The Src-associated in mitosis 68 kDa (Sam68)
protein, also known as KH domain-containing, RNA-binding, signal
transduction-associated protein 1 (KHDRBS1) due to the gene
encoding this protein in humans, was first identified as a molecule
phosphorylated by the tyrosine kinase c-SRC during mitosis
(1). Sam68 belongs to the signal
transduction and activation of RNA metabolism family of RNA-binding
proteins (RBPs). This group of proteins contains a glycine-rich
protein 33/SAM68/GermLine development defective 1 (also known as
GSG) domain for RNA binding and homodimerization (2). Sam68 contains a set of protein
domains that allow protein-protein interactions and modifications
via multiple pathways. These pathways include six proline-rich
sequences enabling interaction with signaling molecules through SRC
homology (SH)3 and WW domains, and a tyrosine-rich region at the
C-terminus, which form docking sites for signaling proteins that
contain SH2 and/or SH3 domains (2).
Due to its high RNA-binding activity and
protein-protein interaction domains, Sam68 is considered a
multifunctional protein. Sam68 is found in a variety of tissues and
cells and has a key role not only in cytosolic cell signaling
(3), but also in numerous other
processes. These processes include both pre-mRNA and microRNA
(miRNA) processing [e.g., spinal muscular atrophy or
spermatogenesis, respectively (4,5)]; RNA transport [e.g., human
immunodeficiency virus type 1 (6)]; signal transduction [e.g.,
polycystic ovary syndrome (7)];
alternative splicing [e.g., multiple sclerosis (8)]; and cell cycle progression [e.g.,
ovarian or prostate cancers (PC) (9,10)]. On another note, Sam68 is also a
component of structures such as ribonucleoprotein complexes or
subnuclear organelles (11,12). In this sense, Sam68 has a key
role in a variety of cellular events, including cell proliferation
and growth, be it as a tumor suppressor or as a proto-oncogene
regulating cell-cycle progression and apoptosis through
RNA-dependent and RNA-independent mechanisms (11-13). Accordingly, Sam68 overexpression
has been found in different types of cancer, which may ultimately
promote disease progression and metastasis.
The activity of Sam68 in cellular processes is
regulated by post-transcriptional modifications, including
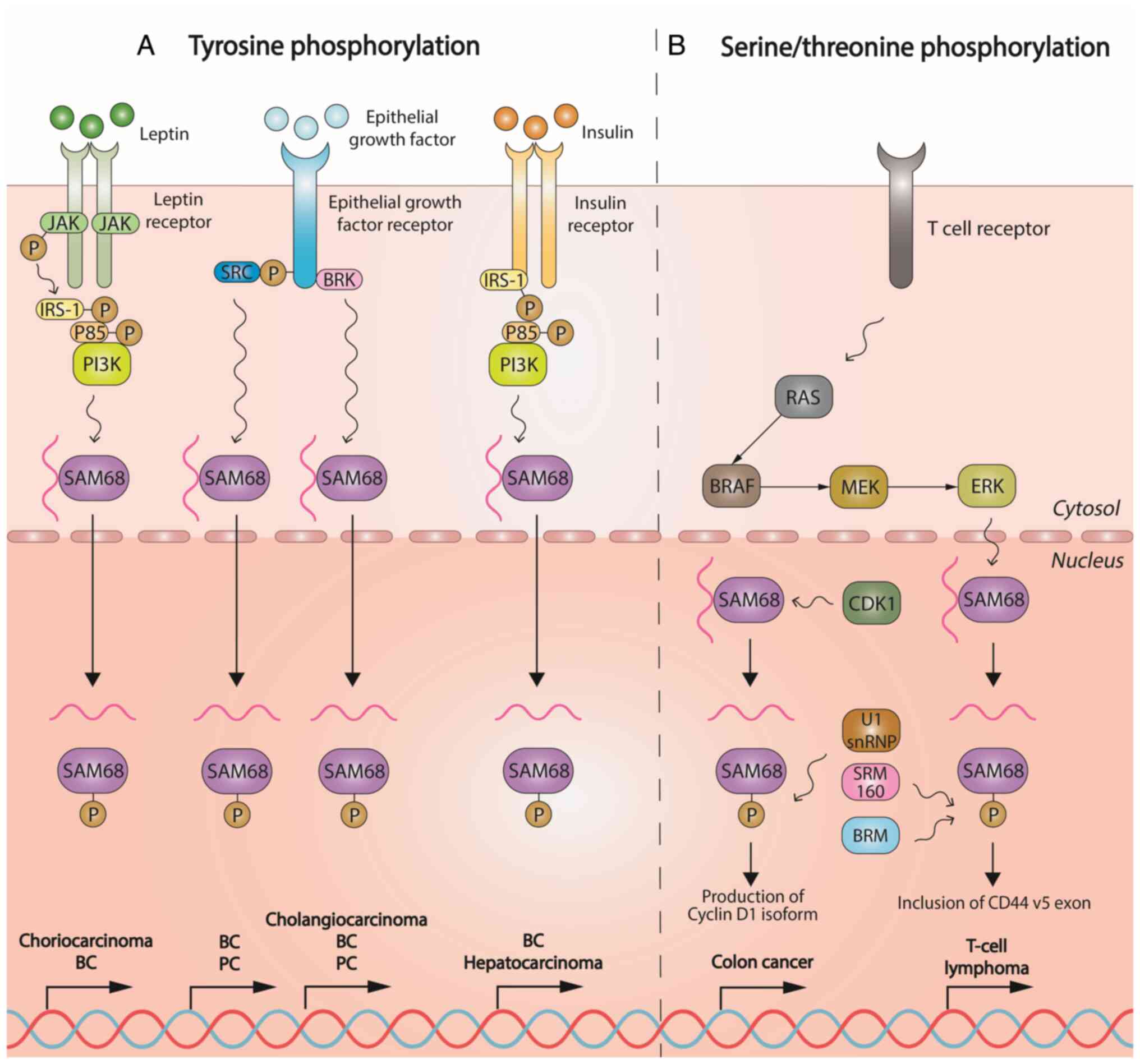
tyrosine, serine and threonine phosphorylation mechanisms (Fig. 1), as well as (de)acetylation,
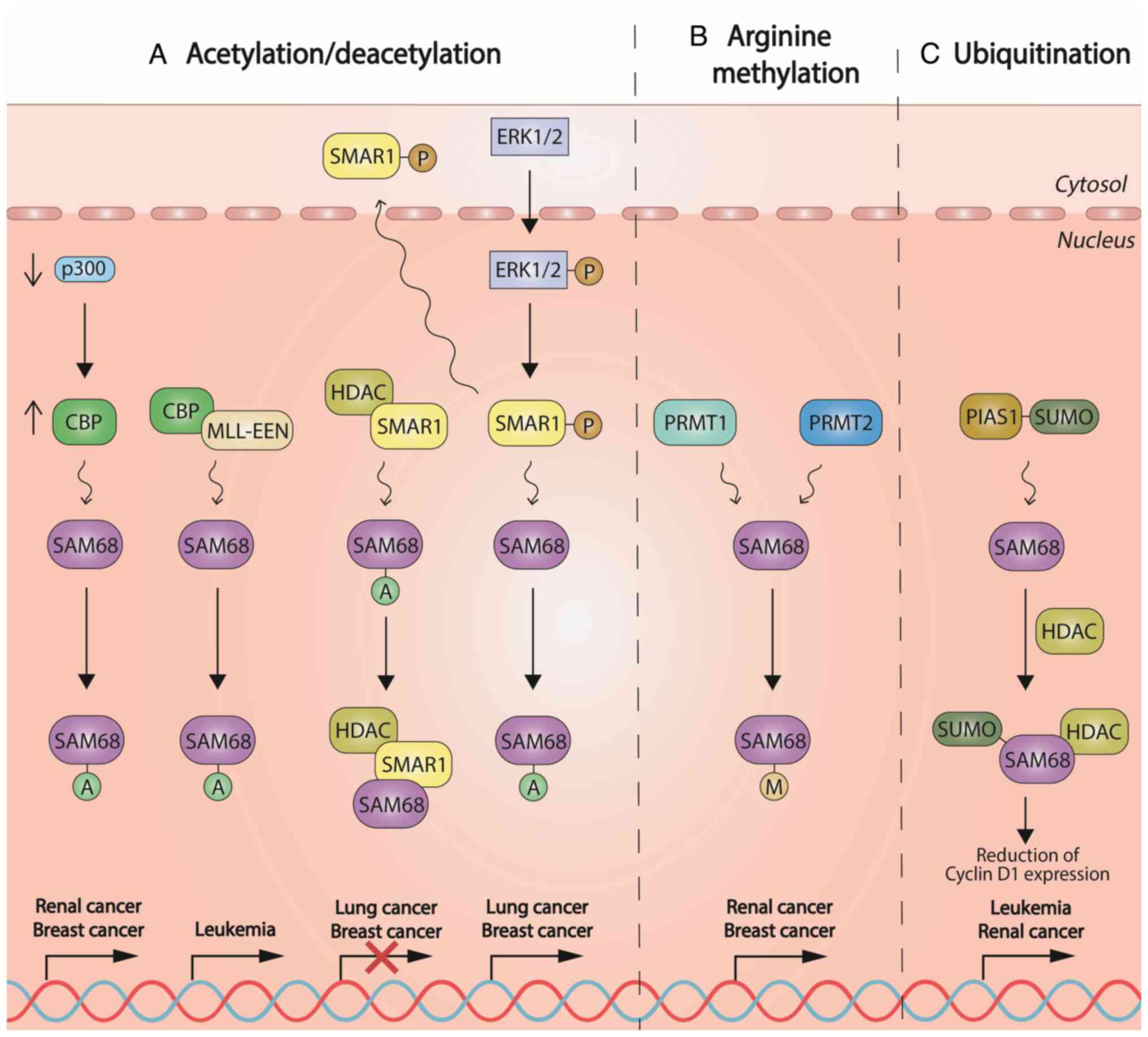
arginine methylation and ubiquitination (Fig. 2) (12). During oncogenesis, these
mechanisms have been shown to ultimately promote the development
and proliferation of malignant cells.
