Introduction
Bone marrow mesenchymal stem cells (BMSCs) possess
robust self-renewal and multi-lineage differentiation potential,
rendering them key cellular resources in bone tissue regeneration
engineering (1). However,
stress-induced senescence leads to SC exhaustion and a marked
decline in osteogenic differentiation, thereby constraining the
application of BMSCs (2,3). Therefore, elucidating the specific
mechanism underlying BMSC senescence and its impact on SC function,
as well as exploring potential intervention strategies, is key.
Cell senescence is a state of cell cycle arrest
caused by endogenous and exogenous stimuli, accompanied by
epigenetic changes (4). The
observed alterations include disruption of heterochromatin
structure (5), aberrations in
DNA methylation and histone modification patterns (6), as well as perturbations in nuclear
membrane integrity (7). ~50% of
eukaryotic genome sequences are repeats and organized into densely
packed and transcriptionally suppressed heterochromatin (8,9).
Under certain pathophysiological conditions, such as cancer,
placental development and aging, heterochromatin is reorganized by
epigenetic modification and accompanied by aberrant transcriptional
activation of repetitive elements (REs) (10-12). REs are classified into tandem and
interspersed repeats based on structural organization. Tandem
repeats consist of satellite and simple repeats, while interspersed
REs primarily refer to transposable elements that actively or
passively integrate into the genome. Transposable elements are
divided into RNA (class I) and DNA transposons (class II elements)
according to the transposition mechanisms. DNA transposons mobilize
their DNA sequence through a 'cut and paste' process, wherein
transposons are excised from the donor site and inserted into a
different genomic location (13). By contrast, retrotransposons
mobilize via a 'copy and paste' manner, which involves reverse
transcription of an RNA intermediate followed by insertion into a
new locus (14).
Retrotransposons are categorized into two subtypes: Long terminal
repeats (LTRs) and non-LTRs. LTRs are commonly known as endogenous
retroviruses (ERVs), whereas non-LTRs include long interspersed
nuclear elements (LINEs) and short interspersed nuclear elements
(15,16). Due to their presence in the host
genome but transcriptional silencing, REs have been regarded as
'junk DNA'. However, advancements in sequencing and computer
analysis technologies have led to a re-evaluation of REs,
particularly transposable elements, enabling more comprehensive
exploration of their biological functions (17). RE transcription leads to
accumulation of endogenous cytosolic self-nucleic acids,
potentially leading to the activation of intracellular signaling
pathways (18-20). Epigenetic resetting and RE
activation during aging and senescence have been reported (11), however, the internal mechanisms
by which they affect the senescence and function of BMSCs remain
poorly understood.
Quercetin, a natural bioactive flavonoid extracted
from a wide variety of fruits, vegetables is known for its
anti-inflammatory, antioxidant and anti-cancer properties (21,22). Quercetin is a well-established
senolytic drug that effectively targets multiple mechanisms to
eliminate senescent cells and suppress the senescence-associated
secretory phenotype (SASP) (23). The activities of this compound
are associated with activation of various pathways, including the
PI3K/Akt and AMPK pathways, and sirtuin 1/PINK1/Parkin-mediated
mitophagy (21). However, the
extent to which quercetin can mitigate or reverse
senescence-related decline of BMSC function and its potential role
in regulating epigenetic mechanisms and inhibiting activation of
REs remain largely unexplored.
The present study investigated the effect of
quercetin against oxidative stress-induced senescence of BMSCs and
its role in reversing senescence-associated decline in osteogenic
capacity. The current study aimed to provide new ideas for the
intervention of SC senescence and development of bone tissue
regeneration.
Materials and methods
Isolation and culture of BMSCs
Animal experiments were conducted according to the
approved guidelines for experimental animal ethics and welfare by
the Institutional Animal Care and Use Committee of Sun Yat-Sen
University (Guangzhou, China) (approval no.
SYSU-IACUC-2024-002262). For BMSC isolation, 10 3-week-old male
Sprague-Dawley rats weighing about 50 g were purchased from the
Laboratory Animal Center of Sun Yat-Sen University. Rats were
sacrificed by cervical dislocation under anesthetization with
isoflurane inhalation (induction, 3%; maintenance, 2%). Animal
death was confirmed by respiratory arrest, cessation of heartbeat
and disappearance of reflexes. Rats were immersed in 75% ethanol at
room temperature for 20 min to sterilize. Tibiae and femurs were
dislocated and both ends were cut off under aseptic conditions. The
bone marrow was flushed out using a syringe, cut into small
fragments and subsequently collected by centrifugation (250 × g, 5
min, 4°C). The fragments were resuspended in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) and seeded into a culture flask. Primary
BMSCs were cultured in DMEM containing 10% fetal bovine serum
(Moregate BioTech) and 100 μg/ml Primocin®
(InvivoGen) in a humidified incubator containing 5% CO2
at 37°C. The culture medium was changed every 3 days. Passage three
BMSCs were used for subsequent experiments.
Chemical compounds
Quercetin was purchased from MedChemExpress and
dissolved in DMSO (Sigma-Aldrich; Merck KGaA).
Poly(I:C)-LMW/LyoVec™ [Poly(I:C)], was purchased from InvivoGen and
dissolved following the manufacturer's instructions. Briefly, to
prepare a stock solution (50 μg/ml), 25 μg
lyophilized Poly(I:C) was added to 500 μl endotoxin-free
water and gently mixed for ≥15 min. The products were aliquoted and
stored at −20°C.
Osteogenic differentiation
For osteogenic induction, BMSCs (1.5×105
cells/well) were seeded in 12-well plates. After reaching 70-80%
confluence, cells were stimulated with 200 μM
H2O2 for 1 h or treated with 0.1 μg/ml
Poly(I:C) for 24 h at 37°C with 5% CO2. Cells were
washed and cultured with osteogenic induction medium (Cyagen
Biosciences Inc.) at 37°C with 5% CO2. Quercetin at
concentration of 1 μM was added to osteogenic induction
medium. The culture medium was refreshed every 3 days. Alkaline
phosphatase (ALP) staining was conducted after 7 days using a
BCIP/NBT ALP Color Development kit (Beyotime Institute of
Biotechnology) according to the manufacturer's instructions.
Alizarin red staining was performed 14-21 days post-induction.
Briefly, BMSCs were washed with PBS and fixed with 4%
paraformaldehyde at room temperature for 30 min. Cells were stained
with alizarin red for 10 min at room temperature. After that, the
images were captured by an inverted light microscope (Zeiss Axio;
magnification, ×100). The stained area was calculated by Image J
1.48v software (National Institutes of Health).
Treatment and transfection
To assess the geroprotective effect of quercetin
treatment and RIG-I knockdown, 2×104 BMSCs were seeded
in 24-well plates and cultured at 37°C with 5% CO2 in
complete DMEM for 24 h. Next, cells were treated with 200 μM
H2O2 for 1 h to induce senescence. For
quercetin treatment, cells were washed with phosphate-buffered
saline (PBS) and cultured with fresh complete DMEM containing
quercetin (0.0, 0.1, 1.0, 10.0 and 100.0 μM) at 37°C with 5%
CO2 for 3 days. For RIG-I knockdown, cells were washed
and subjected to short-interfering RNA (siRNA) transfection using
Lipofectamine® RNAiMAX Reagent (Invitrogen; Thermo
Fisher Scientific, Inc.; cat. no. 13778-150) according to the
manufacturer's instructions. Briefly, cells were transfected with
50 nM siRNA at 37°C with 5% CO2 for 6 h. The si-RIG-I
(5′-CAUUGAAACCAAGAAAUUACC-3′) were synthesized by GENEray Biotech
(Shanghai) Co., Ltd. The si-negative control (NC,
5′-UUCUCCGAACGUGUCACGUTT-3′) was set as the control. To evaluate
knockdown efficiency, cells were harvested for qPCR 48-72 h after
transfection.
Senescence-associated β-galactosidase
(SA-β-gal) staining
Following treatment with quercetin at 37°C with 5%
CO2 for 3 days, senescent cells were assessed using a
SA-β-gal staining kit (Beyotime Institute of Biotechnology)
according to manufacturer's instructions. Briefly, cultured cells
were fixed at room temperature for 15 min, washed with PBS and
incubated with staining solution at 37°C overnight. Images were
captured using an inverted light microscope (Zeiss Axio;
magnification, ×100) and the ratio of stained cells was quantified
using ImageJ 1.48v software (National Institutes of Health).
Cell Counting Kit-8 (CCK-8) assay
The optimal concentration of quercetin was selected
by assessing its toxic effect using CCK-8 assay. Briefly, BMSCs
(3×103 cells/well) were seeded in 96-well plates and
treated with quercetin (0.0, 0.1, 1.0, 10.0, 100.0 and 1,000.0
μM) at 37°C with 5% CO2. After 3 days, cell
viability was measured using CCK-8 (Dojindo Laboratories, Inc.)
according to the manufacturer's protocol. Briefly, cells were
incubated with CCK-8 solution for 2 h at 37°C. Quercetin
concentrations that significantly decreased cell viability compared
with the blank control were considered cytotoxic.
Colony formation assay
The colony formation assay was used to evaluate the
self-renewal ability of BMSCs. In brief, 5×103 BMSCs
were seeded in 12-well plates and stimulated with 200 μM
H2O2 to induce senescence as aforementioned.
Then, BMSCs were cultured in complete DMEM (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C with addition of quercetin (0.0, 0.1,
1.0, 10.0 and 100.0 μM) for 7-10 days. The number of
colonies formed was calculated by crystal violet staining (Beijing
Solarbio Science & Technology Co., Ltd.). Briefly, cells were
fixed with 4% paraformaldehyde for 15 min, then stained with 0.1%
crystal violet for 15 min, both at room temperature. The images
were captured and the number of colonies (>50 cells) were
manually calculated.
