Introduction
CD150, also termed signaling lymphocyte activation
molecule, is a type I transmembrane glycoprotein that belongs to
the CD2/CD150 family of the immunoglobulin (Ig) superfamily of
proteins (1-3). High CD150 expression is frequently
associated with B cell malignancies and is implicated in
immunodeficiency in X-linked lymphoproliferative disease (XLP)
(4-6). CD150 engagement by an anti-CD150
monoclonal antibody (mAb) enhances cell proliferation and cytokine
production and plays either stimulatory or inhibitory roles in
mediating important regulatory signals (1,7,8).
CD150 is not detected on immature dendritic cells
(DCs), granulocytes, monocytes, red blood cells or natural killer
cells. However, it is induced on mature DCs after activation with
CD40L or IL-1β and on monocytes after activation by
lipopolysaccharides (LPS) or Toll-like receptors (9,10). Peripheral blood monocytes can
differentiate into DCs depending on the environmental factors
encountered during migration from blood to tissues (10). Upon contact with IL-4 and
granulocyte-macrophage colony-stimulating factor (GM-CSF),
monocytes differentiate into immature DCs, and the addition of
IL-6, IL-1β or tumor necrosis factor α (TNF-α) leads to the
maturation of DCs (11). DC
maturation is characterized by the upregulated expression of
costimulatory and accessory molecules, enhanced expression of major
histocompatibility complex (MHC) class I and II molecules and
neoexpression of CD83 (12).
CD150 also functions as a receptor for the cellular
entry of a number of morbilliviruses, including the measles virus
(MV) (2,13). CD150-expressing T cells in the
human immune system are important target cells for wild-type MV
(14,15), and infection of these cells may
contribute to immunosuppression through decreased expression of
soluble IL-2 receptor, interferon-γ (IFN-γ), IL-1β and TNF-α
(16,17).
CD150 is also a critical regulator of immune
responses and plays a significant role in the modulation of
cytokine production, particularly in the context of Epstein-Barr
virus (EBV) infection. The activation of CD150 on EBV-transformed B
cells triggers the production of specific cytokines such as IL-1α
and GM-CSF, which are essential for immune cell differentiation and
activation (6). Despite several
studies showing that CD150 has various immunological functions in
cytokine production and intracellular signaling in T cells and
other cell types (2,5,8),
very little is known about the roles and mechanisms that regulate
CD150-induced cytokine secretion in virus-infected B cells or in
B-cell malignancies. CD4+ helper T cells are critical
for germinal center responses, and the activation of these helper T
cells depends on T cell-DC interactions (18-20). Subsequent cognate interactions
between activated B cells and T cells induce germinal center
formation. Serum amyloid P component-deficient mice exhibit marked
defects in B-cell proliferation and germinal-center formation
(21). However, the relationship
between activated B cells and DCs with CD150 still requires
investigation. In the present study, EBV-transformed B cells were
utilized as a model system to improve the understanding of the
effect of cytokines produced by activated B cells on peripheral
blood monocytes. The present study aimed not only to elucidate the
relationship between CD150 and cytokine expression in
EBV-transformed B cells but also to examine the functional role of
various cytokines secreted following cross-linking of CD150 in the
differentiation of peripheral blood monocytes. Additionally, the
complex network of interactions between CD150 activation and
cytokine regulation will be explored, providing insights into the
potential therapeutic targets for EBV-associated conditions.
Materials and methods
Reagents, cell culture and
antibodies
EBV supernatant stock was prepared from an EBV B95-8
marmoset cell line (a gift from Dr B. G. Han, National Genome
Research Institute, National Institute of Health, Seoul, Korea).
Raji cells (cat. no. CCL-86), an EBV+ human Burkitt's
lymphoma cell line, and IM-9 cells (cat. no. CCL-159), an
EBV+ human B lymphoblastoid cell line, were obtained
from the American Type Culture Collection and were maintained in
RPMI-1640 medium (HyClone; Cytiva) supplemented with 10% fetal
bovine serum (FBS; HyClone; Cytiva) and antibiotics in the presence
of 5% CO2 at 37°C. The authenticity of the IM-9 human B
lymphoblastoid cell line utilized in the present study was verified
through human short tandem repeat DNA profiling, and mycoplasma
contamination was tested via PCR at Cosmogenetech, Co., Ltd.
MV core proteins [MV#1: amino acids (a.a) 399-525
(cat. no. GWB-64F3B4); MV#2: a.a 89-165 (cat. no. GWB-9FDC44] and a
recombinant CD150 fusion protein (cat. no. GWB-PPAT45) were
generated by GenWay Biotech, Inc. The purity of the MV core
proteins and recombinant CD150 fusion protein was >95%. An
anti-CD150 mAb (IPO-3; cat. no. MA1-7626) was purchased from
Affinity BioReagents, Inc. UPC-10 (mouse IgG2a; cat. no. M9144) was
purchased from Sigma-Aldrich (Merck KGaA) and used as an isotype
control. PE-conjugated anti-human CD150 (cat. no. 559592),
FITC-conjugated anti-CD20 (cat. no. 555622) and FITC-conjugated
anti-human CD14 (cat. no. 555397), CD1a (cat. no. 555806), CD80
(cat. no. 555683), CD86 (cat. no. 555657), CD83 (cat. no. 556910),
HLA-DR (cat. no. 555811) and CD11c (cat. no. 561355) antibodies
were purchased from BD Biosciences.
Generation of EBV-transformed B
cells
A total of 10 ml peripheral blood was collected from
5 healthy human donors (written informed consent was obtained from
each participant) to establish an EBV-infected B cell line from
normal PBMCs. After 2 h of incubation at 37°C, B cells purified
from normal PBMCs were added to the EBV stock supernatant, and
RPMI-1640 medium was added to achieve 5×105 cells/ml
(22). The cultures were
incubated for 2-4 weeks until clumps of EBV-infected B cells were
visible and the medium turned yellow. Cell phenotypes were
monitored using a FACSCalibur flow cytometer (BD Biosciences)
equipped with CellQuestpro software (version 5.2.1; BD Biosciences)
and confocal laser-scanning microscope (Carl Zeiss AG) after
staining with PE-conjugated anti-human CD150 and FITC-conjugated
anti-CD20 antibodies. For intracellular staining, cells were fixed
with 4% paraformaldehyde at room temperature for 10 min, followed
by permeabilization with 0.1% Triton X-100 in PBS for 5 min at room
temperature. Human blood samples were collected from healthy donors
between January, 2020 and December, 2020 at The Inje University
College of Medicine (Busan, South Korea). The donors ranged in age
from 20 to 50 years and included 4 male and 2 female participants.
Inclusion criteria required that all volunteers be healthy adults
without any chronic illnesses or ongoing infections. The exclusion
criteria included a history of autoimmune diseases, recent
vaccinations or medication use that could affect immune function.
All blood donors provided written consent to participate in the
study. The present study was approved by The Institutional
Bioethics Review Board at the Inje University Busan Paik Hospital
(IRB no. 20-0001). For the activation of purified B cells, the
cells were treated for 24 h with LPS at a concentration of 50
μg/ml, ionomycin at a concentration of 50 ng/ml,
phorbol-12-myristate-13-acetate (PMA) at a concentration of 20
ng/ml, anti-IgM at a concentration of 40 μg/ml or soluble
CD40L (sCD40L) at dilutions of 1:50 and 1:100.
alamarBlue assay
To investigate the effect of CD150 expression on the
viability of EBV-transformed B cells, an alamarBlue assay (Bio-Rad
Laboratories; cat. no. BUF012B) was used to measure cell viability.
EBV-transformed B cells were seeded onto 96-well plates at a
density of 5×104 cells per well and treated with
recombinant CD150 protein at concentrations ranging from 100 ng/ml
to 10 μg/ml for 9 h. As a control, cells were treated with
the UPC-10 isotype control. After 72 h of incubation, 20 μl
alamarBlue solution (10% of the total culture medium) was added to
each well. The fluorescence intensity was measured at 570 nm
(excitation) and 600 nm (emission) using a Fluorometer (Synergy HT;
Bio-Tek Instruments, Inc.) 4 h after the addition of the dye. Each
experiment was performed in triplicate.
CD150 stimulation by antibody
cross-linking, MV proteins and recombinant proteins
After 4 weeks of infection, EBV-transformed B cells
(1×106 cells/ml) were harvested and washed twice with
cold PBS. The cells were resuspended in 100 μl PBS and
incubated with an anti-CD150 mAb (IPO-3; 1 μg/ml) or an
isotype control (UPC-10; 1 μg/ml) at 37°C for 30 min. The
cells were washed with PBS, resuspended in 100 μl PBS and
then incubated with goat anti-mouse IgG (2 μg/ml;
Sigma-Aldrich; Merck KGaA) for 15 min at 37°C. After incubation,
the cells were washed and then cultured in RPMI-1640 medium for an
additional 24 or 48 h at 37°C. For stimulation with MV core
proteins or recombinant CD150, EBV-transformed B cells (4 weeks
after infection, 1×106 cells/ml) were resuspended in 100
μl PBS, incubated with MV#1 (1 μg/ml), MV#2 (1
μg/ml) or recombinant CD150 protein (1 μg/ml) at 37°C
for 1 h, then cultured in RPMI-1640 medium for an additional 24 or
48 h at 37°C.