Sam68 seems to play a critical role in cancer,
particularly by tyrosine phosphorylation-mediated mechanisms. In
1994, phosphoprotein Sam68 was first identified and described as a
mitotic target for tyrosine kinase Src, a mediator of epidermal
growth factor (EGF) receptor signaling in breast cancer (BC) cells
(14). A variety of studies have
demonstrated tyrosine phosphorylation of Sam68 by the Src protein
family in numerous settings, including cancer (15-22). The breast tumor kinase (BRK) is
an intracellular tyrosine kinase that is overexpressed in BC and
boosts EGF response. This kinase was shown to phosphorylate Sam68
on all three tyrosine residues in the nuclear localization signal,
with tyrosine 440 being the main Sam68 modulator in BC cells in
vitro (23). In the same
sense, Sam68 tyrosine phosphorylation was found to be augmented in
cholangiocarcinoma (CC) livers and CC cell lines, as compared to
normal livers and human cholangiocyte cells. Elevated Sam68
tyrosine phosphorylation was also associated with elevated levels
of BRK protein and tumor cell proliferation (24). Of note, a study demonstrated that
BRK remains in the cytoplasm of PC cells. However, while Sam68
phosphorylation remained intact, BRK overexpression was not
sufficient to transport BRK into the nucleus, its most frequent
location in normal prostatic epithelial cells (25). Another study examining prostate
tumors reported that Sam68 was only phosphorylated in those tumors
that expressed a truncated form of the c-Kit tyrosine kinase, which
is found in high-grade tumors (26-28).
Hyperleptinemia and hyperinsulinemia induce a
proinflammatory state in obesity that increases BC risk (29). As Sam68 has been found to be
tyrosine-phosphorylated in BC (27,28), its phosphorylation has also been
investigated in patients with these disorders. It was found that
insulin receptor (IR) tyrosine phosphorylates Sam68, which allows
it to interact with the SH2 domains of p85, the regulatory subunit
of phosphoinositide 3-kinase (PI3K) (30). Leptin has been demonstrated to
increase Sam68 tyrosine phosphorylation via Janus kinase/IR
substrate (IRS-1)/p85/PI3K signaling; consequently, leptin
decreases the ability of Sam68 to bind RNA, while it mediates the
survival, growth and proliferation of BC cell lines (31) and JEG-3 choriocarcinoma cells
(32). Finally, these changes
may ultimately promote abnormal placental development and tumor
growth (32,33). Under insulin conditions, IRS-1,
which has been found in the nucleus of BC cells (34), binds Sam68 following tyrosine
phosphorylation by IR in BC cells in vitro (31). However, it remains elusive
whether Sam68 is directly tyrosine-phosphorylated by IR, as
suggested by the results obtained in HTC hepatoma cells (35), and whether Sam68 binds
intracellular or nuclear IRS-1.
Serine/threonine phosphorylation of Sam68 has also
been studied in cancer. Sam68 has been shown to be
threonine-followed-by-proline phosphorylated by the RAS-MAPK/ERK
kinase (MEK)1/2-ERK1/2 pathway in mouse EL4 T-lymphoma cells to
enhance Sam68-dependent exon v5 inclusion in CD44 pre-mRNA
(36). In addition,
serine/threonine has been demonstrated to be phosphorylated by
cyclin-dependent kinase (CDK)1 at T33 and T317. Furthermore, there
is evidence that this protein kinase reduces the ability of Sam68
to bind RNA, as it downregulates its alternative splicing activity,
thus limiting apoptosis and promoting the proliferation of HCT116
colon cells (37).
Protein acetylation is a reversible
post-transcriptional modification involved in a variety of cellular
processes. Its abnormal action has been associated with multiple
disorders, including cancer (38). Acetylation is widely known to
influence the activity of DNA-binding proteins. Of note, Sam68 was
the first RBP reported to be also acetylated (28). Histone deacetylase (HDAC)
inhibition and the overexpression of the acetyltransferase CREB
binding protein (also known as CBP) promote Sam68 acetylation in
renal and, most frequently, in BC cell lines. Hence, CREB-mediated
Sam68 acetylation enhances the ability of Sam68 to bind RNA for
these cell lines, although other acetyltransferases may be involved
in this process (28). CBP also
takes part in mixed-lineage leukemia (MLL) gene translocations
(39,40) and interacts with the extra
eleven-nineteen gene-MLL complex via Sam68 (41,42). Sam68 has been demonstrated to be
acetylated in a p300-dependent manner (43). In colorectal cancer (CRC) cell
lines, butyrate resistance-associated p300 deficiency has been
shown to favor CBP-induced acetylation. Consequently, acetylation
added to Sam68 activity may affect gene expression and
carcinogenesis in CRC cell lines (44).
Of note, ERK-1/2-mediated phosphorylation of the
Scaffold/matrix-associated region-binding protein 1 (SMAR1)
releases the inhibitory SMAR1-HDAC6-Sam68 complex, facilitating
Sam68 acetylation and alternative splicing in BC cells (45).
Protein methylation is involved in a variety of
cellular processes and its aberrant action is implicated in cancer.
Therefore, protein arginine methyltransferases (PRMTs) have gained
interest in numerous tumors as a therapeutic target (46). Sam68 has been shown to
independently recruit PRMT1 and promote RNA splicing (41). By contrast, loss of PRMT1 in
mouse embryonic fibroblasts led to the downregulation of Sam68
arginine methylation (47). It
has also been demonstrated that PRMT1-mediated methylation may be
modulated by the silencing of the human CCR4-associated factor 1 in
human MCF7 BC cells, which also regulated Sam68 methylation
(48). Of note, the protein
PRMT2 has been shown to interact with both Sam68 and PRMT1 to boost
its activity. In addition, Sam68 regulates subcellular PRMT2
localization via the SH3 domain in HeLa cells. These findings
suggest a role of PRMT2 during inflammation in Sam68-mediated
alternative splicing regulation (49).
Sam68 has been demonstrated to be involved in
tumorigenesis by modulating CSC self-renewal. NF-κB, a DNA
transcription factor acting as a key regulator during oncogenesis
(55,56), is involved in the underlying
mechanism (57).