Reverse transcription-quantitative PCR
(RT-qPCR)
After culturing in DMEM or osteogenic induction
medium (Cyagen Biosciences Inc.), total RNA was extracted from
cells using the RNA-Quick Purification kit (cat. no. RN001; Yishan
Biotechnology Co., Ltd.). RNA was reverse-transcribed into cDNA
using HiScript II Q RT SuperMix according to the manufacturer's
protocol for qPCR (Vazyme Biotech Co., Ltd.). qPCR was performed
using Taq Pro Universal SYBR qPCR Master Mix (Vazyme Biotech Co.,
Ltd.) on a Light Cycler 480 system (Roche Applied Science). The
thermocycling conditions were as follows: Initial denaturation at
95°C for 30 sec, followed by 40 cycles of denaturation at 95°C for
10 sec, annealing and extension at 60°C for 30 sec; and a melting
curve between 60 and 95°C. All results were normalized to GAPDH and
the quantitative method of relative mRNA expression was used
2−ΔΔCq (24). The
senescence marker gene (p21), SASP genes (IL6, TNF-α, IL1α and
IL1β), and osteogenic marker genes [osteoprotegerin (OPG),
osteocalcin (OCN), osteopontin (OPN) and type I collagen α 1
(COL1A1)] were detected. The PCR primers are listed in Table I.
 | Table ISequences for PCR. |
Table I
Sequences for PCR.
| Gene | Sequence,
5′➔3′ |
|---|
| p21 |
F-TGTGATATGTACCAGCCACAGG |
|
R-GCGAAGTCAAAGTTCCACCG |
| IL6 |
F-CTGCTCTGGTCTTCTGGAGT |
|
R-GGTCTTGGTCCTTAGCCACT |
| TNF-α | F:
GGCGTGTTCATCCGTTCTCT |
| R:
CCCAGAGCCACAATTCCCTT |
| IL1α | F:
AGGGCACAGAGGGAGTCAA |
| R:
AGAGACAGATGGTCAATGGCA |
| IL1β | F:
GGGATGATGACGACCTGCTA |
| R:
TGTCGTTGCTTGTCTCTCCT |
| OPG | F:
TGTCCCTTGCCCTGACTACT |
| R:
GTAGCGCCCTTCCTCACATT |
| OCN | F:
CCGTTTAGGGCATGTGTTGC |
| R:
CCGTCCATACTTTCGAGGCA |
| OPN | F:
AAGCGTGGAAACACACAGC |
| R:
CTTTGGAACTCGCCTGACTG |
| COL1A1 | F:
GAGACAGGCGAACAAGGTGA |
| R:
GGGAGACCGTTGAGTCCATC |
| RIG-I | F:
AGCCAATGCGTTCTTACCCA |
| R:
CATCGCCGAGTGCTAAGAGT |
| IFN-β | F:
AGCACTGGGTGGAATGAGAC |
| R:
GACCACCATCCAGGCATAGC |
| GAPDH | F:
TATGACTCTACCCACGGCAAG |
| R:
TACTCAGCACCAGCATCACC |
Cellular immunofluorescence
BMSCs (2×104 cells/well) were seeded on
slides in 24-well plates. Following senescence induction and
quercetin treatment, cellular immunofluorescence was used to
evaluate expression of γ-H2AX, methylation histone H3 Lys9
(H3K9me3), heterochromatin protein 1α (Hp1α), lamina-associated
polypeptide 2 (LAP2), double-stranded (ds)RNA clone rJ2 (rJ2),
LINE-1 open reading frame 1 protein (ORF1p) and dsDNA. BMSCs
cultured on microscope coverslips at 37°C for 3 days were fixed
with 4% paraformaldehyde for 15 min, permeabilized with 0.4% Triton
X-100 in PBS for 30 min and blocked with 10% goat serum (BOSTER
Biological Technology Co., Ltd.) for 1 h, all at room temperature.
Slides were covered with primary antibodies at 4°C overnight
followed by the Alexa Fluor 488 dye-(1:300; cat. no. EM35141-01;
Beijing Emarbio Science & Technology Co., Ltd.) or Alexa Fluor
594 dye-conjugated secondary antibodies (1:300; cat. no.
EM35150-01; Beijing Emarbio Science & Technology Co., Ltd.) for
1 h at room temperature in the dark. Nuclei were stained with DAPI
for 10 min at room temperature and slides were mounted using
anti-fading mounting medium (Vector Laboratories, Inc.; cat. no.
H-1000). Images were acquired using a laser scanning confocal
microscope (Carl Zeiss AG; magnification, ×400). The mean
immunofluorescence intensity or proportion of positive cells was
quantified using ImageJ 1.48v software.
The antibodies used for immunofluorescence staining
were as follows: γ-H2AX (1:400; cat. no. 9718; Cell Signaling
Technology, Inc.), H3K9me3 (1:400; cat. no. ab8898; Abcam), Hp1α
(1:200; cat. no. 2616; Cell Signaling Technology, Inc.), LAP2
(1:100; cat. no. 611000; BD Biosciences), rJ2 (1:100; cat. no.
MABE1134; MilliporeSigma), ORF1p (1:200; cat. no. MABC1152;
MilliporeSigma) and dsDNA (1:400; cat. no. sc-58749; Santa Cruz
Biotechnology, Inc.).
Western blotting
BMSCs (3×105 cells seeded in 6-well
plates) were lysed in RIPA buffer (Beyotime Institute of
Biotechnology) supplemented with protease and phosphatase
inhibitors. Total protein was quantified using BCA Protein Assay
kit (Bio-Rad Laboratories, Inc.). Then, 30 μg/lane protein
were loaded onto SDS-PAGE in 5-15% Bis-Tris precast gel to separate
the different proteins. Subsequently, samples were transferred to a
PVDF membrane (MilliporeSigma) and blocked with 5% non-fat milk for
1 h at room temperature. Then, PVDF membranes were incubated with
primary antibodies [H3K9me3, Hp1α, LAP2, RIG-I, phosphorylated
TANK-binding kinase 1 (p-TBK1), TBK1 or GAPDH] at 4°C overnight
followed by secondary antibodies (1:10,000; HRP AffiniPure Goat
Anti-Rabbit, cat. no. EM35111-01 or HRP AffiniPure Goat Anti-Mouse,
cat. No. EM35110-01; Beijing Emarbio Science & Technology Co.,
LTD.) for 1 h at room temperature. The signals were detected using
the ECL Immobilon Western Chemilum HRP Substrate (cat. no.
WBKLS0500; Merck Millipore) and an ultra-high sensitivity
chemiluminescence imaging system (Bio-Rad Laboratories, Inc.) and
then quantified using ImageJ 1.48v software (National Institutes of
Health).
The antibodies were as follows: H3K9me3 (1:1,000;
cat. no ab8898; Abcam), Hp1α (1:1,000; cat. no. 2616; Cell
Signaling Technology, Inc.), LAP2 (1:500; cat. no. 611000; BD
Biosciences), RIG-I (1:1,000; cat. no. A13407; ABclonal Biotech
Co., Ltd.), p-TBK1 (1:250; cat. no. AP1418; ABclonal Biotech Co.,
Ltd.), TBK1 (1:1,000; cat. no. A3458; ABclonal Biotech Co., Ltd.)
and GAPDH (1:3,000; cat. no. 60004-1-Ig; Proteintech Group,
Inc.).
RNA sequencing (RNA-seq) and data
processing
Following H2O2 stimulation and
quercetin treatment, RNA-seq analysis of BMSCs was conducted by
Shanghai Majorbio Bio-pharm Technology Co., Ltd. Total RNA was
obtained using MJZol total RNA extraction kit (cat. no. T01-500;
Majorbio). RNA quality was determined by 5300 Bioanalyzer (Agilent)
and quantified using ND-2000 (NanoDrop Technologies). The loading
concentration of the final library was 10 pM. Agarose gel (Biowest
Agarose; Biowest) electrophoresis was used to detect the RNA
integrity. mRNA was enriched with Oligo (dT) beads, fragmented into
short fragments (300 bp), and reverse-transcribed into cDNA. After
being linked to adaptor and purified, cDNA fragments were sequenced
on an Illumina NovaSeq Xplus platform (Illumina, Inc.). cDNA
library was prepared following Illumina® Stranded mRNA
Prep, Ligation (cat. no. 20040534; Illumina, Inc.). The sequencing
library was performed on NovaSeq X plus platform (PE150) using
NovaSeq X Series 10B Reagent Kit (300cycles; cat. no. 20085594;
Illumina Inc.) and 300 bp paired-end reads were generated. The
cleaned reads were mapped to Rattus norvegicus genome
(mRatBN7.2; ncbi.nlm.nih.gov/datasets/genome/GCF_015227675.2)
using hisat2 (version 2.1.0). For RE analysis, transposable
elements were quantified and analyzed using TE transcripts (version
2.2.3). Differentially expressed REs were counted using R package
DESeq2 (version 1.38.2) with a cut-off of |log2(fold
change)| >0.3 and P<0.05. The genes significantly affected by
H2O2 stimulation or quercetin were determined
by setting a fold change of ≥1.5 and P<0.05. Venn diagram
analysis of differentially expressed genes, Kyoto Encyclopedia of
Genes and Genomes (KEGG) pathway and Gene Set Enrichment Analysis
(GSEA) were performed on Majorbio Cloud (majorbio.com).
Statistical analysis
The data are presented as the mean ± standard
deviation of three independent experimental repeats. Results were
statistically analyzed using GraphPad Prism (version 5.0;
Dotmatics). Comparisons were conducted with unpaired Student's
t-test or one-way ANOVA followed by Dunnett's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Quercetin alleviates oxidative
stress-induced senescence in BMSCs
The accumulation of reactive oxygen species in BMSCs
causes oxidative stress-induced senescence and impairs SC
properties (25). To induce
cellular senescence in the present study, exogenous
H2O2 was used to stimulate BMSCs. Following
treatment with 200 μM H2O2, the
proportion of SA-β-gal-positive cells significantly increased
(Fig. 1A and B), indicating
cellular senescence was successfully induced under this condition.