Quantification of human cytokines by
ELISA
Culture supernatants from CD150-stimulated
EBV-transformed B cells or EBV+ lymphoma cell lines were
concentrated using Amicon Ultra-15 Centrifugal Filter units
(MilliporeSigma). The concentrations of IL-1α, IL-1β, IL-2, IL-4,
IL-5, IL-6, IL-8, IL-10, IL-12, IL-13, IL-17α and GM-CSF in the
10-fold concentrated cell-free culture supernatants were determined
using multi-cytokine ELISA (Multi-Analyte ELISArray; cat. no.
MER-004A; SABiosciences; Qiagen, Inc.) and single cytokine ELISA
(Single Analyte ELISArray; cat. no. 336151; SABiosciences; Qiagen,
Inc.). The data are presented as the mean of the biological
replicates ± standard deviation (SD).
Reverse transcription (RT)-PCR and
RT-quantitative (q)PCR
Total RNA from PBMCs, naïve B cells or
EBV-transformed B cells was isolated using a RNeasy Mini kit
(Qiagen, Inc.). RNA was reverse transcribed into cDNA using
TOPscript™ cDNA synthesis kit (Enzynomics Co., Ltd.). To
investigate various cytokine levels, PCR amplification was
performed using specific primer sets (Bioneer Corporation) for
CD150, IL-1α, IL-1β, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p35,
IL-12p40, IL-17α, GM-CSF and β-actin (Table I). PCR was performed using Prime
Taq Premix (Genetbio Co., Ltd.). The cDNAs were amplified by PCR
under the following conditions: 30 cycles of denaturation at 95°C
for 20 sec, annealing at 56°C for 30 sec and extension at 72°C for
30 sec in a thermal cycler. The PCR products were resolved using a
2% agarose gel stained with ethidium bromide and visualized under
UV light using the multiple Gel DOC system (FUJIFILM). Band
intensity was quantified using ImageJ software (version 1.54g;
National Institutes of Health).
 | Table ISequences of the oligonucleotide
primers used for RT-PCR and RT-qPCR. |
Table I
Sequences of the oligonucleotide
primers used for RT-PCR and RT-qPCR.
| Target | Oligonucleotide,
5′→3′
| Product size
bp |
|---|
| Sense | Antisense |
|---|
| CD150 |
TATCTACATCTGCACCGTGAGC |
TCCTGAGCTGGGAAGGAGT | 288 |
| IL-1α |
CTGCATGGATCAATCTGT |
CCCATGTCAAATTTCACTGC | 369 |
| IL-1β |
CAGCTACGAATCTCCGACCAC |
GGCAGGGAACCAGCATCTTC | 100 |
| IL-4 |
ATGGGTCTCACCTCCCAACTGCTT |
TTTCCAACGTACTCTGGTTGGC | 355 |
| IL-5 |
TCTGAGGATTCCTGTTCCTG |
TTATCCACTCGGTGTTCATT | 248 |
| IL-6 | GTGTTGCCTGCTGCCTTC
CCTG |
CTCTAGGTATACCTCAAACTCCAA | 321 |
| IL-8 | ATGACTTCCAAGCTGGCC
GTGGCT |
TCTCAGCCCTCTTCAAAAACTTCTC | 292 |
| IL-10 |
CTGAGAACCAAGACCCAGACATCAAGG |
GTCAGCTATCCCAGAGCCCCAGATCCG | 327 |
| IL-12p35 |
CTTCACCACTCCCAAAACCTG |
AGCTCATCACTCTATCAATAG | 532 |
| IL-12p40 |
CATTCGCTCCTGCTGCTTCAC |
TACTCCTTGTTGTCCCCTCTG | 266 |
| IL-17 |
ATGACTCCTGGGAAGACCTCATTG |
TTAGGCCACATGGTGGACAATCGG | 156 |
| GM-CSF |
ATGTGGCTGCAGAGCCTGCTGC |
CTGGCTCCCAGCAGTCAAAGGG | 424 |
| β-actin |
ATCCACGAAACTACCTTCAA |
ATCCACACGGAGTACTTGC | 200 |
qPCR was conducted using a SYBR Green kit (Takara
Bio, Inc.), an iCycler thermal real-time PCR system (Bio-Rad
Laboratories, Inc.) and specific primer sets (the same primer sets
used in the conventional RT-PCR; Table I). The cDNAs were amplified by
PCR under the following conditions: Initial denaturation at 95°C
for 10 min, 40 cycles of 95°C for 30 sec, 55°C for 30 sec and 72°C
for 1 min. Only experiments where a distinct single peak was
observed with a melting temperature different than that of the no
template control were used. The relative gene expression compared
with unstimulated cells was determined by the built in algorithm of
the iCycle iQ real-time detection system software (version 3.10;
Bio-Rad Laboratories, Inc.) using an adaptive baseline to determine
the threshold cycle (Cq). mRNA fold induction values were
calculated using the following equations:
ΔCq=Cqtarget-Cqβ-actin,
Δ(ΔCq)=(ΔCqstimulated-ΔCqcontrol), mRNA fold
change=2−Δ(ΔCq) (23). Experiments were performed in
triplicate and data are presented as the mean ± SD.
Human monocyte isolation, differentiation
and cell surface phenotyping
For monocyte isolation by plastic adherence,
1×107 PBMCs per well were seeded into 6-well plates (BD
Biosciences) and allowed to adhere for 4 h in 2 ml complete medium
(CM), consisting of RPMI-1640 supplemented with 10% FBS and
antibiotics, at 37°C in 5% CO2. The non-adherent cells
were removed and the adherent cells were washed three times with
prewarmed CM. The differentiation of peripheral blood monocytes was
induced by culturing the cells for 7 days in CM supplemented with
the concentrated cell culture supernatant from EBV-transformed B
cells treated with MV core protein (MV#1 or MV#2) or from cells
treated with CD150 protein. The phenotype of the cells was
monitored using a FACSCalibur flow cytometer (BD Biosciences)
equipped with CellQuestpro software (version 5.2.1; BD Biosciences)
and a confocal laser-scanning microscope (Carl Zeiss AG) at a ×400
original magnification, using FITC-conjugated anti-human CD1a,
CD14, CD80, CD86, CD83, HLA-DR and CD11c antibodies (BD
Biosciences). Images were acquired using confocal microscopy
software release 3.0 (Carl Zeiss AG).
Blocking differentiation with
neutralizing antibodies
CD14+ monocytes were incubated for 3 days
with culture supernatant derived from UPC-10 mock stimulated or
CD150-stimulated EBV-transformed B cells with or without
neutralizing antibodies against IL-1α (Abcam; cat. no. ab300501) or
GM-CSF (Abcam; cat. no. ab300495) to investigate the function of
cytokines secreted by CD150-stimulated EBV-transformed B cells. The
cells were then harvested and the expression of various cell
surface markers was analyzed using flow cytometry, as
aforementioned.
Small interfering (si)RNA transfection by
electroporation
In total, three different 19 nucleotide CD150
interfering RNA duplexes (with two 3′ end overhanging dT
nucleotides) and negative control siRNA duplexes with either low or
medium GC contents were obtained from Bioneer Corporation. The
three experimentally verified CD150 siRNA sequences were selected
from the Bioneer siRNA database, and the target sequences are
listed in Table II. A
non-specific siRNA labeled with green fluorescence served as a
control for validating the transfection efficiency in each
experiment. Cells were transiently transfected by electroporation
under optimized conditions. Briefly, cells were electroporated with
200 nM siRNA in serum-free medium in a 0.4 cm electroporation
cuvette using the Bio-Rad Gene Pulser Xcell system (Bio-Rad
Laboratories, Inc.). The electroporation pulse protocol included a
single pulse at room temperature for 20 msec, an applied voltage of
250 V, a current measure of 0.1 mA, and a pulse repetition
frequency of 1. The pulsing buffer used had a conductivity of 1.0
S/m and an osmolarity of 270 mOsm. Post-electroporation, cells were
immediately transferred to a recovery medium composed of RPMI-1640
supplemented with 10% FBS and incubated for 24 h at 37°C with 5%
CO2. In the experiments assessing CD150-induced cytokine
production, 24 h after transfection, the cells were either left
untreated (mock group) or treated with an anti-CD150 mAb for 24 h.
