Introduction
Breast cancer (BC) has ranked as the
second-prevalent malignant tumor worldwide and poses a substantial
threat to women's health (1).
Ahmad (1) reported that 25% of
cancer cases in 2020 were new BC cases. Various treatment methods
were applied in the field of therapy for patients with BC with
different stages (from 0 to IV), including surgery, chemotherapy
and targeted therapy. The following therapy techniques have been
proven to effectively reduce both the occurrence and mortality
rates among patients at all BC stages, ranging from stage 0 to
stage IV (2). Nevertheless, 42% of
patients with BC are found to encounter difficulties in achieving
full recovery since they may experience treatment-related side
effects such as fatigue, hair loss and increased risk of infection,
among others. These side effects can significantly impair the
quality of life for patients (2).
Hence, it is imperative to enhance the efficacy of current
treatment strategies for BC. To attain this goal, numerous
researchers highlighted the potential of circular RNAs (circRNAs)
in BC diagnosis, treatment and prognosis (3).
CircRNAs circular structure (a closed-loop structure
devoid of a 5'-cap or a 3'-polyadenylated tail) is more stable
compared with the linear structure of microRNAs (miRNAs or miRs).
Additionally, circRNAs modulate gene expression and cancer
progression through four major mechanisms, including miRNA
sponging, transcriptional regulation, protein scaffolding and
translation (3-6).
Moreover, numerous researchers have recently focused on the sponge
function of circRNAs. For example, circRNAs act as competing
endogenous RNAs, sequestering miRNAs to prevent degradation of
targeted mRNAs by bonding with them. Exon-derived circRNAs (for
example, ciRS-7 and circ-ITCH) could act as miRNA sponge for RNA
binding, resulting in negative regulation of miRNA and downstream
target genes (7,8).
In the BC field, researchers have found pathways,
such as the circRNA-miRNA-mRNA, that are associated with
tumorigenesis, progression and metastasis. For example,
hsa_circ_0003645-miR-139-3p/high mobility group box 1 (HMGB1) has
been shown to promote the proliferation of BC cells (9). Moreover, the Gene Expression Omnibus
Database (GEO; https://www.ncbi.nlm.nih.gov/geo/ is an authority
database supported by the National Center for Biotechnology
Information (NCBI), and it can be used to characterize gene
function (10,11). Via GEO analysis, a new circRNA was
found, namely hsa_circ_0001508, showing high expression in BC
tissue. To the best of our knowledge, there is no research focusing
on this new gene across PubMed and Embase databases. Consequently,
hsa_circ_0001508 has become a focus of the present study.
Additionally, TargetScan (https://www.targetscan.org/) is an online available
bioinformatics tool useful to predict pathways by finding possible
binding sites, and its application only has a low false positive
rate of 30% (12). The present
study identified two possible pathways through the application of
TargetScan, including hsa_circ_0001508/miR-505-3p/HMGB1 as well as
hsa_circ_0001508/miR-505-3p/vestigial-like family member 3 (VGLL3),
in which there are some possible binding sites between these
genes.
Moreover, previous research indicates that
miR-505-3p (tumor suppressor in BC), HMGB1 (oncogene) and VGLL3
(oncogene) are associated with BC (13,14).
miR-505-3p was found to have binding sites for HMGB1 mRNA [for
example, 3'-untranslated regions (3'-UTR) in HMGB1 mRNA position
417-424]; miR-505-3p can also negatively regulate the expression of
HMGB1, leading to hepatocellular carcinoma cell decreasing
(15). However, to the best of our
knowledge, there are no previous studies focused on the binding
sites between miR-505-3p and VGLL3.
In summary, in the BC field, the
circRNA-miRNA-targeted gene axis has already been studied for a
long time, but several new genes still have not been investigated.
Based on the results of previous predictions via TargetScan and GEO
analysis, a pilot study was conducted, including the biological
function of hsa_circ_0001508 in BC and its possible mechanism in
hsa_circ_0001508/miR-505-3p/HMGB1 as well as
hsa_circ_0001508/miR-505-3p/VGLL3.
Materials and methods
Gene Chip data collection and
analyses
CircRNAs data of BC were obtained from the NCBI
database (accession no. GSE182471; https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE182471).
The raw data obtained were subjected to two types of analysis:
Functional enrichment analysis and biological information analysis
using various software tools including CRAFT v1.0 (https://github.com/annadalmolin/CRAFT),
miRbase (https://www.mirbase.org), the Circular
RNA interactome database (https://circinteractome.nia.nih.gov), circMIR
(https://www.bio-inf.cn/circmir/) and
TargetScan (https://www.targetscan.org/).