To investigate the effect of quercetin on senescent BMSCs,
quercetin (0, 0.1, 1, 10 and 100 μM) were used to treat
H2O2-induced BMSCs for 72 h. The ratio of
SA-β-gal-positive cells was reduced with an increase in quercetin
concentration, especially when the concentration >1 μM
(Fig. 1A and B). Additionally,
uninduced BMSCs were treated with quercetin to assess cytotoxicity.
Quercetin had no significant effect on cell viability at
concentrations of 0.0, 0.1, 1.0 and 10.0 μM. However, it
exerted toxic effects on BMSCs when the concentration reached 100
and 1,000 μM (Fig. 1C).
To identify the optimal concentration, the restorative effect of
quercetin on proliferation of senescent BMSCs was further
investigated. The colony formation assay demonstrated the impaired
proliferative ability of senescent BMSCs was rescued by quercetin
at 1 μM (Fig. 1D and E).
Consequently, subsequent experiments were conducted using this
concentration. H2O2 resulted in upregulation
of p21 mRNA expression, which was decreased upon treatment with
quercetin (Fig. 1F). Senescent
cells do not divide but retain the ability to secrete bioactive
molecules known as SASP, which alters the microenvironment for both
senescent and surrounding cells (26). SASP-associated IL6, TNF-α, IL1α
and IL1β showed increased mRNA expression following
H2O2 treatment; however, expression decreased
following quercetin treatment (Fig.
1G). These data aligned with the SA-β-gal staining and colony
formation results showing that quercetin alleviated cellular
senescence phenotype induced by H2O2.
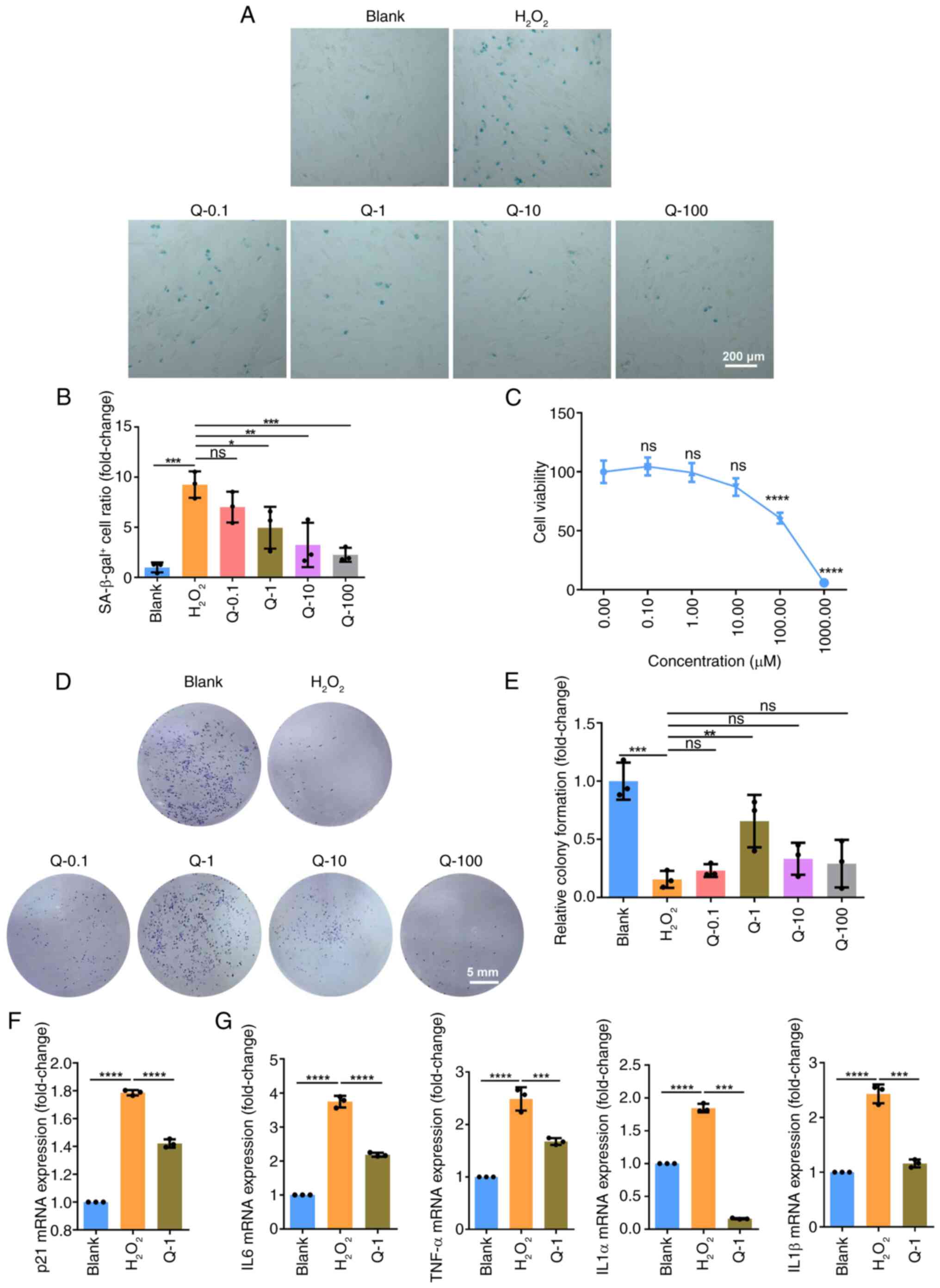 | Figure 1Optimal concentration of Q for
alleviating senescence in BMSCs. (A) BMSCs were exposed to 200
μM H2O2 for 1 h to induce senescence,
followed by treatment with Q for 3 days. Subsequently, SA-β-gal
staining was performed. (B) Ratio of senescent cells was
quantitatively analyzed. (C) Cell Counting Kit-8 assay was
conducted to evaluate the cytotoxic effects of Q on BMSCs. (D)
Colony formation assay and (E) quantitative analysis were used to
investigate the effect of Q on proliferation of senescent BMSCs.
(F) Reverse transcription-quantitative PCR was used to detect mRNA
expression levels of the senescence marker p21 as well as those of
the senescence-associated secretory phenotype-associated (G) IL6,
TNF-α, IL1α and IL1β. Data were quantified as fold-change.
*P<0.05, **P<0.01, ***P<0.001,
****P<0.0001 vs. H2O2. ns,
non-significant; BMSC, bone marrow mesenchymal stem cell; SA-β-gal,
senescence-associated β-galactosidase; Q, quercetin. |
Quercetin restores genomic and epigenomic
instability in senescent BMSCs
The instability of the genome caused by persistent
DNA damage and accumulating mutations, along with epigenomic
instability characterized by loss of heterochromatin structure and
RE suppression are mechanisms underlying senescent phenotypes
(27). The dsDNA break marker
γ-H2AX is a notable molecular indicator for DNA damage and serves
as a hallmark of genomic instability (28). Immunofluorescence revealed a
significant increase in nuclear γ-H2AX signal in
H2O2-induced senescent BMSCs. Quercetin
resulted in a reduced expression of γ-H2AX, demonstrating that
quercetin stabilized the genome of BMSCs (Fig. 2A). In eukaryotic cells, most of
the genome is packaged into condensed and transcriptionally
suppressed heterochromatin. H3K9me2 and 3 are epigenetic markers of
heterochromatin found on transposable elements to ensure their
transcriptional suppression (29). There was a noticeable decrease in
expression of H3K9me3 in H2O2-induced
senescent BMSCs; however, this decrease was rescued by quercetin
(Fig. 2B). Consistently, western
blotting demonstrated a decrease in expression of H3K9me3 in
H2O2-induced senescent BMSCs, which was
rescued by quercetin (Fig. 2E).
The HP1 family binds to H3K9me2 and 3 and interacts with other
proteins to maintain chromatin condensation (30). Both immunofluorescence staining
and western blotting revealed that Hp1α expression decreased in the
H2O2 group but recovered in the quercetin
group (Fig. 2C and E).
Additionally, LAP2, involved in the compaction of heterochromatin
by anchoring it on the inner nuclear membrane (30), exhibited similar expression
patterns as H3K9me3 and Hp1α and was downregulated in the
H2O2 but rescued in the quercetin group
(Fig. 2D and E). These data
demonstrated a decrease in the structural integrity of
heterochromatin during senescence, with quercetin serving a key
role in safeguarding genomic and epigenomic stability, thereby
mitigating senescence.