For certain experiments, the extracted RNA was analyzed using
RT-qPCR.
| Table IIsiRNAs used in transfection. |
Table II
siRNAs used in transfection.
| Target | Oligonucleotide,
5′→3′
| siRNA
identifier |
|---|
| Sense | Antisense |
|---|
| CD150 |
GUGUCAUCAUGAUUCUCAU(dTdT) |
AUGAGAAUCAUGAUGACAC(dTdT) | siRNA#1 |
| CD150 |
GGUACCUUAUGACCCUGGA(dTdT) |
UCCAGGGUCAUAAGGUACC(dTdT) | siRNA#2 |
| CD150 |
GAGAUCGCUACAAGUUUUA(dTdT) |
UAAAACUUGUAGCGAUCUC(dTdT) | siRNA#3 |
| Negative control
siRNA |
CUAUCAAGUGUCUCGUUGUTT |
ACAACGAGACACUUGAUAGTT | si-control |
Statistical analysis
All data are presented as the mean ± SD, and each
value represents at least three different experiments. The
Shapiro-Wilk test was used to check the normality of the data and
Levene's test verified the homogeneity of variance before one-way
analysis of variance (ANOVA). ANOVA followed by Scheffe's test was
used for the analysis of differences between each treatment
condition. P<0.05 was considered to indicate a statistically
significant difference. Statistical analyses were performed using
SPSS software (version 26.0; IBM Corp.).
Results
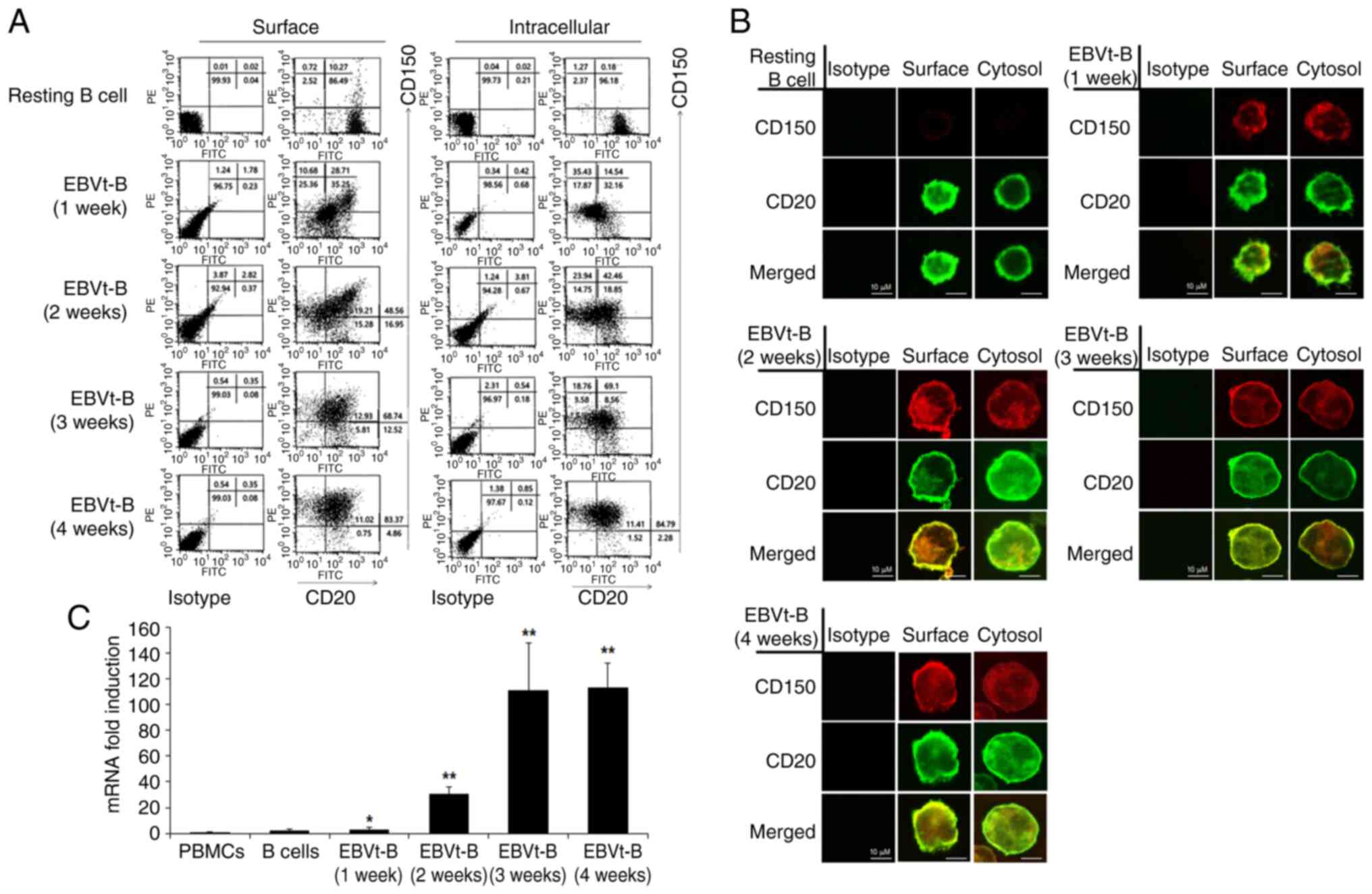
CD150 expression on B cells is induced by
EBV infection
EBV-infected B cells were harvested every week for 4
weeks after the transformation process was complete to determine
CD150 expression patterns using flow cytometry and confocal
microscopy. CD150 was minimally expressed on the surface and in the
cytoplasm of CD20+ resting B cells. The expression of
CD150 slightly increased 1 week after infection and elevated
quickly thereafter. After 4 weeks of EBV infection, most of the
EBV-transformed B cells strongly expressed CD150 molecules on the
cell surface and in the cytoplasm (Fig. 1A and B). We acknowledge that
Total Internal Reflection Fluorescence microscopy could offer more
precise surface signal imaging; however, we chose confocal
microscopy to capture both surface and intracellular details. The
images presented in Fig. 1B are
single plane images, allowing for detailed visualization of the
distribution of the target molecules. This approach complemented
the FACS results shown in Fig.
1A, which quantitatively validated the surface expression.
CD150 mRNA was weakly expressed until 1 week after infection but
was likewise significantly upregulated 4 weeks after EBV infection
(Fig. 1C). It was also observed
that CD150 expression in resting B cells purified from human blood
was induced after treatment with LPS, sCD40L, anti-IgM, PMA and
ionomycin (Fig. S1A). Moreover,
the other EBV+ lymphoma cell lines, IM-9 and Raji, also
expressed high levels of CD150 on their cell surfaces (data not
shown). These results suggested that the expression of CD150 was
induced on B cells over time after EBV infection and could be
upregulated by different immune stimuli, highlighting its potential
role in the immune response to EBV.
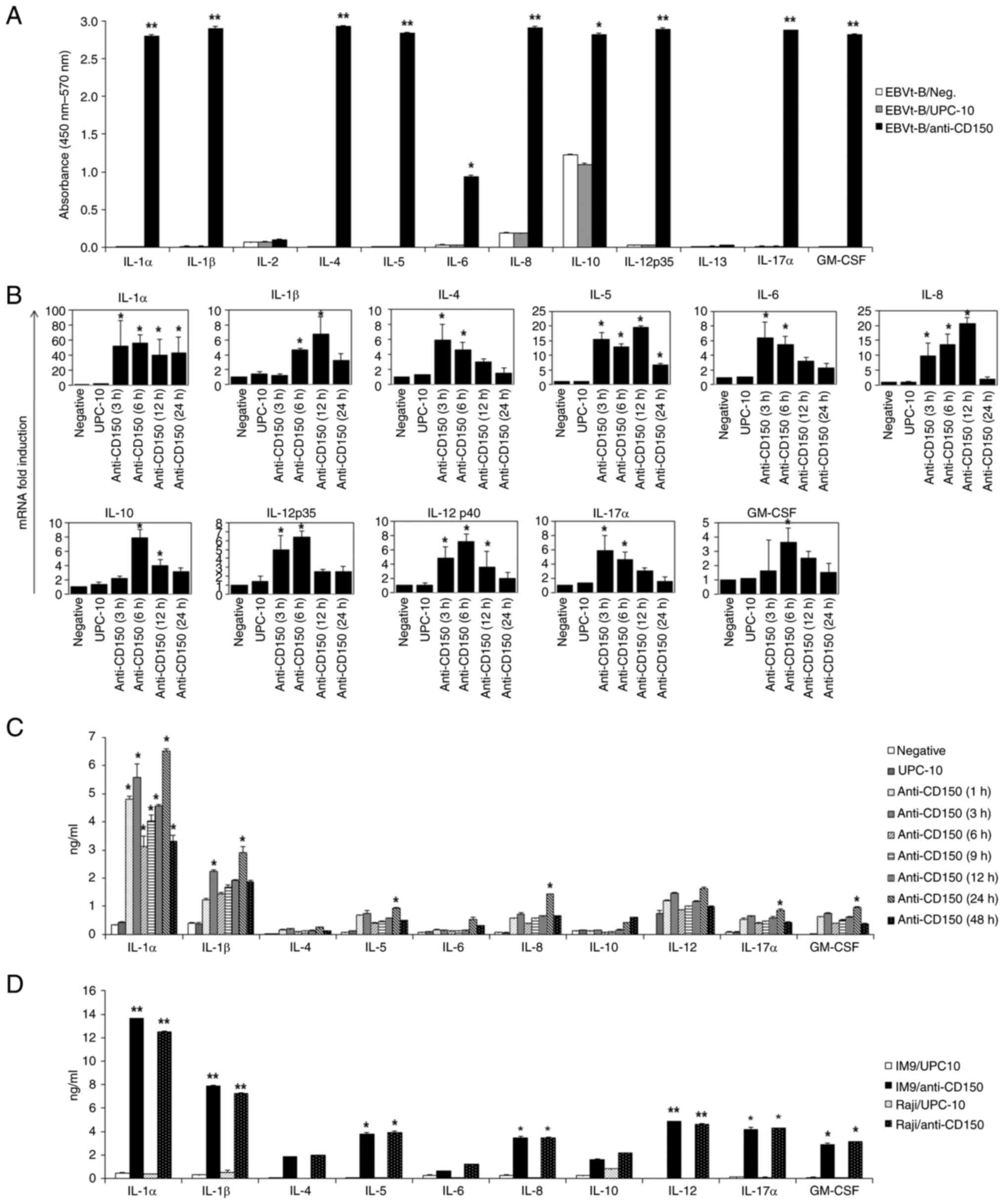
CD150 stimulation by cross-linking
induces the production of various cytokines in EBV-transformed B
cells and EBV+ lymphoma cell lines
The effect of CD150 stimulation on the viability of
EBV-transformed B cells was next investigated. After stimulation
with CD150, EBV-transformed B cells were incubated with an
anti-CD150 mAb (IPO-3), followed by cross-linking with a secondary
antibody. These cells were cultured in the presence of alamarBlue
to detect cell viability with a colorimetric analysis. It was
observed that CD150 expression had a marked effect on the viability
of EBV-transformed B cells (Fig.