Cell lines
BC cell lines, including HCC1806, HCC1937 and
CAL-51, and MCF10A cells (non-cancerous cell line), were obtained
from The Cell Bank of Type Culture Collection of The Chinese
Academy of Sciences. Cells were cultured in a medium containing
RPMI-1640 supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.), 1% penicillin and 1% streptomycin for Cell
Culture (Beyotime Institute of Biotechnology) in an incubator at
37˚C with 5% CO2.
The HCC1937 cell line was chosen for the next
experimental steps since hsa_circ_0001508 in HCC1937 cells
presented the highest expression compared with the rest of the cell
lines (HCC1806, CAL-51 and MCF10A cells). Furthermore, the HCC1937
cell line is BRCA1-mutated, which sets it apart from the HCC1806
and CAL-51 cell lines. Additionally, the combined expression of
miR-505-3p and BRCA1 could be utilized to differentiate between
different types of BC, indicating its potential clinical diagnostic
value (16). As a result, the
present study focused on the HCC1937 cell line due to its potential
clinical implications.
Based on the results of, the HCC1937 cell line was
chosen as a major cell line for the next experimental steps as the
expression of hsa_circ_0001508 in HCC1937 cells exhibited the
highest expression compared with the other cell lines (HCC1806,
CAL-51 and MCF10A cells). Furthermore, the HCC1937 cell line is
BRCA1-mutated, which distinguishes it from the HCC1806 and CAL-51
cell lines. Additionally, the combined expression of miR-505-3p and
BRCA1 could be utilized to differentiate between different types of
BC, indicating its potential clinical diagnostic value (16). As a result, the present study
focused on the HCC1937 cell line due to its potential clinical
implications. Consequently, the present study focused on the
HCC1937 cell line due to its potential clinical implications.
Transfection
Aiming to knockdown hsa_circ_0001508, HCC1937 cells
were transfected with small interfering (si)RNAs for circ_0001508
(si-circ_0001508; 5'-ACATAAATG-ATAAATCTGTATTATTTGG-3'; 10 nM) and
si-negative control (siNC; 5'-TTCTCC-GAACGTGT-CACGT-3'; 10 nM). All
vectors were purchased from Sangon Biotech Co., Ltd., and all
transfections were performed using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.), following the
manufacturer's protocol. Subsequently, the cells were incubated at
37˚C with 5% CO2 for 12 h, the culture medium was
replaced with fresh medium containing 10% FBS and the subsequent
experimental process was conducted within 24 h.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted using the TriQuick reagent
(Thermo Fisher Scientific, Inc.) and reverse-transcribed into cDNA
using the Quantscript RT Kit (Tiangen Biotech Co., Ltd.), according
to the manufacturer's protocol. The reaction condition for the
reverse transcription was 37˚C for 60 min. Subsequently, qPCR was
performed using RealMaster Mix (SYBR Green; Tiangen Biotech Co.,
Ltd.) on an ABI 7500 system (Thermo Fisher Scientific, Inc.) with
the following thermocycling conditions: Initial denaturation at
95˚C for 10 min followed by 40 cycles at 94˚C for 15 sec and 60˚C
for 60 sec. The expression of genes was calculated by using the
2-ΔΔCq method (17).
Both U6 and GAPDH were used as references for the expression of
hsa_circ_0001058 and miRNA, respectively. The sequences of the
primers used for qPCR were as follows: Circ_0001508 forward,
5'-AATGGAAGGCACCCTGTGATT-3' and reverse,
5'-CAGCTTTTGTGCTTCCAACCT-3'; miR-505-3p forward,
5-CGTCAACACTTGCTGGTTTCCTA-3' and reverse,
5'-ACGAATTTGCGTGTCATCC-3'; HMGB1 forward,
5'-TCAAAGGAGAACATCCTGGCCTGT-3' and reverse,
5'-CTGCTTGTCATCTGCAGCAGTGTT-3'; VGLL3 forward,
5'-CCAACTACAGTCACCTCTGCTAC-3' and reverse,
5'-ACCACGGTGATTCCTTACTCTTG-3'; GAPDH forward,
5'-GGAGTCCACTGGCGTCTTCA-3' and reverse, 5'-GTCATGAGTCCTTCCA
CGATACC-3'; and U6 forward, 5'-CTCGCTTCGGCAGCACA-3'and reverse,
5'-AACGCTTCACGAATTTGCGT-3'.
Cell counting MTT assay
HCC1937 cells were seeded into 96-well plates
(1.0x103 cells/well) and incubated at 37˚C with 5%
CO2 for 12, 24, 48, 72 and 96 h. A total of 20 µl of MTT
reagent (Procell Life Science & Technology Co., Ltd.) was added
into each well and subsequently incubated for 4 h at 37˚C. The
absorbance of each cell was measured at a wavelength of 490 nm by
using a microplate reader (Thermo Fisher Scientific, Inc.).