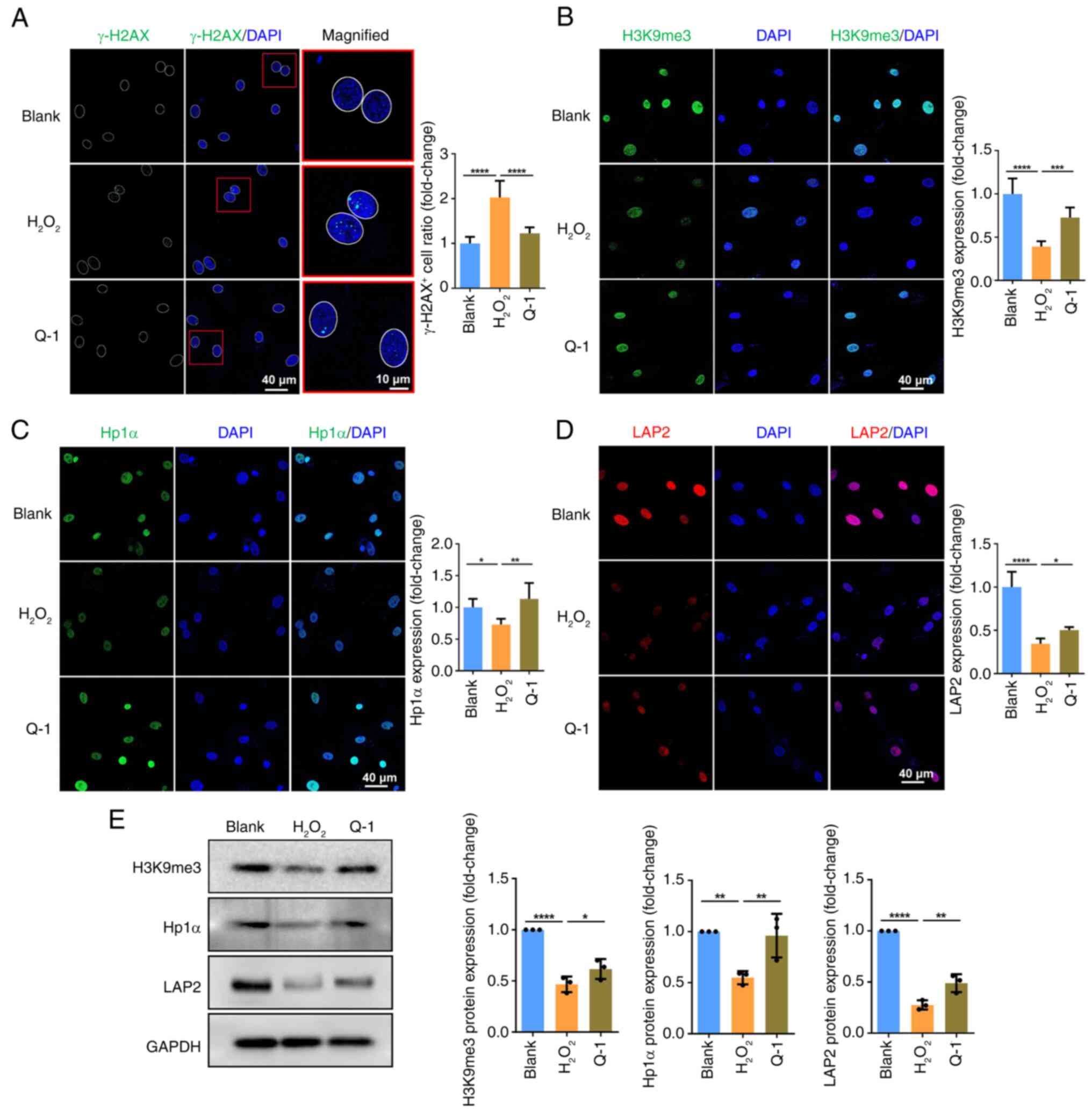 | Figure 2Q decreases DNA damage response and
restores epigenetic characteristics of senescent BMSCs.
Immunofluorescence staining of (A) γ-H2AX, (B) H3K9me3, (C) Hp1α
and (D) LAP2 was performed in BMSCs. (E) Western blotting of
H3K9me3, Hp1α and LAP2 protein expression in BMSCs. Data were
quantified as fold-change. *P<0.05,
**P<0.01, ***P<0.001,
****P<0.0001. BMSC, bone marrow mesenchymal stem
cell; H3K9me3, methylation histone H3 Lys9; LAP2, lamina-associated
polypeptide 2; Hp1α, heterochromatin protein 1α; Q, quercetin. |
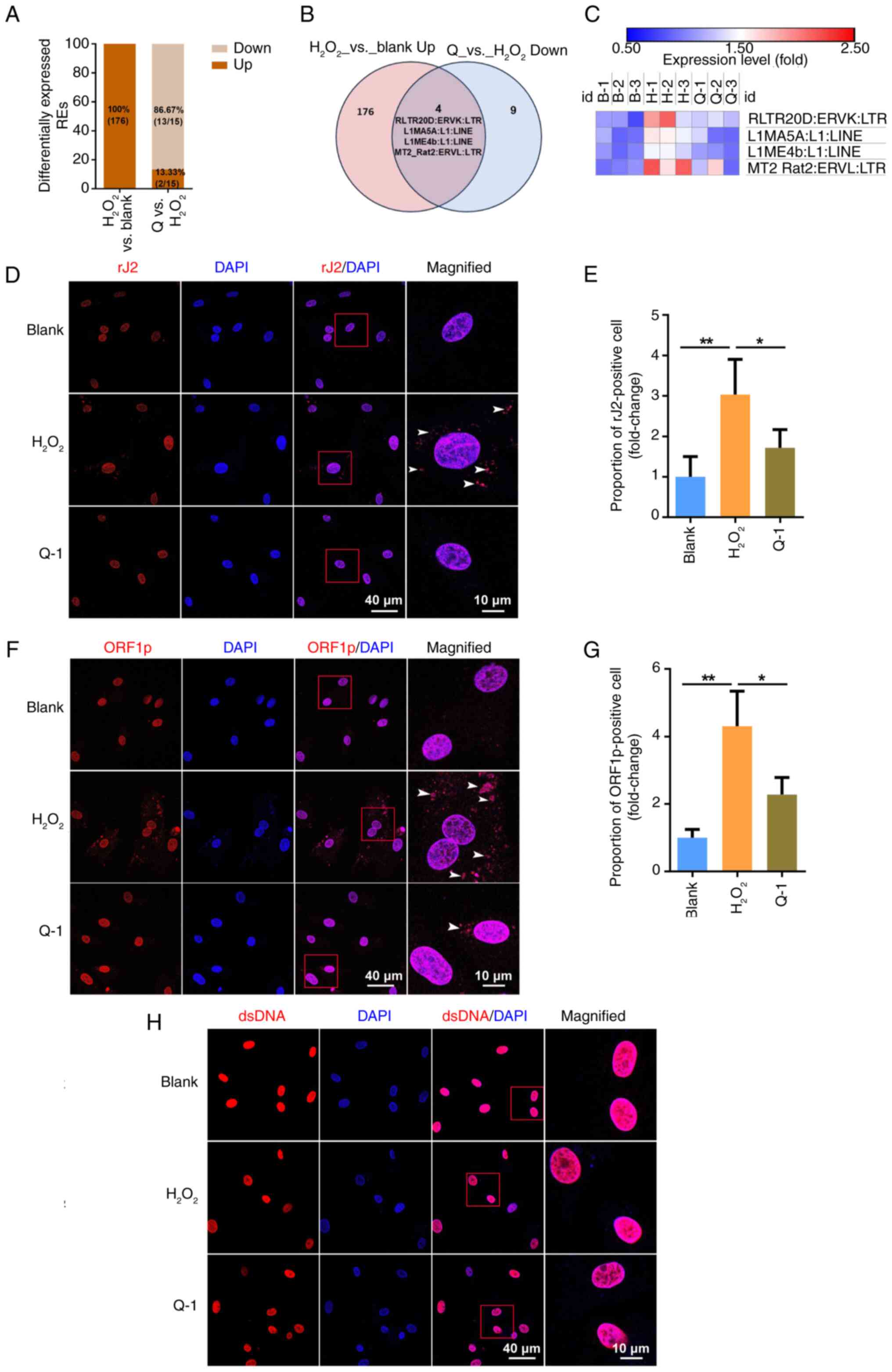
Quercetin ameliorates senescence by
inhibiting the activation of REs and accumulation of cytoplasmic
dsRNA in BMSCs
Loss of suppressive epigenetic regulation and
heterochromatin structure during cell senescence process may elicit
transcriptional activation of REs (9). To elucidate the mechanism by which
quercetin alleviates senescence of BMSCs, bulk mRNA sequencing was
employed to assess the transcriptional levels of REs. A total of
176 REs was differentially expressed after
H2O2 treatment. All differentially expressed
REs were transcribed at elevated levels in
H2O2-induced senescent BMSCs. Quercetin
resulted in differential expression of 15 repeated sequences in
senescent BMSCs, among which 86.67% (13/15) were downregulated
(Fig. 3A). To identify REs
upregulated by H2O2 and rescued by quercetin,
Venn diagram analysis was used. The rescued REs primarily included
the LINE-1 and ERV families (Fig.
3B). The heatmap revealed augmented expression of these REs in
the H2O2 group, whereas down-regulation was
observed in the quercetin group, suggesting the involvement of
LINE-1 and ERVs in quercetin-mediated alleviation of cell
senescence (Fig. 3C). Activation
of RE transcription results in translocation of endogenous RNA to
the cytoplasm and generation of DNA through reverse transcriptase
activity (31).
Immunofluorescence using rJ2 antibody confirmed the accumulation of
cytosolic dsRNA in senescent BMSCs. Quercetin decreased the
proportion of cytoplasmic rJ2-positive BMSCs (Fig. 3D and E). In accordance with
endogenous expression of dsRNA, there was elevated cytoplasmic
protein expression of LINE-1 ORF1p in
H2O2-induced senescent BMSCs, which was be
reversed by administration of quercetin (Fig. 3F and G). RNA derived from
retroelements can also be converted to dsDNA in cytoplasm through
endogenous reverse transcriptase activity; however, no cytoplasmic
dsDNA was detected in any group, indicating that dsDNA was not
involved in the mechanism underlying
H2O2-induced BMSC senescence (Fig. 3H).