S1B). Subsequently, it was next examined whether the improved
cell viability was related to cytokine production, since numerous
cytokines are potent effectors of the maintenance of cell survival
(24). EBV-transformed B cells
were stimulated with IPO-3 or mock stimulated with the UPC-10
isotype control, followed by cross-linking with a secondary
antibody. Multiplex cytokine ELISA, which enabled monitoring of the
changes in the levels of multiple cytokines in the culture
supernatant, revealed significant increases in the secretion of a
number of cytokines, such as IL-1α, IL-1β, IL-4, IL-5, IL-6, IL-8,
IL-10, IL-12A, IL-17α and GM-CSF, after stimulation with CD150, but
the secreted IL-2 and IL-13 levels did not increase (Fig. 2A).
The changes in the expression of these secreted
cytokines were also quantitatively measured using RT-qPCR and
single cytokine ELISA. The cytokine mRNA levels increased at 3 or 6
h in EBV-transformed B cells after stimulation with the anti-CD150
mAb (Fig. 2B). Compared with the
levels of cytokines secreted by EBV-transformed B cells after mock
stimulation with UPC-10, the protein levels of most cytokines were
also notably increased at 24 h following CD150 stimulation
(P<0.05; Fig. 2C). Notably,
the production of IL-1α following CD150 stimulation was markedly
greater than that of other cytokines at the mRNA and protein
levels. Similarly, the EBV+ lymphoma cell lines, IM-9
and Raji, displayed increased production of various cytokines after
24 h of stimulation with CD150. Although the levels of individual
cytokines varied slightly between the cell lines, the overall
pattern of cytokine increases in response to CD150 stimulation was
consistent with that observed in the EBV-transformed B cells
(Fig. 2D). These results
suggested that stimulation of CD150 on B cells infected with EBV by
antibody cross-linking might result in the production of multiple
cytokines, which may affect immune cell proliferation.
Stimulation with MV core proteins or
recombinant CD150 protein induces the production of multiple
cytokines in EBV-transformed B cells and EBV+ lymphoma
cell lines
CD150 acts as a receptor for MV and mediates virus
uptake (14). Therefore, the
impact of CD150 activation on cytokine production was explored by
treating EBV-transformed B cells, Raji cells and IM-9 cells with MV
core protein fragments (MV#1 and #2) or recombinant CD150 protein.
After 24 h of treatment, the cytokine levels in the cell culture
supernatants were measured using a multiple cytokine ELISA kit. The
findings showed that MV#1 (a.a 399-525; 1 μg/ml) and MV#2
(a.a 89-165; 1 μg/ml), as with anti-CD150 mAb, significantly
induced the production of various cytokines, with IL-1α, IL-1β,
IL-10 and IL-12 being notably elevated (Fig. 3A-C). Additionally, recombinant
CD150 protein (1 μg/ml), a natural ligand of CD150, resulted
in cytokine profiles like those observed with anti-CD150 mAb
treatment (Fig. 3D). These
results indicate that CD150 activation, whether by its natural
ligand or by MV core protein fragments, can regulate cytokine
production. This suggests that CD150 potentially functions as a
receptor for MV, mediating similar signaling pathways involved in
cytokine regulation (25).
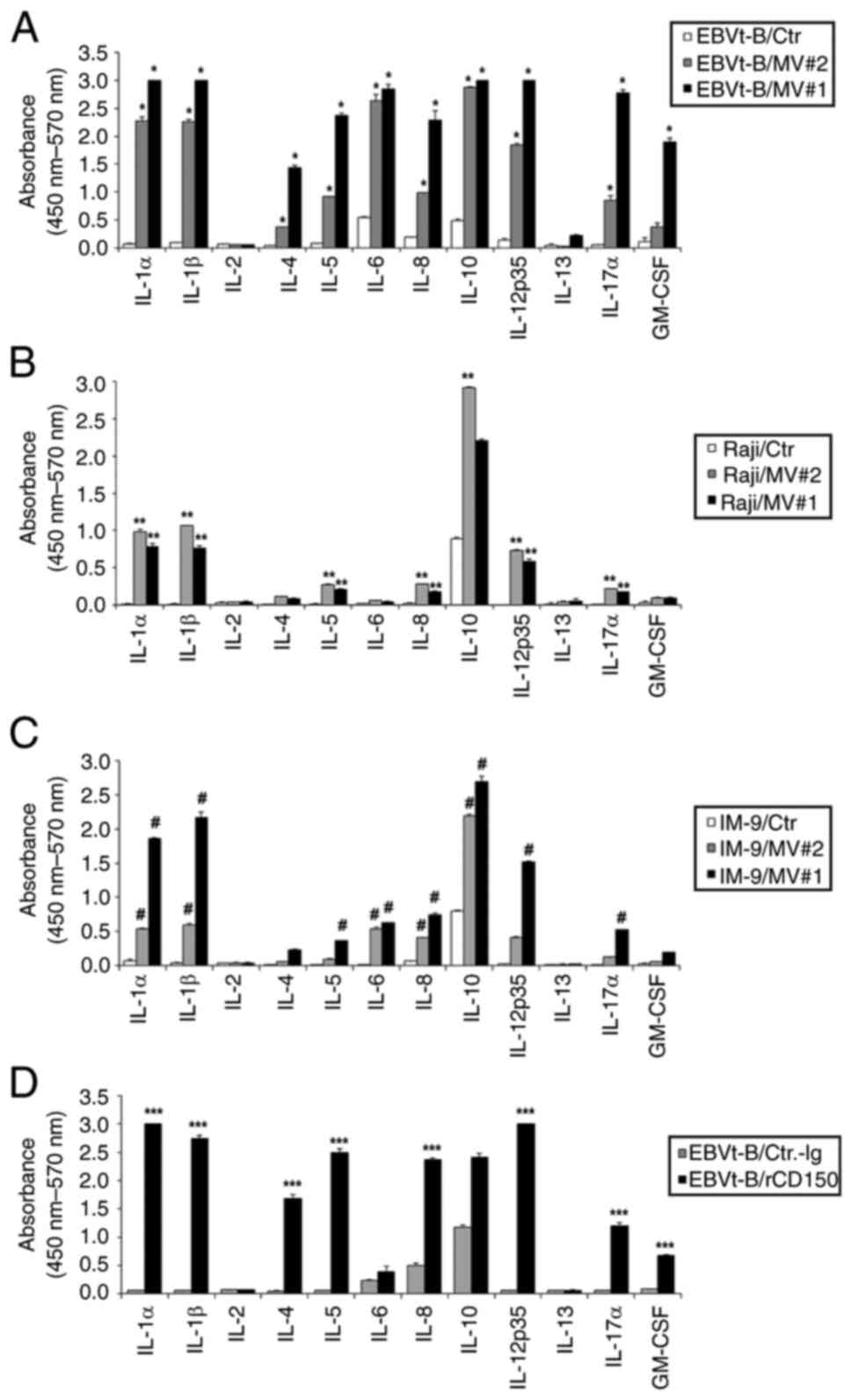 | Figure 3Production of multiple inflammatory
cytokines after treatment with MV or recombinant CD150 protein. The
effects of MV#1 and MV#2 on cytokine secretion in (A) EBVt-B cells
(*P<0.05, treated with MV#2 vs. unstimulated EBV-tB
cells), (B) Raji cells (**P<0.01, treated with MV#1
vs. unstimulated Raji cells) and (C) IM-9 cells
(#P<0.05, treated with MV#2 vs. unstimulated IM-9
cells), as determined using multiplex cytokine ELISA. (D) The
effects of rCD150 on cytokine secretion in EBVt-B cells, as
determined using multiplex cytokine ELISA. Data are presented as
the mean of two independent experiments, and the error bars
represent the SD of the mean. (***P<0.001, rCD150 vs.
unstimulated EBV-tB cells). Comparisons between all individual data
were performed using one-way ANOVA followed by Scheffe's test. Ctr,
control; EBVt-B, Epstein-Barr virus-transformed B (cells); GM-CSF,
granulocyte-macrophage colony-stimulating factor; MV, measles
virus; MV#1, MV core protein amino acids 399-525; MV#2, MV core
protein amino acids 89-165; rCD150, recombinant CD150. |
Cytokines produced following CD150
stimulation induce the differentiation of monocytes into DCs
CD150 has been shown to be involved in the
regulation of a variety of cellular functions, including
proliferation and the expression of certain cytokines (7,8).