Cell invasion assay
The invasion of HCC1937 cells was observed by using
a 24-well Transwell chamber. A total of 10 µl Matrigel (BD
Biosciences) was placed in the lower compartment of a Transwell
chamber and incubated at 37˚C for 2 h. HCC1937 cells
(1x104 cells/well) were cultured for 12 h at 37˚C in the
upper compartment. Subsequently, 100 µl FBS was added to the lower
compartment culture medium. After 24 h, the cell invasion in the
lower surface of the compartment was observed with a light
microscope (magnification, x100; Olympus Corporation) after
staining with crystal violet staining solution (cat. no. C0121;
Beyotime Institute of Biotechnology). A total of five visual fields
(upper, lower, central, left, and right) were randomly selected for
counting. The pore diameter of Transwell membrane is 0.8 µm.
Wound healing assay
The transfected HCC1937 cells were cultured in a
6-well plate with serum-free culture medium. After scratching, the
cells were incubated for 24 h at 37˚C. Images were captured at the
same site at the start of the experiment and after 24 h.
Flow cytometry assay
Cells (3x105) were prepared following the
instructions provided in the reagent kit (BD Biosciences).
Subsequently, the cells were resuspended after mixed 100 µl of
binding buffer. A total of 4-5 µl of FITC-Annexin V and 5 µl of PI
working solution was used to stain the cells, which were stored on
ice with dark conditions for 15 min. Cell apoptosis analysis was
performed using a flow cytometer (DxFLEX Flow Cytometer;
CytExpert™ Software; Beckman Coulter, Inc.).
Western blot assay
The protein extraction buffer (cell lysis buffer for
Western and IP, Biosharp; PMSF, Beijing Solarbio Science &
Technology Co., Ltd.) was used. Total protein was collected and
transferred onto a PVDF membrane, which was subsequently blocked
for 1-2 h. The membrane was then incubated overnight at 4˚C with
the primary antibodies (HMGB1 from rabbit and VGLL3 from mouse)
followed by incubation at room temperature with the secondary
antibody for 2 h. The bicinchoninic acid method was used to
determine protein concentration. A total of 20 µg of protein were
loaded per lane and subjected to SDS-PAGE (percentage of gel, 20%).
Blocking was performed using non-fat dried milk in blocking reagent
(5% TBST with 05% Tween) at 37˚C. The visualisation reagent used
was SuperKine™ West Femto Maximum Sensitivity Substrate
Abbkine Scientific Co., Ltd.) and Gel imaging system, Tanon 4600SF
(Tanon Science and Technology Co., Ltd.) was used for densitometric
analysis. Antibodies used were as follows: HMGb1 (cat. no. 3935s;
Cell Signaling Technology, Inc.), secondary antibody anti-rabbit
IgG (HRP-linked; cat. no. 7074; Cell signaling Technology, Inc.),
VGLL3 (cat. no. ab68262; Abcam), secondary antibody goat Anti-Mouse
IgG H&L (HRP-linked; cat. no. ab205719; Abcam) and β-actin
[(13E5) rabbit mAb (HRP Conjugate); cat. no. 5125; Cell Signaling
Technology, Inc.]. The dilution of primary and secondary antibodies
was 1:1,000 and 1:2,000, respectively.
Statistical analysis
All data were collected and analyzed with SPSS
software (version 26.0; IBM Corp.) and GraphPad Prism software
(version 5.0; Dotmatics). Data are presented as the mean ± standard
deviation. Differences in the two groups were assessed using
one-way ANOVA followed by LSD (≤3 groups) or Bonferroni (>3
groups) as post hoc tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
Gene Chip analysis
BC-related data were retrieved from the NCBI website
and gene expression data from the GSE182471 dataset was extracted.