Induction of dsRNA triggers RIG-I
receptor signaling and innate immune response activation, whereas
quercetin inhibits this process
Endogenous cytosolic self-nucleic acids can directly
trigger innate immune receptor signaling and induce inflammatory
responses via recognition of RNA or DNA (15,20). To identify the downstream effect
of dsRNA, Venn diagram analysis was employed to identify
quercetin-rescued genes that exhibited differential expression in
response to H2O2 and displayed similar
expression patterns as dsRNA transcripts. A total of 20 genes were
upregulated by H2O2 and differential
expression was reversed by quercetin (Fig. 4A). KEGG analysis revealed that
these genes were significantly enriched in pathways including
'viral protein interaction with cytokine and cytokine receptor',
'cytosolic DNA-sensing pathway', 'RIG-I-like receptor signaling
pathway' and 'toll-like receptor signaling pathway' (Fig. 4B), which are associated with
innate immune response. Moreover, GSEA demonstrated activation of
innate immune response in H2O2-induced
senescence of BMSCs (Fig. 4C).
RIG-I-like receptor signaling pathway serves a key role in
recognizing cytoplasmic dsRNA and initiating an IFN response. To
validate the activation of this pathway, western blotting was
performed, revealing H2O2 treatment
upregulated expression of RIG-I and innate immunity-related protein
p-TBK1. Conversely, quercetin downregulated these proteins
(Fig. 4D and E). To investigate
the role of RIG-I-like receptor signaling pathway in senescence
induction, siRNA was used to knock down RIG-I expression in
rat-derived BMSCs. The effectiveness of knockdown was confirmed by
RT-qPCR and western blotting (Fig.
4F). RIG-I knockdown resulted decreased expression of
downstream innate immune effector IFN-β which was upregulated in
H2O2-induced senescence (Fig. 4G). SA-β-gal staining was used to
assess cell senescence. Notably, RIG-I knockdown led to a decrease
in the proportion of SA-β-gal-positive cells following
H2O2 treatment, thereby confirming the
functional impact of RIG-I on cell senescence (Fig. 4H and I). Overall, the present
findings suggested that cytoplasmic dsRNAs induced by REs
contributed to activation of RIG-I receptor signaling and
subsequent innate immune response. This activation lead to type I
IFN response and proinflammatory cytokine production, ultimately
promoting senescence.
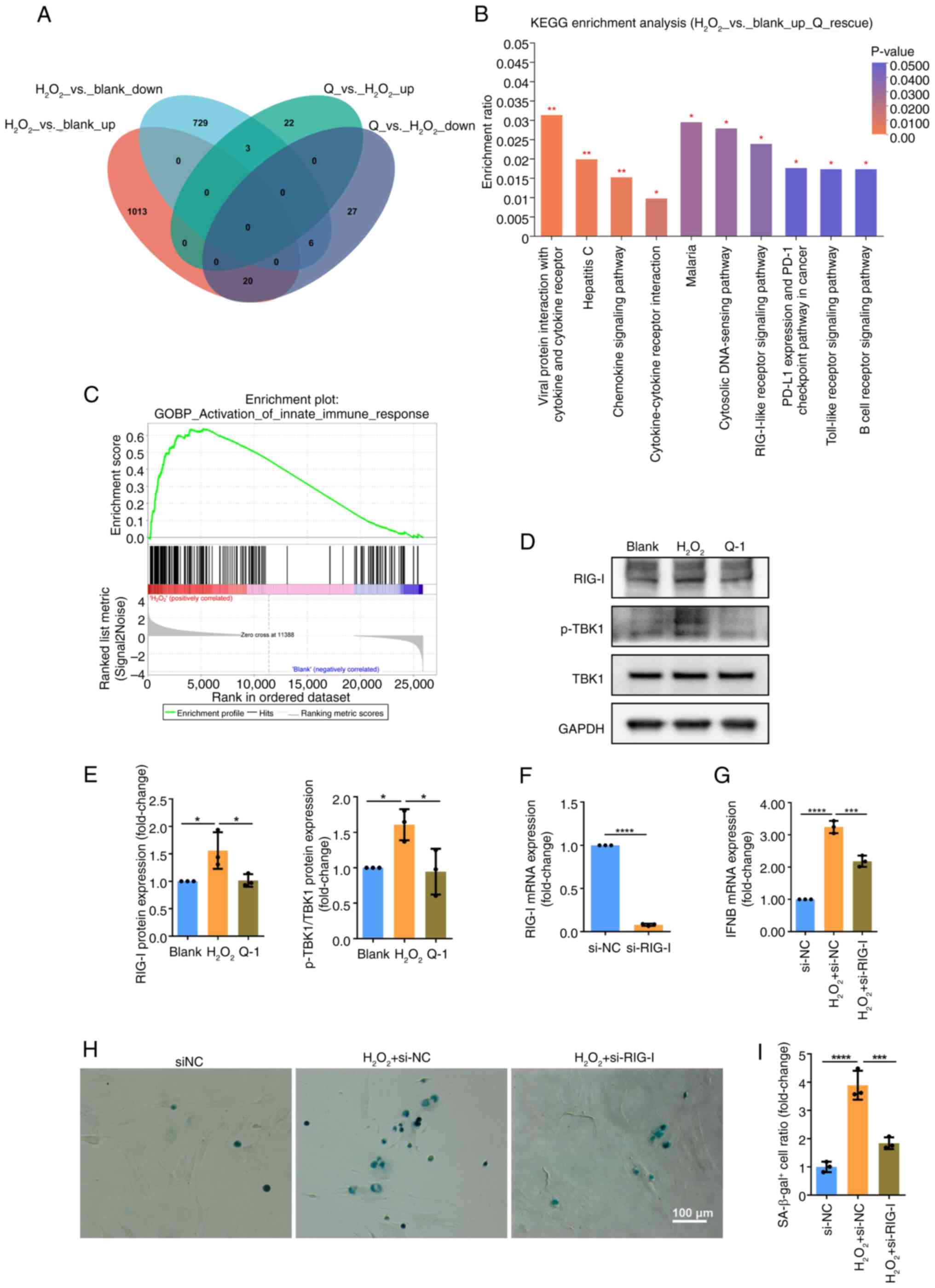 | Figure 4Cytoplasmic double-stranded RNA is
recognized by RIG-I-like receptor signaling pathway and initiates
innate immune response, leading to senescence of BMSCs. (A) Venn
diagram was conducted to identify the genes that showed
differential expression following H2O2
induction and were subsequently rescued by quercetin. (B) KEGG
pathway analysis was used to analyze the differentially expressed
genes that were upregulated by H2O2 induction
and rescued by quercetin. (C) Gene set enrichment analysis was
performed to analyze innate immune response (|normalized enrichment
score|>1, P<0.05, false discovery rate <0.25). (D) Western
blotting was used to assess (E) protein expression of RIG-I, p-TBK1
and TBK1. (F) Validation of RIG-I knockdown efficiency by RT-qPCR
and Western blotting. (G) RT-qPCR detected the mRNA expression of
IFNB. (H) SA-β-gal staining of BMSCs following
H2O2 induction and RIG-I knockdown. (I)
Proportion of SA-β-gal positive cells was quantified. Data were
quantified as fold-change. *P<0.05,
***P<0.001, ****P<0.0001. BMSC, bone
marrow mesenchymal stem cell; KEGG, Kyoto Encyclopedia of Genes and
Genomes; RIG-1, retinoic acid-inducible gene-1; p-TBK1,
phosphorylated TANK-binding kinase 1; SA-β-gal,
senescence-associated β-galactosidase; RT-qPCR, reverse
transcription-quantitative PCR; siNC, small interfering negative
control; Q, quercetin. |
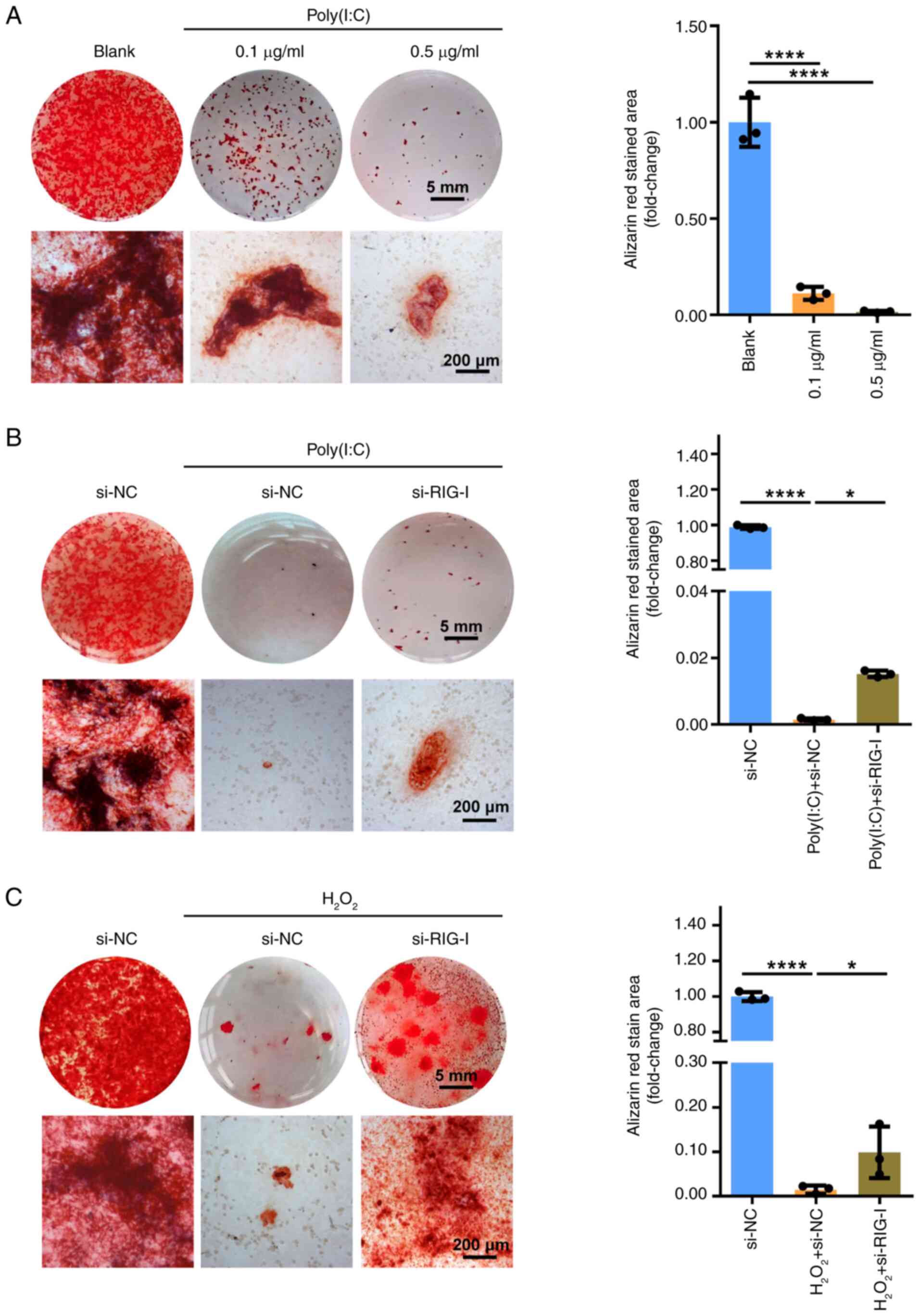
Quercetin promotes osteogenesis in
senescent BMSCs by inhibiting the dsRNA-induced RIG-I receptor
signaling pathway
To evaluate the effect of quercetin on osteogenic
differentiation potential of senescent BMSCs, BMSCs were cultured
in an osteoblast differentiation medium for 7-21 days. Oxidative
stress-induced senescence impaired osteogenic potential of BMSCs,
as indicated by a decrease in ALP and alizarin red staining.
