Introduction
Prostate cancer is a heterogeneous disease due to
which the search for genetic causes involved in the pathogenesis
remains a challenge. Deregulated expression of several genes such
as EZR and OCT1 were identified, and the involvement
of the aberrant expression of these genes was reported in the
pathogenesis of PCa (1,2). The variety in biological behaviour of
PCa demands identification of biomarkers that may distinguish a
slow growing cancer from a more aggressive cancer with a potential
to metastasize (3).
The androgen receptor is a ligand-dependent zinc
finger DNA-binding protein that is involved in the regulation of
transcription of a variety of gene derivatives (4). The unique feature of PCa is its
dependency of androgen for its growth and survival. Several novel
androgen-regulated genes have been identified, some of which may be
important in the regulation of prostate cell invasiveness (5). In general, androgens activate the
androgen receptors which in turn control the expression of androgen
receptor response elements (ARE) containing genes due to which
current research targets androgen-based therapies for PCa.
Epigenetic factors are also known to mediate the
expression of several genes. DNA methylation is one of the
epigenetic mechanisms (6) and it
occurs in mammals mostly at cytosines within CpG dinucleotide.
Several studies have been proposed that DNA hypomethylation can
cause activation of oncogenes and genetic instability, whilst
hypermethylation is associated with inappropriate gene silencing
(7). For instance, Lin et
al reported the role of hypermethylation in the silence of
glutathione-S-transferase P1 (GSTP1) expression in PCa
(8). It is reported that
GSTP1 is hypermethylated in nearly all human prostate
cancers and its promoter DNA methylation level is able to
differentiate between BPH and different grades of prostate
adenocarcinoma (9–11).
PRAC is a novel gene encoding for the 382
nucleotide RNA, and it specifically expressed in prostate tissue,
rectum and colon. The sequence tag database is a potential source
for discovery of new genes (12,13),
and it was used to find the PRAC gene (14). The PRAC gene is located on
chromosome 17 at position 17q21, 4 kbp downstream from the
homeodomain Hoxb-13 gene. To date, there is no specific
study on the prognostic role and regulatory factors that govern the
expression of PRAC gene. In this study, we have identified
the distinct difference in the expression patterns of PRAC protein
between PCa and BPH tissues. Additionally, regulatory role of
methylation in the expression of PRAC gene was
demonstrated.
Materials and methods
Cell lines and clinical tissues
In total, five PCa cell lines including DU145, PC3,
LNCaP, LNCaP-R and CWR22R were obtained from the American Type
Culture Collection (Manassas VA, USA). DU145 and PC3 are known as
aggressive and androgen insensitive cell lines whereas LNCaP, LNCaP
and CWR22R are relatively less aggressive and androgen sensitive
cells. Immunohistochemical analysis was carried for the prostate
specimen of 54 patients with equal number of BPH and PCa tissues
were represented (Table I). All
the clinical samples were approved by the Research Ethics Committee
of Chang Gung Memorial Hospital (Tao-Yuan, Taiwan) with the
approval no. 95-0345B.
 | Table I.PRAC immunohistochemical score from
benign and cancer prostate tissues. |
Table I.
PRAC immunohistochemical score from
benign and cancer prostate tissues.
| Clinical
samples | Immunostaining
| P-value |
|---|
| Strong | Moderate | Weak |
|---|
| Tissues | | | | |
| BPH (n=27) | 21 (77.7%) | 6 (22.2%) | 0 | <0.0001 |
| PCa (n=27) | 2 (7.4%) | 7 (25.9%) | 18 (66.7%) | |
| Pathological
GS | | | | |
| ≤3+4 (n=7) | 2 (28.5%) | 3 (42.8%) | 2 (28.5%) | |
| ≥4+3 (n=20) | 0 | 4 (20%) | 16 (80%) | |
Cell culture
The cells were cultured in RPMI-1640 containing 10%
FBS, 50 mg/ml each of penicillin and streptomycin and the medium
was replaced every alternative day. To synchronize the cell cycle,
all prostate cells used in this study were incubated in RPMI media
without serum for 24 h. These cancer cell lines were further used
to analyse the expression of PRAC gene both at mRNA and protein
levels.