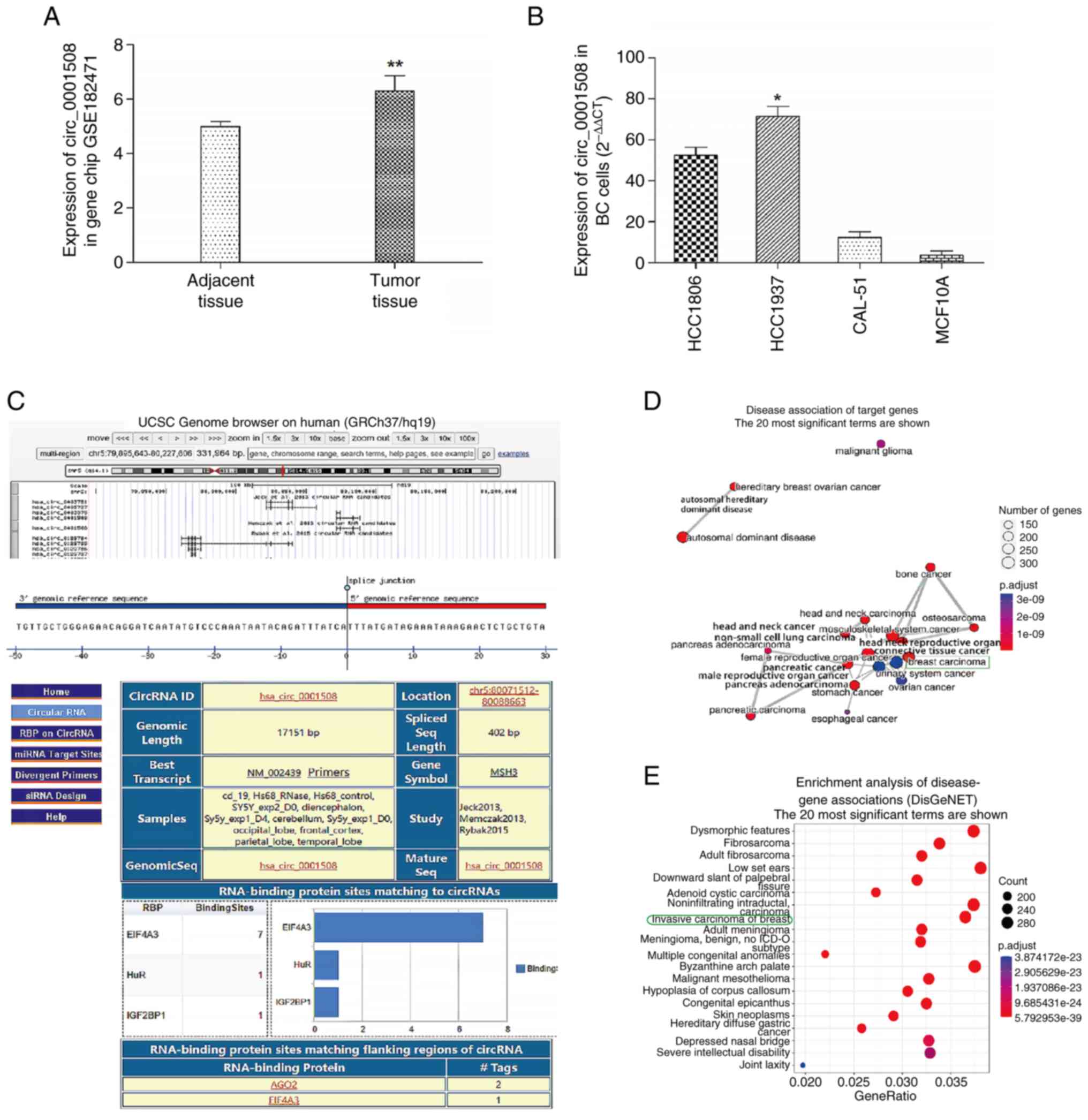
The expression levels of hsa_circ_0001508 were significantly higher
in BC tissues compared with adjacent BC normal tissues with fold
changes of 7 and 4.5, respectively (Fig. 1A).
hsa_circ_0001508 basic
information
The genomic location of hsa_circ_0001508 on
chromosome 5 (chr5.80071513-80088663) and its length of 17,151 base
pairs is demonstrated in Fig. 1C.
These results were sourced from the Circular RNA interactome
database (https://circinteractome.nia.nih.gov/api/v2/circsearch?circular_rna_query=hsa_circ_0001508+&gene_symbol_query=&submit=circRNA+Search).
Additionally, the functional analysis of hsa_circ_0001508 performed
by using cluster analysis software (CRAFT v1.0; https://github.com/annadalmolin/CRAFT)
revealed a higher relevance of hsa_circ_0001508 to BC compared with
other malignant tumors (Fig. 1D
and E).
Identification of hsa_circ_0001508
expression in each cell line
The results of the RT-qPCR assay showed various
expression levels for hsa_circ_0001508 in different cell lines,
including the BC cell lines (HCC1806, HCC1937 and CAL-51) and the
non-cancerous cell line MCF10A. The HCC1937 cells showed the
highest expression of hsa_circ_0001508 among all the cell lines
used in the present study (Fig.
1B). For example, the expression level of hsa_circ_0001508 in
HCC1937 cells was 25-fold higher than that observed in MCF10A
cells. Consequently, HCC1937 cells were selected for subsequent
experiments.