However, quercetin restored the osteogenic differentiation ability
of senescent BMSCs (Fig. 5A and
B). The results were further supported by RT-qPCR of osteogenic
marker genes, including OPG, OCN, OPN and COL1A1. mRNA levels of
osteogenic marker genes (OPG, OCN, OPN, COL1A1) were decreased in
senescent BMSCs, whereas they increased after quercetin
administration (Fig. 5C). The
aforementioned results demonstrated that dsRNA contributes to the
mechanism by which quercetin ameliorates BMSC senescence.
Furthermore, the role of dsRNA and downstream RIG-I receptor
signaling pathway was investigated by adding synthetic dsRNA
analogue Poly(I:C), which activates the RIG-I signaling pathway, to
the osteoblast differentiation medium. Poly(I:C) resulted in a
concentration-dependent decrease in alizarin red staining (Fig. 6A), confirming that dsRNA
decreased osteoblast differentiation potential of BMSCs. Moreover,
the positive area of alizarin red staining improved with si-RIG-I
compared with the si-NC control (Fig. 6B), indicating knockdown of RIG-I
partially rescued the impaired osteoblast differentiation ability
of BMSCs caused by dsRNA. Similarly, knockdown of RIG-I improved
osteogenic differentiation ability of BMSCs impaired by
H2O2 (Fig.
6C). These data indicated that quercetin restored the
osteogenic differentiation ability of senescent BMSCs, potentially
due to the inhibition of the dsRNA-triggered RIG-I receptor
signaling pathway.
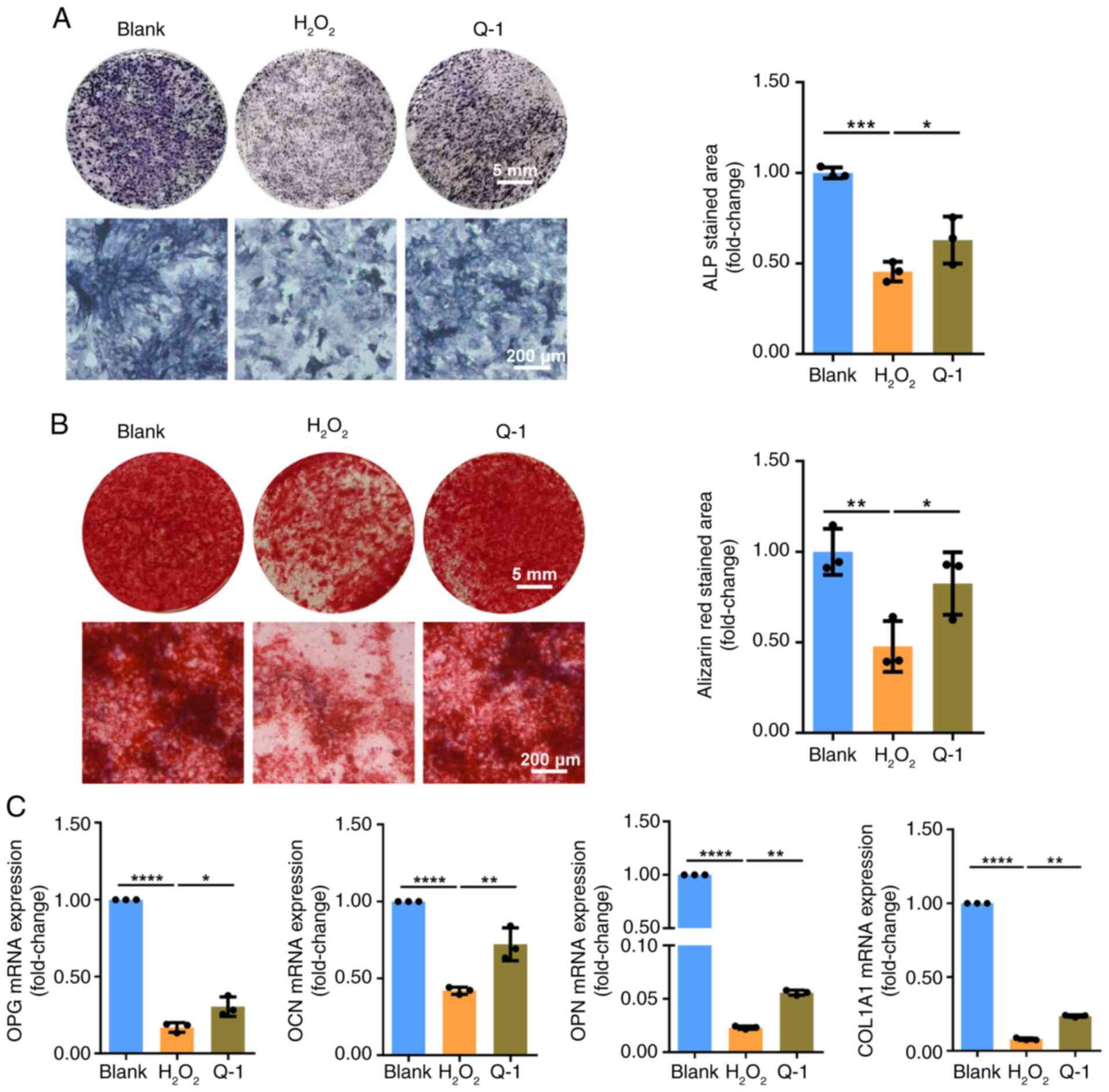 | Figure 5Q restores osteogenic ability of
H2O2-induced senescent BMSCs. (A) Following 7
days of osteogenic induction, ALP staining was used to evaluate the
effect of Q on impaired osteogenic differentiation ability of BMSCs
caused by H2O2. (B) After 14-21 days of
osteogenic induction, alizarin red staining assay was performed.
(C) mRNA levels of osteogenic differentiation-related gene
including OPG, OCN, OPN and COL1A1 were detected by reverse
transcription-quantitative PCR. Data were quantified as
fold-change. *P<0.05, **P<0.01
***P<0.001, ****P<0.0001. OPG,
osteoprotegerin; OCN, osteocalcin; OPN, osteopontin; COL1A1, type I
collagen A1; BMSC, bone marrow mesenchymal stem cell; ALP, alkaline
phosphatase; Q, quercetin. |
Discussion
The senescence of mesenchymal SCs markedly impairs
their ability to replicate and differentiate, thereby decreasing
their potential for tissue regeneration and repair (2,3).
In the present study, epigenomic rearrangements occurred during
cellular senescence in BMSCs, as evidenced by a decrease in the
expression of heterochromatin-related indicators Hp1α, H3K9me3 and
LAP2. This leads to transcriptional activation of REs and
subsequent induction of downstream RNA-sensing pathways. Here,
quercetin stabilizes heterochromatin, inhibits the release of REs
and suppresses the activation of RNA-sensing signaling pathways. As
a result, it mitigates oxidative stress-induced senescence in BMSCs
and enhances their osteogenic capability.
Retrotransposons have coevolved with their host
genomes since the emergence of life (12). Certain beneficial interactions
are established during development, such as facilitating necessary
chromatin status before implantation in mouse embryos through
transcriptional activation of specific LTR and growth LINE-1
elements (31) and using LTR
enhancers to regulate key genes involved in human innate immune
pathways (32). However,
inadvertent activation of retrotransposons in adult somatic tissue
is associated with potential deleterious effects. Both LTR and
LINE-1 RNA and protein are upregulated with age in both
Drosophila (33,34) and mammals, including mice, monkey
and human (35,36). Reactivation of LTR and LINE-1 is
observed in fibroblasts and keratinocytes in various forms of
cellular senescence, including replicative and oncogene- and
irradiation-induced senescence (37-39). Here,
H2O2-induced senescence in BMSCs resulted in
the upregulation of LTR and LINE-1. The present data suggested that
the reactivation of REs serves a crucial role in stem cell
senescence while quercetin reverses this effect.
