Introduction
Renal cell carcinoma (RCC) is a complex disease
entity comprising different types of cancer occurring in the
kidney, each of which results from a different gene, with different
histological characteristics and a clinical course that responds
differently to therapy (1). Over
the past two decades, the incidence of RCC has increased due to an
increasing number of patients diagnosed with small kidney tumor
types (2). The overall lifetime
risk for RCC development is ~1 in 67, and it is higher in men
compared with women (3). Due to
its gradual onset and progression, RCC may be difficult to detect
and ~80% of renal tumor types are discovered incidentally during
unassociated abdominal imaging examinations (4). Among all RCCs, clear cell RCC is the
most frequently occurring type, accounting for >90% of all cases
(5). The risk factors for RCC may
vary, including smoking, obesity, hypertension, dietary habits and
living environment (6). Diverse
treatments for RCC have been reported, including the use of drugs,
virus-based gene therapies and ablative therapies, in addition to
preoperative tumor embolization (7-10).
However, novel therapies are required due to the complexity and
poor prognosis of RCC.
The lysyl oxidase-like 2 (LOXL2) protein is a type
of human paralogue of lysyl oxidase (LOX) that functions as an
amine oxidase and aids the formation of lysine-derived cross-links
in collagen and elastin (11). At
present, LOXL2 is considered to be responsible for a diverse range
of cellular functions, including cell adhesion, migration and
invasion, metastasis, epithelial-to-mesenchymal transition (EMT)
and malignant transformation (12). Downregulated LOXL2 expression is
reported to inhibit cell migration and invasion in RCC (13). The steroid receptor coactivator
(Src)/focal adhesion kinase (FAK) signaling pathway is known to be
involved in the migration and invasion of various tumor cell types
(14). A previous study reported
that the inhibition of Src may represent a promising option for the
treatment of RCC (15).
Accumulating evidence indicates a potential association between
LOXL2 and the Src/FAK signaling pathway, as tumor-secreted LOXL2 is
able to activate fibroblasts through FAK signaling, and LOXL2 is
capable of activating the FAK/protein kinase B (AKT)/mammalian
target of rapamycin signaling pathways (16,17).
However, few studies investigating LOXL2 and the Src-AKT signaling
pathway in RCC have been conducted to date. Therefore, the aim of
the present study was to investigate the effect of LOXL2 on the
invasion, migration and EMT of RCC cells through the Src/FAK
signaling pathway, in order to identify novel potential targets for
the treatment of RCC.
Materials and methods
Ethics statement
The study protocol was approved by the Ethics
Committee of Shanghai Jiao Tong University Affiliated Sixth
People's Hospital (Shanghai, China) and Shanghai Jiao Tong
University Affiliated Sixth People's Hospital South Campus
(Shanghai, China), and all the patients signed informed consent
forms. The present study obtained the approval of the Ethics Review
Committee of Shanghai Jiao Tong University Affiliated Sixth
People's Hospital and Shanghai Jiao Tong University Affiliated
Sixth People's Hospital South Campus (approval no. 201407004).
Study subjects
RCC tissues and adjacent normal tissues were
collected from 80 patients with RCC, including 46 men and 30 women,
who were hospitalized at the Shanghai Jiao Tong University
Affiliated Sixth People's Hospital and Shanghai Jiao Tong
University Affiliated Sixth People's Hospital South Campus between
September 2014 and March 2017. The age of the patients ranged from
14 to 61 years, with a mean of 50.99 years. Among the patients, 25
were aged <50 years and 55 were aged ≥55 years. According to the
pathological diagnosis and classification criteria for RCC
published by the World Health Organization, 38 cases were diagnosed
as Fuhrman grade I-II and 42 cases as III-IV (18), 42 cases were diagnosed as
Tumor-Node-Metastasis (TNM) stage pT1-pT2 and 38 cases as pT3-pT4
(19); in addition, 36 cases had a
tumor diameter of <4 cm and 44 had a tumor diameter of ≥4 cm.
None of the patients had been subjected to radiotherapy or
chemotherapy.
Hematoxylin and eosin (H&E)
staining
The tissue specimens were fixed in 10% neutral
formaldehyde for 16-18 h at room temperature, dehydrated through
graded ethanol (50, 70, 80, 90 and 100%), transparentized in xylene
at 35°C for 30 min and embedded in paraffin at 60°C for 30 min.
Subsequently, the specimens were sliced into serial sections at a
thickness of 5 µm and allowed to stand at 45°C. Next, the
sections were extracted, baked at 60°C for 1 h, deparaffinized by
washing with xylene and rehydrated. Then, H&E staining was
performed at a temperature of 35°C as follows: the sections were
stained by hematoxylin for 10 min, washed with water for 15 min,
differentiated with 1% hydrochloric acid-ethanol for 3 sec, washed
with water for 10 sec and stained with 0.5% eosin for 3 min
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China). Then, the sections were dehydrated with graded ethanol,
transparentized with xylene and sealed with neutral gum. The
pathological changes of the RCC tissues and adjacent normal tissues
were observed under an optical microscope (magnification, ×200)
(CX31-LV320; Olympus Corporation, Tokyo, Japan).
Immunohistochemistry
The tissues were embedded in paraffin and sliced
into 3-4-µm sections. The sections were deparaffinized and
hydrated at room temperature for 20 min. Subsequently, the
endogenous peroxidase activity was blocked in 3%
H2O2 at room temperature for 10 min while the
sections were processed for antigen retrieval in a microwave at a
power of 800 W at 90°C for 5 min. Next, the sections were blocked
with 10% goat serum and incubated at room temperature for 10 min.
Primary polyclonal rabbit anti-human LOXL2 antibody (1:100; cat no.
ab179810; Abcam, Cambridge, MA, USA) was added to the sections and
left overnight at 4°C. Secondary biotinylated goat anti-rabbit
immunoglobulin G (IgG) antibody (1:1,000; cat no. ab6789; Abcam)
was added to the sections for 30 min at 37°C. Next, hematoxylin
(cat no. C0105. Beyotime Institute of Biotechnology, Shanghai,
China) was used to counterstain the nuclei for 30 sec at 37°C,
followed by 3,3′-diaminobenzidine (cat no. P0202; Beyotime
Institute of Biotechnology) staining at 37°C for 10 min.
Subsequently, the sections were dehydrated with hydrochloric acid
ethanol to clear and sealed with gum. Images were captured under an
optical microscope (magnification, ×200) (Olympus Corporation), and
analyzed using Image-Pro Plus 6.0 software (Media Cybernetics,
Inc., Rockville, MD, USA). Brown-yellow cytoplasmic staining was
classified as a positive expression.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Frozen tissues (100 mg) were obtained and a
TRIzol® kit (cat no. 16096020; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) was employed to extract total RNA,
according to the manufacturer's protocol. The primers were designed
by the biological software packages Primer Premier 5 (Premier
Biosoft International, Palo Alto, CA, USA) and Oligo 6.0 software
(Molecular Biology Insights Inc., Cascade, CO, USA) and synthesized
by Takara Biotechnology Co., Ltd. (Dalian, China), and are
presented in Table I. RNA (10
µl) was extracted and diluted by RNase-free ultra-pure water
(×20). The optical density (OD) at 260 nm and 280 nm was recorded
with an ultraviolet/visible spectrophotometer, and the
concentration and purity of the total RNA were measured. Next, 5
µl Mix reagent (cat no. 4368702; Tideradar Beijing
Technology Co., Ltd., Beijing, China), 5 µl total RNA and 10
µl RNase-free H2O were added into an Eppendorf
tube, mixed by centrifugation (7,200 × g at 4°C for 1 min) and
placed in a qPCR instrument. The reaction conditions were as
follows: 37°C for 15 min and 85°C for 5 sec; and the reaction was
terminated at 4°C. The cDNA obtained by RT was stored at -20°C. The
ABI7500 qPCR instrument (ABI 7500; Applied Biosystems; Thermo
Fisher Scientific, Inc.) was used for RT-qPCR and the reaction
conditions were as follows: Pre-denaturation at 95°C for 10 min and
a total of 40 cycles of denaturation at 95°C for 10 sec, annealing
at 60°C for 20 sec and extension at 72°C for 34 sec. SYBR Green
fluorescence dye (Takara Biotechnology Co., Ltd.) was applied for
RT-qPCR, which was also suitable for cell detection at 48 h of
transfection. The expression levels were evaluation by
2−ΔΔCq method (20).
 | Table IPrimer sequences of associated genes
for reverse transcription-quantitative polymerase chain
reaction. |
Table I
Primer sequences of associated genes
for reverse transcription-quantitative polymerase chain
reaction.
| Gene | Sequence |
|---|
| LOXL2 | F:
5′-GGGTGGAGGTGTACTATGATGG-3′ |
| R:
5′-CTTGCCGTAGGAGGAGCTG-3′ |
| FAK | F:
5′-GCTTACCTTGACCCCAACTTG-3′ |
| R:
5′-ACGTTCCATACCAGTACCCAG-3′ |
| Src | F:
5′-GAGCGGCTCCAGATTGTCAA-3′ |
| R:
5′-CTGGGGATGTAGCCTGTCTGT-3′ |
| MMP-9 | F:
5′-TGTACCGCTATGGTTACACTCG-3′ |
| R:
5′-GGCAGGGACAGTTGCTTCT-3′ |
| E-cadherin | F:
5′-GGAAGTCAGTTCAGACTCCAGCC-3′ |
| R:
5′-AGGCCTTTTGACTGTAATCACACC-3′ |
| N-cadherin | F:
5′-AACCCTTATTTTGCCCCCAAT-3′ |
| R:
5′-AAGGTCATTAAACAACTTCTGAGC-3′ |
| Vimentin | F:
5′-GACGCCATCAACACCGAGTT-3′ |
| R:
5′-CTTTGTCGTTGGTTAGCTGGT-3′ |
| GAPDH | F:
5′-GGTGAAGGTCGGAGTCAACGG-3′ |
| R:
5′-CCTGGAAGATGGTGATGGGATT-3′ |
Western blot analysis
The RCC tissues and adjacent normal tissues were
added to liquid nitrogen and ground into a homogeneous fine powder.
Protein lysate was added (cat no. C0481; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) to the tissues at 4°C for 30 min and
centrifugation was conducted at 29,000 × g at 4°C for 20 min. Next,
the supernatant was collected and packaged separately for further
use. The bicinchoninic acid method was used to measure the protein
concentration of each sample and deionized water was used for
adjustment, in order to maintain consistency with the volume of the
loading sample. Subsequently, proteins were separated by SDS-PAGE
(10% gels). The sample was mixed with the loading buffer and boiled
at 100°C for 5 min, then the denatured protein was preserved at
-70°C. The protein sample (50 µg/lane) was subjected to
electrophoretic separation and the protein was transferred to a
nitrocellulose membrane and sealed using 5% skimmed milk powder at
4°C overnight. Diluted primary antibodies [rabbit anti-human LOXL2,
FAK, phosphorylated (p)-FAK, Src, p-Src, matrix metalloproteinase
(MMP)-9, epithelial (E)-cadherin, neuronal (N)-cadherin and
vimentin (1:1,000; cat nos. ab179810, ab40794, ab81298, ab47405,
ab40660, ab73734, ab1416, ab18203 and ab92547, respectively; Abcam)
and rabbit anti-human GAPDH (1:2,000; cat no. ab8226; Abcam) were
added to the samples, incubated at room temperature for 1 h and
washed with phosphate-buffered saline (PBS) three times, for 5 min
each time. Secondary goat anti-rabbit IgG antibody (1:1,000; cat
no. ab6721; Abcam) was added, incubated at room temperature for 1 h
and washed with PBS at room temperature three times, for 5 min each
time. Immunoreactive bands were visualized by enhanced
chemiluminescence (Pierce; Thermo Fisher Scientific, Inc.) at room
temperature for 1 min. Next, the liquid was removed and the
membrane was covered with a plastic wrap, photographed by X-ray and
observed. Densiometric analysis was performed using Image J
software (version 1.37; National Institutes of Health, Bethesda,
MD, USA). GAPDH was used as the internal reference. The ratio of
the gray value of the target band to that of the internal reference
band was used as the relative protein expression.