Other studies have shown the influence of Sam68 in
the self-renewal capacity of breast CSCs; these studies focused on
the tumor-derived spheroids responsible for the enrichment of CSCs
or cells with stem cell-related characteristics (58,59). Investigation of mammosphere
formation in NOD/SCID mice revealed that SKBR-3-Sam68 BC cells had
a significantly higher weight compared to SKBR-3-vector tumor
cells. In addition, Sam68 activated the Wnt/β-catenin pathway and
Sam68 protein expression was negatively correlated with miRNA-204
expression (60). MiRNA-204 is a
tumor suppressor miRNA that inhibits cancer cell proliferation and
invasion in multiple types of cancer (61-63), which may be useful for diagnosis
(64). In the same sense, the
Wnt/β-catenin signaling pathway has been found to be activated by
Sam68 in MDA-MB-231 BC-bearing mice (a breast CSC-like model),
compared to MCF-7 BC-bearing mice (a breast non-CSC-like model)
(51). This finding suggests a
stronger role of Sam68 during tumor initiation.
Similarly, stem-like cells from spheroids in
triple-negative BC-bearing mice expressed high levels of Myc, which
required the presence of Sam68 for DNA-damage repair (65). Also, Sam68 gene inhibition caused
defects in the poly(ADP-ribose) polymerase (PARP)-induced PAR chain
synthesis upon DNA damage. This resulted in cancer cell death,
delayed tumor growth and improved survival rates (65), thus demonstrating a key role of
Sam68 in CSC-mediated oncogenesis.
Of note, certain proteins with tumor-promoting
properties have been found to be associated with Sam68 in patients
with BC but not in BC murine models. In this sense, disease-free
survival of patients with BC has been associated with BRK6
expression in tumor tissue. In turn, BRK6 expression was
significantly correlated with the expression of Sam68 and other
signaling proteins such as phosphatase and tensin homolog, ERK and
p-ERK (70). MMP-9 has been
proven to correlate with Sam68 expression along with lymph node
metastasis, suggesting a role in epithelial-mesenchymal transition
(69).
In light of the evidence available, Sam68 plays a
major role in all stages of oncogenesis. Indeed, Sam68 has been
identified as an independent negative prognostic factor for BC
(71), particularly in luminal A
and triple-negative BC (65),
and emerges as a promising therapeutic target in this disease.
The association between Sam68 overexpression and PC
cell proliferation and poor outcomes has been consistently
demonstrated in different clinical settings (72,73). However, there is a paucity of
data from murine models as compared to patients (10,26,74-76). Sam68 has been documented to be
phosphorylated by BRK in PC-bearing mice to inhibit the Sam68
RNA-binding capacity and lead to uncontrolled cell cycle
progression (74). Of note,
Sam68 has been detected in the nuclei of luminal epithelial cells
and to be unaltered in high-grade prostate tumors. By contrast,
nuclear localization of BRK correlated with higher BRK activity in
a more differentiated prostate tumor cell line (LNCaP, poorly
differentiated tumorigenic cells), whereas cytoplasmic localization
of BRK correlated with decreased BRK activity in a poorly
differentiated prostate tumor cell line (PC3, more aggressive cell
line) (74).
Regarding LNCaP PC cells, Sam68 has been
phosphorylated and detected in grade 7-9 hypertrophic prostate
tumors that expressed a truncated form of the c-Kit tyrosine kinase
receptor (26). Of note, Sam68
may also interact with the androgen receptor (AR) to co-regulate
AR-dependent transcription and modulate AR-dependent alternative
splicing. These effects are exerted by enhancing the recruitment of
a Sam68-responsive exon transcribed to androgen response elements
within the promoter region of the prostate-specific antigen gene
(76). This activity ultimately
promotes cancer development and progression. In fact,
downregulation of Sam68 has resulted in the reduction of both cell
cycle progression and PC cell proliferation (75,77).
Similarly, in a PC3 murine model, an antitumor
hemisynthetic cardenolide called UNBS1450 showed effectiveness
in vitro and in vivo in impairing c-Myc expression
and disrupting Sam68 nuclear bodies. These nuclear bodies consisted
of Sam68 protein, nucleolar proteins and nucleic acids. According
to the authors, Sam68 nuclear bodies delayed the proliferation of
PC3 cells but not of normal cells in vitro (78). These findings suggest a critical
role of Sam68 in PC, emerging as a promising therapeutic target in
the disease.
High Sam68 expression has also been found in
non-small cell lung cancer (NSCLC) tissue compared to adjacent
non-cancerous tissue. In addition, Sam68 expression correlated with
lymph node metastasis, advanced tumor grade and poor prognosis
(80,83). Similarly, NSCLC cell lines show
high Sam68 expression and its knockdown inhibited tumor cell
proliferation, colony formation and cell cycle progression via
Wnt/β-catenin pathway inhibition (83). By contrast, in lung squamous cell
carcinoma, a type of NSCLC, Sam68 expression was similar in normal
and tumor tissue; however, Sam68 was spliced by the uc001bub
isoform in normal tissue, whereas it was spliced by the uc001bua
and uc001buc isoforms in tumor tissue, suggesting a potential role
for Sam68 as a prognostic marker in this disease (84).
Of note, an electrochemical immunosensor has been
developed for Sam68 protein quantification. The sensor was
successfully tested in lung cancer patients, showing a good
analytical performance and sensitivity, as compared to ELISA kits
(79). This means that Sam68
protein can be tested and quickly quantified in other types of
tumors.
As in BC, Wnt/β-catenin signaling has been
demonstrated to be involved in CSC-mediated tumor initiation via
Sam68 in patients with colon cancer (85). This signaling pathway was found
to be strongly activated by Sam68 in the HT29 colon CSC-like model,
as compared to the SW480 non-CSC-like model (51). In line with these results, Sam68
upregulation was correlated with both increased PAR production and
NF-κB-mediated anti-apoptotic transcription. Furthermore, Sam68
genetic deletion limited the tumor burden. These results suggest a
novel role for this RBP in genotoxic stress-initiated nuclear
signaling, which is crucial for colon tumorigenesis (86). Consistently with those notions
and similarly to LUAD (see above), Sam68 drove glycolysis in CRC
cells via alternative splicing of PKM2, which resulted in CRC cell
proliferation (87).
However, the protumoral role of Sam68 in CRC and
colon tumors may not be as evident as in other types of cancer. In
a study, the lysine-specific methyltransferase Set7/9 methylated
Sam68 and its knockout reduced the levels of Sam68 protein in human
colon cancer cells. This activity resulted in an altered regulation
of the cell cycle and apoptosis, which explains the association
between high levels of Sam68-Set7/9 co-expression and improved
survival rates (88). On another
note, it should be taken into account that the p53 status may
influence the role of Sam68 in tumor development (89). At least in colon cancer in
vivo, Sam68 may not only have a tumor-promoting role in
p53-deficient mice, but also a tumor-suppressive role in mice
expressing wild-type p53 (90).
To a lesser extent, Sam68 has also been studied in
other types of tumors in both mice and humans, and its expression
has been unfailingly associated with cell proliferation,
transformation, tumorigenesis and metastasis.
There is evidence of a high expression of the Sam68
isoform with a shortened 3'untranslated region (3'UTRs) due to
alternative polyadenylation in gastric cancer tissue, as compared
to paracancer tissue. Overexpression of Sam68 3'UTRs drives tumor
progression, as it helps Sam68 miRNA escape from miRNA-mediated
gene inhibition (91).
Therefore, Sam68 3'UTR expression is elevated in gastric cancer,
being associated with a higher grade of malignancy and, ultimately,
with poor prognosis (92). Of
note, in vitro Sam68 knockdown reduced cell cycle
progression and gastric cancer cell migration and invasion
(92). The circular RNA
Epstein-Barr virus (ebv)-circular RNA ribosomal protein S13
(circRPMS1) was overexpressed in EBV-associated gastric carcinoma
(EBVaGC). In addition, the binding of ebv-circRPMS1 to Sam68
activated m6A methyltransferase (METTL3) transcription, resulting
in EBVaGC cell proliferation, migration and invasion. This
mechanism was ultimately associated with distant metastasis and
poor prognosis in clinical EBVaGC samples (93).
In oral tongue squamous cell carcinoma (OTSCC),
Sam68 overexpression was significantly associated with
lymphovascular invasion and pathological cervical lymph node
metastasis in cancer patients (94). Sam68 was also shown to play a
role in an OTSCC murine model. According to a study, high Sam68
expression significantly inhibited cisplatin-induced apoptosis.
This resulted in the induction of anti-apoptotic proteins such as
caspase-9, caspase-3 and PARP, whereas Sam68 silencing markedly
boosted the sensitivity of cancer cells to apoptosis (95).