Retrotransposons are suppressed through a variety of
mechanisms, with the most extensively studied being the formation
of heterochromatin via epigenetic regulation (8,9,34). This occurs by recruiting
chromatin remodeling enzymes and effector proteins to
retrotransposon DNA sequences. Levels of heterochromatin or factors
involved in establishing heterochromatin decrease with age, leading
to an increase in retrotransposon activity (40,41). The present study revealed that
markers of heterochromatin such as H3K9me3 and HP1α, along with
LAP2, were downregulated following
H2O2-indcued senescence of BMSCs. Meanwhile,
transcripts of LINE-1 and ERVs were upregulated in the present
study, indicating that retrotransposons are activated due to
epigenetic reset and loss of heterochromatin structure.
Administration of quercetin rescues these alterations, suggesting
its protective role in stabilizing heterochromatin.
Recently, activation of retrotransposons has been
shown to exert a detrimental impact on aging and disease, primarily
attributed to recognition of nucleic acids derived from
retrotransposons (11,38). dsDNA or dsRNA in the cytoplasm is
generally perceived as an intruding pathogen, triggering innate
immune pathways (18).
Endogenous dsRNA can arise from bidirectional transcription of
retrotransposons or through imperfect base pairing of identical or
similar elements. These molecules are detected by members of the
RIG-I-like receptor family, namely RIG-I, melanoma
differentiation-associated protein 5 and laboratory of genetics and
physiology 2, which bind mitochondrial antiviral signaling (MAVS)
protein. This leads to TBK1 phosphorylation and activation of
downstream IFN-I response as well as release of proinflammatory
cytokines (15) that promote
SASP and cell senescence (42).
RE-derived RNA can be converted to dsDNA by endogenous reverse
transcriptase activity. Cytoplasmic DNA is sensed by cyclic GMP-AMP
synthase, resulting in formation of cyclic GMP-AMP, which
subsequently binds to stimulator of IFN genes thereby initiating
innate immune pathways (20). In
the present study, the cytoplasmic accumulation of dsRNA in
senescent BMSCs was observed, while no dsDNA was detected in the
cytoplasm, indicating that REs induced senescence through the
dsRNA-mediated RIG-I pathway. Additionally, upregulation of RIG-I
and p-TBK1 in senescent BMSCs was revealed. Quercetin effectively
decreased cytoplasmic dsRNA aggregation and expression levels of
RIG-I and p-TBK1. Moreover, knockdown of RIG-I significantly
reversed the senescence of BMSCs and restored the impaired
osteogenic function.
There is substantial evidence indicating a notable
decline in the osteogenic differentiation capacity of BMSCs with
aging, which consequently affects bone regeneration potential
(43,44). However, limited research has been
conducted on the impact of dsRNA and its activation pathway on stem
cell differentiation: The existing studies have yielded conflicting
results, highlighting the need for further investigation (45,46). Here, osteogenic differentiation
ability of BMSCs was significantly decreased after induction of
senescence by H2O2 or administration of
Poly(I:C), a synthetic dsRNA analogue. However, blocking the RNA
sensing pathway via quercetin or knockdown of RIG-I effectively
improved the compromised osteogenic differentiation in senescent
BMSCs. Lou et al (47)
demonstrated that overexpression of RIG-I impaired the
clonogenicity and osteogenic potential of BMSCs. Overall,
involvement of a non-canonical pathway, dsRNA/RIG-I, in the
regulation of osteogenic differentiation in BMSCs was elucidated
and the underlying regulatory mechanism of quercetin in this
signaling cascade was unraveled.
Collectively, the findings of the present study
demonstrated that the release of REs during senescence triggered
activation of the RIG-I RNA sensing pathway and subsequent
downstream innate immune response, resulting in decreased
osteogenic capacity of BMSCs. Quercetin restored epigenetic
regulation and stabilized heterochromatin, thereby inhibiting
release of REs and preventing the decline in osteogenic capacity
(Fig. 7). The present study
offers novel strategies and targets for alleviating senescence of
BMSCs and promoting bone regeneration.
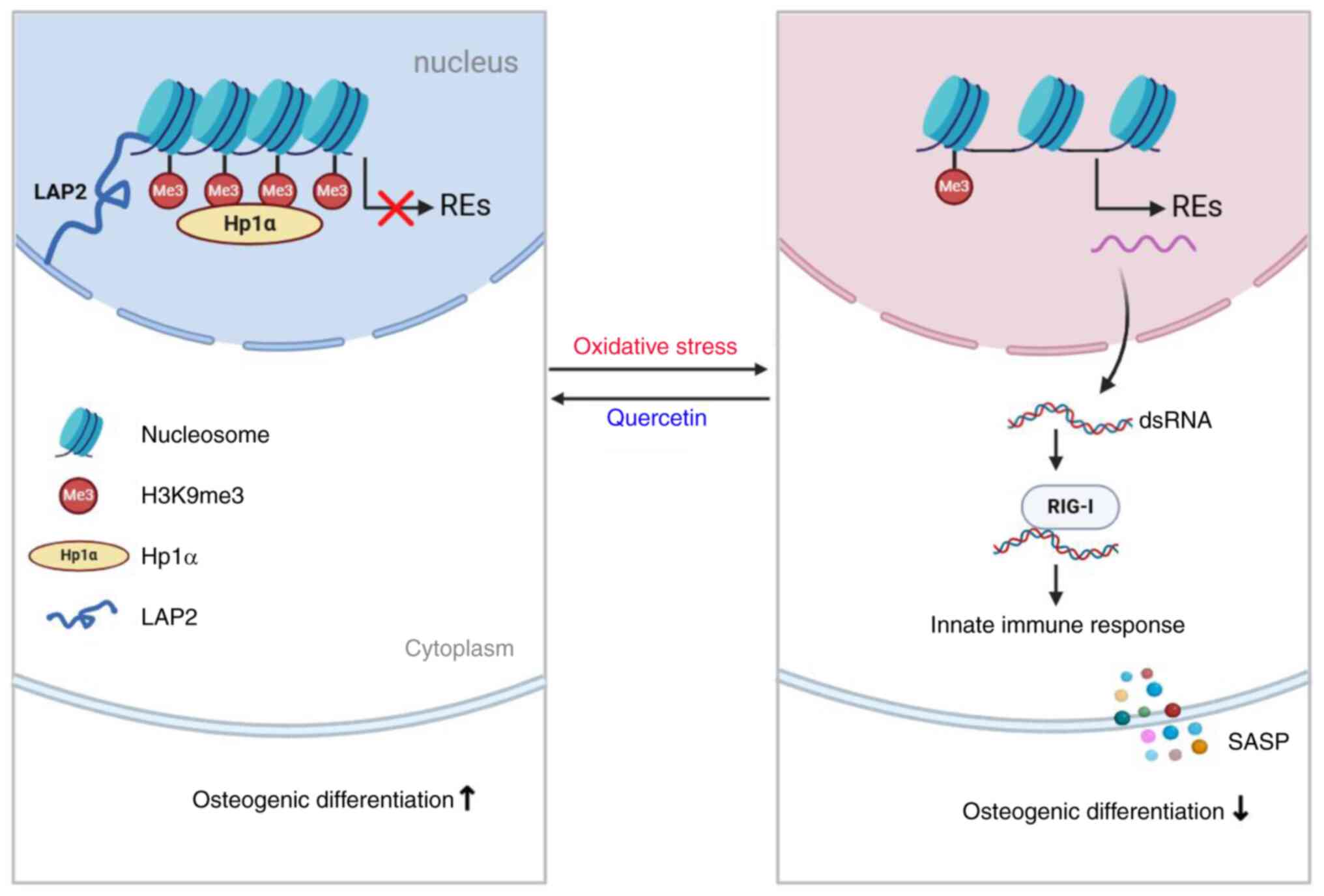 | Figure 7Mechanism by which quercetin
ameliorates senescence and promotes osteogenic differentiation of
bone marrow mesenchymal stem cells. The epigenetic modifications
(H3K9me3, HP1α, and LAP2) undergo changes in the BMSC genome under
oxidative stress. This leads to a loss of heterochromatin structure
and activation of RE transcription. RE transcripts are released
into the cytoplasm, causing an accumulation of dsRNA that triggers
RIG-I receptor signaling and activates the innate immune response.
Subsequently, proinflammatory cytokines released may function as
SASP, promoting BMSC senescence and inhibiting osteogenic
differentiation. However, quercetin can effectively stabilize
heterochromatin structure, thus alleviating cell senescence and
restoring osteogenic differentiation capability.RE, repetitive
elements; H3K9me3, trimethylated histone H3 lysine 9; HP1α,
heterochromatin protein 1α; LAP2, lamina-associated polypeptide 2;
ds, double-stranded; SASP, senescence-associated secretory
phenotype; RIG-I, retinoic acid-inducible gene I. |
Availability of data and materials
The data generated in the present study may be found
in the Sequence Read Archive under accession number PRJNA1156890 or
at the following URL: (https://ncbi.nlm.nih.gov/bioproject/PRJNA1156890).
Authors' contributions
JMP, BC and YTS conceived and designed the
experiments. YTS, CYW, LLW, ZHL and JX performed the experiments
and analyzed the data. JX, BC and JMP confirm the authenticity of
all the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experimental procedures were approved by
the Institutional Animal Care and Use Committee of Sun Yat-Sen
University (Guangzhou, China; approval no.
SYSU-IACUC-2024-002262).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
Funding
The present study was supported by National Natural Science
Foundation of China (grant nos. 82101015, 82202843 and 82301080),
Natural Science Foundation of Guangdong Province (grant nos.
2022A1515012493 and 2024A1515010971) and Basic and Applied Basic
Research Foundation of Guangdong Province (grant no.
2023A1515010272).