RNA extraction and qRT-PCR
Total RNA from the cultured PCa cell lines were
extracted by TRIzol (Invitrogen, Carlsbad, CA, USA) using the
protocol recommended by the manufacturers. Total RNA was quantified
and analysed by spectrophotometry (NanoDrop Technology Inc.,
Wilmington, DE, USA). The cDNA was prepared by using SuperScript™
III First-Strand Synthesis SuperMix kit (Invitrogen). qRT-PCR
reactions were performed using SYBR-Green SuperMix (Bio-Rad,
Hercules, CA, USA) in 20 μl total volume and a Bio-Rad
iCycler iQ Real-Time Detection System according to the
manufacturer’s instructions. Primers for target genes including
PRAC (forward, 5′-GCCCATTTCTCAGATCA AGG-3′; reverse,
5′-GGTCTCGCCCAGTAGATGTT-3′), PSA (forward,
5′-AGGTCAGCCACAGCTTCCCA-3′; reverse, 5′-GGGCAGGTCCATGACCTTCA-3′),
GSTP1 (forward, 5′-CAATACCATCCTGCGTCACCT-3′; reverse,
5′-GCAAG ACCTTCATTGTGGGAG-3′) and β-actin (forward, 5′-CATG
TACGTTGCTATCCAGGC-3′; reverse, 5′-ATCGTGCGTGA CATTAAGGAG-3′) were
designed using Primer 3 online tool (15). PCR reactions were performed in
triplicate, and relative expression level of target genes in all
the cell lines was calculated by normalizing to β-actin
expression levels using the comparative threshold cycle (CT)
method. CT represents the cycle numbers at which the amplification
reaches a threshold level chosen to lie in the exponential phase of
all PCR reactions. Data were analysed using the iCycle iQ system
software (Bio-Rad).
Protein extraction and immunoblot assay
of human PRAC, AR and β-actin
Cultured cells were lysed in Pro-Prep™ Protein
Extraction Solution (Intron Biotechnology, Seoul, Korea) according
to the manufacturer’s instructions. Proteins were analysed by
spectrophotometry (NanoDrop Technology Inc.). For western blot
analysis, cell lysates were separated by using 4–20% Tris-glycine
precast gel (Bio-Rad) and transfered to 0.2-μm Immobilon-PSQ
PVDF membrane (Millipore, Billerica, MA, USA). Later, membrane was
blocked with 5% non-fat milk in TBS-T buffer (150 mM NaCl, 10 mM
Tris/pH 8.0 and 0.05% Tween-20) at room temperature for 1 h or
overnight at 4°C. Then, the membranes were immunoblotted with
diluted PRAC primary antibodies (1:500) (Abnova, Walnut, CA, USA)
for 1 h at room temperature or overnight at 4°C, followed by
incubation with secondary antibodies (AP124P, Chemicon, Millipore)
for 1 h at room temperature. Blots were visualized by a
chemiluminescence ECL system (Millipore).
Immunohistochemistry
IHC analysis was performed after approval from
institutional review board. PCa and BPH tissues embedded in
paraffin were cut into 5-mm sections. The sections were dewaxed in
xylene and rehydrated in ethanol (Sigma Chemical Co., St. Louis,
MO, USA). For antigen retrieval, paraffin sections were boiled for
20 min in 10 mM sodium citrate buffer. Endogenous peroxidase
activity was blocked by treatment with 3% hydrogen peroxide (Merck,
Hohenbrunn, Germany) for 10 min and tissue sections were then
incubated with 1% bovine serum albumin (Invitrogen) for 30 min. The
sections were then incubated with the diluted PRAC primary antibody
(1:200) (Abnova). After washing in TBS containing 0.1% Tween-20,
the sections were then further incubated with SuperPicture HRP
Polymer conjugate antibody (Zymed/Invitrogen, Carlsbad, CA, USA)
for 30 min. The peroxidase reaction was visualised using a liquid
DAB substrate kit (Dako, Carpinteria, CA, USA). All sections were
counterstained with hematoxylin (Dako) for 20 sec. Finally, the
sections were microscopically observed. Immunostaining of PRAC was
scored independently. IHC stains were scored: 0, no staining; 1,
weak staining; 2, moderate staining; and 3, strong staining; based
on their intensity. In addition, IHC stains were graded on a
semi-quantitative scale according to the prevalence of nuclear
fraction positivity within the tumor cells (0, <10%; 1+, 10–25%;
2+, 25–50%; 3+, 50–75%; and 4+, >75%). Nuclear positivity of
PRAC protein was scored by multiplying the percentage of positive
cells (P) by the intensity (I).