Sam68 has also been found to be highly expressed in
skin lesions from glioma-associated oncogene family zinc finger
2-transgenic mice with nonmelanoma skin cancer. A study suggested
that Sam68 regulates discoidin domain receptor in keratinocytes and
promotes the growth and survival of cancer cells via the NF-κB
signaling pathway (96). In
addition, Sam68 deficiency in mouse testis has been associated with
infertility and defects in spermatogenesis due to the
downregulation of genes involved in the cell cycle, cell death,
cell-to-cell signaling and interaction, and cancer. This evidence
suggests that Sam68 not only plays a role in the development of
functional male gametes, but it may also be involved in testis
cancer (97). However, the role
of Sam68 in skin and testis cancer has not yet been assessed in
clinical patients.
Sam68 expression has also been analyzed in certain
types of human tumors rather than murine tumors. In these studies,
Sam68 was shown to act as a regulator of human CSC vulnerability
and increase tumor cell proliferation, migration and invasion.
Thus, Sam68 has been identified as a promising prognostic factor
for leukemia (51,98); neuroblastoma (99,100); epithelial ovarian cancer
(9,101); PC (102); cervical cancer lymph node
metastasis (103); muscle
invasive bladder cancer (104);
endometrial carcinoma (105);
esophageal squamous cell carcinoma (106); non-Hodgkin's lymphoma (107); sacral chordoma (108); and hepatocellular carcinoma
(109).
Sam68 also plays a key role in the development of
renal cell carcinoma (RCC). Evidence suggests that Sam68 is
overexpressed in RCC cell lines and cancer tissue, is positively
associated with disease stage and severity, and is linked to
shorter overall survival rates (110). Sunitinib downregulated
phosphorylated Sam68 expression and, in turn, inhibited RCC cell
apoptosis in murine models (111). Similarly, Sam68 has been found
to be upregulated in human glioblastoma tumor tissue, and its
expression was associated with poor prognosis; therefore, it has
been suggested as a therapeutic target in this disease (112). By contrast, Sam68 deficiency
has been associated with neoplastic transformation of murine NIH3T3
fibroblasts, defective contact inhibition and the development of
metastatic tumors in glioblastoma-bearing nude mice after tyrosine
phosphorylation (113).
As outlined in section 2, Sam68 is regulated
post-translationally via multiple pathways, including tyrosine
phosphorylation by the Src tyrosine kinase (114), or SUMOylation by the SUMO-1
protein (51). This regulation
provides Sam68 with oncogenic properties that regulate cell cycle
and promote tumor cell survival, growth, proliferation and
metastasis. Thus, Sam68 has been suggested as a promising
therapeutic target in different types of tumors.
From a traditional perspective, chemotherapy (CT) is
very effective in increasing the immunogenicity of malignant cells
and inhibiting immunosuppressive pathways in cancer (115). However, tumor cell eradication
by CT was not as effective as initially expected. Later,
combinatorial regimens were developed and successfully tested, with
improved clinical safety and efficacy (115,116). On another note, certain tumors
do not respond properly to CT due to the development of drug
resistance. Resistance may be induced either by pharmacological and
physiological factors, such as drug metabolism or inadequate drug
access to tumor sites, or by cell- and tissue-specific factors,
including gene overexpression and phosphorylation (117,118). In this sense, Sam68
phosphorylation was shown to induce CT resistance in vitro
and in vivo in a variety of tumors, including OTSCC and
prostate, breast and colon cancer. Thus, silencing of Sam68
phosphorylation enhanced the sensitivity of tumor cells to
apoptosis induced by chemotherapeutic agents such as cisplatin,
etoposide or camptothecin (65,75,86,95). This evidence suggests that Sam68
knockdown may improve the cytotoxic effects of CT.
In most types of tumors, there is strong evidence
that elevated levels of the protein Sam68, along with Sam68 gene
overexpression, play a major role in oncogenesis. Thus, Sam68 has
been proven to promote tumor cell development, proliferation,
progression and metastasis in vitro and in vivo.
These findings identify Sam68 as a key regulator in cancer that
should be considered as a clinical biomarker and a promising
therapeutic target. This conclusion is also supported by the fact
that this RBP is directly involved in the self-renewal and survival
capacity of CSCs, which are essential for the activation of tumor
growth and promote treatment failure and tumor relapse (126).
Of note, post-transcriptional modifications, added
to Sam68's characteristics, lead to an increased number of
interactions with multiple genes involved in cancer. Some of these
genes include (but are not limited to) NIMA related kinase 2 or
EPHA3 in BC (69,127); Rad51 in acute myeloid leukemia
(123); lysine-specific
methyltransferase Set7/9 in colon cancer (88); or METTL3 in gastric carcinoma
(88). Thus, the evidence
available suggests that Sam68 may also need the involvement of
other actors with specific roles in these diseases.
The regulation of cancer driven by Sam68 may depend,
at least in part, on i) the type of tissue in which the tumor
develops; ii) the tumor niche; and iii) the interactions between
Sam68 with different RNAs or other proteins via different signaling
pathways, as previously suggested (114). For these reasons, further
research is needed to elucidate the potential role of Sam68 and its
interactions in these settings in vivo and in
vitro.
Based on previous experience of our group with
Sam68, it is strongly suggested that high levels/expression of
Sam68 have an important role not only in cancer (31-33,128), but also in other diseases
(7,129,130). There is cumulative evidence
supporting this idea, which has been particularly evaluated in BC;
CRC (85-88); gastric cancer (91-93); lung cancer (79-84); and PC (10,26,72-76) tumors, among many others (Table II). Different mechanisms of the
role of Sam68 on tumorigenesis in these tumors have been found,
such as regulation of alternative splicing of glycolytic enzymes
(82,87) and proto-oncogenes (72,73,76), the Wnt/β-catenin pathway
(83), genotoxic stress-induced
NF-κB activation (86), DNA
methylation (93) and
modification of miRNAs (91).
However, no meta-analyses or clinical trials have been developed to
elucidate the exact mechanism of Sam68 mediated tumorigenesis.
Meta-analyses may be useful to synthesize the existing data for
diseases in which Sam68 is involved, including cancer. On the other
hand, there is only one clinical trial, carried out by Awe et
al (131), who reported in
2020 that Sam68 is a key determinant of vascular endothelial growth
factor receptor 1 isoform expression, extensively found in cancer
(132). In light of the
cumulative evidence available on the potential role of Sam68 as a
therapeutic target in cancer, further studies are needed to provide
conclusive evidence.
Not applicable.
CJ-C, FS-J, LC-M and VS-M contributed to the
conceptualization, literature search and reviewing of the draft.
CJ-C wrote the draft. All authors have read and agreed to the final
version of the manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
CJ-C is supported by a Margarita Salas fellowship, granted by
the University of Seville (Seville, Spain). The group is supported
by Plan Andaluz Investigación y Desarrollo, Junta de Andalucia,
Spain (grant no. CTS-151 to VS-M).
|
1
|
Courtneidge SA and Fumagalli S: A mitotic
function for Src? Trends Cell Biol. 4:345–347. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Frisone P, Pradella D, Di Matteo A,
Belloni E, Ghigna C and Paronetto MP: SAM68: Signal transduction
and RNA metabolism in human cancer. Biomed Res Int.
2015:5289542015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Najib S, Martín-Romero C, González-Yanes C
and Sánchez-Margalet V: Role of Sam68 as an adaptor protein in
signal transduction. Cell Mol Life Sci. 62:36–43. 2005. View Article : Google Scholar
|
|
4
|
Pagliarini V, Jolly A, Bielli P, Di Rosa
V, De la Grange P and Sette C: Sam68 binds Alu-rich introns in SMN
and promotes pre-mRNA circularization. Nucleic Acids Res.
48:633–645. 2020. View Article : Google Scholar :
|
|
5
|
Messina V, Meikar O, Paronetto MP,
Calabretta S, Geremia R, Kotaja N and Sette C: The RNA binding
protein SAM68 transiently localizes in the chromatoid body of male
germ cells and influences expression of select microRNAs. PLoS One.
7:e397292012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li J, Liu Y, Kim BO and He JJ: Direct
participation of Sam68, the 68-kilodalton Src-associated protein in
mitosis, in the CRM1-mediated Rev nuclear export pathway. J Virol.
76:8374–8382. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vilariño-García T, Pérez-Pérez A,
Santamaría-López E, Prados N, Fernández-Sánchez M and
Sánchez-Margalet V: Sam68 mediates leptin signaling and action in
human granulosa cells: Possible role in leptin resistance in PCOS.