Adenovirus expression vector construction
and screening
The RNA interference technique was applied and the
specific target sequences of small interfering RNA (siRNA) of LOXL2
were designed according to the mRNA sequence of nucleotide LOXL2
from the National Center for Biotechnology Information (NCBI;
https://www.ncbi.nlm.nih.gov/).
Adenovirus expression vectors targeting LOXL2 were constructed,
including vectors expressing siRNA-LOXL2-1, siRNA-LOXL2-2 and
siRNA-LOXL2-3 (Table II), LOXL2
overexpression adenovirus vector Ad-CMV-LOXL2-eGFP and blank
adenovirus vector Ad-CMV-eGFP (Beijing Shengke Boyuan Biotechnology
Co., Ltd., Beijing, China). The vector with the highest silencing
efficiency was screened for subsequent cell experiments. The
plasmid pLenR-GPH vector (Engreen Biosystem New Zealand Ltd.,
Auckland, New Zealand) was used to construct the siRNA adenovirus
expression vector siRNA-LOXL2, the LOXL2 overexpression adenovirus
vector Ad-CMV-LOXL2-eGFP and the blank adenovirus vector
Ad-CMV-eGFP. Double-stranded DNA oligo containing interference
sequence was synthesized and directly linked to the vectors
digested by the restriction endonucleases NheI and
KpnI. The ligation product was transformed into E.
coli DH5α cells with the purposes of amplifying plasmid, and
then plasmid was extracted and identified through restriction
endonucleases NheI and KpnI digestion.
| Table IISilencing sequences. |
Table II
Silencing sequences.
| Expression
vector | Target
sequences |
|---|
| siRNA-LOXL2-1 |
CCTDTTCCAGGTTGTTATT |
| siRNA-LOXL2-2 |
CCGATTACTCCAACAACAT |
| siRNA-LOXL2-3 |
CCAGATAGAGAACCTGAAT |
| siRNA-NC |
TTTATAGAGGTTGTACTCC |
Cell grouping and transfection
HK-2 (normal renal tubular epithelial cells; cat no.
CRL-2190; American Type Culture Collection, Manassas, VA, USA) and
the RCC cell lines 786-0, ACHN, Caki1 and A498 (Cell Resource
Centre, Institute of Basic Medical Sciences, Peking Union Medical
College, Beijing, China) were cultured in RPMI-1640 culture medium
(cat no. 22400089; Gibco; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.). The cells were seeded into a 6-well plate
(1×105/well) at 37°C in a humidified atmosphere
containing 5% CO2. The culture medium was changed every
2-3 days. The cells were subcultured until they reached 80-90%
confluence. The culture medium was then removed and the cells were
washed with PBS twice, digested with 0.25% trypsin for 2-5 min at
37°C, resuspended and subcultured in 5 ml RPMI-1640 containing 10%
FBS.
Cells in the logarithmic growth phase were extracted
and assigned into the following groups: i) 786-O cell line, blank
group (no transfection), siRNA negative control (si-NC) group
(cells transfected with si-NC) and si-LOXL2 group (cells
transfected with si-LOXL2); ii) Caki1 cell line, blank group (no
transfection), LOXL2 empty vector group (cells transfected with the
empty adenovirus vector Ad-CMV-eGFP), LOXL2 vector group (cells
transfected with Ad-CMV-LOXL2-eGFP), PP2 group [cells transfected
with 20 µmol/l of the signaling pathway inhibitor PP2
(Selleck Chemicals, Houston, TX, USA)] and the LOXL2 vector + PP2
group (cells transfected with Ad-CMV-LOXL2-eGFP and 20
µmol/l PP2). Prior to transfection, the cells were passaged
and seeded into a 6-well plate (1×105/well). The cell
confluence reached 70-80% on the day of transfection. The cells
were transfected using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. Subsequently, 250 µl serum-free Opti-MEM (Gibco;
Thermo Fisher Scientific, Inc.) was used to dilute 100 pmol blank
adenovirus vector Ad-CMV-eGFP, siRNA-LOXL2 and Ad-CMV-LOXL2-eGFP
solutions (final concentration, 50 nM), which were gently mixed and
incubated at room temperature for 5 min. Next, 250 µl
serum-free Opti-MEM was used to dilute 5 µl Lipofectamine
2000 and the two solutions were mixed and incubated at room
temperature for 5 min. Subsequent to combining the two mixtures,
the solution was incubated at room temperature for 20 min and added
into the wells of a cell culture plate. The transfected cells were
then cultured in an incubator at 37°C in a humidified atmosphere
containing 5% CO2. After 6-8 h of culture, cells were
resuspended in complete medium. The eGFP protein expression in the
cells was observed under a fluorescence microscope (magnification,
×100) (M30C; Shanghai Wan Heng Precision Instrument Co., Ltd.,
Shanghai, China) after a 24-h culture; two random fields were
selected for image capture and the mean value was calculated. The
transfection efficiency was equal to the ratio of the number of
transfected cells to that of total cells. After 24-48-h culturing,
the cells were used for further experiments.
MTT assay
Following transfection for 24 h, RCC cells in the
logarithmic growth phase were collected and RPMI-1640 containing
10% FBS was used to prepare the cell suspension
(2.5×105/ml). The cells were seeded into a 96-well
plate, and 8-wells were set for each group (100 µl/well);
the plate was then incubated at 37°C in a humidified atmosphere
containing 5% CO2 for 24, 48 and 72 h. Next, 10
µl MTT solution (5 mg/ml; Sigma-Aldrich; Merck KGaA) was
added to each well for another 4-h culture. Once the culture was
terminated, the supernatant was discarded and 150 µl
dimethyl sulfoxide solution was added to the plate and vibrated for
10 min to dissolve crystals at room temperature. A microplate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA) was used to
measure the optical density (OD) value of each well at 490 nm.
Three parallel wells were set for each group and the mean value was
calculated. The experiment was repeated three times. The curve of
cell proliferation was drawn with time as the abscissa and the OD
value as the ordinate.
Transwell assay
The RCC cells were dissolved with Matrigel (cat no.
356234; BD Biosciences, San Jose, CA, USA) overnight, and diluted
with serum-free DMEM (Gibco; Thermo Fisher Scientific, Inc.) at
dilution ratio of 1:3. Next, 50 µl diluted Matrigel was
added into the upper Transwell chamber and allowed to stand in an
incubator for 30 min at 37°C. The upper chamber (with serum-free
medium) was inoculated with a ~1×105/ml cell suspension
and the DMEM medium containing 10% FBS was added to the basolateral
Transwell chamber. After a 24-h culture at 37°C, the chambers were
washed twice with PBS. Cells were removed from the top of the
membranes with a cotton swab. Invading cells were fixed with 5%
glutaraldehyde at 4°C for 30 min and stained with 0.1% crystal
violet (Sigma-Aldrich; Merck KGaA) at 37°C for 10 min. The number
of invading cells was counted and used as the indicator to evaluate
cell invasion and migration ability. Images were captured under an
optical microscope (magnification, ×200; Olympus Corporation). The
experiment was repeated three times.
Wound healing assay
The RCC cells were seeded in a 6-well plate.
Serum-free DMEM was used following cell attachment. When the cell
confluence reached 90-100%, a pipette tip (10 µl) was used
to scratch the 6-well plate perpendicular to the bottom of the
plate, with 4-5 lines per well, ensuring that the width of all
scratches was uniform. Once the cells were rinsed using PBS three
times, the scratched cells were removed and cultured in an
incubator. Migration distance at the scratch region was determined
under an optical microscope (magnification, ×100) at 0 and 24 h,
and six horizontal lines were randomly drawn and then photographed.
The scratch gap confluence rate was calculated as follows: Gap
confluence rate = (gap width at 0 h - gap width at 24 h)/gap width
at 0 h. Three duplicated wells were set in each well and the
experiment was repeated three times.
Flow cytometry
The culture medium was discarded 48 h after
transfection. A total of 1×105/ml RCC cells were then
washed once with PBS balanced salt solution and digested using
0.25% trypsin. When the cells acquired a round shape, trypsin was
removed and the DMEM medium containing 10% FBS was added to the
cells to terminate digestion. Next, the cells were mixed to prepare
a mixed cell suspension, which was centrifuged at 200 × g for 5 min
at 4°C, and then the supernatant was discarded. PBS balanced salt
solution was used to wash the cells twice and precooled 70% ethanol
was used to fix the cells for 30 min at 4°C, following which the
cells were collected by centrifugation at 200 × g for 5 min at 4°C.
Subsequently, PBS solution was added to wash the cells twice, and 1
ml propidium iodide (PI) with RNAse was used for staining for 30
min at 37°C. The cells were washed with PBS balanced salt solution
twice and PI was removed. The volume of the mixture solution was
adjusted to 1 ml with the addition of PBS solution. The samples
were transferred to a flow cytometry instrument (BD-Aria; FACS
Calibur; Beckman Coulter, Miami, FL, USA) in order to record cell
cycle at the activating wavelength of 488 nm detected by red
fluorescence. Three samples were set in each group and the
experiment was repeated three times.
At 48 h after transfection, trypsin without
ethylenediaminetetraacetic acid was applied to digest the cells for
5 min at 37°C, and the cells were then collected into a flow tube.
Next, centrifugation was performed and the supernatant was removed.
The cells were washed with cold PBS three times, followed by
centrifugation at 200 × g for 5 min at 4°C and supernatant removal.
Subsequently, Annexin V-fluorescein isothiocyanate (FITC) dye
solution was prepared by Annexin V-FITC, PI and HEPES buffer
solution at 1:2:50 using the Annexin-V-FITC apoptosis detection kit
(cat no. C1065; Beyotime Institute of Biotechnology) according to
the manufacturer's protocol. A total of 100 µl dye solution
was added to 1×106 cells for resuspension. Following
oscillation and mixing, the cells were incubated at room
temperature for 15 min, followed by the addition of 1 ml HEPES
buffer solution and oscillation. The samples were transferred on
flow cytometry instrument (BD-Aria), then, bandpass filters of 525
and 620 nm were activated by 488 nm wavelength to detect FITC and
PI fluorescence, in addition to cell apoptosis. Three samples were
set for each group and the experiment was repeated three times.
Statistical analysis
All data were processed using SPSS 21.0 statistical
software (IBM Corp., Armonk, NY, USA). The measurement data were
subject to normal distribution, which was expressed as the mean ±
standard deviation. The comparison of RCC and normal adjacent
tissues was performed using a paired Student's t-test; the
comparison between two groups was performed by an independent
samples t-test and that among multiple groups by one-way analysis
of variance. Tukey's post hoc test was used following analysis of
variance. Numerical data were expressed as percentage or rate and
analyzed using a χ2 test. P<0.05 was considered to
indicate statistically significant differences.