Hormonal treatment
LNCaP cells were seeded at low density (about
1×106 cells) in 100-mm cell culture plates. The medium
was then substituted with phenol red-free RPMI-1640 with 5% Dextran
coated charcoal-treated FBS (HyClone Inc., Logan, UT, USA) in order
to avoid any interfering factor that might modify the metabolic
ability of the cells. The cells were preconditioned for 24 h in
medium containing two different flasks prior to exposure to 1 nM
DHT and 1 μM flutamide (Sigma Chemical Co.). In one flask, 1
μM of flutamide was added into the medium 1 h before the DHT
inclusion. Control flasks received vehicle (medium with ethanol
only). Following 24 h of incubation in androgen containing medium,
the cells were washed twice with phosphate-buffered saline (PBS, pH
7.4) and harvested by scraping and transferred to sterile plastic
tubes, and stored at −80°C until analysis. Cells from the control
flasks were harvested in the same way.
5-aza-CdR treatment
DU145 and PC3 cells were seeded at low density
(about 1-1.5×106 cells) in 100-mm cell culture plates.
After 24 h, the cell lines were treated with different final
concentrations of 5-aza-CdR (Sigma Chemical Co.) at 1, 5 and 10
μM. Since the downregulation of PRAC was identified
in well known aggressive cell lines including DU145 and PC3,
restoration of PRAC mRNA was analysed after 4 days of
treatment with 5-aza-CdR. GSTP1 expression was
simultaneously analysed as positive control.
Statistical analysis
The data, representative of three or more indepen
dent experiments are presented as the mean ± SEM. To reveal the
statistical significance, unpaired t-test was carried out using
GraphPad software in which p-value <0.05 was considered as
significant.
Results
Expression analysis of PRAC in PCa cell
lines
Initially, PRAC expression was screened in
five PCa cell lines including DU145, PC3, LNCaP, LNCaP-R and CW22R.
The results of qRT-PCR and immunoblot assays showed the high
expression level of PRAC in all the androgen sensitive cells
including LNCaP, LNCaP-R and CW22R, whereas its expression was
significantly downregulated (p<0.05) in the androgen insensitive
cell lines DU145 and PC3 (Fig. 1).
Moreover, immunoblot assay revealed a strong association of PRAC
protein levels with the levels of AR protein (Fig. 1B). These results initiate a
speculation that PRAC might be regulated by androgens.