Endocr Connect. 9:479–488. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Adinolfi A, Di Sante G, Rivignani Vaccari
L, Tredicine M, Ria F, Bonvissuto D, Corvino V, Sette C and Geloso
MC: Regionally restricted modulation of Sam68 expression and
Arhgef9 alternative splicing in the hippocampus of a murine model
of multiple sclerosis. Front Mol Neurosci. 15:10736272022.
View Article : Google Scholar
|
|
9
|
Wang Y, Zhang W, Wang X, Wang D, Xie J,
Tang C, Xi Q, Zhong J and Deng Y: Expression of Sam68 correlates
with cell proliferation and survival in epithelial ovarian cancer.
Reprod Sci. 24:97–108. 2017. View Article : Google Scholar
|
|
10
|
Pieraccioli M, Caggiano C, Mignini L,
Zhong C, Babini G, Lattanzio R, Di Stasi S, Tian B, Sette C and
Bielli P: The transcriptional terminator XRN2 and the RNA-binding
protein Sam68 link alternative polyadenylation to cell cycle
progression in prostate cancer. Nat Struct Mol Biol. 29:1101–1112.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Taylor SJ, Resnick RJ and Shalloway D:
Sam68 exerts separable effects on cell cycle progression and
apoptosis. BMC Cell Biol. 5:52004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sanchez-Jimenez F and Sanchez-Margalet V:
Role of Sam68 in post-transcriptional gene regulation. Int J Mol
Sci. 14:23402–23419. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Richard S, Vogel G, Huot ME, Guo T, Muller
WJ and Lukong KE: Sam68 haploinsufficiency delays onset of mammary
tumorigenesis and metastasis. Oncogene. 27:548–556. 2008.
View Article : Google Scholar
|
|
14
|
Irwin ME, Bohin N and Boerner JL: Src
family kinases mediate epidermal growth factor receptor signaling
from lipid rafts in breast cancer cells. Cancer Biol Ther.
12:718–726. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pillay I, Nakano H and Sharma SV:
Radicicol inhibits tyrosine phosphorylation of the mitotic Src
substrate Sam68 and retards subsequent exit from mitosis of
Src-transformed cells. Cell Growth Differ. 7:1487–1499.
1996.PubMed/NCBI
|
|
16
|
Barlat I, Maurier F, Duchesne M, Guitard
E, Tocque B and Schweighoffer F: A role for Sam68 in cell cycle
progression antagonized by a spliced variant within the KH domain.
J Biol Chem. 272:3129–3132. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Paronetto MP, Achsel T, Massiello A,
Chalfant CE and Sette C: The RNA-binding protein Sam68 modulates
the alternative splicing of Bcl-x. J Cell Biol. 176:929–939. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zambuzzi WF, Granjeiro JM, Parikh K,
Yuvaraj S, Peppelenbosch MP and Ferreira CV: Modulation of Src
activity by low molecular weight protein tyrosine phosphatase
during osteoblast differentiation. Cell Physiol Biochem.
22:497–506. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Horn S, Meyer J, Stocking C, Ostertag W
and Jücker M: An increase in the expression and total activity of
endogenous p60(c-Src) in several factor-independent mutants of a
human GM-CSF-dependent leukemia cell line (TF-1). Oncogene.
22:7170–7180. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brignatz C, Paronetto MP, Opi S,
Cappellari M, Audebert S, Feuillet V, Bismuth G, Roche S, Arold ST,
Sette C and Collette Y: Alternative splicing modulates
autoinhibition and SH3 accessibility in the Src kinase Fyn. Mol
Cell Biol. 29:6438–48. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gorla L, Cantù M, Miccichè F, Patelli C,
Mondellini P, Pierotti MA and Bongarzone I: RET oncoproteins induce
tyrosine phosphorylation changes of proteins involved in RNA
metabolism. Cell Signal. 18:2272–2282. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mamidipudi V, Dhillon NK, Parman T, Miller
LD, Lee KC and Cartwright CA: RACK1 inhibits colonic cell growth by
regulating Src activity at cell cycle checkpoints. Oncogene.
26:2914–2924. 2007. View Article : Google Scholar
|
|
23
|
Lukong KE, Larocque D, Tyner AL and
Richard S: Tyrosine phosphorylation of sam68 by breast tumor kinase
regulates intranuclear localization and cell cycle progression. J
Biol Chem. 280:38639–38647. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mizuguchi Y, Specht S, Isse K, Sasatomi E,
Lunz JG III, Takizawa T and Demetris AJ: Breast tumor
kinase/protein tyrosine kinase 6 (Brk/PTK6) activity in normal and
neoplastic biliary epithelia. J Hepatol. 63:399–407. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Brauer PM, Zheng Y, Wang L and Tyner AL:
Cytoplasmic retention of protein tyrosine kinase 6 promotes growth
of prostate tumor cells. Cell Cycle. 9:4190–4199. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Paronetto MP, Farini D, Sammarco I, Maturo
G, Vespasiani G, Geremia R, Rossi P and Sette C: Expression of a
truncated form of the c-Kit tyrosine kinase receptor and activation
of Src kinase in human prostatic cancer. Am J Pathol.
164:1243–1251. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Locatelli A and Lange CA: Met receptors
induce Sam68-dependent cell migration by activation of alternate
extracellular signal-regulated kinase family members. J Biol Chem.
286:21062–21072. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Babic I, Jakymiw A and Fujita DJ: The RNA
binding protein Sam68 is acetylated in tumor cell lines, and its
acetylation correlates with enhanced RNA binding activity.
Oncogene. 23:3781–3789. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sánchez-Jiménez F, Pérez-Pérez A, de la
Cruz-Merino L and Sánchez-Margalet V: Obesity and breast cancer:
Role of leptin. Front Oncol. 9:5962019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sanchez-Margalet V and Najib S: p68 Sam is
a substrate of the insulin receptor and associates with the SH2
domains of p85 PI3K. FEBS Lett. 455:307–310. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pérez-Pérez A, Sánchez-Jiménez F,
Vilariño-García T, de la Cruz L, Virizuela JA and Sánchez-Margalet
V: Sam68 mediates the activation of insulin and leptin signalling
in breast cancer cells. PLoS One. 11:e01582182016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sánchez-Jiménez F, Pérez-Pérez A,
González-Yanes C, Najib S, Varone CL and Sánchez-Margalet V: Leptin
receptor activation increases Sam68 tyrosine phosphorylation and
expression in human trophoblastic cells. Mol Cell Endocrinol.
332:221–227. 2011. View Article : Google Scholar
|
|
33
|
Sánchez-Jiménez F, Pérez-Pérez A,
González-Yanes C, Varone CL and Sánchez-Margalet V: Sam68 mediates
leptin-stimulated growth by modulating leptin receptor signaling in
human trophoblastic JEG-3 cells. Hum Reprod. 26:2306–2315. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Reiss K, Del Valle L, Lassak A and
Trojanek J: Nuclear IRS-1 and cancer. J Cell Physiol.
227:2992–3000. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sung CK, Choi WS and Sanchez-Margalet V:
Guanosine triphosphatase-activating protein-associated protein, but
not src-associated protein p68 in mitosis, is a part of insulin
signaling complexes. Endocrinology. 139:2392–2398. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Matter N, Herrlich P and König H:
Signal-dependent regulation of splicing via phosphorylation of
Sam68. Nature. 420:691–695. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Malki I, Liepina I, Kogelnik N, Watmuff H,
Robinson S, Lightfoot A, Gonchar O, Bottrill A, Fry AM and
Dominguez C: Cdk1-mediated threonine phosphorylation of Sam68
modulates its RNA binding, alternative splicing activity and
cellular functions. Nucleic Acids Res. 50:13045–13062. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang J, Song C and Zhan X: The role of
protein acetylation in carcinogenesis and targeted drug discovery.
Front Endocrinol (Lausanne). 13:9723122022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sobulo OM, Borrow J, Tomek R, Reshmi S,
Harden A, Schlegelberger B, Housman D, Doggett NA, Rowley JD and
Zeleznik-Le NJ: MLL is fused to CBP, a histone acetyltransferase,
in therapy-related acute myeloid leukemia with a t(11;16)
(q23;p13.3). Proc Natl Acad Sci USA. 94:8732–8737. 1997. View Article : Google Scholar
|
|
40
|
Lavau C, Du C, Thirman M and Zeleznik-Le
N: Chromatin-related properties of CBP fused to MLL generate a
myelodysplastic-like syndrome that evolves into myeloid leukemia.