Results
Histopathological examination of RCC
tissues and adjacent normal tissues
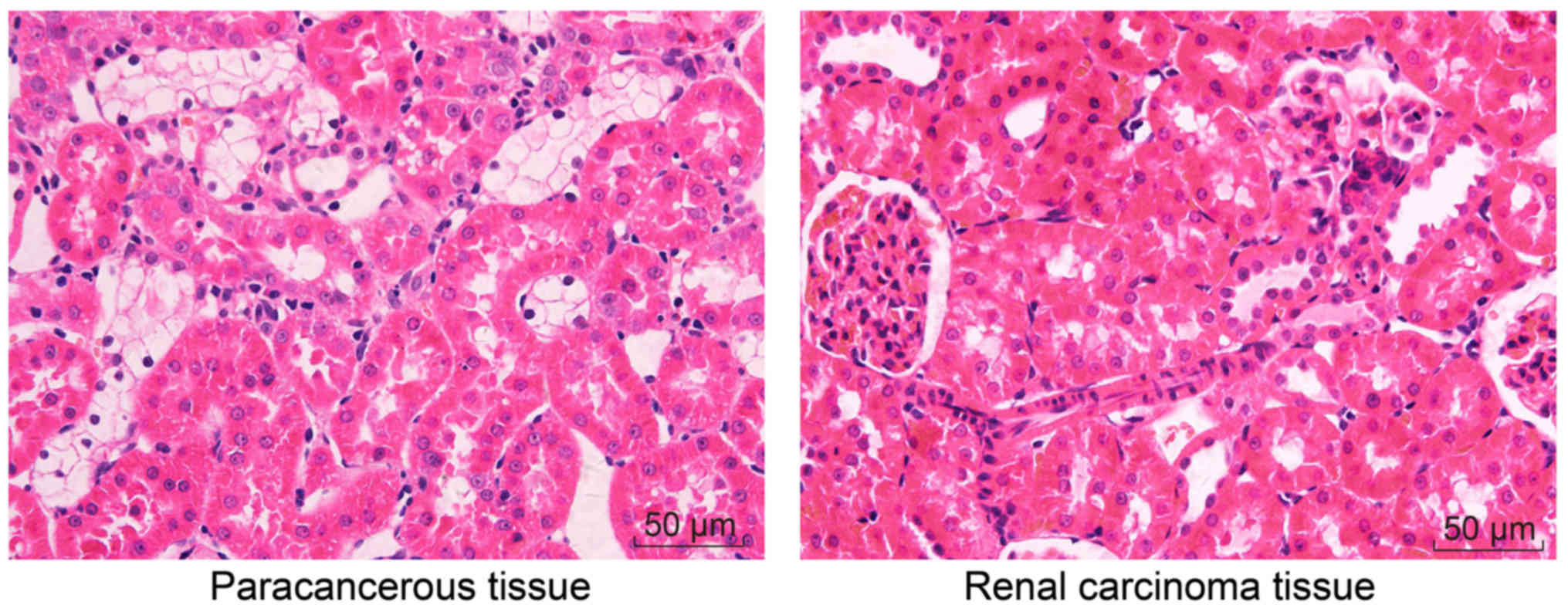
H&E staining was performed to observe
histopathological changes. In the adjacent normal tissues, the
cells exhibited normal morphology and uniform arrangement, without
inflammatory infiltration. In the RCC tissues, however, the cells
exhibited dedifferentiated morphology and disorderly arrangement;
notable tumor cell infiltration was detected, around which normal
kidney tissues were observed (Fig.
1). In conclusion, the cells exhibited dedifferentiated
morphology and were arranged in a disorderly manner in RCC tissues
compared with adjacent normal tissues.
LOXL2 protein expression is high in RCC
tissues
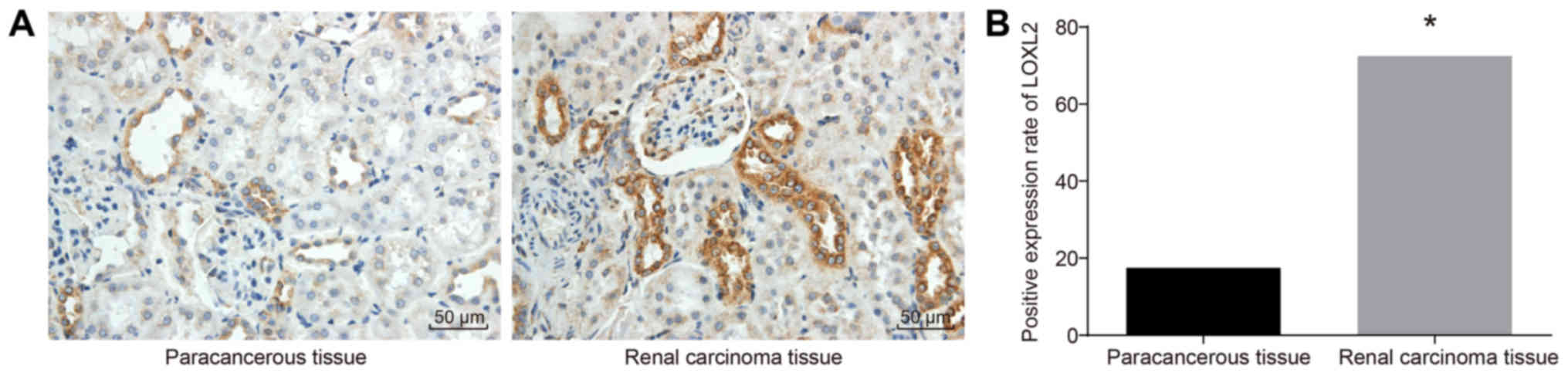
In order to measure the cell protein expression of
LOXL2, immunohistochemistry was performed. The results (Fig. 2) revealed that LOXL2-positive cells
were located in the cytoplasm, presenting as brown-yellow granules.
The protein expression rate of LOXL2 in the RCC tissues was 72.5%
(58/80), which was significantly higher compared with 17.5% (14/80)
in adjacent normal tissues (P<0.05). Taken together, these
results demonstrated that LOXL2 protein expression was higher in
RCC tissues compared with that in adjacent normal tissues.
Associations between L OX L 2 expression
and clinicopathological characteristics of RCC
The associations between LOXL2 expression and the
clinicopathological characteristics of RCC is presented in Table III. LOXL2 expression was
independent of the sex and age of patients with RCC, but was
significantly associated with stage [Fuhrman (P=0.001) and TNM
(P=0.009)], histological type (P=0.001), diameter (P=0.001) and
venous invasion (P=0.002). The expression of LOXL2 was
significantly elevated in patients with Fuhrman grade III + IV and
those with clear cell RCC with a tumor diameter ≥4 cm (all
P<0.05). Therefore, LOXL2 expression was significantly
associated with tumor grade, type, diameter and venous invasion in
RCC.
| Table IIIAssociation between LOXL2 expression
and clinicopathological characteristics of renal cell
carcinoma. |
Table III
Association between LOXL2 expression
and clinicopathological characteristics of renal cell
carcinoma.
|
Characteristics | Case number
(n) | LOXL2 expression
| P-value |
|---|
| Positive | Negative |
|---|
| Age (years) | | | | 0.567 |
| <50 | 25 | 11 | 14 | |
| ≥50 | 55 | 28 | 27 | |
| Sex | | | | 0.476 |
| Male | 46 | 24 | 22 | |
| Female | 34 | 15 | 19 | |
| Fuhrman grade | | | | 0.001 |
| I+II | 38 | 11 | 27 | |
| III+IV | 42 | 28 | 14 | |
|
Tumor-Node-Metastasis stage | | | | 0.009 |
| pT1+pT2 | 42 | 13 | 29 | |
| pT3+pT4 | 38 | 26 | 18 | |
| Histological
type | | | | 0.001 |
| Clear cell | 43 | 13 | 30 | |
| Papillary | 36 | 25 | 11 | |
| Tumor diameter | | | | 0.001 |
| <4 cm | 36 | 10 | 26 | |
| ≥4 cm | 44 | 29 | 15 | |
| Venous
invasion | | | | 0.002 |
| Yes | 45 | 29 | 16 | |
| No | 35 | 10 | 25 | |
LOXL2 and FAK/Src signaling pathway genes
are highly expressed and EMT-associated gene expression is affected
in RCC tissues
Subsequently, RT-qPCR and western blot analysis were
performed. The results (Fig. 3)
demonstrated that, compared with the adjacent normal tissues, the
mRNA expression levels of LOXL2, FAK, Src, MMP-9, N-cadherin and
vimentin were significantly increased and the mRNA expression of
E-cadherin was significantly decreased in RCC tissues (all
P<0.05), whereas the protein expression levels of LOXL2,
p-FAK/FAK, p-Src/Src, MMP-9, N-cadherin and vimentin were
significantly increased and the protein expression of E-cadherin
significantly decreased in RCC tissues (all P<0.05). Taken
together, these data indicate that genes involved in LOXL2 and
FAK/Src signaling pathways are highly expressed in RCC, and
EMT-associated gene expression may be affected in RCC tissues.