Binding interaction between
hsa_circ_0001508 and miR-505-3p
The potential sequestering of binding sites of
miR-505-3p within target genes were predicted using the circMIR
software (https://www.bio-inf.cn/circmir/). It was found that
there are two binding sites between hsa_circ_0001508 and
miR-505-3p, including 3'-UTR of miR-505 positions 223 and 224
(Table I).
 | Table Ihsa_circ_0001508-miR-505-3p
interactions were predicted using the circMIR. |
Table I
hsa_circ_0001508-miR-505-3p
interactions were predicted using the circMIR.
| Circular RNA ID | miRNA ID | Number of binding
sites predicted | Specific binding
sites predicted |
|---|
| hsa_circ_0001508 | miR-505-3p | 2 | 223,224 |
hsa_circ_0001508 knockdown inhibits
the proliferation and motility of HCC1937 cells
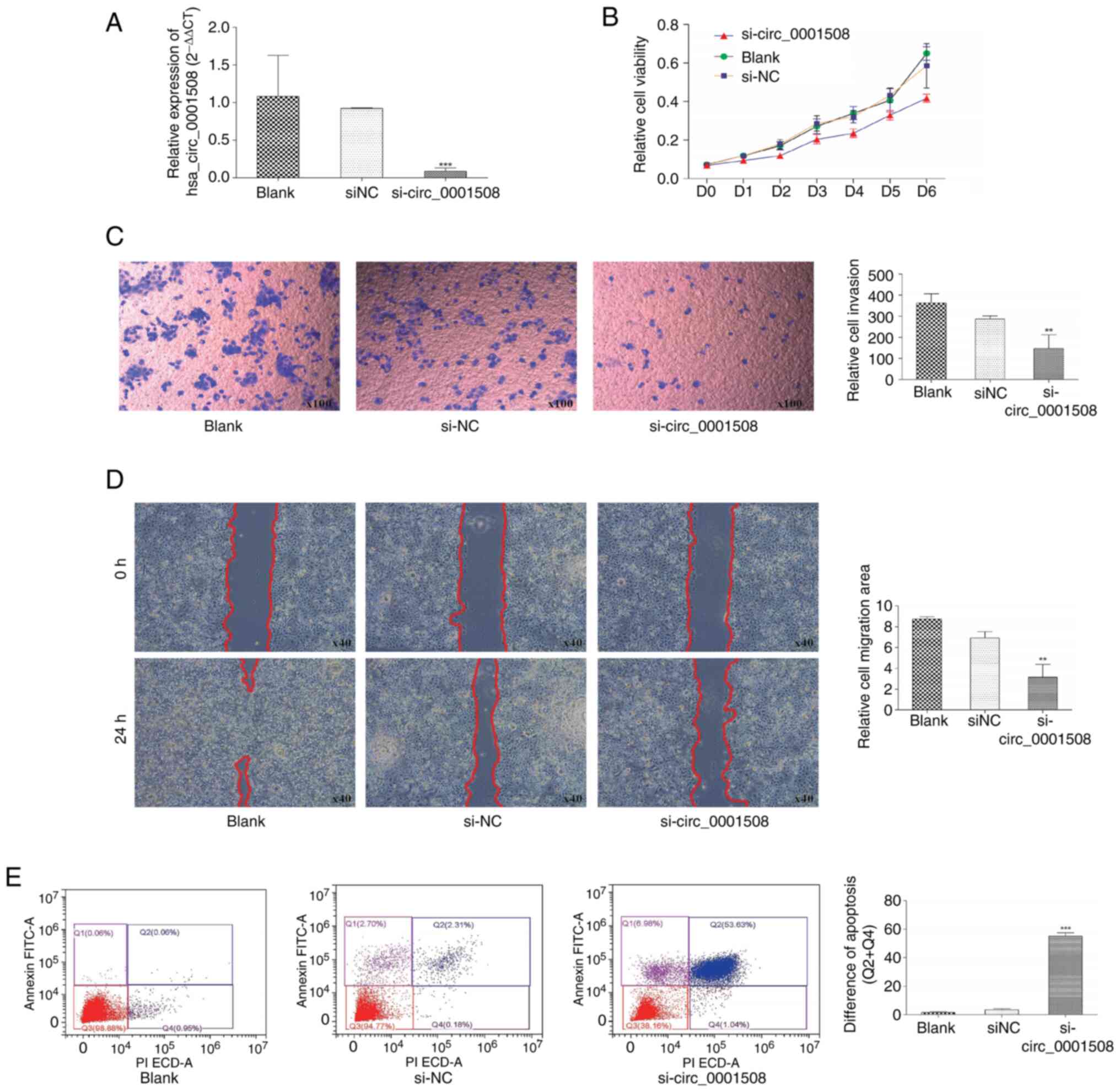
Knockdown experiments were conducted in the present
study using si-circ_0001508. Subsequently, to explore the
biological function of hsa_circ_0001508 in BC cells, the expression
of hsa_circ_0001508 was detected using RT-qPCR in three
experimental groups: si-circ_0001508; si-NC and blank. A
significant decrease was observed in the expression of
hsa_circ_0001508 in the si-circ_0001508 group compared with both
the blank and si-NC groups (Fig.
2A).
MTT analysis results revealed a significant decrease
in the proliferation of HCC1937 cells when hsa_circ_0001508 was
knocked down (si-circ_0001508 group), compared with the si-NC and
Blank group (Fig. 2B). Moreover,
Transwell and wound-healing assays indicated the involvement of
hsa_circ_0001508 in HCC1937 cell migration. For instance, the
si-circ_0001508 group exhibited fewer positively stained cells
(Fig. 2C), suggesting the invasion
of the cells in the si-circ_0001508 group was inhibited.