However, to the best of our knowledge, the ability of CD150 on
EBV-transformed B cells to regulate the function of immune cells
has not been reported. The effect of the multiple cytokines
secreted after CD150 stimulation on the differentiation of
peripheral blood monocytes was next examined. DCs generated from
monocytes (MoDCs) were used as a model to study the biology of DCs
in vitro. Monocytes were isolated from healthy human PBMCs
and subsequently incubated with culture supernatant derived from
CD150-stimulated EBV-transformed B cells for 7 days to examine MoDC
differentiation. Exposure of these monocytes to culture supernatant
derived from CD150-stimulated EBV-transformed B cells resulted in
cluster formation in a dose-dependent manner (data not shown). The
morphology of cell clusters changed to that typical of DCs on day
3, and the cells retained their DC morphology for at least 7 days
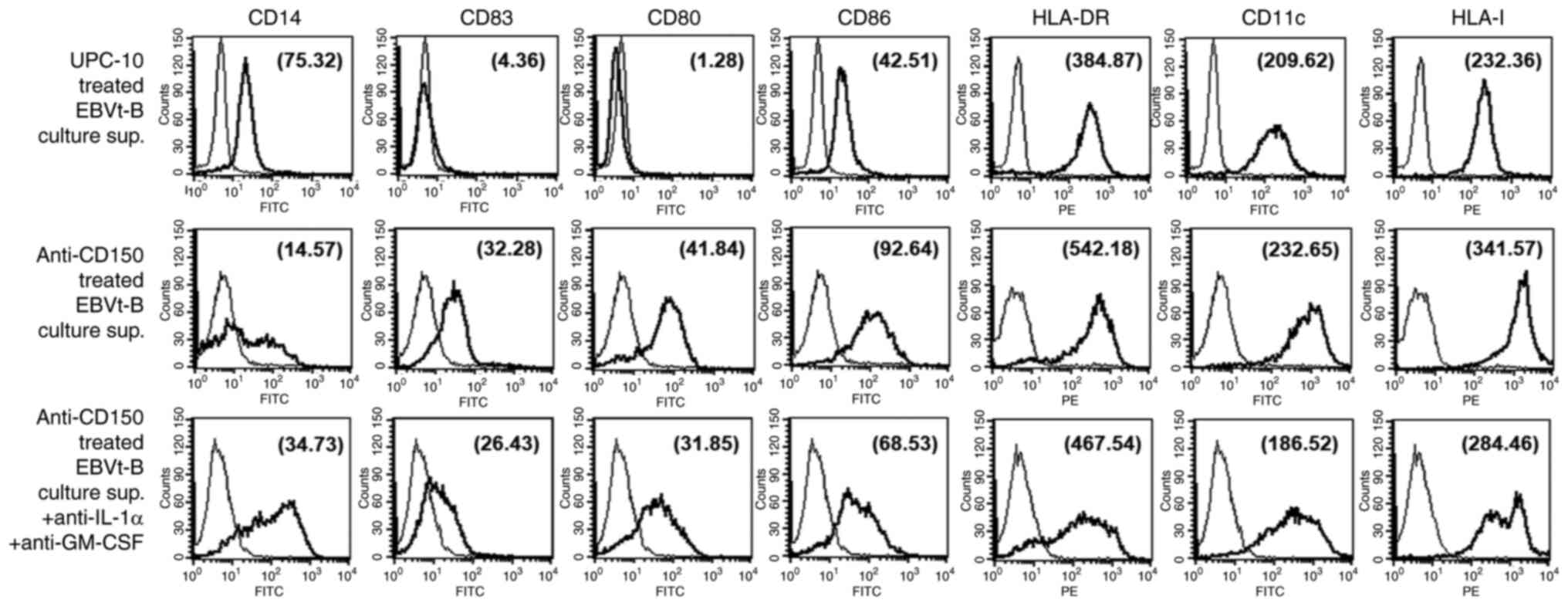
(Fig. 4A). Peripheral blood
monocytes displayed a broad range of elevated surface markers and
morphologies consistent with those of fully mature DCs
(CD83+CD14−CD80+CD86highHLA-DRhigh)
after culture for 3 days in the presence of culture supernatant
derived from CD150-stimulated EBV-transformed B cells. By contrast,
monocytes incubated with culture supernatant derived from
unstimulated EBV-transformed B cells expressed almost no detectable
CD83, CD1a or CD80 and retained relatively high levels of CD14, a
marker of peripheral blood monocytes (Fig. 4B). The phenotypes and morphology
of differentiated monocytes were also observed via fluorescence
microscopy, which revealed decreased CD14 expression and high CD83
expression, consistent with mature DC development (Fig. 4C). It was also observed that
peripheral blood monocytes exhibited increased expression of CD1a,
CD80, CD83, CD86 and HLA-DR and decreased expression of CD14 after
incubation with culture supernatant derived from MV#2-treated
EBV-transformed B cells for 7 days (Fig. 4D). These results suggested that
the cytokines produced by CD150 stimulation may be potent mediators
of the transformation of peripheral blood monocytes into DCs with
mature phenotypes.