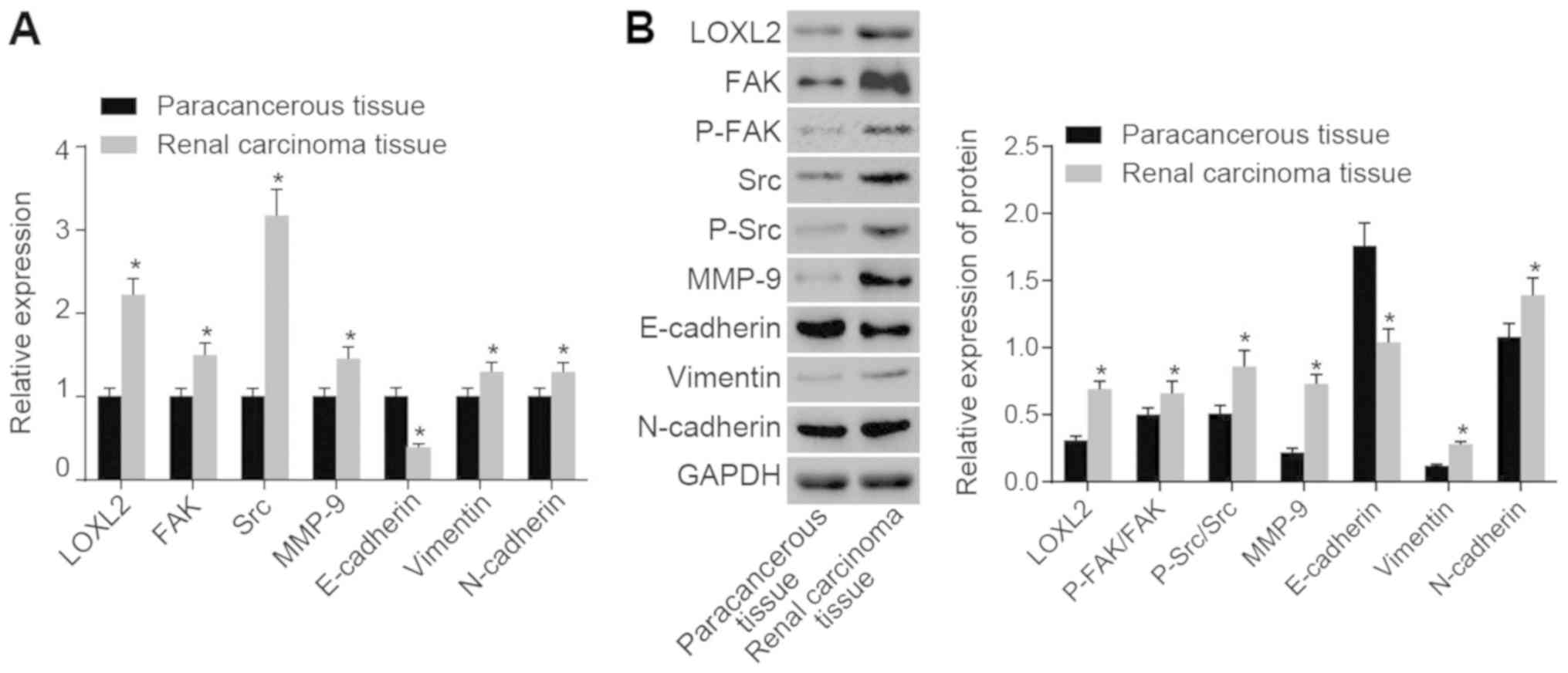 | Figure 3According to reverse
transcription-quantitative polymerase chain reaction and western
blot analysis, LOXL2 and FAK/Src signaling pathway-associated genes
are highly expressed in RCC tissues, and genes associated with the
epithelial-to-mesenchymal transition are affected. (A) mRNA
expression of LOXL2, FAK, Src, MMP-9, N-cadherin and vimentin was
increased in RCC tissues, whereas that of E-cadherin was decreased
compared with adjacent normal tissues. (B) Protein expression of
LOXL2, FAK, Src, MMP-9, N-cadherin and vimentin were increased in
RCC tissues, whereas that of E-cadherin was decreased, in contrast
to the adjacent normal tissues; n=80. (C) Quantified protein
expression of LOXL2, FAK, P-FAK, Src, P-Src, MMP-9, E-cadherin,
vimentin and N-cadherin. Paired t-test was used for analysis;
*P<0.05 vs. adjacent normal tissues. LOXL2, lysyl
oxidase-like 2; Src, steroid receptor coactivator; FAK, focal
adhesion kinase; RCC, renal cell carcinoma; MMP, matrix
metalloproteinase; P-, phosphorylated; N-cadherin, neuronal
cadherin; E-cadherin, epithelial cadherin. |
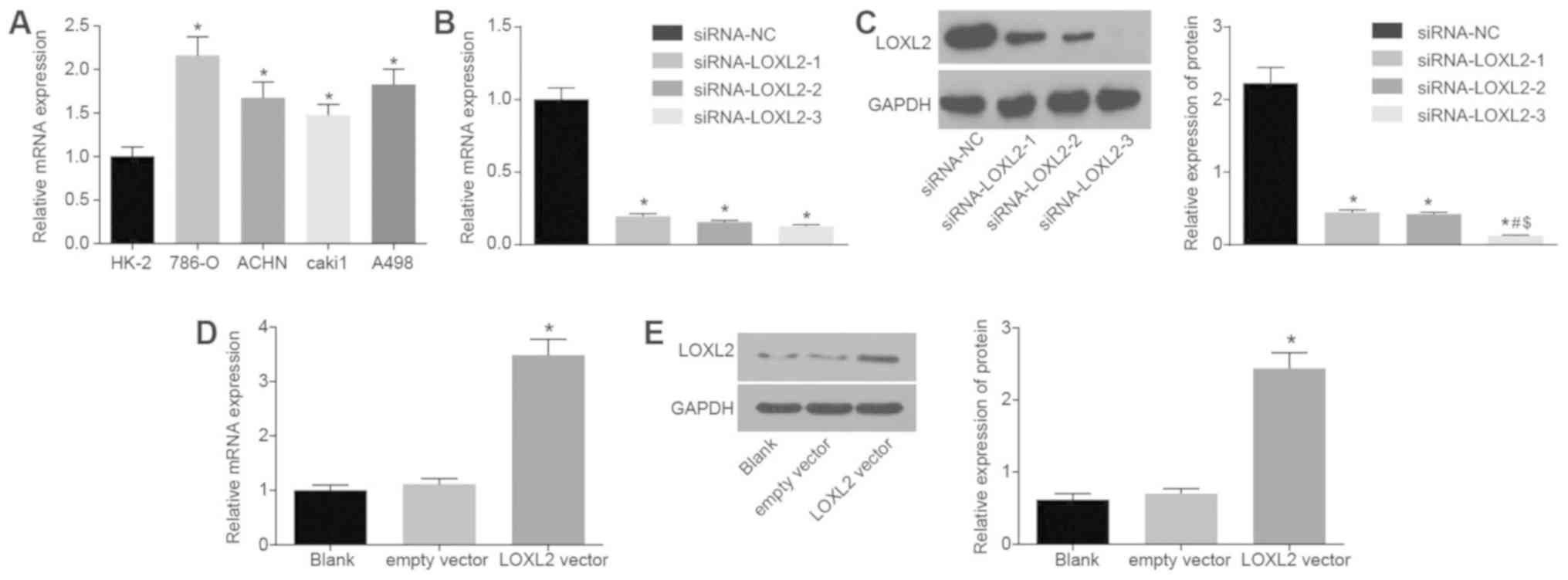
LOXL2 downregulation in 786-0 and Cak1
cells
In order to study the expression of LOXL2 in normal
renal tubular epithelial cells (HK-2) and RCC cell lines, LOXL2
mRNA expression in HK-2, 786-O, ACHN, Caki1 and A498 cells was
detected by RT-qPCR. Compared with HK-2 cells, the expression of
LOXL2 mRNA in RCC cell lines was significantly increased
(P<0.05; with the highest expression in 786-O cells and the
lowest expression in Caki1 cells, compared with all other RCC cell
lines; Fig. 4A). Thus, 786-O cells
were selected for siRNA transfection experiments and Caki1 cells
for overexpression experiments. The efficiency detection results of
each group following 786-O cell transfection with interference
plasmids are presented in Fig. 4B.
Compared with the siRNA-NC group, the siRNA-LOXL2-1, siRNA-LOXL2-2
and siRNA-LOXL2-3 groups exhibited significantly decreased LOXL2
mRNA expression (P<0.05; Fig.
4B) and protein expression (P<0.05; Fig. 4C). The lowest protein expression of
LOXL2 was observed in the siRNA-LOXL2-3 group, with a significantly
lower expression of LOXL2 compared with the other two siRNA groups
(P<0.05), indicating that the siRNA-LOXL2-3 group had the
highest interference efficiency. Therefore, the plasmid in the
siRNA-LOXL2-3 group was selected for further experimentation.
Following Caki1 cell transfection with LOXL2 overexpression
plasmids, the mRNA and protein expression of LOXL2 increased
significantly compared with the blank control (P<0.05; Fig. 4D and E). Therefore, 786-O cells
were selected for siRNA experiments and Caki1 cells for
overexpression experiments.
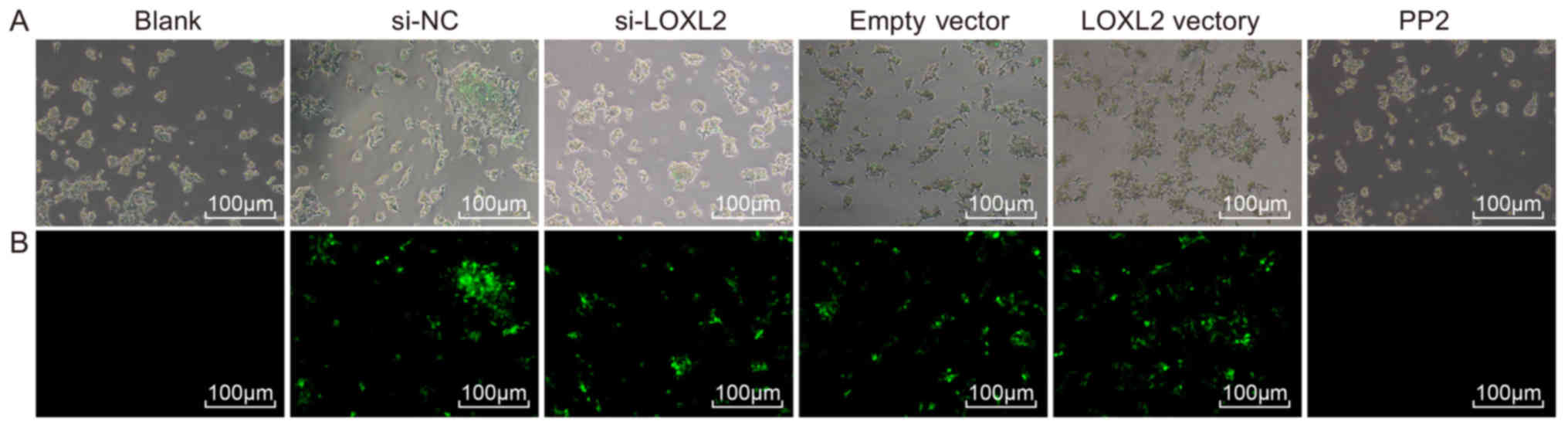
Transfection efficiency
Cell transfection was performed to measure green
fluorescence in cells. Following cell transfection, no green
fluorescence was observed in the normal, blank and PP2 groups under
a fluorescence microscope. However, intense green fluorescence was
observed in the cells of the si-NC, si-LOXL2, empty vector and
LOXL2 vector groups (expression rate, >80%), indicating that the
adenovirus vector was successfully transfected into the RCC cells
and kidney tubular epithelial cells and effectively expressed in
these cells (Fig. 5).
Silencing LOXL2 inhibits the activation
of the FAK/Src pathway and EMT in RCC cells
Next, RT-qPCR and western blot analysis were used to
determine mRNA and protein expression of associated genes and
proteins, respectively. As presented in Fig. 6, the results demonstrated that
following the transfection of 786-O cells, the mRNA and protein
expression of LOXL2, FAK, Src, MMP-9, N-cadherin and vimentin were
significantly reduced and that of E-cadherin was significantly
increased in the si-LOXL2 group compared with the blank and si-NC
groups (P<0.05). Following the transfection of Caki1 cells, the
mRNA and protein expression of LOXL2, FAK, Src, MMP-9, N-cadherin
and vimentin were significantly increased and that of E-cadherin
was significantly decreased in the LOXL2 vector group compared with
the blank and empty vector groups (all P<0.05; Fig. 6). In the PP2 group, gene expression
changes exhibited the opposite trend compared with the LOXL2 vector
group, except for LOXL2 which demonstrated a similar trend. In
comparison with the cells in the LOXL2 vector group, the mRNA and
protein expressions of p-FAK/FAK, p-Src/Src, MMP-9, N-cadherin and
vimentin were significantly decreased, and that of E-cadherin was
significantly increased in the LOXL2 vector + PP2 group (all
P<0.05; Fig. 6). The
aforementioned data indicated that silencing LOXL2 inhibits the
activation of the FAK/Src signaling pathway and of MMP-9 and EMT.