Additionally, would-healing assays which were conducted 24 h after
transfection indicated a significant delay in wound closure in the
hsa_circ_0001508 knockdown group compared with the si-NC group
(Fig. 2D).
The result of flow cytometry (Fig. 2E) revealed that the percentage of
apoptotic cells, including early and late apoptotic, in the
si-circ_0001508 group was significantly higher than in the si-NC
group and blank group. Additionally, when the hsa_circ_0001508 was
silenced, the percentage of early and late apoptotic cells was
found to be 53.63 and 1.04%, respectively. By contrast, the si-NC
group showed an early apoptotic rate of 2.31% and a late apoptotic
rate of 0.18%. These findings suggested that hsa_circ_0001508 may
play an oncogenic role in the progression of BC.
The aforementioned findings suggested an association
between hsa_circ_0001508 expression levels and the proliferation
and migration of BC cells. Notably, hsa_circ_0001508 may play an
oncogenic effect on HCC1937 BC cells, meaning that its
overexpression may promote BC tumorigenesis.
hsa_circ_0001508 affects the
expression of target genes HMGB1 and VGLL3 by regulating
miR-505-3p
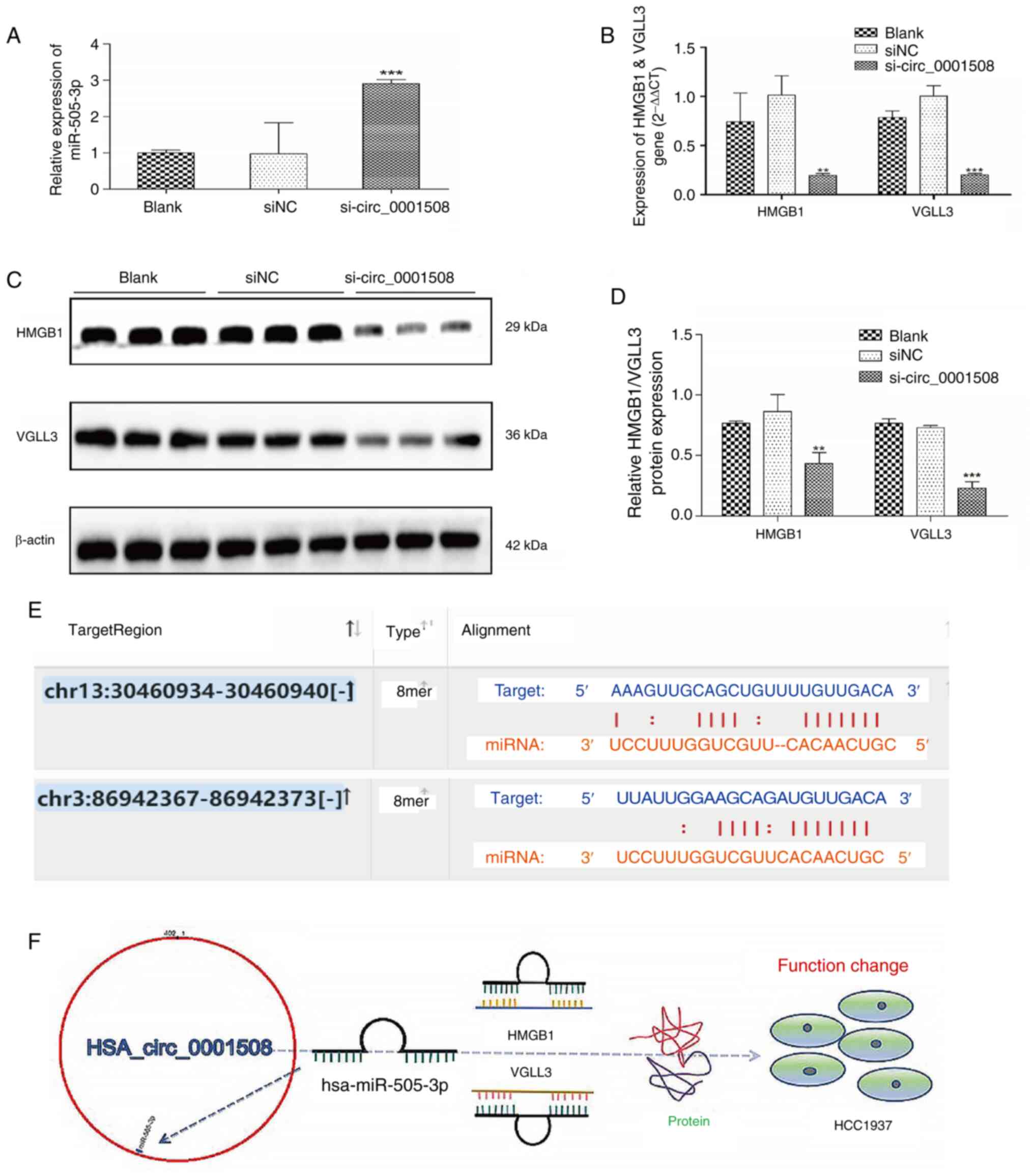
Bioinformatics analysis results were conducted with
the TargetScan tool, which indicated the putative binding sites of
miR-505-3p for the HMGB1 and VGLL3 genes (Fig. 3E). Additionally, the expression of
miR-505-3p was significantly increased in the si-circ_0001508 group
compared with the si-NC and blank groups (Fig. 3A). Similarly, the gene expression
analysis performed by RT-qPCR showed that the expression levels of
HMGB1 and VGLL3 were significantly lower in the si-circ_0001508
group compared with the si-NC and blank groups (Fig. 3B). The western blotting
demonstrated a significant decrease in the protein expression
levels of HMGB1 and VGLL3 in the si-circ_0001508 group (Fig. 3C and D).