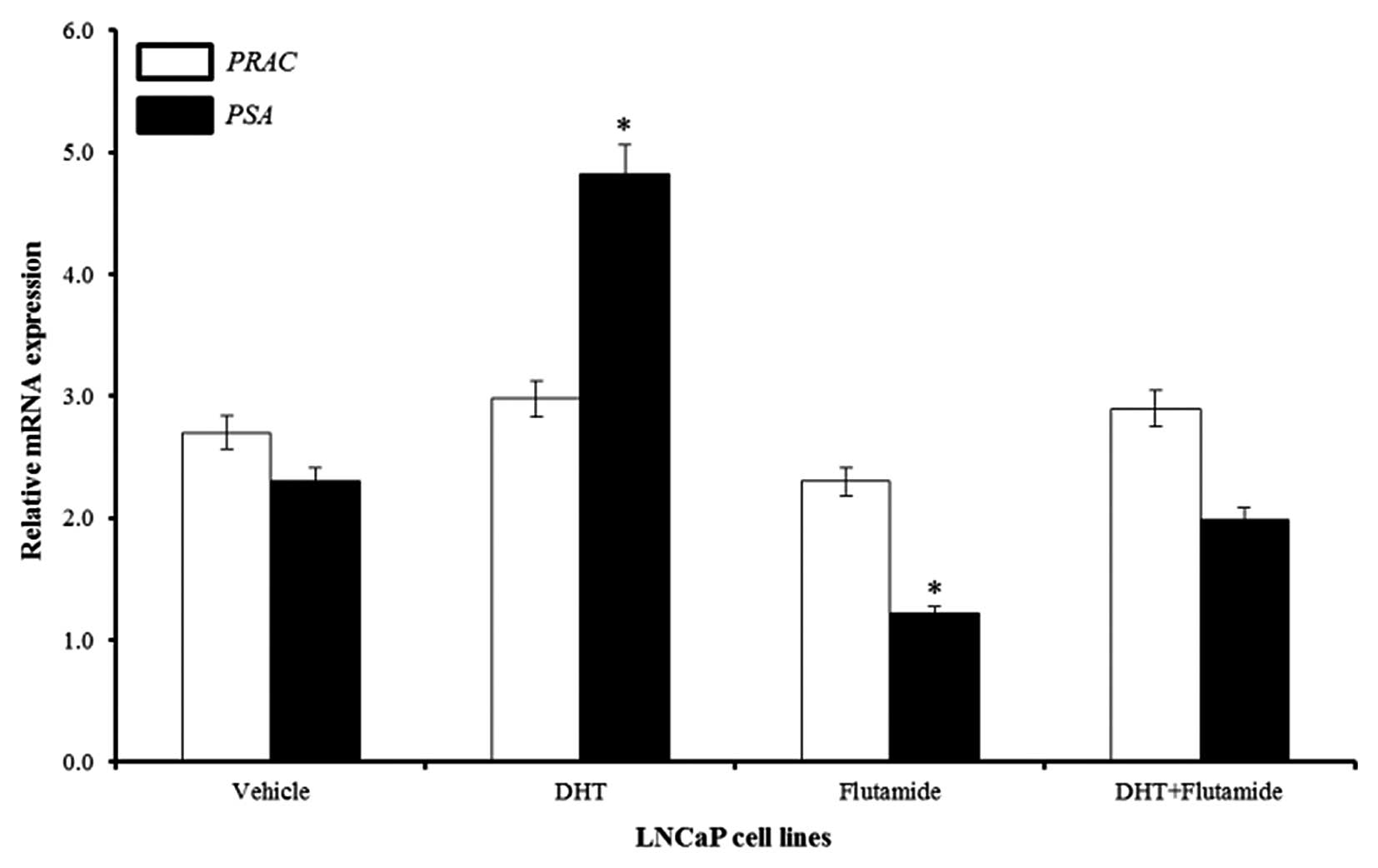
Evaluation of the androgen effect on PRAC
expression
Since higher PRAC levels were identified in
androgen sensitive cell lines, we treated LNCaP cell lines with the
various concentrations of DHT. Our quantitative expression analysis
showed slight changes in the expression of PRAC after the
DHT treatment. To further clarify this difference in PRAC
expression, we treated the LNCaP cell line with flutamide, an
anti-androgen. However, there was no significant change in the
expression of PRAC after the treatment with flutamide,
whereas the well known androgen regulated candidate PSA,
showed significant response to the androgen treatment (p<0.05)
(Fig. 2). Our results indicate
that androgens do not have significant role in the regulation of
PRAC expression, and thus the PRAC is expressed in an
androgen-independent manner.
The PRAC gene contains a CpG island near
the transcription start site
Analysis of the PRAC gene (near the
transcription start site) using MethPrimer CpG Island finder
(16) revealed the presence of two
CpG islands. The first island totals 6 CpG pairs and spans a region
of 110 bp (+27 to +136). This CpG island covers the region
downstream of the transcription start site, the first exon (which
is translatable), and part of the first intron adjacent to exon 1.