Additionally, PP2, as a FAK/Src signaling pathway inhibitor,
reversed the effect of LOXL2 vector on Caki1 cells. Consequently,
activation of the FAK/Src pathway and EMT may be inhibited by
silencing LOXL2 in RCC cells.
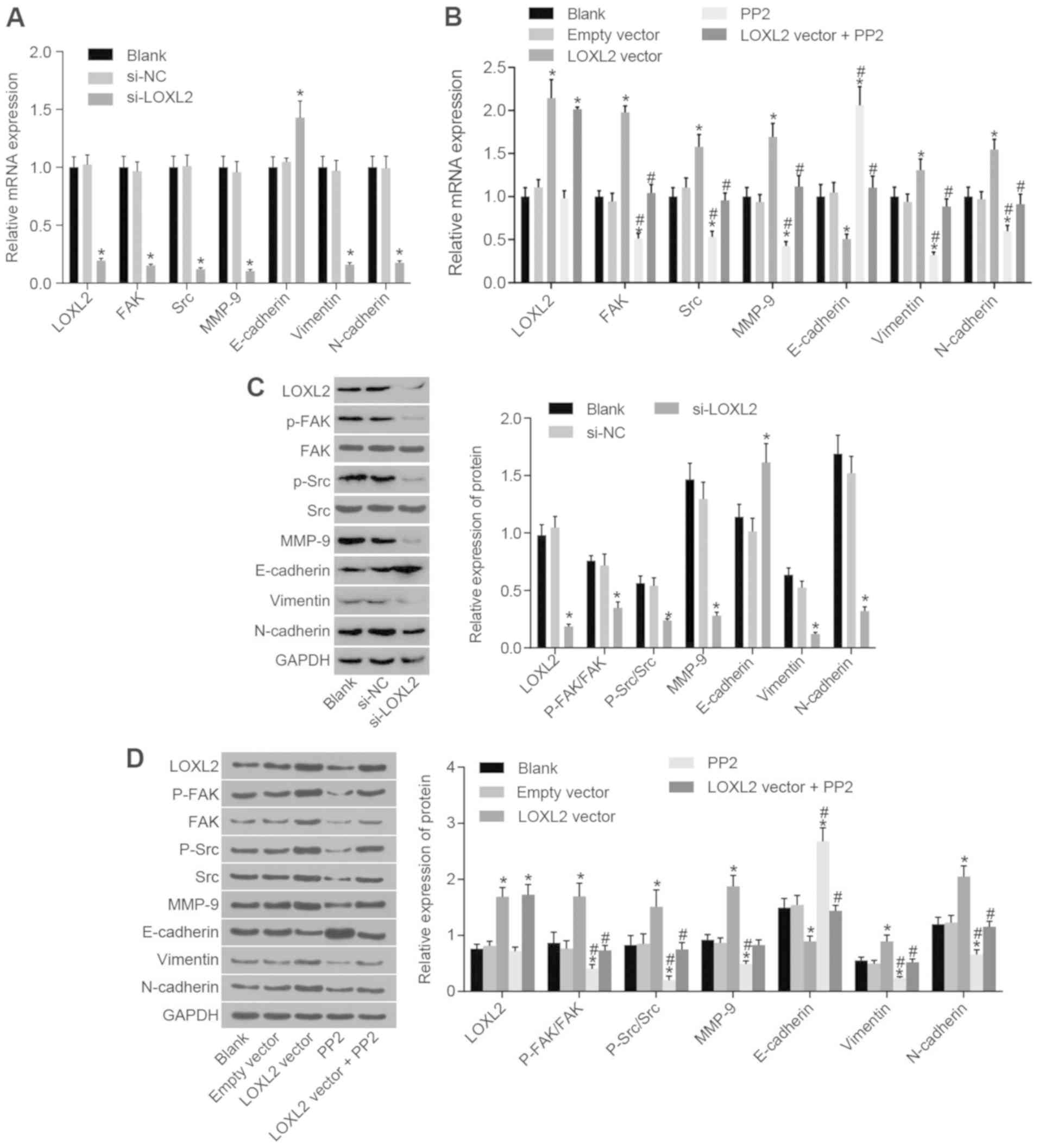 | Figure 6Silencing of LOXL2 inhibited
activation of the FAK/Src pathway and affected factors associated
with epithelial-to-mesenchymal transition. (A) Once 786-O cells
were transfected with si-LOXL2, the mRNA expression of FAK, Src,
MMP-9, N-cadherin and vimentin were significantly decreased and
that of E-cadherin was significantly increased;
*P<0.05 vs. blank and si-NC groups. (B) Once Caki1
cells were transfected with the LOXL2 vector, the mRNA expression
of FAK, Src, MMP-9, N-cadherin and vimentin were significantly
increased and that of E-cadherin was significantly decreased. In
the PP2 group, mRNA expression changes exhibited the opposite
trend. *P<0.05 vs. the blank and empty vector groups;
#P<0.05 vs. the LOXL2 vector group. (C) Once 786-O
cells were transfected with si-LOXL2, the protein expression of
FAK, Src, MMP-9, N-cadherin and vimentin was significantly
decreased and that of E-cadherin was significantly increased.
*P<0.05 compared with the blank and si-NC groups. (D)
Once Caki1 cells were transfected with LOXL2 vector, the protein
expressions of FAK, Src, MMP-9, N-cadherin and vimentin were
significantly increased and that of E-cadherin was significantly
decreased. In the PP2 group, protein expression changes exhibited
the opposite trend. *P<0.05 vs. the blank and empty
vector groups; #P<0.05 vs. the LOXL2 vector group.
One-way analysis of variance was used for comparison and the
experiment was repeated three times. LOXL2, lysyl oxidase-like 2;
Src, steroid receptor coactivator; FAK, focal adhesion kinase; MMP,
matrix metalloproteinase; NC, negative control; P-, phosphorylated;
si-, small interfering RNA; N-cadherin, neuronal cadherin;
E-cadherin, epithelial cadherin. |
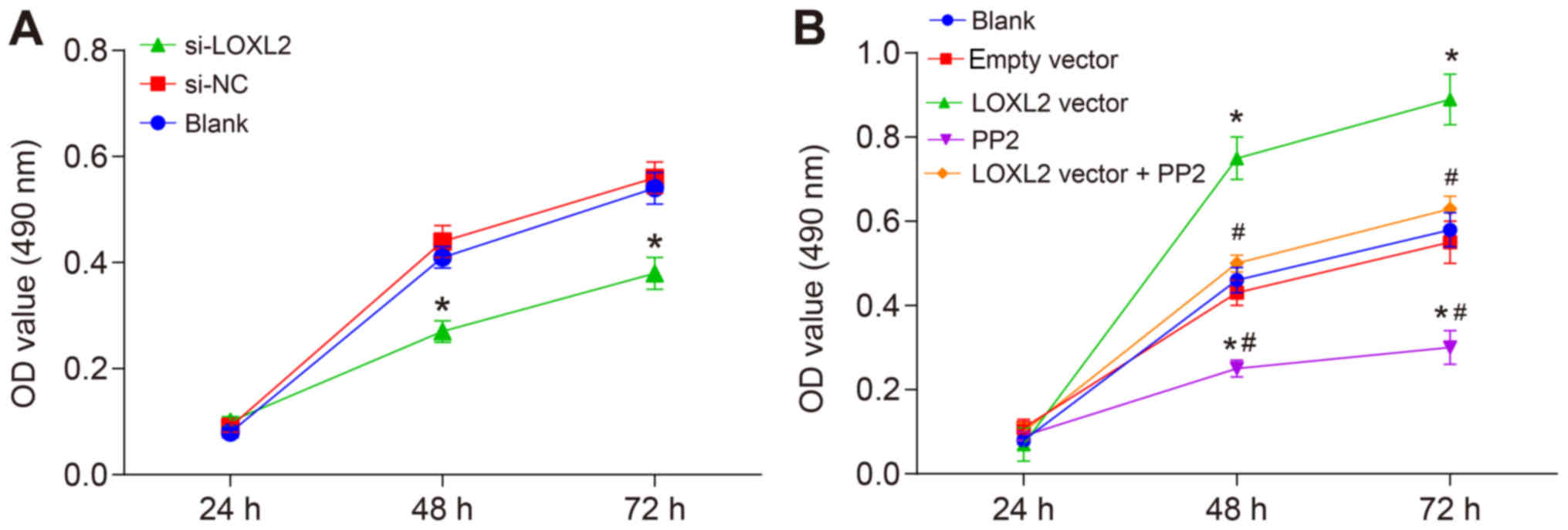
Silencing of LOXL2 inhibits the
proliferation of RCC cells
Cell proliferation was assessed with an MTT assay
and the results revealed differences in cell proliferation at 24,
48 and 72 h in each group. Compared with the cells in the blank and
si-NC groups, the cell proliferation was significantly inhibited at
all time points once 786-O cells were transfected with si-LOXL2
(P<0.05; Fig. 7A). This
indicates that the cell proliferation ability was repressed when
LOXL2 expression was inhibited. In comparison with the cells in the
blank and empty vector groups, once Caki1 cells were transfected
with the LOXL2 vector, cell proliferation was significantly
increased, while in the PP2 group it was significantly decreased
(P<0.05; Fig. 7B).
Additionally, the cell proliferation ability in the LOXL2 vector +
PP2 group was significantly decreased compared with cells in the
LOXL2 vector group (P<0.05; Fig.
7B). These data indicate that cell proliferation decreases when
LOXL2 expression and the activation of the FAK/Src pathway are
inhibited, and PP2 may reverse the effects of LOXL2 on the
proliferation of RCC cells. In brief, silencing LOXL2 represses RCC
cell proliferation.