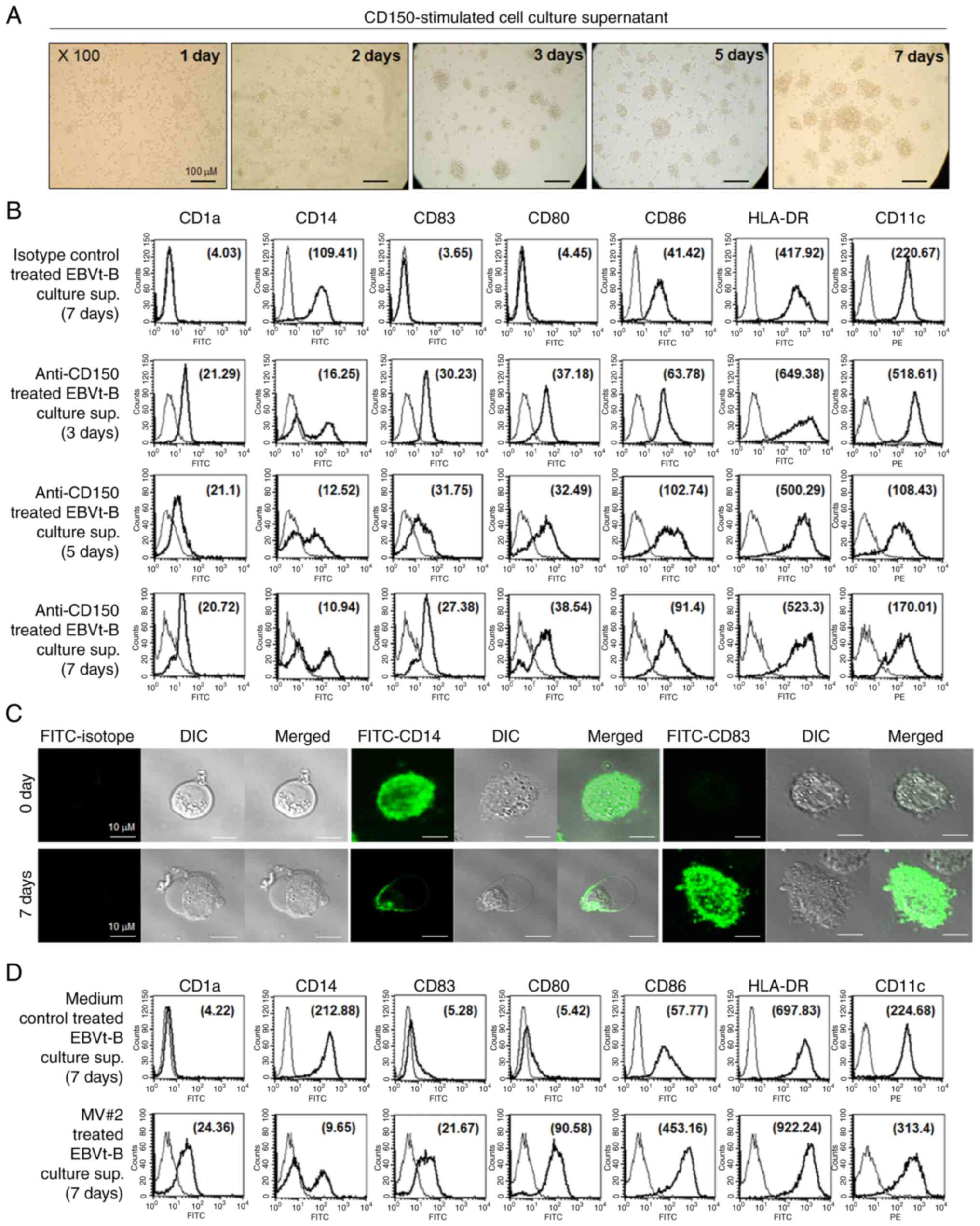 | Figure 4Rapid induction of monocyte
differentiation into DCs by cytokines secreted from
CD150-stimulated EBVt-B cells. (A) Monocytes were incubated with
CD150-stimulated EBVt-B cell culture supernatant (total protein
concentration, 20 ng/ml) for 7 days and examined for the extent of
clustering. Microscopic examination of monocyte clustering and
differentiation into DCs over 7 days (original magnification, ×100;
scale bars, 100 μm). (B) Flow cytometry analysis of surface
markers expressed on peripheral blood monocytes after a 48 h
incubation with culture supernatant derived from CD150-stimulated
EBVt-B cells. Culture supernatant derived from EBVt-B cells mock
stimulated with an isotype control antibody (UPC-10) was used as a
control. The MFI was determined to quantitate the changes in
expression levels of cell surface markers on monocytes in a
time-dependent manner and is indicated in parentheses within the
histogram. (C) Confocal microscopy analysis of monocytes showing
changes in the expression of CD14 and CD83, as well as in
morphology, after an incubation with CD150-stimulated EBVt-B cell
culture supernatant. Scale bars, 10 μm. (D) Flow cytometry
analysis of surface markers on peripheral blood monocytes after a
7-day incubation with culture supernatant derived from
MV#2-stimulated EBVt-B cells (total protein concentration, 20
ng/ml). The MFIs (in parentheses) are denoted. Thin curves
represent the fluorescence of the isotype control. The results
represent four independent experiments. DCs, dendritic cells; DIC,
differential interference contrast; EBVt-B, Epstein-Barr
virus-transformed B (cells); MFI, mean fluorescence intensity;
MV#2, measles virus core protein amino acids 89-165. |
Cytokine-induced differentiation of
monocytes into DCs by CD150-stimulated EBV-transformed B cells is
partially due to IL-1a and GM-CSF
Rapid DC maturation from monocytes can also be
induced by the addition of proinflammatory mediators together with
GM-CSF and IL-4 at the initiation of culture (26,27). The functional role of cytokines
produced by CD150-stimulated EBV-transformed B cells was
investigated using neutralizing antibodies against IL-1α and
GM-CSF, which are the cytokines secreted most abundantly by
EBV-transformed B cells following stimulation with an anti-CD150
antibody and are essential for the differentiation of monocytes
into DCs. The differentiation of monocytes treated with anti-IL-1α
and anti-GM-CSF neutralizing antibodies was partially inhibited, as
shown by the significant reduction in the expression of the DC
surface markers, CD1a, CD83 and CD86 compared with the control
(Fig. 5). This result suggested
that the differentiation of peripheral blood monocytes into DCs
requires IL-1α and GM-CSF as well as other cytokines (such as
IL-1β, IL-4 and IL-6).
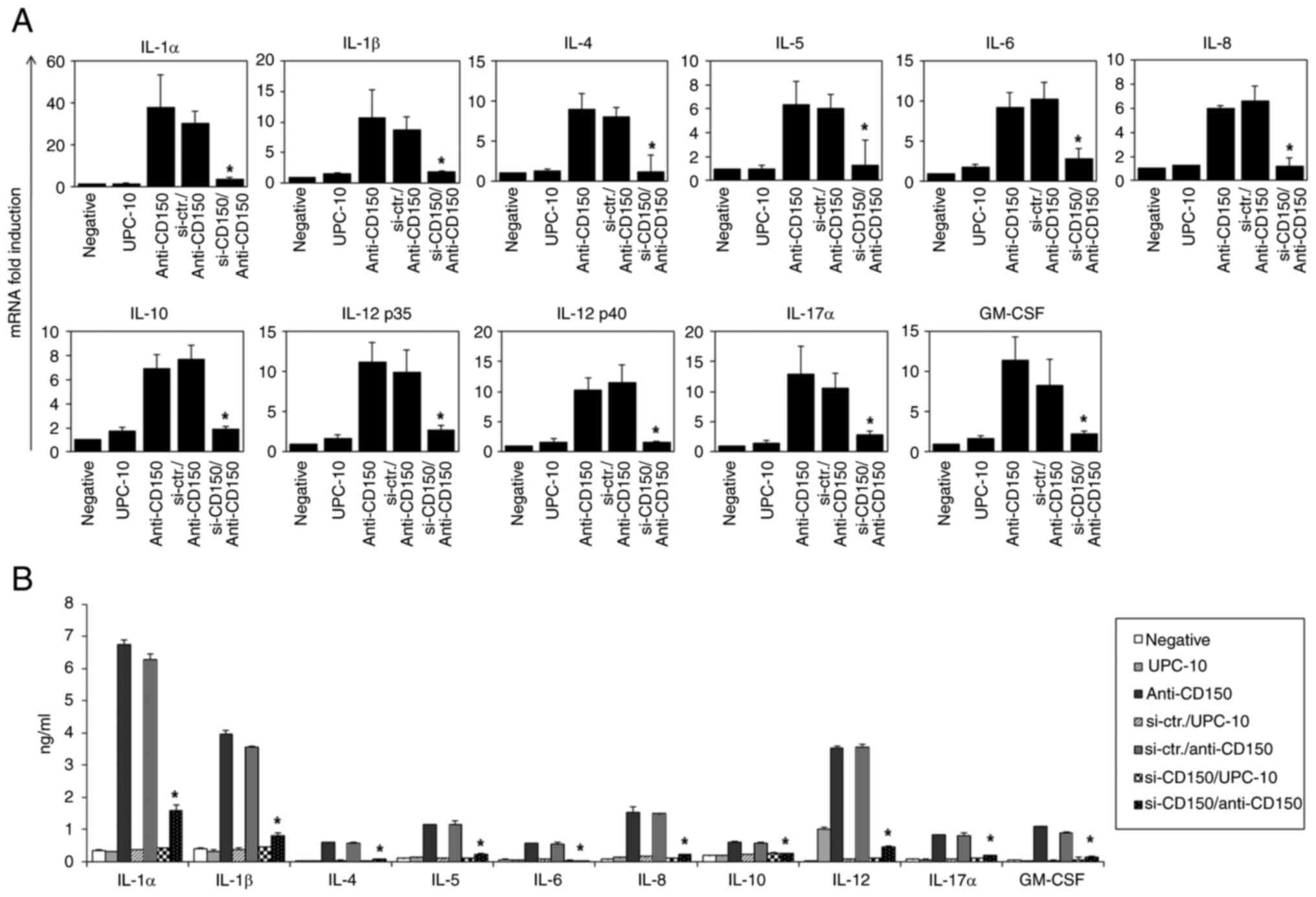
CD150 knockdown in EBV-transformed B
cells results in decreased cytokine production
To determine the role of CD150 in cytokine
regulation, siRNAs were used to knockdown CD150 expression in
EBV-transformed B cells. The efficacy and specificity of three
different siRNAs (si-CD150 #1, #2 and #3) were evaluated and
si-CD150 #3 was selected for further experiments due to its more
notable effect in reducing CD150 expression (Fig. S2). Following CD150 knockdown,
the cells were stimulated with anti-CD150 mAb or isotype control,
and the cytokine levels were measured using RT-qPCR (Fig. 6A) and ELISA (Fig. 6B). The results showed that CD150
knockdown significantly suppressed cytokine production compared
with the control groups, highlighting the critical role of CD150 in
modulating cytokine responses in EBV-transformed B cells (25).
Discussion
The results of the present study revealed that CD150
activation in EBV-transformed B cells triggered the production of
cytokines such as IL-1α and GM-CSF, directly impacting immune cell
differentiation and activation. CD150 activation also affected the
broader cytokine milieu, influencing both pro-inflammatory and
anti-inflammatory responses. The elevation of IL-1α and GM-CSF
suggested that CD150 enhanced the immune response against EBV,
potentially aiding the proliferation and survival of
EBV-transformed B cells. This dual role of CD150 in balancing
immune responses and contributing to EBV persistence underscores
the need for further investigations into targeting this pathway for
therapeutic interventions in EBV-associated conditions.