The second island totals 9 CpG pairs and spans a region of 142 bp
(+213 to + 354) and covers a part of intron 1 (Fig. 3A). This identification led us to
further study the regulatory role of methylation on PRAC
expression.
PRAC expression analysis in
methyltransferase inhibitor treated DU145 and PC3 cell lines
We treated DU145 and PC3 cell lines with the
5-aza-CdR to identify the effect of methylation in the control of
PRAC expression. Interestingly, significant increment in the
expression of PRAC was found when the DU145 and PC3 cell
lines were treated with higher concentration (10 μM)
compared to lower concentrations of 5-aza-CdR treated and wild-type
cell lines (p<0.05). GSTP1 expression was simultaneously
analysed as positive control, and the distinct difference in the
expression of GSTP1 was also identified (p<0.05)
(Fig. 3B). These results provide
initial evidence for the role of methylation in the regulation of
PRAC expression.
IHC analysis revealed aberrant expression
of PRAC in prostate cancer
IHC analysis was carried to evaluate the correlation
of the PRAC immunoreactivity in BPH and PCa tissues (Fig. 4A). We noted that there is an
apparent trend in the reduction of PRAC expression in aggressive
cancer. BPH tissues from most patients (77.7%) frequently showed
profound nuclear immunostaining, whereas a majority of cancer
tissues (66.7%) showed weak immunoreaction which is supporting the
downregulation of PRAC protein in cancer tissue. Additionally, most
of cancer patients (80%) with high Gleason scores (≥4+3) showed
weak nuclear positivity of PRAC protein compared to cancer tissues
with low Gleason score (≤3+4) (Table
I). The difference between PRAC staining intensity in cancer
and benign samples was found to be significant (P<0.0001)
(Fig. 4B).
Discussion
Clinical therapeutic effectiveness of PCa has been
challenged by significant cellular heterogeneity and limited
understanding of the genetic elements governing disease progression
(17). Although, it was found that
PRAC is expressed specifically in prostate, rectum and
colon, there were no studies on the importance of the PRAC
gene in the PCa pathogenesis and regulatory factors that govern the
expression of the gene. Therefore, we analysed its expression
patterns in human benign and malignant prostate tissues. Initial
expression analysis demonstrated the expression of PRAC protein is
possibly related to the AR expression (Fig. 1B) which delineates that androgen
receptor (AR) may have an essential role in the regulation of
PRAC expression. It is well known that AR plays a central
role in regulating the growth of the PCa cells (18). Therefore, research has been
focusing on the role of AR regulated genes during PCa progression.
Several previous findings indicated that the expression of the
PSA gene in LNCaP cells is regulated by androgens (19,20).
Due to its tissue specificity and androgen inducibility, the
PSA gene has been used as a reference gene to study androgen
action in PCa. In the present study, we treated the LNCaP cell line
(endogenous AR containing line) with DHT and flutamide. Contrary to
expectation, PRAC was found to be regulated in an
androgen-independent manner (Fig.
2). PRAC is located at chromosome band 17q21 immediately
adjacent to the PRAC2 and Hoxb-13 genes, which have
also been proposed as markers of human PCa (14,21).
In addition, Hoxb-13 was reported to be expressed only in
the prostate and colon of mice in an androgen-independent manner,
which is consistent with our expression studies of PRAC in
LNCaP cell lines (22). This
increases the possibility that PRAC and Hoxb-13 are
under the same transcriptional control.
Studies have proven that epigenetic alterations are
common events in cancer including PCa, which may lead to aberrant
expression of critical genes such as tumor suppressors and
oncogenes (23,24). Moreover, it was found that various
epigenetic inhibitors such as DNA methyltransferase inhibitors,
5-aza-cytidine (5-aza-CR or Vidaza) and its more potent analogue
5-aza-CdR or decitabine can chemically reverse the expression of
genes altered due to the epigenetic alterations (23,24).