The possible mechanism of action of hsa_circ_0001508
in HCC1937 cells is shown in Fig.
3F; it is suggested that hsa_circ_0001508 may act as
carcinogenic based on biological function and related molecular
regulation.
Discussion
The present results suggested that hsa_circ_0001508
serves as an oncogene in promoting BC progression. The findings of
the Transwell and wound-healing assays, MTT analysis, as well as
flow cytometry, revealed different expression levels of target
proteins in cells within three different experiment groups:
si-hsa_circ_0001508, si-NC and blank. These results demonstrate the
cancer-promoting function of hsa_circ_0001508 in BC. Furthermore,
there are no previous studies regarding the biological roles of
hsa_circ_0001508 in major biomedical databases (PubMed). The
present study provides empirical evidence to support the
cancer-promoting function of hsa_circ_0001508 in BC. Therefore, the
oncogene function of hsa_circ_0001508 could be a valuable focus for
future research in the BC field, including diagnosis, treatment and
prognosis.
hsa_circ_0001508 may play a vital role in regulating
HCC1937 cells through the hsa_circ_0001508/miR-505-3p/HMGB1 axis.
When hsa_circ_0001508 is silenced, the expression of the targeted
gene HMGB1 decreases. While other researchers have reported a
binding position [for example, 417-424(15)] between miR-505-3p and HMGB1, there
is no research focusing on possible binding sites in HCC1973 cell
lines. Additionally, the TargetScan results (Fig. 3E) predict one possible binding
site, such as 3'UTR of HMGB1 mRNA position 934-940. In conclusion,
it would be valuable to conduct a dual-luciferase assay to identify
the binding site.
Furthermore, the expression of VGLL3 was
significantly lower when hsa_circ_0001508 was silenced.
Additionally, TargetScan (Fig. 3E)
also indicated a potential binding site between miR-505-3p and
VGLL3, specifically within the 3'UTR of HMGB1 mRNA at position
367-373. As there is no existing research on miR-505-3p/VGLL3 in
BC, these findings could be valuable for future research. In
addition to miR-505-3p and its targeted possible binding sites, the
circMIR software (Table I)
reported that there are two binding sites located at position 223
and 224 in the 3'UTR of miR-505, although there is no research
supporting the competitive endogenous RNA crosstalk between
hsa_circ_0001508 and miR-505-3p, resulting in negative regulation
of miR-505 (inhibition) via sponge function to upstream target
genes (HMGB1 and VGLL3) in the development of BC.
Nevertheless, the present study does provide some
empirical evidence (for example, the expression of miR-505, HMGB1
and VGLL3 changed after hsa_circ_001508 silencing) to support the
research value of hsa_circ_0001508 and its related axis (for
example, hsa_circ_0001508/miR-505-3p/HMGB1 and
hsa_circ_0001508/miR-505-3p/VGLL3).
The study provides empirical evidence using
biomedical experimental tools and computer tools (for example,
target prediction, and miRNA and mRNA interaction analysis) to
support the biological function of hsa_circ_0001508 (BC activator)
and two possible circRNAs/miRNAs/mRNA axes. These findings are
valuable for researchers looking to expand their knowledge of the
BC field. However, due to financial limitations, there is a lack of
validation experiments in the present pilot study to investigate
these signaling pathways, such as dual-luciferase assay. However,
hsa_circ_0001508/miR-505-3p/HMGB1 and
hsa_circ_0001508/miR-505-3p/VGLL3 have research value for further
research.