CD150 is a pivotal signaling molecule to produce
various cytokines, and the present study provides promising and
novel data showing the expression of a variety of cytokine genes
and the secretion of multiple cytokines by EBV-transformed B cells,
which is likely to contribute to the activation of host immune
responses. In humans, CD150 is constitutively expressed on immature
thymocytes and CD45RO+ memory T cells and is rapidly
induced on naïve B and T cells following activation (28,29). CD150 is a self-ligand considered
to have a notable role in adhesion and in signaling in the immune
synapse between T cells and antigen-presenting cells (30). Moreover, CD150 may be involved in
cancer malignancy and autoimmune disorders, including inflammatory
bowel syndrome, rheumatoid arthritis, multiple sclerosis,
hepatitis, systemic lupus erythematosus and XLP (3-5,31). The results of the present study
demonstrated that CD150 was gradually expressed on the B cell
surface during EBV infection and was fully expressed on the surface
after 4 weeks of EBV exposure. Therefore, EBV does appear to
regulate the expression of CD150. It was also observed that CD150
expression in resting B cells purified from human blood was induced
after treatment with LPS, CD40L, anti-IgM, PMA and ionomycin. These
results were consistent with the findings of previous studies that
indicate that CD150 expression is enhanced by mitogens and is
elevated in EBV+ Burkitt's lymphomas and human
T-lymphotropic virus type 1-transformed T cell lines (32,33). The findings of the present study
suggested that EBV infection triggers CD150 expression, similar to
how B cell activation occurs in response to antigens. This insight
into the involvement of CD150 in cytokine production and its
increased expression during EBV infection highlights CD150 as a
potential target for treating EBV-related conditions. Cytokines
play a pivotal role in immune responses and inflammation,
suggesting that strategies targeting CD150 could be effective in
managing EBV-induced B cell transformations and associated cancer
types.
In the present study, it was expected that the
anti-CD150 mAb would increase the apoptosis of EBV-transformed B
cells. This assumption was based on the antiproliferative effects
of CD150, which inhibits the proliferation of Hodgkin's lymphoma
cell lines and induces cell death following stimulation (34). However, the results of the
present study indicated that CD150 stimulation led to an increase
in EBV-transformed B cell viability and the secretion of multiple
cytokines, including both proinflammatory and anti-inflammatory
cytokines, under optimal conditions. Although the expression of
some of these genes (such as IL-10) (35) by EBV-associated LCL cells has
already been reported in the literature, the results of the present
study provide evidence that EBV-transformed B cells potentially
produce a variety of regulatory cytokines. Certain cytokines are
able to regulate the proliferation of EBV-infected B cells directly
(such as IL-1 and IL-10) (35),
while others are notable regulators of leukocyte activity (such as
IL-4, IL-5 and IL-10) (10) or
the differentiation of peripheral blood monocytes (such as IL-4,
IL-12 and GM-CSF) (26,27). This unexpected outcome highlights
the complexity of the interaction of EBV with host cell signaling
pathways, specifically through CD150, suggesting that EBV may
exploit these mechanisms to modulate the immune environment.
Several viruses, including MV, human herpesvirus 6
and mouse hepatitis virus, utilize CD150 receptors to maintain
equilibrium with their host and evade immune surveillance. These
viruses have developed a variety of immune evasion programs based
on the regulation of cell surface molecules that serve as viral
receptors and on the regulation of cytokine expression (36,37). CD150, which possesses Ig
superfamily domains or complement control protein domains, acts as
a receptor for MV and mediates virus uptake. In the present study,
incubation of EBV-transformed B cells, Raji cells and IM-9 cells
with MV core protein #1 or #2 caused the cells to produce multiple
cytokines. Specifically, IL-1α, IL-1β, IL-10 and IL-12 levels were
highly increased, while the levels of secreted IL-5, IL-8 and
IL-17α increased slightly. IL-4 and GM-CSF levels were also
slightly elevated, while IL-2 was not detected and the IL-13 level
was only slightly increased following stimulation with MV #2. In
the present study, it was also determined that inhibition of CD150
using siRNAs might be a potential strategy for blocking the
production of multiple cytokines, although further research is
needed to fully understand the implications of these interventions
for both EBV-associated diseases and normal immune functions.
Certain cancer cells have the capacity to produce
multiple cytokines as growth factors, and at least some of these
growth factors and their receptors may act in tumor cells via
paracrine and/or autocrine loop mechanisms, either by extracellular
release of the growth factor or by action of the tumor itself
(38). For instance, GM-CSF, a
key growth factor for the differentiation of DCs, stimulates the
proliferation and growth of some malignant hematopoietic cell
types, including acute leukemia and lymphomas (39), and receptors for GM-CSF are
present on some leukemic cell lines (40). Typically, DCs used in
experimental and clinical fusion trials are obtained primarily from
standard in vitro 7-day cultures. The standard 7-day culture
protocol yields mature DCs with the ability to stimulate primary
antigen-specific Th1-type immune responses, induce the production
of IFN-γ and activate autologous naïve T cells (41). Moreover, Hanazawa et al
(42) reported that IL-1α and
IL-1β potently induce the differentiation of the macrophage-like
tumor cell line, P388D1. However, in the present study, typical DC
features were observed within 2 days after normal monocytes were
exposed to the CD150-stimulated EBV-transformed B cell culture
supernatant. In addition, IL-1α was the cytokine secreted at the
highest level by EBV-transformed B cells. It was also found that
the differentiation of monocytes incubated with anti-IL-1α and
anti-GM-CSF neutralizing antibodies was partially inhibited. These
results suggest that the differentiation of peripheral blood
monocytes into DCs requires IL-1α and GM-CSF, as well as other
cytokines (such as IL-1β, IL-4 and IL-6). This accelerated
differentiation, driven by cytokines such as IL-1α and GM-CSF,
suggests that EBV may exploit CD150 signaling to alter the immune
landscape, promoting an environment conducive to its persistence
and the proliferation of transformed cells.
In conclusion, the findings of the present study
highlight the crucial role of CD150 in regulating cytokine
production and immune responses in EBV-transformed B cells. The
observed elevation of cytokines such as IL-1α and GM-CSF suggests
that targeting CD150 could be a promising strategy for managing
EBV-related conditions. Despite these insights, the complexity of
the functions of CD150 requires further research to fully explore
its therapeutic potential in combating EBV-related diseases.
Moreover, the experimental design of the present study did not vary
the infection titer of EBV, focusing instead on observing changes
over time post-infection. This limitation suggests that future
studies should consider varying the infection titer to fully
understand its impact on CD150 expression and cytokine
regulation.
Supplementary Data
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
All authors made a significant contribution to the
present study. IKS performed the investigation and data curation.
DYH acquired funding, wrote the original draft of the manuscript
and contributed to the conception or design of the study and the
acquisition of data. HYK contributed significantly to the
conception and design of the study, edited and revised the
manuscript and read and approved the final version of the
manuscript. All authors read and approved the final version of the
manuscript. HYK and DYH confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
The present study using human blood was approved by
The Institutional Bioethics Review Board at the Inje University
Busan Paik Hospitals (Busan, South Korea; IRB no. 20-0001), and all
donors gave written informed consent for the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have a competing
interest related to the following patent: US patent no. 9845456B2,
'Composition containing complex cytokines derived from EBV-infected
B cells for inducing the maturation of dendritic cells', published
on December 19, 2017. The patent details methods for using CD150 to
modulate immune responses, which are relevant to the findings
presented in this manuscript.
Acknowledgements
Not applicable.
Funding
This study was supported by The National Research Foundation of
Korea grant funded by the Korea government (MSIT) (grant no.
RS-2024-00354069) and supported by Basic Science Research Program
through the National Research Foundation of Korea funded by the
Ministry of Education (grant no. RS-2023-00248268).
References
|
1
|
Cocks BG, Chang CC, Carballido JM, Yssel
H, de Vries JE and Aversa G: A novel receptor involved in T-cell
activation. Nature. 376:260–263. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sidorenko SP and Clark EA: The
dual-function CD150 receptor subfamily: The viral attraction. Nat
Immunol. 4:19–24. 2003. View Article : Google Scholar
|
|
3
|
Farhangnia P, Ghomi SM, Mollazadehghomi S,
Nickho H, Akbarpour M and Delbandi AA: SLAM-family receptors come
of age as a potential molecular target in cancer immunotherapy.
