preservation of corneas. Cryobiology. 23:323–353. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Paolillo M, Galiazzo MC, Daga A, Ciusani
E, Serra M, Colombo L and Schinelli S: An RGD small-molecule
integrin antagonist induces detachment-mediated anoikis in glioma
cancer stem cells. Int J Oncol. 53:2683–2694. 2018.PubMed/NCBI
|
|
143
|
Bourguignon LYW: Matrix Hyaluronan-CD44
interaction activates MicroRNA and LncRNA signaling associated with
chemoresistance, invasion, and tumor progression. Front Oncol.
9:4922019. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Zhu P, Lu H, Wang M, Chen K, Chen Z and
Yang L: Targeted mechanical forces enhance the effects of tumor
immunotherapy by regulating immune cells in the tumor
microenvironment. Cancer Biol Med. 20:44–55. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Tong J, Lan XT, Zhang Z, Liu Y, Sun DY,
Wang XJ, Ou-Yang SX, Zhuang CL, Shen FM, Wang P and Li DJ:
Ferroptosis inhibitor liproxstatin-1 alleviates metabolic
dysfunction-associated fatty liver disease in mice: Potential
involvement of PANoptosis. Acta Pharmacol Sin. 44:1014–1028. 2023.
View Article : Google Scholar :
|