EMBO J. 19:4655–4664. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cheung N, Chan LC, Thompson A, Cleary ML
and So CW: Protein arginine-methyltransferase-dependent
oncogenesis. Nat Cell Biol. 9:1208–1215. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bielli P, Busà R, Paronetto MP and Sette
C: The RNA-binding protein Sam68 is a multifunctional player in
human cancer. Endocr Relat Cancer. 18:R91–R102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Siam A, Baker M, Amit L, Regev G, Rabner
A, Najar RA, Bentata M, Dahan S, Cohen K, Araten S, et al:
Regulation of alternative splicing by p300-mediated acetylation of
splicing factors. RNA. 25:813–824. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bordonaro M: Hypothesis: Sam68 and Pygo2
mediate cell type-specific effects of the modulation of CBP-Wnt and
p300-Wnt activities in colorectal cancer cells. J Cancer.
12:5046–5052. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nakka KK, Chaudhary N, Joshi S, Bhat J,
Singh K, Chatterjee S, Malhotra R, De A, Santra MK, Dilworth FJ and
Chattopadhyay S: Nuclear matrix-associated protein SMAR1 regulates
alternative splicing via HDAC6-mediated deacetylation of Sam68.
Proc Natl Acad Sci USA. 112:E3374–E3383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hwang JW, Cho Y, Bae GU, Kim SN and Kim
YK: Protein arginine methyltransferases: Promising targets for
cancer therapy. Exp Mol Med. 53:788–808. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yu Z, Chen T, Hébert J, Li E and Richard
S: A mouse PRMT1 null allele defines an essential role for arginine
methylation in genome maintenance and cell proliferation. Mol Cell
Biol. 29:2982–2996. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Robin-Lespinasse Y, Sentis S, Kolytcheff
C, Rostan MC, Corbo L and Le Romancer M: hCAF1, a new regulator of
PRMT1-dependent arginine methylation. J Cell Sci. 120:638–647.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Vhuiyan MI, Pak ML, Park MA, Thomas D,
Lakowski TM, Chalfant CE and Frankel A: PRMT2 interacts with
splicing factors and regulates the alternative splicing of BCL-X. J
Biochem. 162:17–25. 2017.PubMed/NCBI
|
|
50
|
Deng L, Meng T, Chen L, Wei W and Wang P:
The role of ubiquitination in tumorigenesis and targeted drug
discovery. Signal Transduct Target Ther. 5:112020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Benoit YD, Mitchell RR, Risueño RM,
Orlando L, Tanasijevic B, Boyd AL, Aslostovar L, Salci KR,
Shapovalova Z, Russell J, et al: Sam68 allows selective targeting
of human cancer stem cells. Cell Chem Biol. 24:833–844.e9. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Babic I, Cherry E and Fujita DJ: SUMO
modification of Sam68 enhances its ability to repress cyclin D1
expression and inhibits its ability to induce apoptosis. Oncogene.
25:4955–4964. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
La Rosa P, Bielli P, Compagnucci C, Cesari
E, Volpe E, Farioli Vecchioli S and Sette C: Sam68 promotes
self-renewal and glycolytic metabolism in mouse neural progenitor
cells by modulating Aldh1a3 pre-mRNA 3'-end processing. Elife.
5:e207502016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Mao P, Joshi K, Li J, Kim SH, Li P,
Santana-Santos L, Luthra S, Chandran UR, Benos PV, Smith L, et al:
Mesenchymal glioma stem cells are maintained by activated
glycolytic metabolism involving aldehyde dehydrogenase 1A3. Proc
Natl Acad Sci USA. 110:8644–8649. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rinkenbaugh AL and Baldwin AS: The NF-κB
pathway and cancer stem cells. Cells. 5:162016. View Article : Google Scholar
|
|
56
|
Vazquez-Santillan K, Melendez-Zajgla J,
Jimenez-Hernandez L, Martínez-Ruiz G and Maldonado V: NF-κB
signaling in cancer stem cells: A promising therapeutic target?
Cell Oncol (Dordr). 38:327–339. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Vazquez-Santillan K, Melendez-Zajgla J,
Jimenez-Hernandez LE, Gaytan-Cervantes J, Muñoz-Galindo L,
Piña-Sanchez P, Martinez-Ruiz G, Torres J, Garcia-Lopez P,
Gonzalez-Torres C, et al: NF-kappaB-inducing kinase regulates stem
cell phenotype in breast cancer. Sci Rep. 6:373402016. View Article : Google Scholar
|
|
58
|
Ishiguro T, Ohata H, Sato A, Yamawaki K,
Enomoto T and Okamoto K: Tumor-derived spheroids: Relevance to
cancer stem cells and clinical applications. Cancer Sci.
108:283–289. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Manuel Iglesias J, Beloqui I,
Garcia-Garcia F, Leis O, Vazquez-Martin A, Eguiara A, Cufi S, Pavon
A, Menendez JA, Dopazo J and Martin AG: Mammosphere formation in
breast carcinoma cell lines depends upon expression of E-cadherin.
PLoS One. 8:e772812013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang L, Tian H, Yuan J, Wu H, Wu J and Zhu
X: CONSORT: Sam68 is directly regulated by MiR-204 and promotes the
Self-renewal potential of breast cancer cells by activating the
Wnt/beta-catenin signaling pathway. Medicine (Baltimore).
94:e22282015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hong BS, Ryu HS, Kim N, Kim J, Lee E, Moon
H, Kim KH, Jin MS, Kwon NH, Kim S, et al: Tumor suppressor
miRNA-204-5p regulates growth, metastasis, and immune
microenvironment remodeling in breast cancer. Cancer Res.
79:1520–1534. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Li P, Wang Q and Wang H: MicroRNA-204
inhibits the proliferation, migration and invasion of human lung
cancer cells by targeting PCNA-1 and inhibits tumor growth in vivo.
Int J Mol Med. 43:1149–1156. 2019.PubMed/NCBI
|
|
63
|
Imam JS, Plyler JR, Bansal H, Prajapati S,
Bansal S, Rebeles J, Chen HI, Chang YF, Panneerdoss S, Zoghi B, et
al: Genomic loss of tumor suppressor miRNA-204 promotes cancer cell
migration and invasion by activating AKT/mTOR/Rac1 signaling and
actin reorganization. PLoS One. 7:e523972012. View Article : Google Scholar
|
|
64
|
Liang CY, Li ZY, Gan TQ, Fang YY, Gan BL,
Chen WJ, Dang YW, Shi K, Feng ZB and Chen G: Downregulation of
hsa-microRNA-204-5p and identification of its potential regulatory
network in non-small cell lung cancer: RT-qPCR, bioinformatic- and
meta-analyses. Respir Res. 21:602020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Turdo A, Gaggianesi M, Di Franco S, Veschi
V, D'Accardo C, Porcelli G, Lo Iacono M, Pillitteri I, Verona F,
Militello G, et al: Effective targeting of breast cancer stem cells
by combined inhibition of Sam68 and Rad51. Oncogene. 41:2196–2209.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Mehner C, Hockla A, Miller E, Ran S,
Radisky DC and Radisky ES: Tumor cell-produced matrix
metalloproteinase 9 (MMP-9) drives malignant progression and
metastasis of basal-like triple negative breast cancer. Oncotarget.
5:2736–2749. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
London M and Gallo E: Critical role of
EphA3 in cancer and current state of EphA3 drug therapeutics. Mol
Biol Rep. 47:5523–5533. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Lv XY, Wang J, Huang F, Wang P, Zhou JG,
Wei B and Li SH: EphA3 contributes to tumor growth and angiogenesis
in human gastric cancer cells. Oncol Rep. 40:2408–2416.