As it was well established that these inhibitors effectively
restore the expression of GSTP1 in PCa cells, herein to
investigate the efficacy of 5-aza-CdR in PCa cells, we used
GSTP1 as reference gene (25). Interestingly, significant increment
in the expression of PRAC was found when the DU145 and PC3
cell lines were treated with higher concentration of (10 μM)
compared to lower concentrations of 5-aza-CdR treated cell line and
wild-type cell line (p<0.05). To further support the efficacy of
5-aza-CdR in PCa cells, the distinct difference in the expression
of GSTP1 was identified (p<0.05) in DU145 and PC3 cells
(Fig. 3B). These findings indicate
that understanding the molecular mechanism for the methylation of
the PRAC gene may provide insights into the development of
PCa.
To our knowledge, only one study with a few samples
has demonstrated expression pattern of PRAC in PCa (14). Moreover, it has been identified
that location of PRAC gene has been shown to undergo loss of
heterozygosity (LOH) in PCa (26,27).
If the PRAC genes were located within the LOH region, there
could possibly be a reduction in PRAC/PRAC2 expression in
PCa. These findings urged us to analyse the expression patterns of
PRAC protein in cancerous versus benign prostate tissue using a
larger cohort of well-defined PCa patients. Interestingly, PRAC
protein was found to be downregulated in cancerous tissue as
compared to BPH (p<0.0001) (Fig.
4B). These results highlight the possible correlation between
PRAC expression and invasiveness of PCa. Moreover, we are
the first to demonstrate the epigenetic factor, methylation,
effects on PRAC gene expression in prostate carcinoma cells,
and also androgen-independent regulation of PRAC expression.
The decreased expression of PRAC protein could be due to the effect
of methylation, which needs to be further studied. In conclusion,
these results suggest that PRAC protein may play an important role
in the pathogenesis and probably can be a biomarker for PCa.
Acknowledgements
This study was supported partly by the
grant from National Science Council, Taiwan: 101-2221-E-027-001 and
101-2314-B-182A-019, and Chang Gung Memorial Hospital, Taiwan:
CMRPG391781-CMRPG391782, CMRPG3B1601 and NMRPD1A1271. The authors
also cordially acknowledge Mr. Yu-Hsin Chang, Ms. Nuan-Yu Huang,
Ms. Ya-Ping Liu and Ms. Pei-Yi Wu for the technical support.
References
|
1.
|
Pang ST, Fang X, Valdman A, Norstedt G,
Pousette A, Egevad L and Ekman P: Expression of ezrin in prostatic
intraepithelial neoplasia. Urology. 63:609–612. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Obinata D, Takayama K, Urano T, et al:
Oct1 regulates cell growth of LNCaP cells and is a
prognostic factor for prostate cancer. Int J Cancer. 130:1021–1028.
2012. View Article : Google Scholar
|
|
3.
|
Garnick MB and Fair WR: Combating prostate
cancer. Sci Am. 279:74–83. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Gelmann EP: Molecular biology of the
androgen receptor. J Clin Oncol. 20:3001–3015. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Pang ST, Dillner K, Wu X, Pousette A,
Norstedt G and Flores-Morales A: Gene expression profiling of
androgen deficiency predicts a pathway of prostate apoptosis that
involves genes related to oxidative stress. Endocrinology.
143:4897–4906. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Sharma S, Kelly TK and Jones PA:
Epigenetics in cancer. Carcinogenesis. 31:27–36. 2010. View Article : Google Scholar
|
|
7.
|
Feinberg AP, Ohlsson R and Henikoff S: The
epigenetic progenitor origin of human cancer. Nat Rev Genet.
7:21–33. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Lin X, Tascilar M, Lee WH, et al:
GSTP1 CpG island hypermethylation is responsible for the
absence of GSTP1 expression in human prostate cancer cells. Am J
Pathol. 159:1815–1826. 2001. View Article : Google Scholar
|
|
9.
|
Perry AS, Foley R, Woodson K and Lawler M:
The emerging roles of DNA methylation in the clinical management of
prostate cancer. Endocr Relat Cancer. 13:357–377. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Hanson JA, Gillespie JW, Grover A, et al:
Gene promoter methylation in prostate tumor-associated stromal
cells. J Natl Cancer Inst. 98:255–261. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Singal R, van Wert J and Bashambu M:
Cytosine methylation represses glutathione S-transferase P1
(GSTP1) gene expression in human prostate cancer cells.