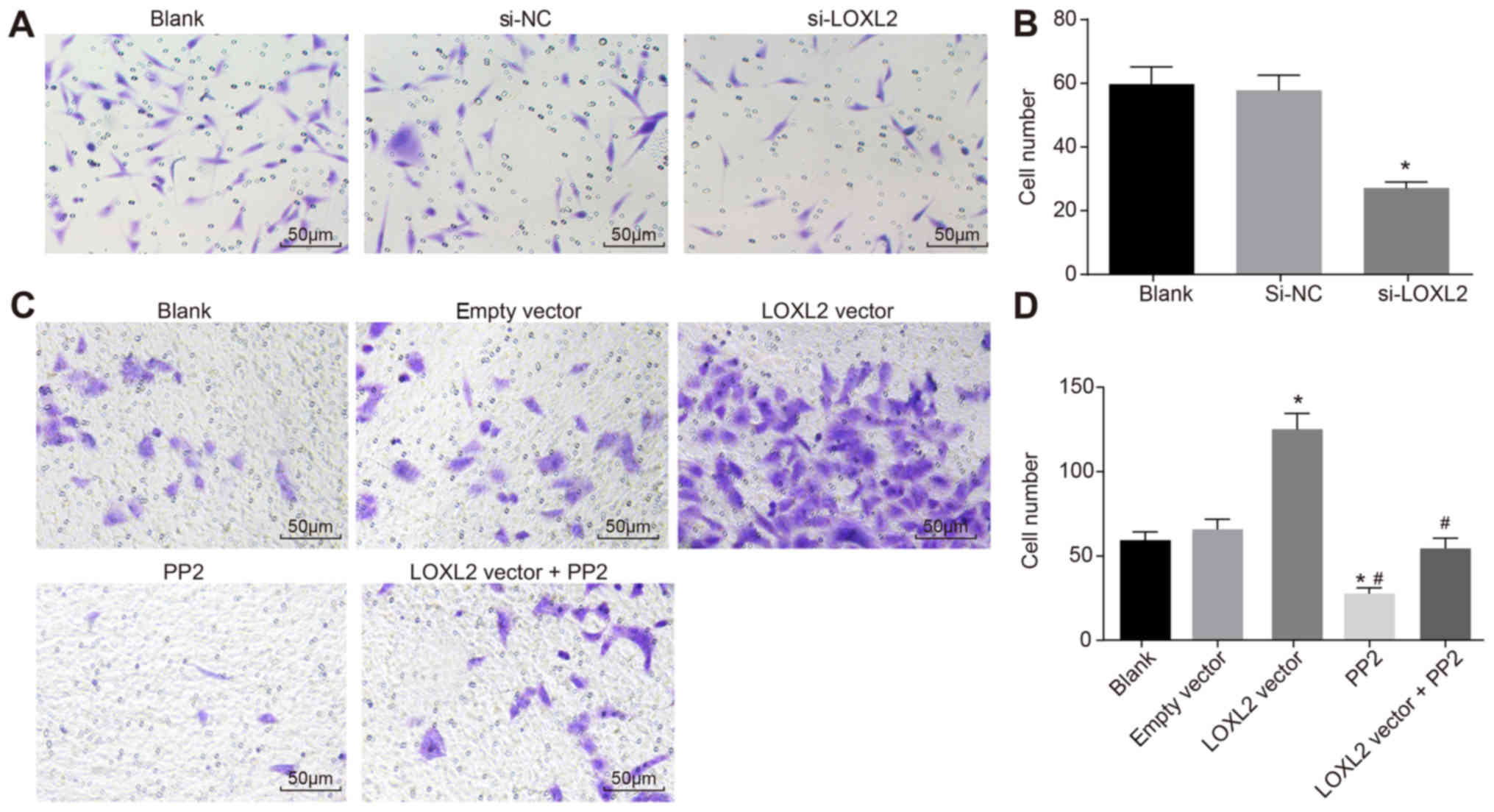
Silencing LOXL2 inhibits the invasion of
RCC cells
A Transwell assay was used to analyze the invasion
of cells in each group. Compared with cells in the blank group, the
number of invading cells was decreased significantly once 786-O
cells were transfected with si-LOXL2 (P<0.05; Fig. 8A and B). The results demonstrated
that silencing LOXL2 inhibited the invasion of RCC cells. Compared
with cells in the blank and empty vector groups, the number of
invading cells was significantly increased once Caki1 cells were
transfected with LOXL2 vector (P<0.05), whereas that in the PP2
group was significantly decreased (P<0.05). In comparison with
cells in the LOXL2 vector group, the number of invading cells in
the LOXL2 vector + PP2 group was decreased significantly
(P<0.05; Fig. 8C and D). The
results revealed that cell invasion ability decreases when LOXL2
expression and the activation of the FAK/Src pathway are inhibited.
In addition, PP2 is able to reverse the effects of LOXL2 on the
invasion ability of RCC cells. In conclusion, LOXL2 silencing
inhibits RCC cell invasion.
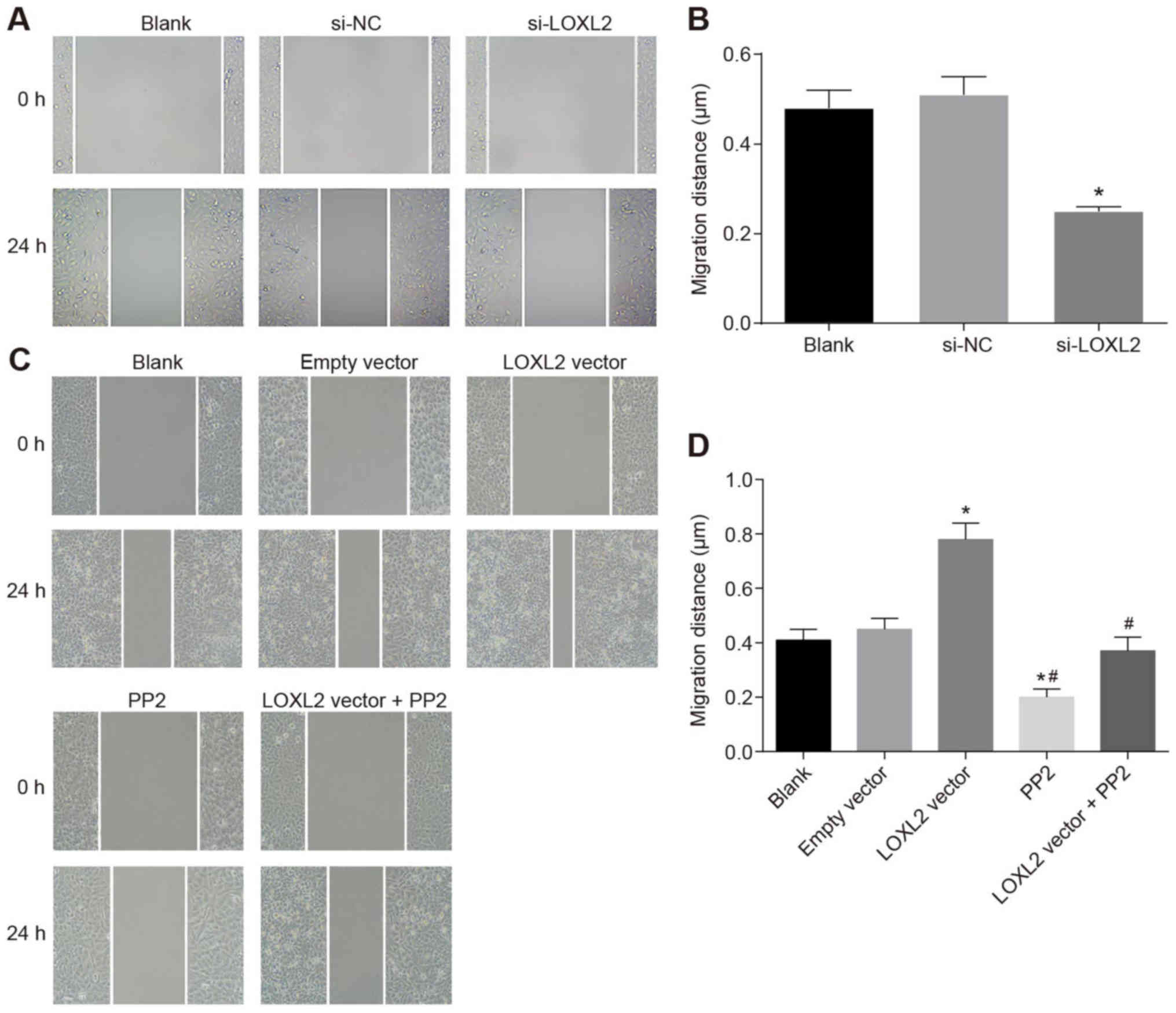
Silencing of LOXL2 inhibits the migration
of RCC cells
A wound healing test was applied to detect cell
migration. The results are presented in Fig. 9A and B. Once the 786-O cells were
transfected with si-LOXL2, the cell migration ability was
significantly weakened (P<0.05). Therefore, silencing LOXL2 may
inhibit the migration of RCC cells. By contrast, once Caki1 cells
were transfected with LOXL2 vector, cell migration ability was
enhanced significantly compared with cells in the blank and empty
vector groups (P<0.05), whereas the cell migration ability in
the PP2 and PP2 + LXOL2 vector groups was significantly reduced
(both P<0.05). Compared with cells in the LOXL2 vector group,
the cell migration ability in the LOXL2 vector + PP2 group was
significantly decreased (P<0.05; Fig. 9C and D). The results demonstrated
that inhibition of LOXL2 expression and inactivation of the FAK/Src
pathway inhibited the migration of RCC cells, and PP2 was able to
reverse the effects of LOXL2 on cell migration. In conclusion, the
experimental data indicate that the migration of RCC cells may be
suppressed by LOXL2 silencing.
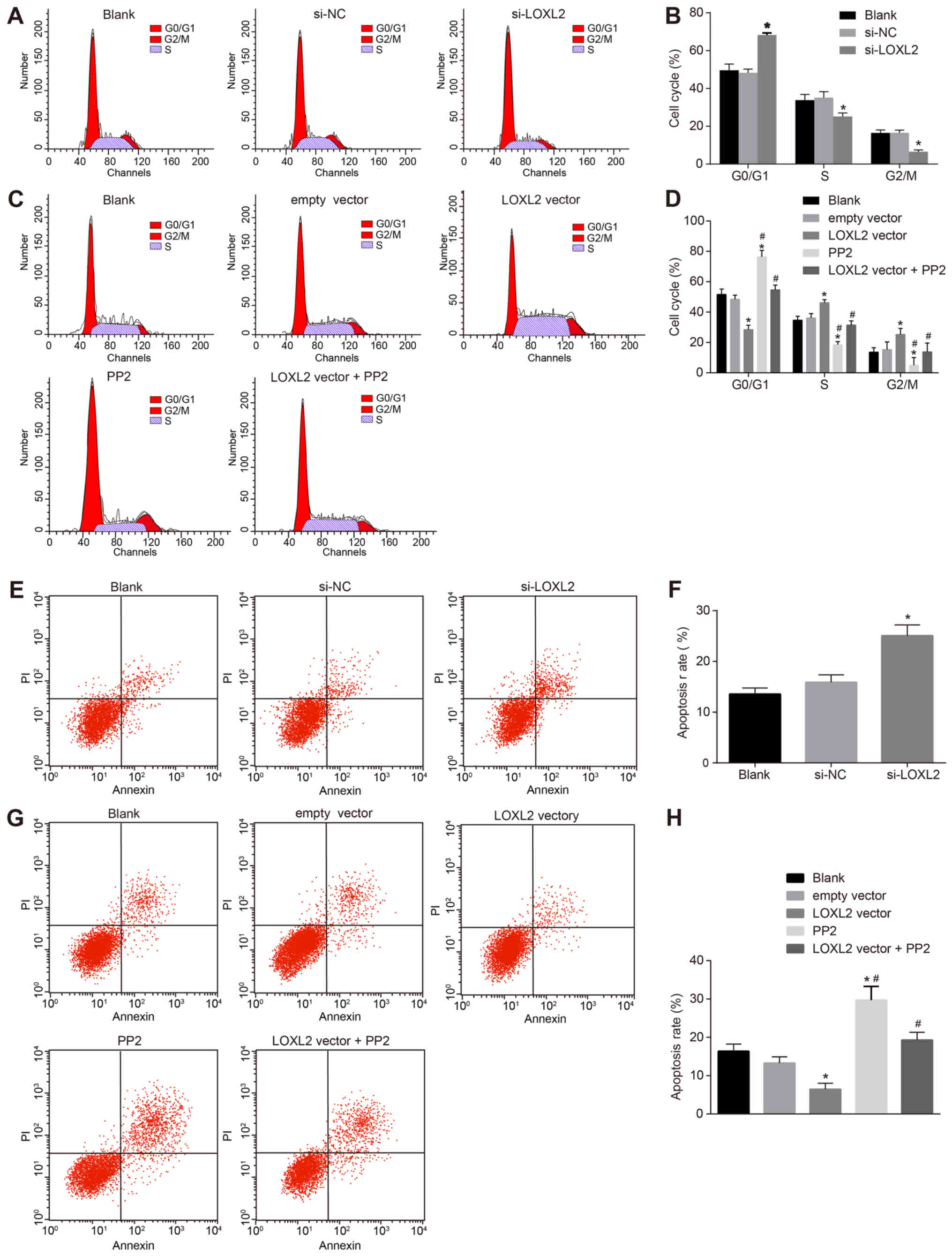
Silencing of LOXL2 inhibits the cell
cycle distribution and promotes the apoptosis of RCC cells
The cell cycle detection of RCC cells was performed
by flow cytometry (Fig. 10A and
B). The results demonstrated that, compared with cells in the
blank group, once 786-O cells were transfected with si-LOXL2, there
were a significantly higher number of cells at the G1 phase, and a
significantly lower number in the S and G2/M phases (P<0.05),
indicating that silencing LOXL2 may inhibit cell cycle distribution
in RCC cells. In comparison with cells in the blank and empty
vector groups, once Caki1 cells were transfected with LOXL2 vector,
a significantly fewer number of cells were arrested at the G1 phase
and a significantly greater number of cells were arrested at the S
and G2/M phases (all P<0.05). The number of cells at the G1
phase increased significantly, and decreased significantly at the S
and G2/M phases, in the PP2 group compared with the blank, empty
vector and LOXL2 vector groups (all P<0.05). Compared with cells
in the LOXL2 vector group, in the LOXL2 vector + PP2 group, a
significantly greater number of cells were arrested at the G1 phase
and significantly fewer cells at the S and G2/M phases (all
P<0.05; Fig. 10C and D),
indicating that suppression of LOXL2 expression and inactivation of
the FAK/Src pathway may reduce the percentage of RCC cells at the S
phase.