2018.PubMed/NCBI
|
|
69
|
Chen X, Zhang L, Yuan M, Kuang Z, Zou Y,
Tang T, Zhang W, Hu X, Xia T, Cao T and Jia H: Sam68 Promotes the
progression of human breast cancer through inducing activation of
EphA3. Curr Cancer Drug Targets. 20:76–83. 2020. View Article : Google Scholar
|
|
70
|
Aubele M, Walch AK, Ludyga N, Braselmann
H, Atkinson MJ, Luber B, Auer G, Tapio S, Cooke T and Bartlett JM:
Prognostic value of protein tyrosine kinase 6 (PTK6) for long-term
survival of breast cancer patients. Br J Cancer. 99:1089–1095.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Song L, Wang L, Li Y, Xiong H, Wu J, Li J
and Li M: Sam68 up-regulation correlates with, and its
down-regulation inhibits, proliferation and tumourigenicity of
breast cancer cells. J Pathol. 222:227–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Paronetto MP, Cappellari M, Busà R,
Pedrotti S, Vitali R, Comstock C, Hyslop T, Knudsen KE and Sette C:
Alternative splicing of the cyclin D1 proto-oncogene is regulated
by the RNA-binding protein Sam68. Cancer Res. 70:229–239. 2010.
View Article : Google Scholar :
|
|
73
|
Caggiano C, Pieraccioli M, Panzeri V,
Sette C and Bielli P: c-MYC empowers transcription and productive
splicing of the oncogenic splicing factor Sam68 in cancer. Nucleic
Acids Res. 47:6160–6171. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Derry JJ, Prins GS, Ray V and Tyner AL:
Altered localization and activity of the intracellular tyrosine
kinase BRK/Sik in prostate tumor cells. Oncogene. 22:4212–4220.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Busà R, Paronetto MP, Farini D,
Pierantozzi E, Botti F, Angelini DF, Attisani F, Vespasiani G and
Sette C: The RNA-binding protein Sam68 contributes to proliferation
and survival of human prostate cancer cells. Oncogene.
26:4372–4382. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Rajan P, Gaughan L, Dalgliesh C, El-Sherif
A, Robson CN, Leung HY and Elliott DJ: The RNA-binding and adaptor
protein Sam68 modulates signal-dependent splicing and
transcriptional activity of the androgen receptor. J Pathol.
215:67–77. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Uddin MH, Li Y, Khan HY, Muqbil I,
Aboukameel A, Sexton RE, Reddy S, Landesman Y, Kashyap T, Azmi AS
and Heath EI: Nuclear export inhibitor KPT-8602 synergizes with
PARP inhibitors in escalating apoptosis in castration resistant
cancer cells. Int J Mol Sci. 22:66762021. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Mijatovic T, De Nève N, Gailly P, Mathieu
V, Haibe-Kains B, Bontempi G, Lapeira J, Decaestecker C, Facchini V
and Kiss R: Nucleolus and c-Myc: Potential targets of
cardenolide-mediated antitumor activity. Mol Cancer Ther.
7:1285–1296. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Sumithra B, Jayanthi VSPKSA, Manne HC,
Gunda R, Saxena U and Das AB: Antibody-based biosensor to detect
oncogenic splicing factor Sam68 for the diagnosis of lung cancer.
Biotechnol Lett. 42:2501–2509. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zhang Z, Xu Y, Sun N, Zhang M, Xie J and
Jiang Z: High Sam68 expression predicts poor prognosis in Non-small
cell lung cancer. Clin Transl Oncol. 16:886–891. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Lin CH, Liao CC, Wang SY, Peng CY, Yeh YC,
Chen MY and Chou TY: Comparative O-GlcNAc proteomic analysis
reveals a role of O-GlcNAcylated SAM68 in lung cancer
aggressiveness. Cancers (Basel). 14:2432022. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhu S, Chen W, Wang J, Qi L, Pan H, Feng Z
and Tian D: SAM68 promotes tumorigenesis in lung adenocarcinoma by
regulating metabolic conversion via PKM alternative splicing.
Theranostics. 11:3359–3375. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Li X, Zhou X, Hua F, Fan Y, Zu L, Wang Y,
Shen W, Pan H and Zhou Q: The RNA-binding protein Sam68 is critical
for non-small cell lung cancer cell proliferation by regulating
Wnt/β-catenin pathway. Int J Clin Exp Pathol. 10:8281–8291.
2017.
|
|
84
|
Sumithra B, Saxena U and Das AB: A
comprehensive study on genome-wide coexpression network of
KHDRBS1/Sam68 reveals its cancer and Patient-specific association.
Sci Rep. 9:110832019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Masibag AN, Bergin CJ, Haebe JR, Zouggar
A, Shah MS, Sandouka T, Mendes da Silva A, Desrochers FM,
Fournier-Morin A and Benoit YD: Pharmacological targeting of Sam68
functions in colorectal cancer stem cells. iScience. 24:1034422021.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Fu K, Sun X, Wier EM, Hodgson A, Liu Y,
Sears CL and Wan F: Sam68/KHDRBS1 is critical for colon
tumorigenesis by regulating genotoxic stress-induced NF-κB
activation. Elife. 5:e150182016. View Article : Google Scholar
|
|
87
|
Zhao J, Li J, Hassan W, Xu D, Wang X and
Huang Z: Huang, Sam68 promotes aerobic glycolysis in colorectal
cancer by regulating PKM2 alternative splicing. Ann Transl Med.
8:4592020. View Article : Google Scholar
|
|
88
|
Vasileva E, Shuvalov O, Petukhov A,
Fedorova O, Daks A, Nader R and Barlev N: KMT Set7/9 is a new
regulator of Sam68 STAR-protein. Biochem Biophys Res Commun.
525:1018–1024. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Li N and Richard S: Sam68 functions as a
transcriptional coactivator of the p53 tumor suppressor. Nucleic
Acids Res. 44:8726–8741. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Li N, Ngo CT, Aleynikova O, Beauchemin N
and Richard S: The p53 status can influence the role of Sam68 in
tumorigenesis. Oncotarget. 7:71651–71659. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yu X, Kang W, Zhang J, Chen C and Liu Y:
Shortening of the KHDRBS1 3'UTR by alternative cleavage and
polyadenylation alters miRNA-mediated regulation and promotes
gastric cancer progression. Am J Transl Res. 14:6574–6585.
2022.PubMed/NCBI
|
|
92
|
Xiao J, Wang Q, Yang Q, Wang H, Qiang F,
He S, Cai J, Yang L and Wang Y: Clinical significance and effect of
Sam68 expression in gastric cancer. Oncol Lett. 15:4745–4752.
2018.PubMed/NCBI
|
|
93
|
Zhang JY, Du Y, Gong LP, Shao YT, Pan LJ,
Feng ZY, Pan YH, Huang JT, Wen JY, Sun LP, et al: ebv-circRPMS1
promotes the progression of EBV-associated gastric carcinoma via
Sam68-dependent activation of METTL3. Cancer Lett. 535:2156462022.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Komiyama T, Kuroshima T, Sugasawa T,
Fujita SI, Ikami Y, Hirai H, Tsushima F, Michi Y, Kayamori K,
Higashino F and Harada H: High expression of Sam68 contributes to
metastasis by regulating vimentin expression and a motile phenotype
in oral squamous cell carcinoma. Oncol Rep. 48:1832022. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Chen S, Li H, Zhuang S, Zhang J, Gao F,
Wang X, Chen W and Song M: Sam68 reduces Cisplatin-induced
apoptosis in tongue carcinoma. J Exp Clin Cancer Res. 35:1232016.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Fu K, Sun X, Xia X, Hobbs RP, Guo Y,
Coulombe PA and Wan F: Sam68 is required for the growth and
survival of nonmelanoma skin cancer. Cancer Med. 8:6106–6113. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Paronetto MP, Messina V, Bianchi E, Barchi
M, Vogel G, Moretti C, Palombi F, Stefanini M, Geremia R, Richard S
and Sette C: Sam68 regulates translation of target mRNAs in male
germ cells, necessary for mouse spermatogenesis. J Cell Biol.