Cancer Res. 61:4820–4826. 2001.PubMed/NCBI
|
|
12.
|
Okubo K and Matsubara K: Complementary DNA
sequence (EST) collections and the expression information of the
human genome. FEBS Lett. 403:225–229. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Burke J, Wang H, Hide W and Davison DB:
Alternative gene form discovery and candidate gene selection from
gene indexing projects. Genome Res. 8:276–290. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Liu XF, Olsson P, Wolfgang CD, Bera TK,
Duray P, Lee B and Pastan I: PRAC: A novel small nuclear
protein that is specifically expressed in human prostate and colon.
Prostate. 47:125–131. 2001. View Article : Google Scholar
|
|
15.
|
Rozen S and Skaletsky H: Primer3 on the
WWW for general users and for biologist programmers. Methods Mol
Biol. 132:365–386. 2000.PubMed/NCBI
|
|
16.
|
Li LC and Dahiya R: MethPrimer: designing
primers for methylation PCRs. Bioinformatics. 18:1427–1431. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Andreoiu M and Cheng L: Multifocal
prostate cancer: biologic, prognostic, and therapeutic
implications. Hum Pathol. 41:781–793. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Visakorpi T, Hyytinen E, Koivisto P, et
al: In vivo amplification of the androgen receptor gene and
progression of human prostate cancer. Nat Genet. 9:401–406. 1995.
View Article : Google Scholar
|
|
19.
|
Montgomery BT, Young CY, Bilhartz DL,
Andrews PE, Prescott JL, Thompson NF and Tindall DJ: Hormonal
regulation of prostate-specific antigen (PSA) glycoprotein in the
human prostatic adenocarcinoma cell line, LNCaP. Prostate.
21:63–73. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Lee C, Sutkowski DM, Sensibar JA, et al:
Regulation of proliferation and production of prostate-specific
antigen in androgen-sensitive prostatic cancer cells, LNCaP, by
dihydrotestosterone. Endocrinology. 136:796–803. 1995.PubMed/NCBI
|
|
21.
|
Olsson P, Motegi A, Bera TK, Lee B and
Pastan I: PRAC2: a new gene expressed in human prostate and
prostate cancer. Prostate. 56:123–130. 2003. View Article : Google Scholar
|
|
22.
|
Sreenath T, Orosz A, Fujita K and
Bieberich CJ: Androgen-independent expression of hoxb-13 in
the mouse prostate. Prostate. 41:203–207. 1999. View Article : Google Scholar
|
|
23.
|
Jones PA and Taylor SM: Cellular
differentiation, cytidine analogs and DNA methylation. Cell.
20:85–93. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Bender CM, Gonzalgo ML, Gonzales FA,
Nguyen CT, Robertson KD and Jones PA: Roles of cell division and
gene transcription in the methylation of CpG islands. Mol Cell
Biol. 19:6690–6698. 1999.PubMed/NCBI
|
|
25.
|
Chiam K, Centenera MM, Butler LM, Tilley
WD and Bianco-Miotto T: GSTP1 DNA methylation and expression
status is indicative of 5-aza-2′-deoxycytidine efficacy in human
prostate cancer cells. PLoS One. 6:e256342011. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Cheng L, Bostwick DG, Li G, Wang Q, Hu N,
Vortmeyer AO and Zhuang Z: Allelic imbalance in the clonal
evolution of prostate carcinoma. Cancer. 85:2017–2022. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Uchida T, Wang C, Sato T, et al:
BRCA1 gene mutation and loss of heterozygosity on chromosome
17q21 in primary prostate cancer. Int J Cancer. 84:19–23. 1999.
View Article : Google Scholar
|