Apoptosis of RCC cells was assessed by flow
cytometry. The results indicated that, compared with cells in the
blank group, once 786-O cells were transfected with si-LOXL2, cell
apoptosis increased significantly (P<0.05; Fig. 10E and F). Once Caki1 cells were
transfected with the LOXL2 vector, cell apoptosis was significantly
decreased (P<0.05), whereas that in the PP2 and PP2 + LOXL2
vector groups was significantly increased (P<0.05) compared with
the blank and empty vector groups. Additionally, compared with
cells in the LOXL2 vector group, cell apoptosis in the LOXL2 vector
+ PP2 group was significantly increased (Fig. 10G and H). These data suggest that,
if the LOXL2 expression and activation of the FAK/Src pathway are
inhibited, the apoptosis of RCC cells is promoted. Therefore, PP2
reverses the effect of LOXL2 on RCC cell apoptosis. In conclusion,
LOXL2 silencing inhibits the cell cycle distribution and promotes
the apoptosis of RCC cells.
Discussion
RCC is commonly a fatal malignancy due to late
diagnosis and poor treatment options (21). The aim of the present study was to
investigate the effects and potential mechanisms of action of LOXL2
on the EMT and proliferation and invasion of RCC cells through the
Src/FAK signaling pathway.
The results demonstrated that the expression of
LOXL2 in RCC tissues was increased compared with that in adjacent
normal tissues. The expression of LOXL2 was detected in renal
tissue and in tubular epithelial cells in human kidneys, and LOXL2
functions as a promoter for the progression of tubulointerstitial
fibrosis (22). A previous study
reported that, in comparison with adjacent non-cancerous renal
tissues, LOXL2 was substantially elevated in specimens of human
clear cell RCC (23). Furthermore,
the expression of LOX in the kidney was revealed to be markedly
elevated in hyperuricemia, which is associated with renal diseases
(24).
According to the results of the clinicopathological
characteristics analysis, LOXL2 expression was revealed to be
associated with tumor grade, stage (Fuhrman and TNM), type,
diameter and venous invasion. The expression of LOXL2 was increased
in patients with tumor grade III-IV, a tumor diameter ≥4 cm and
venous invasion. RCC staging is mainly based on the TNM staging
system, which, together with tumor grade, provides necessary
information regarding treatment and prognosis (25). At the time of symptomatic
diagnosis, 30-40% of patients with RCC will have already developed
metastasis to the lymph nodes or other organs (4). Although patients with RCC have a
5-year survival rate of 85%, ~1/3 of patients are diagnosed with a
metastatic or advanced stage of the disease, with a 5-year survival
rate of 10% (26). A previous
study reported that high LOX protein expression was associated with
the occurrence and number of lymph node metastases (27). In addition, the high expression of
LOXL2 has been reported to be associated with the poor prognosis of
patients with colon cancer, and it may be applied in clinical
practice for risk classification (28).
The Src/FAK signaling pathway-associated genes FAK
and Src exhibited increased expression in RCC cells compared with
in normal renal tubular epithelial cells. Src is a non-receptor
tyrosine kinase that serves a key role in regulating multiple
cellular functions, including cell proliferation, adhesion and
survival (29), whereas FAK
participates in tumor cell migration and metastasis (30). The mRNA expression of Src was high
in RCC samples compared with that in normal kidney samples, and Src
may promote the occurrence of malignant phenotypes of RCC due to
the resistance to apoptosis induced by B-cell lymphoma-extra large
and angiogenesis by Src/signal transducer and activator of
transcription 3 (STAT3)/vascular endothelial growth factor
signaling (31). FAK has been
detected in adult RCC tissues, and it is has also been hypothesized
to be involved in pediatric renal tumor types through affecting
cell survival (32).
A key result of the present study was that, compared
with the blank group, the expression of FAK, Src, MMP-9, N-cadherin
and vimentin was substantially decreased in the si-LOXL2 and PP2
groups, while the expression of E-cadherin was notably increased,
indicating that the silencing of LOXL2 may inactivate the Src/FAK
signaling pathway and inhibit EMT. LOXL2, through biochemical and
biomechanical mechanisms, is capable of stimulating tumor
development, potentially by activating a diverse range of signaling
pathways (12). According to a
previous report, secreted LOXL2 and LOXL4 were revealed to regulate
the Src/FAK signaling pathway in gastric cancer (33,34).
LOX may also affect colorectal cancer by mechanisms involving Src
and FAK (35,36). MMP-9 serves a pivotal role in tumor
angiogenesis, cell invasion and migration, in addition to EMT
regulation (37,38). EMT is generally considered to
participate in cancer progression and metastasis (39). N-cadherin is considered to be the
mesenchymal marker in the process of EMT; E-cadherin loss and
vimentin gain are two crucial steps in EMT (40,41).
LOXL2 promotes invasion by regulating the expression and activity
of the extracellular protein tissue inhibitor of MMP-9 (42). Furthermore, accumulating evidence
indicates that LOXL2 downregulates E-cadherin expression and
stimulates EMT (43). The Src/FAK
signaling pathway may also participate in the EMT of thyroid cancer
cells (44). LOXL2 in breast
cancer cells has been demonstrated to activate oncogenic signaling
pathways, resulting in EMT by extracellular in addition to
intracellular mechanisms (45).
Therefore, it has been hypothesized that LOXL2 silencing may
inhibit EMT in RCC cells via downregulating the Src/FAK signaling
pathway.
Another notable observation of the present study was
that silencing of LOXL2 inhibits RCC cell migration and invasion
though the inactivation of the Src/FAK signaling pathway. The
inactivation of the Src/STAT3 signaling pathway may induce
apoptosis of human renal carcinoma Caki1 cells (46). FAK/Src signaling serves a crucial
role in the invasion of neoplastic cells (47). LOXL2, a member of an amine oxidase
family, has been suggested to be implicated in cross-link formation
in stromal collagens and elastin, cell motility, and tumor
formation and progression (48).
The inhibition of LOXL2 substantially inhibits cancer cell
migration and invasion in RCC (49). LOXL2 expression in stromal
fibroblasts was also revealed to be associated with the depth of
tumor invasion, lymphatic and venous invasion, in addition to
peritoneal dissemination in gastric cancer (50).
In conclusion, the results of the present study
indicated that LOXL2 silencing inhibits the EMT, proliferation and
invasion of RCC cells through the Src/FAK signaling pathway, which
may provide a novel approach to the treatment of RCC. However, the
specific mechanisms have yet to be fully elucidated and the sample
size of the present study was limited; therefore, further
investigation is required.
Funding
Not applicable.
Availability of data and materials
The analysed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
XH conceived the study and XH and JJY designed the
study. JJY was involved in data collection. XH performed the
statistical analysis and preparation of figures. XH drafted the
paper. JJY contributed substantially to its revision. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The study protocol was approved by the Ethics
Committee of Shanghai Jiao Tong University Affiliated Sixth
People's Hospital and Shanghai Jiao Tong University Affiliated
Sixth People's Hospital South Campus, and all the patients signed
informed consent forms. This study obtained the approval of the
Ethics Review Committee of the Ethics Committee of Shanghai Jiao
Tong University Affiliated Sixth People's Hospital and Shanghai
Jiao Tong University Affiliated Sixth People's Hospital South
Campus (approval no. 201407004).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Linehan WM, Srinivasan R and Schmidt LS:
The genetic basis of kidney cancer: A metabolic disease. Nat Rev
Urol. 7:277–285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tan HJ, Norton EC, Ye Z, Hafez KS, Gore JL
and Miller DC: Long-term survival following partial vs radical
nephrectomy among older patients with early-stage kidney cancer.
JAMA. 307:1629–1635. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gohara A, Eltaki N, Sabry D, Murtagh D Jr,
Jankun J, Selman SH and Skrzypczak-Jankun E: Human 5-, 12- and
15-lipoxygenase-1 coexist in kidney but show opposite trends and
their balance changes in cancer. Oncol Rep. 28:1275–1282. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morrissey JJ, London AN, Luo J and
Kharasch ED: Urinary biomarkers for the early diagnosis of kidney
cancer. Mayo Clin Proc. 85:413–421. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu SS, Quinn DI and Dorff TB: Clinical use
of cabozantinib in the treatment of advanced kidney cancer:
Efficacy, safety, and patient selection. OncoTargets Ther.
9:5825–5837. 2016. View Article : Google Scholar
|
|
6
|
Chow WH, Dong LM and Devesa SS:
Epidemiology and risk factors for kidney cancer. Nat Rev Urol.
7:245–257. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jonasch E: Incorporating New Systemic
Therapies in Kidney Cancer Treatment. J Natl Compr Canc Netw.
15(5S): 703–705. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dent P, Curiel DT and Fisher PB: The
potential of virus-based gene therapies for treatment of metastatic
kidney cancer. Expert Rev Anticancer Ther. 11:809–811. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Froger L, Neuzillet Y and Lebret T:
Indications for ablative treatment in kidney cancer in the elderly.
Prog Urol. 22:1004–1009. 2012.In French. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jaganjac S, Schefe L, Avdagić E, Spahović
H and Hiros M: Preoperative kidney tumor embolization as procedure
for therapy of advanced kidney cancer. Acta Inform Med. 22:302–305.
2014. View Article : Google Scholar
|
|
11
|
Kim YM, Kim EC and Kim Y: The human lysyl
oxidase-like 2 protein functions as an amine oxidase toward
collagen and elastin. Mol Biol Rep. 38:145–149. 2011. View Article : Google Scholar
|
|
12
|
Wu L and Zhu Y: The function and
mechanisms of action of LOXL2 in cancer (Review). Int J Mol Med.