BC has become a focal point in the biomedical field,
with several researchers focusing on circRNAs, especially
circRNAs/miRNAs/mRNA axis. The present study discovered a new
circRNA (hsa_circ_0001508) using data from Gene Chip analysis and
its novelty makes it valuable for research in the BC field. Certain
potential binding sites were found between
hsa_circ_0001508/miR-505-3p/HMGB1 and
hsa_circ_0001508/miR-505-3p/VGLL3, providing empirical evidence to
support that hsa_circ_0001508 could influence the expression of
miR-505-3p, HMGB1 and VGLL3. Based on these findings,
hsa_circ_0001508/miR-505-3p/HMGB1 and
hsa_circ_0001508/miR-505-3p/VGLL3 are considered valuable pathways
for future research to contribute more knowledge to the BC
field.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The data generated in the present study may be found
in the Gene Expression Omnibus under accession number GSE182471 or
at the following URL: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE182471.
The data generated in the present study may be found in the
Circular RNA Interactome database under accession number
hsa_circ_0001508 or at the following URL: https://circinteractome.nia.nih.gov/api/v2/circsearch?circular_rna_query=hsa_circ_0001508+&gene_symbol_query=&submit=circRNA+Search.
Authors' contributions
ZS played a leadership position, being responsible
for conducting the experimental process. FG was in another
leadership position (principal investigator), developing research
protocols and overseeing the data analysis. WD conducted
bioinformatics analysis. JQ and LC performed statistical analysis
and visual presentations of data. All authors read and approved the
final version of the manuscript. ZS and FG confirm the authenticity
of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ahmad A: Breast cancer: Current
perspectives on the disease status. In: Breast Cancer Metastasis
and Drug Resistance. Springer International Publishing AG, pp51-64,
2019.
|
|
2
|
Caswell-Jin JL, Sun LP, Munoz D, Lu Y, Li
Y, Huang H, Hampton JM, Song J, Jayasekera J, Schechter C, et al:
Analysis of breast cancer mortality in the US-1975 to 2019. JAMA.
331:233–241. 2024.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Huang X, Song C, Zhang J, Zhu L and Tang
H: Circular RNAs in breast cancer diagnosis, treatment and
prognosis. Oncol Res. 32:241–249. 2023.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Zhao X, Zhong Y, Wang X, Shen J and An W:
Advances in circular RNA and its applications. Int J Med Sci.
19:975–985. 2022.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Lei M, Zheng G, Ning Q, Zheng J and Dong
D: Translation and functional roles of circular RNAs in human
cancer. Mol Cancer. 19(30)2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Barbagallo D, Caponnetto A, Brex D,
Mirabella F, Barbagallo C, Lauretta G, Morrone A, Certo F, Broggi
G, Caltabiano R, et al: CircSMARCA5 regulates VEGFA mRNA splicing
and angiogenesis in glioblastoma multiforme through the binding of
SRSF1. Cancers (Basel). 11(194)2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7(11215)2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yang C, Yuan W, Yang X, Li P, Wang J, Han
J, Tao J, Li P, Yang H, Lv Q and Zhang W: Circular RNA circ-ITCH
inhibits bladder cancer progression by sponging miR-17/miR-224 and
regulating p21, PTEN expression. Mol Cancer. 17(19)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Zhang M, Bai X, Zeng X, Liu J, Liu F and
Zhang Z: circRNA-miRNA-mRNA in breast cancer. Clin Chim Acta.
523:120–130. 2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Clough E and Barrett T: The gene
expression omnibus database. In: Methods in molecular biology.
Springer, New York, NY, pp93-110, 2016.
|
|
11
|
Li J, Zhang Y, Gao Y, Cui Y, Liu H, Li M
and Tian Y: Downregulation of HNF1 homeobox B is associated with
drug resistance in ovarian cancer. Oncol Rep. 32:979–988.
2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Rasheed Z: Bioinformatics approach: A
powerful tool for microRNA research. Int J Health Sci (Qassim).
11:1–3. 2017.PubMed/NCBI
|
|
13
|
Dhumale SS, Waghela BN and Pathak C:
Quercetin protects necrotic insult and promotes apoptosis by
attenuating the expression of RAGE and its ligand HMGB1 in human
breast adenocarcinoma cells. IUBMB Life. 67:361–373.
2015.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Takakura Y, Hori N, Terada N, Machida M
and Yamaguchi N, Takano H and Yamaguchi N: VGLL3 activates
inflammatory responses by inducing interleukin-1α secretion. FASEB
J. 35(e21996)2021.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yan J, Ying S and Cai X: MicroRNA-mediated
regulation of HMGB1 in human hepatocellular carcinoma. Biomed Res
Int. 2018(2754941)2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Tanic M, Yanowski K, Gómez-López G,
Socorro Rodriguez-Pinilla M, Marquez-Rodas I, Osorio A, Pisano DG,
Martinez-Delgado B and Benítez J: MicroRNA expression signatures
for the prediction of BRCA1/2 mutation-associated hereditary breast
cancer in paraffin-embedded formalin-fixed breast tumors. Int J
Cancer. 136:593–602. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|