References
|
1
|
Zheng C, Chen J, Liu S and Jin Y: Stem
cell-based bone and dental regeneration: A view of
microenvironmental modulation. Int J Oral Sci. 11:232019.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Al-Azab M, Safi M, Idiiatullina E,
Al-Shaebi F and Zaky MY: Aging of mesenchymal stem cell: Machinery,
markers, and strategies of fighting. Cell Mol Biol Lett. 27:692022.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin H, Sohn J, Shen H, Langhans MT and
Tuan RS: Bone marrow mesenchymal stem cells: Aging and tissue
engineering applications to enhance bone healing. Biomaterials.
203:96–110. 2019. View Article : Google Scholar :
|
|
4
|
Lopez-Otin C, Blasco MA, Partridge L,
Serrano M and Kroemer G: The hallmarks of aging. Cell.
153:1194–1217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang W, Li J, Suzuki K, Qu J, Wang P,
Zhou J, Liu X, Ren R, Xu X, Ocampo A, et al: Aging stem cells. A
Werner syndrome stem cell model unveils heterochromatin alterations
as a driver of human aging. Science. 348:1160–1163. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Soto-Palma C, Niedernhofer LJ, Faulk CD
and Dong X: Epigenetics, DNA damage, and aging. J Clin Invest.
132:e1584462022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pathak RU, Soujanya M and Mishra RK:
Deterioration of nuclear morphology and architecture: A hallmark of
senescence and aging. Ageing Res Rev. 67:1012642021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stamidis N and Żylicz JJ: RNA-mediated
heterochromatin formation at repetitive elements in mammals. EMBO
J. 42:e1117172023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McCarthy RL, Kaeding KE, Keller SH, Zhong
Y, Xu L, Hsieh A, Hou Y, Donahue G, Becker JS, Alberto O, et al:
Diverse heterochromatin-associated proteins repress distinct
classes of genes and repetitive elements. Nat Cell Biol.
23:905–914. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Anwar SL, Wulaningsih W and Lehmann U:
Transposable Elements in human cancer: Causes and consequences of
deregulation. Int J Mol Sci. 18:9742017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gorbunova V, Seluanov A, Mita P, McKerrow
W, Fenyö D, Boeke JD, Linker SB, Gage FH, Kreiling JA, Petrashen
AP, et al: The role of retrotransposable elements in ageing and
age-associated diseases. Nature. 596:43–53. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang J, Lu X, Zhang W and Liu GH:
Endogenous retroviruses in development and health. Trends
Microbiol. 32:342–354. 2024. View Article : Google Scholar
|
|
13
|
Ågren JA and Clark AG: Selfish genetic
elements. PLoS Genet. 14:e10077002018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang CR, Burns KH and Boeke JD: Active
transposition in genomes. Annu Rev Genet. 46:651–675. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dumetier B, Sauter C, Hajmirza A, Pernon
B, Aucagne R, Fournier C, Row C, Guidez F, Rossi C, Lepage C, et
al: Repeat element activation-driven inflammation: Role of NFκB and
implications in normal development and cancer? Biomedicines.
10:31012022. View Article : Google Scholar
|
|
16
|
Lanciano S and Cristofari G: Measuring and
interpreting transposable element expression. Nat Rev Genet.
21:721–736. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Copley KE and Shorter J: Repetitive
elements in aging and neurodegeneration. Trends Genet. 39:381–400.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gazquez-Gutierrez A, Witteveldt J, Heras
SR and Macias S: Sensing of transposable elements by the antiviral
innate immune system. RNA. 27:735–752. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Evans TA and Erwin JA:
Retroelement-derived RNA and its role in the brain. Semin Cell Dev
Biol. 114:68–80. 2021. View Article : Google Scholar
|
|
20
|
Miller KN, Victorelli SG, Salmonowicz H,
Dasgupta N, Liu T, Passos JF and Adams PD: Cytoplasmic DNA:
Sources, sensing, and role in aging and disease. Cell.
184:5506–5526. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cui Z and Zhao X, Amevor FK, Du X, Wang Y,
Li D, Shu G, Tian Y and Zhao X: Therapeutic application of
quercetin in aging-related diseases: SIRT1 as a potential
mechanism. Front Immunol. 13:9433212022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Peng D, Chen L, Sun Y, Sun L, Yin Q, Deng
S, Niu L, Lou F, Wang Z, Xu Z, et al: Melanoma suppression by
quercein is correlated with RIG-I and type I interferon signaling.
Biomed Pharmacother. 125:1099842020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nieto M, Konigsberg M and Silva-Palacios
A: Quercetin and dasatinib, two powerful senolytics in age-related
cardiovascular disease. Biogerontology. 25:71–82. 2024. View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Lee SS, Vũ TT, Weiss AS and Yeo GC:
Stress-induced senescence in mesenchymal stem cells: Triggers,
hallmarks, and current rejuvenation approaches. Eur J Cell Biol.
102:1513312023. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ritschka B, Storer M, Mas A, Heinzmann F,
Ortells MC, Morton JP, Sansom OJ, Zender L and Keyes WM: The
senescence-associated secretory phenotype induces cellular
plasticity and tissue regeneration. Genes Dev. 31:172–183. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Korotkov A, Seluanov A and Gorbunova V:
Sirtuin 6: Linking longevity with genome and epigenome stability.
Trends Cell Biol. 31:994–1006. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Feng JX and Riddle NC: Epigenetics and
genome stability. Mamm Genome. 31:181–195. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Padeken J, Methot SP and Gasser SM:
Establishment of H3K9-methylated heterochromatin and its functions
in tissue differentiation and maintenance. Nat Rev Mol Cell Biol.
23:623–640. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shevelyov YY and Ulianov SV: The nuclear
lamina as an organizer of chromosome architecture. Cells.
8:1362019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cosby RL, Chang NC and Feschotte C:
Host-transposon interactions: Conflict, cooperation, and cooption.
Genes Dev. 33:1098–1116. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chuong EB, Elde NC and Feschotte C:
Regulatory evolution of innate immunity through co-option of
endogenous retroviruses. Science. 351:1083–1087. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li W, Prazak L, Chatterjee N, Grüninger S,
Krug L, Theodorou D and Dubnau J: Activation of transposable
elements during aging and neuronal decline in Drosophila. Nat
Neurosci. 16:529–531. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wood JG, Jones BC, Jiang N, Chang C,
Hosier S, Wickremesinghe P, Garcia M, Hartnett DA, Burhenn L,
Neretti N and Helfand SL: Chromatin-modifying genetic interventions
suppress age-associated transposable element activation and extend
life span in Drosophila. Proc Natl Acad Sci USA. 113:11277–11282.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
De Cecco M, Criscione SW, Peterson AL,
Neretti N, Sedivy JM and Kreiling JA: Transposable elements become
active and mobile in the genomes of aging mammalian somatic
tissues. Aging (Albany NY). 5:867–883. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu X, Liu Z, Wu Z, Ren J, Fan Y, Sun L,
Cao G, Niu Y, Zhang B, Ji Q, et al: Resurrection of endogenous
retroviruses during aging reinforces senescence. Cell.
186:287–304.e26. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Colombo AR, Elias HK and Ramsingh G:
Senescence induction universally activates transposable element
expression. Cell Cycle. 17:1846–1857. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Di Giorgio E, Ranzino L, Tolotto V, Dalla
E, Burelli M, Gualandi N and Brancolini C: Transcription of
endogenous retroviruses in senescent cells contributes to the
accumulation of double-stranded RNAs that trigger an anti-viral
response that reinforces senescence. Cell Death Dis. 15:1572024.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Touma F, Lambert M, Martinez Villarreal A,
Gantchev J, Ramchatesingh B and Litvinov IV: The ultraviolet
irradiation of keratinocytes induces ectopic expression of LINE-1
retrotransposon machinery and leads to cellular senescence.
Biomedicines. 11:30172023. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
De Cecco M, Ito T, Petrashen AP, Elias AE,
Skvir NJ, Criscione SW, Caligiana A, Brocculi G, Adney EM, Boeke
JD, et al: L1 drives IFN in senescent cells and promotes
age-associated inflammation. Nature. 566:73–78. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wood JG, Hillenmeyer S, Lawrence C, Chang
C, Hosier S, Lightfoot W, Mukherjee E, Jiang N, Schorl C, Brodsky
AS, et al: Chromatin remodeling in the aging genome of Drosophila.
Aging Cell. 9:971–978. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pérez-Mancera PA, Young ARJ and Narita M:
Inside and out: The activities of senescence in cancer. Nat Rev
Cancer. 14:547–558. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hu M, Xing L, Zhang L, Liu F, Wang S, Xie
Y, Wang J, Jiang H, Guo J, Li X, et al: NAP1L2 drives mesenchymal
stem cell senescence and suppresses osteogenic differentiation.
Aging Cell. 21:e135512022. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xie Y, Han N, Li F, Wang L, Liu G, Hu M,
Wang S, Wei X, Guo J, Jiang H, et al: Melatonin enhances
osteoblastogenesis of senescent bone marrow stromal cells through
NSD2-mediated chromatin remodelling. Clin Transl Med. 12:e7462022.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Niepmann ST, Willemsen N, Boucher AS, Stei
M, Goody P, Zietzer A, Bulic M, Billig H, Odainic A, Weisheit CK,
et al: Toll-like receptor-3 contributes to the development of
aortic valve stenosis. Basic Res Cardiol. 118:62023. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Khokhani P, Rahmani NR, Kok A, Öner FC,
Alblas J, Weinans H, Kruyt MC and Croes M: Use of therapeutic
pathogen recognition receptor ligands for osteo-immunomodulation.
Materials (Basel). 14:11192021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lou Q, Jiang K, Xu Q, Yuan L, Xie S, Pan
Y, Chen J, Wu J, Zhu J, Jiang L and Zhao M: The RIG-I-NRF2 axis
regulates the mesenchymal stromal niche for bone marrow
transplantation. Blood. 139:3204–3221. 2022. View Article : Google Scholar : PubMed/NCBI
|