185:235–249. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Wang Q, Li Y, Cheng J, Chen L, Xu H, Li Q
and Pang T: Sam68 affects cell proliferation and apoptosis of human
adult T-acute lymphoblastic leukemia cells via AKT/mTOR signal
pathway. Leuk Res. 46:1–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zhao D, Tian Y, Li P, Wang L, Xiao A,
Zhang M and Shi T: MicroRNA-203 inhibits the malignant progression
of neuroblastoma by targeting Sam68. Mol Med Rep. 12:5554–5560.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Zhao X, Li Z, He B, Liu J, Li S, Zhou L,
Pan C, Yu Z and Xu Z: Sam68 is a novel marker for aggressive
neuroblastoma. Onco Targets Ther. 6:1751–1760. 2013.PubMed/NCBI
|
|
101
|
Dong L, Che H, Li M and Li X: Sam68 is
overexpressed in epithelial ovarian cancer and promotes tumor cell
proliferation. Med Sci Monit. 22:3248–3256. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Chen ZY, Cai L, Zhu J, Chen M, Chen J, Li
ZH, Liu XD, Wang SG, Bie P, Jiang P, et al: Fyn requires HnRNPA2B1
and Sam68 to synergistically regulate apoptosis in pancreatic
cancer. Carcinogenesis. 32:1419–1426. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Li Z, Yu CP, Zhong Y, Liu TJ, Huang QD,
Zhao XH, Huang H, Tu H, Jiang S, Zhang Y, et al: Sam68 expression
and cytoplasmic localization is correlated with lymph node
metastasis as well as prognosis in patients with Early-stage
cervical cancer. Ann Oncol. 23:638–646. 2012. View Article : Google Scholar
|
|
104
|
Zhang Z, Yu C, Li Y, Jiang L and Zhou F:
Utility of SAM68 in the progression and prognosis for bladder
cancer. BMC Cancer. 15:3642015. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Wang Q, Li Y, Zhou J, Liu J, Qin J, Xing
F, Zhang J and Cheng J: Clinical significance of Sam68 expression
in endometrial carcinoma. Tumour Biol. 36:4509–4518. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wang Y, Liang L, Zhang J, Li M, Zhu J,
Gong C, Yang L, Zhu J, Chen L and Ni R: Sam68 promotes cellular
proliferation and predicts poor prognosis in esophageal squamous
cell carcinoma. Tumour Biol. 36:8735–8745. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Wu Y, Xu X, Miao X, Zhu X, Yin H, He Y, Li
C, Liu Y, Chen Y, Lu X, et al: Sam68 regulates cell proliferation
and cell adhesion-mediated drug resistance (CAM-DR) via the AKT
pathway in non-Hodgkin's lymphoma. Cell Prolif. 48:682–690. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Wen H, Li P, Ma H, Zheng J, Yu Y and Lv G:
High expression of Sam68 in sacral chordomas is associated with
worse clinical outcomes. Onco Targets Ther. 10:4691–4700. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zhang T, Wan C, Shi W, Xu J, Fan H, Zhang
S, Lin Z, Ni R and Zhang X: The RNA-binding protein Sam68 regulates
tumor cell viability and hepatic carcinogenesis by inhibiting the
transcriptional activity of FOXOs. J Mol Histol. 46:485–497. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Zhang Z, Li J, Zheng H, Yu C, Chen J, Liu
Z, Li M, Zeng M, Zhou F and Song L: Expression and cytoplasmic
localization of SAM68 is a significant and independent prognostic
marker for renal cell carcinoma. Cancer Epidemiol Biomarkers Prev.
18:2685–2693. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Wu Z, Peng Y, Xiong L, Wang J, Li Z, Ning
K, Deng M, Wang N, Wei W, Li Z, et al: Role of Sam68 in Sunitinib
induced renal cell carcinoma apoptosis. Cancer Med. 11:3674–3686.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Feng J, Ren X, Fu H, Li D, Chen X, Zu X,
Liu Q and Wu M: LRRC4 mediates the formation of circular RNA CD44
to inhibitGBM cell proliferation. Mol Ther Nucleic Acids.
26:473–487. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Liu K, Li L, Nisson PE, Gruber C, Jessee J
and Cohen SN: Neoplastic transformation and tumorigenesis
associated with sam68 protein deficiency in cultured murine
fibroblasts. J Biol Chem. 275:40195–40201. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Lukong KE and Richard S: Targeting the
RNA-binding protein Sam68 as a treatment for cancer? Future Oncol.
3:539–544. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Galluzzi L, Buqué A, Kepp O, Zitvogel L
and Kroemer G: Immunological effects of conventional chemotherapy
and targeted anticancer agents. Cancer Cell. 28:690–714. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Behranvand N, Nasri F, Zolfaghari Emameh
R, Khani P, Hosseini A, Garssen J and Falak R: Chemotherapy: A
double-edged sword in cancer treatment. Cancer Immunol Immunother.
71:507–526. 2022. View Article : Google Scholar
|
|
117
|
Raguz S and Yagüe E: Resistance to
chemotherapy: New treatments and novel insights into an old
problem. Br J Cancer. 99:387–391. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Brown FC, Still E, Koche RP, Yim CY, Takao
S, Cifani P, Reed C, Gunasekera S, Ficarro SB, Romanienko P, et al:
MEF2C phosphorylation is required for chemotherapy resistance in
acute myeloid leukemia. Cancer Discov. 8:478–497. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Saqub H, Proetsch-Gugerbauer H, Bezrookove
V, Nosrati M, Vaquero EM, de Semir D, Ice RJ, McAllister S,
Soroceanu L, Kashani-Sabet M, et al: Dinaciclib, a cyclin-dependent
kinase inhibitor, suppresses cholangiocarcinoma growth by targeting
CDK2/5/9. Sci Rep. 10:184892020. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Savage G and Antman KH: Imatinib
mesylate-a new oral targeted therapy. N Engl J Med. 346:683–693.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Zhong L, Li Y, Xiong L, Wang W, Wu M, Yuan
T, Yang W, Tian C, Miao Z, Wang T and Yang S: Small molecules in
targeted cancer therapy: Advances, challenges, and future
perspectives. Signal Transduct Target Ther. 6:2012021. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Wang Z, Jia R, Wang L, Yang Q, Hu X, Fu Q,
Zhang X, Li W and Ren Y: The emerging roles of Rad51 in cancer and
its potential as a therapeutic target. Front Oncol. 12:9355932022.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Huang F and Mazin AV: A small molecule
inhibitor of human RAD51 potentiates breast cancer cell killing by
therapeutic agents in mouse xenografts. PLoS One. 9:e1009932014.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Zhang L, Xu L, Zhang F and Vlashi E:
Doxycycline inhibits the cancer stem cell phenotype and
Epithelial-to-mesenchymal transition in breast cancer. Cell Cycle.
16:737–745. 2017. View Article : Google Scholar :
|
|
125
|
Hirai H, Sootome H, Nakatsuru Y, Miyama K,
Taguchi S, Tsujioka K, Ueno Y, Hatch H, Majumder PK, Pan BS and
Kotani H: MK-2206, an allosteric Akt inhibitor, enhances antitumor
efficacy by standard chemotherapeutic agents or molecular targeted
drugs in vitro and in vivo. Mol Cancer Ther. 9:1956–1967. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Marzagalli M, Fontana F, Raimondi M and
Limonta P: Cancer stem Cells-key players in tumor relapse. Cancers
(Basel). 13:3762021. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Naro C, Barbagallo F, Caggiano C, De Musso
M, Panzeri V, Di Agostino S, Paronetto MP and Sette C: Functional
interaction between the oncogenic kinase NEK2 and Sam68 promotes a
splicing program involved in migration and invasion in
Triple-negative breast cancer. Front Oncol. 12:8806542022.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Quintana-Portillo R, Canfrán-Duque A,
Issad T, Sánchez-Margalet V and González-Yanes C: Sam68 interacts
with IRS1. Biochem Pharmacol. 83:78–87. 2012. View Article : Google Scholar
|
|
129
|
Vilariño-García T, Guadix P, Dorado-Silva
M, Sánchez-Martín P, Pérez-Pérez A and Sánchez-Margalet V:
Decreased expression of Sam68 is associated with insulin resistance
in granulosa cells from PCOS patients. Cells. 11:28212022.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Najib S, Rodríguez-Baño J, Ríos MJ,
Muniain MA, Goberna R and Sánchez-Margalet V: Sam68 is tyrosine
phosphorylated and recruited to signalling in peripheral blood
mononuclear cells from HIV infected patients. Clin Exp Immunol.
141:518–525. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Awe O, Sinkway JM, Chow RP, Wagener Q,
Schulz EV, Yu JY, Nietert PJ, Wagner CL and Lee KH: Differential
regulation of a placental SAM68 and sFLT1 gene pathway and the
relevance to maternal vitamin D sufficiency. Pregnancy Hypertens.
22:196–203. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Shibuya M: Involvement of Flt-1 (VEGF
receptor-1) in cancer and preeclampsia. Proc Jpn Acad Ser B Phys
Biol Sci. 87:167–178. 2011. View Article : Google Scholar : PubMed/NCBI
|