36:1200–1204. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nishikawa R, Chiyomaru T, Enokida H,
Inoguchi S, Ishihara T, Matsushita R, Goto Y, Fukumoto I, Nakagawa
M and Seki N: Tumour-suppressive microRNA-29s directly regulate
LOXL2 expression and inhibit cancer cell migration and invasion in
renal cell carcinoma. FEBS Lett. 589:2136–2145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu D, Yao Q, Zhan C, Le-Meng Z, Liu H, Cai
Y, Tu C, Li X, Zou Y and Zhang S: MicroRNA-146a promote cell
migration and invasion in human colorectal cancer via
carboxypeptidase M/src-FAK pathway. Oncotarget. 8:22674–22684.
2017.PubMed/NCBI
|
|
15
|
Suwaki N, Vanhecke E, Atkins KM, Graf M,
Swabey K, Huang P, Schraml P, Moch H, Cassidy AM, Brewer D, et al:
A HIF-regulated VHL-PTP1B-Src signaling axis identifies a
therapeutic target in renal cell carcinoma. Sci Transl Med.
3:85ra472011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Barker HE, Bird D, Lang G and Erler JT:
Tumor-secreted LOXL2 activates fibroblasts through FAK signaling.
Mol Cancer Res. 11:1425–1436. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim BR, Dong SM, Seo SH, Lee JH, Lee JM,
Lee SH and Rho SB: Lysyl oxidase-like 2 (LOXL2) controls
tumor-associated cell proliferation through the interaction with
MARCKSL1. Cell Signal. 26:1765–1773. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Deckers IA, Schouten LJ, Van Neste L, van
Vlodrop IJ, Soetekouw PM, Baldewijns MM, Jeschke J, Ahuja N, Herman
JG, van den Brandt PA, et al: Promoter methylation of cdo1
identifies clear-cell renal cell cancer patients with poor survival
outcome. Clin Cancer Res. 21:3492–3500. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Brookman-May SD, May M, Wolff I, Zigeuner
R, Hutterer GC, Cindolo L, Schips L, De Cobelli O, Rocco B, De
Nunzio C, et al: CORONA Project; European Association of Urology
(EAU) Young Academic Urologists (YAU) Renal Cancer Group:
Evaluation of the prognostic significance of perirenal fat invasion
and tumor size in patients with pT1-pT3a localized renal cell
carcinoma in a comprehensive multicenter study of the CORONA
project. Can we improve prognostic discrimination for patients with
stage pT3a tumors? Eur Urol. 67:943–951. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kotsopoulos J, Zhang S, Akbari M, Salmena
L, Llacuachaqui M, Zeligs M, Sun P and Narod SA: BRCA1 mRNA levels
following a 4-6-week intervention with oral 3,3'-diindolylmethane.
Br J Cancer. 111:1269–1274. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mickley A, Kovaleva O, Kzhyshkowska J and
Gratchev A: Molecular and immunologic markers of kidney
cancer-potential applications in predictive, preventive and
personalized medicine. EPMA J. 6:202015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Choi SE, Jeon N, Choi HY, Shin JI, Jeong
HJ and Lim BJ: Lysyl oxidase like 2 is expressed in kidney tissue
and is associated with the progression of tubulointerstitial
fibrosis. Mol Med Rep. 16:2477–2482. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hase H, Jingushi K, Ueda Y, Kitae K, Egawa
H, Ohshio I, Kawakami R, Kashiwagi Y, Tsukada Y, Kobayashi T, et
al: LOXL2 status correlates with tumor stage and regulates integrin
levels to promote tumor progression in ccRCC. Mol Cancer Res.
12:1807–1817. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang Z, Xiaohua W, Lei J, Ruoyun T,
Mingxia X, Weichun H, Li F, Ping W and Junwei Y: Uric acid
increases fibronectin synthesis through upregulation of lysyl
oxidase expression in rat renal tubular epithelial cells. Am J
Physiol Renal Physiol. 299:F336–F346. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fiore DC and Fox CL: Urology and
nephrology update: Bladder and kidney cancer. FP Essent. 416:26–29.
2014.PubMed/NCBI
|
|
26
|
Bianchi L, Rossi L, Tomao F, Papa A,
Zoratto F and Tomao S: Thyroid dysfunction and tyrosine kinase
inhibitors in renal cell carcinoma. Endocr Relat Cancer.
20:R233–R245. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sakai M, Kato H, Sano A, Tanaka N, Inose
T, Kimura H, Sohda M, Nakajima M and Kuwano H: Expression of lysyl
oxidase is correlated with lymph node metastasis and poor prognosis
in esophageal squamous cell carcinoma. Ann Surg Oncol.
16:2494–2501. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Torres S, Garcia-Palmero I, Herrera M,
Bartolomé RA, Peña C, Fernandez-Aceñero MJ, Padilla G,
Peláez-García A, Lopez-Lucendo M, Rodriguez-Merlo R, et al: LOXL2
Is Highly Expressed in Cancer-Associated Fibroblasts and Associates
to Poor Colon Cancer Survival. Clin Cancer Res. 21:4892–4902. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chojnacka K and Mruk DD: The Src
non-receptor tyrosine kinase paradigm: New insights into mammalian
Sertoli cell biology. Mol Cell Endocrinol. 415:133–142. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pei G, Lan Y, Chen D, Ji L and Hua ZC: FAK
regulates E-cadherin expression via p-SrcY416/p-ERK1/2/p-Stat3Y705
and PPARγ/miR-125b/Stat3 signaling pathway in B16F10 melanoma
cells. Oncotarget. 8:13898–13908. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Majid S, Saini S, Dar AA, Hirata H,
Shahryari V, Tanaka Y, Yamamura S, Ueno K, Zaman MS, Singh K, et
al: MicroRNA-205 inhibits Src-mediated oncogenic pathways in renal
cancer. Cancer Res. 71:2611–2621. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Megison ML, Gillory LA, Stewart JE, Nabers
HC, Mrozcek-Musulman E and Beierle EA: FAK inhibition abrogates the
malignant phenotype in aggressive pediatric renal tumors. Mol
Cancer Res. 12:514–526. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peng L, Ran YL, Hu H, Yu L, Liu Q, Zhou Z,
Sun YM, Sun LC, Pan J, Sun LX, et al: Secreted LOXL2 is a novel
therapeutic target that promotes gastric cancer metastasis via the
Src/FAK pathway. Carcinogenesis. 30:1660–1669. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li RK, Zhao WY, Fang F, Zhuang C, Zhang
XX, Yang XM, Jiang SH, Kong FZ, Tu L, Zhang WM, et al: Lysyl
oxidase-like 4 (LOXL4) promotes proliferation and metastasis of
gastric cancer via FAK/Src pathway. J Cancer Res Clin Oncol.
141:269–281. 2015. View Article : Google Scholar
|
|
35
|
Baker AM, Cox TR, Bird D, Lang G, Murray
GI, Sun XF, Southall SM, Wilson JR and Erler JT: The role of lysyl
oxidase in SRC-dependent proliferation and metastasis of colorectal
cancer. J Natl Cancer Inst. 103:407–424. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Baker AM, Bird D, Lang G, Cox TR and Erler
JT: Lysyl oxidase enzymatic function increases stiffness to drive
colorectal cancer progression through FAK. Oncogene. 32:1863–1868.
2013. View Article : Google Scholar
|
|
37
|
Raghu H, Sodadasu PK, Malla RR, Gondi CS,
Estes N and Rao JS: Localization of uPAR and MMP-9 in lipid rafts
is critical for migration, invasion and angiogenesis in human
breast cancer cells. BMC Cancer. 10:6472010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Asuthkar S, Nalla AK, Gondi CS, Dinh DH,
Gujrati M, Mohanam S and Rao JS: Gadd45a sensitizes medulloblastoma
cells to irradiation and suppresses MMP-9-mediated EMT.
Neuro-oncol. 13:1059–1073. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Iwatsuki M, Mimori K, Yokobori T, Ishi H,
Beppu T, Nakamori S, Baba H and Mori M: Epithelial-mesenchymal
transition in cancer development and its clinical significance.
Cancer Sci. 101:293–299. 2010. View Article : Google Scholar
|
|
40
|
Zeng T, Peng L, Chao C, Fu B, Wang G, Wang
Y and Zhu X: miR-451 inhibits invasion and proliferation of bladder
cancer by regulating EMT. Int J Clin Exp Pathol. 7:7653–7662.
2014.
|
|
41
|
Myong NH: Loss of E-cadherin and
Acquisition of Vimentin in Epithelial-Mesenchymal Transition are
Noble Indicators of Uterine Cervix Cancer Progression. Korean J
Pathol. 46:341–348. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Barker HE, Chang J, Cox TR, Lang G, Bird
D, Nicolau M, Evans HR, Gartland A and Erler JT: LOXL2-mediated
matrix remodeling in metastasis and mammary gland involution.
Cancer Res. 71:1561–1572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Park JS, Lee JH, Lee YS, Kim JK, Dong SM
and Yoon DS: Emerging role of LOXL2 in the promotion of pancreas
cancer metastasis. Oncotarget. 7:42539–42552. 2016.PubMed/NCBI
|
|
44
|
Baquero P, Jiménez-Mora E, Santos A, Lasa
M and Chiloeches A: TGFβ induces epithelial-mesenchymal transition
of thyroid cancer cells by both the BRAF/MEK/ERK and Src/FAK
pathways. Mol Carcinog. 55:1639–1654. 2016. View Article : Google Scholar
|
|
45
|
Moon HJ, Finney J, Xu L, Moore D, Welch DR
and Mure M: MCF-7 cells expressing nuclear associated lysyl
oxidase-like 2 (LOXL2) exhibit an epithelial-to-mesenchymal
transition (EMT) phenotype and are highly invasive in vitro. J Biol
Chem. 288:30000–30008. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Park JE, Park B, Chae IG, Kim DH, Kundu J,
Kundu JK and Chun KS: [Corrigendum] Carnosic acid induces apoptosis
through inactivation of Src/STAT3 signaling pathway in human renal
carcinoma Caki cells. Oncol Rep. 36:37172016. View Article : Google Scholar
|
|
47
|
Tai G, Hohenstein P and Davies JA: FAK-Src
signalling is important to renal collecting duct morphogenesis:
Discovery using a hierarchical screening technique. Biol Open.
2:416–423. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhan P, Shen XK, Qian Q, Zhu JP, Zhang Y,
Xie HY, Xu CH, Hao KK, Hu W, Xia N, et al: Down-regulation of lysyl
oxidase-like 2 (LOXL2) is associated with disease progression in
lung adenocarcinomas. Med Oncol. 29:648–655. 2012. View Article : Google Scholar
|
|
49
|
Kurozumi A, Kato M, Goto Y, Matsushita R,
Nishikawa R, Okato A, Fukumoto I, Ichikawa T and Seki N: Regulation
of the collagen cross-linking enzymes LOXL2 and PLOD2 by
tumor-suppressive microRNA-26a/b in renal cell carcinoma. Int J
Oncol. 48:1837–1846. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kasashima H, Yashiro M, Kinoshita H,
Fukuoka T, Morisaki T, Masuda G, Sakurai K, Kubo N, Ohira M and
Hirakawa K: Lysyl oxidase-like 2 (LOXL2) from stromal fibroblasts
stimulates the progression of gastric cancer. Cancer Lett.
354:438–446. 2014. View Article : Google Scholar : PubMed/NCBI
|