Front Immunol. 14:11741382023. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kis LL, Nagy N, Klein G and Klein E:
Expression of SH2D1A in five classical Hodgkin's disease-derived
cell lines. Int J Cancer. 104:658–661. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sayos J, Wu C, Morra M, Wang N, Zhang X,
Allen D, van Schaik S, Notarangelo L, Geha R, Roncarolo MG, et al:
The X-linked lymphoproliferative-disease gene product SAP regulates
signals induced through the co-receptor SLAM. Nature. 395:462–469.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shcherbina V, Gordiienko I, Shlapatska L,
Gluzman D and Sidorenko S: CD150 and CD180 are negative regulators
of IL-10 expression and secretion in chronic lymphocytic leukemia B
cells. Neoplasma. 68:760–769. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sidorenko SP and Clark EA:
Characterization of a cell surface glycoprotein IPO-3, expressed on
activated human B and T lymphocytes. J Immunol. 151:4614–4624.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aversa G, Chang CC, Carballido JM, Cocks
BG and de Vries JE: Engagement of the signaling lymphocytic
activation molecule (SLAM) on activated T cells results in
IL-2-independent, cyclosporin A-sensitive T cell proliferation and
IFN-gamma production. J Immunol. 158:4036–4044. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Howie D, Okamoto S, Rietdijk S, Clarke K,
Wang N, Gullo C, Bruggeman JP, Manning S, Coyle AJ, Greenfield E,
et al: The role of SAP in murine CD150 (SLAM)-mediated T-cell
proliferation and interferon gamma production. Blood.
100:2899–2907. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Farina C, Theil D, Semlinger B, Hohlfeld R
and Meinl E: Distinct responses of monocytes to Toll-like receptor
ligands and inflammatory cytokines. Int Immunol. 16:799–809. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chapuis F, Rosenzwajg M, Yagello M, Ekman
M, Biberfeld P and Gluckman JC: Differentiation of human dendritic
cells from monocytes in vitro. Eur J Immunol. 27:431–441. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Randolph GJ, Beaulieu S, Lebecque S,
Steinman RM and Muller WA: Differentiation of monocytes into
dendritic cells in a model of transendothelial trafficking.
Science. 282:480–483. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tatsuo H, Ono N and Yanagi Y:
Morbilliviruses use signaling lymphocyte activation molecules
(CD150) as cellular receptors. J Virol. 75:5842–5850. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
de Swart RL, Ludlow M, de Witte L, Yanagi
Y, van Amerongen G, McQuaid S, Yüksel S, Geijtenbeek TB, Duprex WP
and Osterhaus AD: Predominant infection of CD150+ lymphocytes and
dendritic cells during measles virus infection of macaques. PLoS
Pathog. 3:e1782007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Amurri L, Reynard O, Gerlier D, Horvat B
and Iampietro M: Measles virus-induced host immunity and mechanisms
of viral evasion. Viruses. 14:26412022. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hirsch RL, Griffin DE, Johnson RT, Cooper
SJ, Lindo de Soriano I, Roedenbeck S and Vaisberg A: Cellular
immune responses during complicated and uncomplicated measles virus
infections of man. Clin Immunol Immunopathol. 31:1–12. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ward BJ, Johnson RT, Vaisberg A, Jauregui
E and Griffin DE: Cytokine production in vitro and the
lymphoproliferative defect of natural measles virus infection. Clin
Immunol Immunopathol. 61:236–248. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Allen CDC, Okada T and Cyster JG:
Germinal-center organization and cellular dynamics. Immunity.
27:190–202. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Garside P, Ingulli E, Merica RR, Johnson
JG, Noelle RJ and Jenkins MK: Visualization of specific B and T
lymphocyte interactions in the lymph node. Science. 281:96–99.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yin X, Chen S and Eisenbarth SC: Dendritic
cell regulation of T helper cells. Annu Rev Immunol. 39:759–790.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Crotty S, Kersh EN, Cannons J,
Schwartzberg PL and Ahmed R: SAP is required for generating
long-term humoral immunity. Nature. 421:282–287. 2003.PubMed/NCBI
|
|
22
|
Park GB, Kim YS, Lee HK, Song H, Kim S,
Cho DH and Hur DY: Reactive oxygen species and p38 MAPK regulate
Bax translocation and calcium redistribution in salubrinal-induced
apoptosis of EBV-transformed B cells. Cancer Lett. 313:235–248.
2011.PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
|
|
24
|
Ottaviani E, Malagoli D and Franchini A:
Invertebrate humoral factors: Cytokines as mediators of cell
survival. Prog Mol Subcell Biol. 34:1–25. 2004.PubMed/NCBI
|
|
25
|
Hur DY, Park GB, Kim YS, Lee HK and Kim
DJ: Composition containing complex cytokines derived from
EBV-infected B cells for inducing the maturation of dendritic
cells. US Patent US9845456B2. Filed March 20, 2014; issued December
19, 2017.
|
|
26
|
Zou GM and Tam YK: Cytokines in the
generation and maturation of dendritic cells: Recent advances. Eur
Cytokine Netw. 13:186–199. 2002.PubMed/NCBI
|
|
27
|
Fricke I, Mitchell D, Petersen F, Böhle A,
Bulfone-Paus S and Brandau S: Platelet factor 4 in conjunction with
IL-4 directs differentiation of human monocytes into specialized
antigen-presenting cells. FASEB J. 18:1588–1590. 2004.PubMed/NCBI
|
|
28
|
Isomäki P, Aversa G, Cocks BG, Luukkainen
R, Saario R, Toivanen P, de Vries JE and Punnonen J: Increased
expression of signaling lymphocytic activation molecule in patients
with rheumatoid arthritis and its role in the regulation of
cytokine production in rheumatoid synovium. J Immunol.
159:2986–2993. 1997.PubMed/NCBI
|
|
29
|
Minagawa H, Tanaka K, Ono N, Tatsuo H and
Yanagi Y: Induction of the measles virus receptor SLAM (CD150) on
monocytes. J Gen Virol. 82:2913–2917. 2001.PubMed/NCBI
|
|
30
|
Bleharski JR, Niazi KR, Sieling PA, Cheng
G and Modlin RL: Signaling lymphocytic activation molecule is
expressed on CD40 ligand-activated dendritic cells and directly
augments production of inflammatory cytokines. J Immunol.
167:3174–3181. 2001.PubMed/NCBI
|
|
31
|
Tojjari A, Giles FJ, Vilbert M, Saeed A
and Cavalcante L: SLAM modification as an immune-modulatory
therapeutic approach in cancer. Cancers (Basel).
15:48082023.PubMed/NCBI
|
|
32
|
Mavaddat N, Mason DW, Atkinson PD, Evans
EJ, Gilbert RJ, Stuart DI, Fennelly JA, Barclay AN, Davis SJ and
Brown MH: Signaling lymphocytic activation molecule (CDw150) is
homophilic but self-associates with very low affinity. J Biol Chem.
275:28100–28109. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sidorenko SP, Vetrova EP, Yurchenko OV,
Berdova AG, Shlapatskaya LN and Gluzman DF: Monoclonal antibodies
of IPO series against B cell differentiation antigens in leukemia
and lymphoma immunophenotyping. Neoplasma. 39:3–9. 1992.PubMed/NCBI
|
|
34
|
Yurchenko MY, Kovalevska LM, Shlapatska
LM, Berdova GG, Clark EA and Sidorenko SP: CD150 regulates JNK1/2
activation in normal and Hodgkin's lymphoma B cells. Immunol Cell
Biol. 88:565–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Burdin N, Péronne C, Banchereau J and
Rousset F: Epstein-Barr virus transformation induces B lymphocytes
to produce human interleukin 10. J Exp Med. 177:295–304. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tatsuo H, Ono N, Tanaka K and Yanagi Y:
SLAM (CDw150) is a cellular receptor for measles virus. Nature.
406:893–897. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nédellec P, Dveksler GS, Daniels E,
Turbide C, Chow B, Basile AA, Holmes KV and Beauchemin N: Bgp2, a
new member of the carcinoembryonic antigen-related gene family,
encodes an alternative receptor for mouse hepatitis viruses. J
Virol. 68:4525–4537. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rowe JM and Rapoport AP: Hemopoietic
growth factors: A review. J Clin Pharmacol. 32:486–501. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vellenga E, Young DC, Wagner K, Wiper D,
Ostapovicz D and Griffin JD: The effects of GM-CSF and G-CSF in
promoting growth of clonogenic cells in acute myeloblastic
leukemia. Blood. 69:1771–1776. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gasson JC, Kaufman SE, Weisbart RH,
Tomonaga M and Golde DW: High-affinity binding of
granulocyte-macrophage colony-stimulating factor to normal and
leukemic human myeloid cells. Proc Natl Acad Sci USA. 83:669–673.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dauer M, Obermaier B, Herten J, Haerle C,
Pohl K, Rothenfusser S, Schnurr M, Endres S and Eigler A: Mature
dendritic cells derived from human monocytes within 48 h: A novel
strategy for dendritic cell differentiation from blood precursors.
J Immunol. 170:4069–4076. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hanazawa S, Hanaizumi C, Amano S, Hirose
K, Ohmori Y, Kumegawa M, Yamaura K and Kitano S: Inductive effect
of recombinant human interleukin-1 alpha and beta on
differentiation of macrophage-like tumor cell line P388D1. J Cell
Physiol. 136:543–546. 1988.PubMed/NCBI
|