Introduction
Atherosclerosis is a major pathological process
related to a number of important adverse vascular events, including
coronary artery disease, stroke, and peripheral arterial disease
(1). Atherosclerosis is a chronic
inflammatory disease of the arterial wall, characterized by the
formation of lipid-laden lesions (2). Reducing the inflammatory reaction at
these sites may ameliorate atherosclerosis.
Peroxisome proliferator-activated receptors (PPARs)
are nuclear hormone receptors that act as ligand-activated
transcription factors to transactivate or transrepress target genes
via distinct mechanisms (3). Three
subtypes, PPAR-α, PPARβ/δ and PPARγ, have been identified and are
differentially expressed in distinct tissues. Numerous studies
provided evidence that peroxisome proliferator-activated receptor α
(PPAR-α), mainly expressed in high energy-requiring tissues such as
skeletal muscle, heart and liver (4), can limit inflammation and regulate
lipid metabolism (5,6). The beneficial effects of PPAR-α have
been highlighted in acute pancreatitis (7). However, the role of PPAR-α in
modulating inflammatory progression of atherosclerosis has been
rarely reported. A number of studies have suggested that PPAR-α
activation plays a pivotal role in regulating fatty acid oxidation,
lipid metabolism, vascular responses, and various metabolic and
intracellular signaling pathways that lead to microvascular
complications (8). As a general
modulator of the inflammatory response, PPAR-α was first reported
to be involved in a prolonged inflammatory response to leukotriene
B4 (LTB4) in a PPAR-α-null mice model (9). Moreover, increasing evidence suggests
that the effects of PPAR-α in inflammation are associated with
Alzheimer’s disease (AD) and Crohn’s disease (10,11).
Inflammation in atherosclerosis is characterized by a sequential
release of anti- and pro-inflammatory cytokines, including
interleukin-10 (IL-10), tumor necrosis factor-α (TNF-α) and
P-selectin (12–14). In this study, we investigated the
role of PPAR-α in atherosclerosis and in the induction of
expression of these inflammatory factors.
Materials and methods
Animal experiments
Adult male New Zealand White rabbits weighing
2.0–2.5 kg (n=25), were purchased from the Experimental Animal
Breeding Co. (Shanghai, China). Following a one-week adaptation
period, they were randomly divided into 5 groups of 5 rabbits each:
Control, high-fat diet + balloon injury, high-fat diet + balloon
injury + placebo, high-fat diet + balloon injury + fenofibrate,
high-fat diet + balloon injury + WY-14643. Rabbits were housed in a
temperature-, humidity- and light-controlled room with free access
to water. The control group was fed with the standard diet; the
high-fat + balloon injury group was fed with the high-fat diet
(cholesterol 2%, lard 10%, and standard diet 88%) and was subjected
to balloon injury. Rabbits in the placebo, fenofibrate and WY-14643
groups were subjected to balloon injury and then received placebo,
fenofibrate or WY-14643, respectively. Specifically, the placebo
group was fed with cholesterol diet and was intraperitoneally
injected with saline (1 mg/kg/day). The fenofibrate group was fed
with cholesterol diet containing fenofibrate pellets (130
mg/kg/day), as previously used by Jeanpierre et al (15). The WY-14643 group was fed with
cholesterol diet and received an intraperitoneal injection of
WY-14643 (1 mg/kg/day) (16). All
experiments were performed according to the guidelines of the
Experimental Animal Center of Shanghai First People’s Hospital
(SYXK, Shanghai, 2009–0086). The study was approved by the Ethics
Committee of Shanghai Jiaotong University (Shanghai, China)
Angioplasty
Rabbits (n=20) were left to adapt to the environment
for a week, were fed with high-fat diet for 4 weeks and were then
subjected to femoral artery balloon injury. Animals were
anesthetized with intravenous infusion of ketamine (0.5 ml/kg
Ketalar; Pfizer Inc., New York, NY, USA) and xylazine (0.25 ml/kg
Rompun, Bayer Healthcare, Whippany, NJ, USA). Femoral artery
de-endothelialization was induced using a 3F Fogarty®
balloon catheter (Baxter, Deerfield, IL, USA). Using ophthalmic
scissors, a ‘V’-shaped small hole was created in the artery wall,
followed by insertion of a balloon catheter (1:15 diluted heparin
saline infiltration) into the iliac artery (~15 cm). The balloon
was connected to a 20 ml injector and ~10 ml air (~2 atm) was
injected into the balloon. The balloon was slowly withdrawn from
the femoral artery, causing injury, and reinserted. This stretching
was repeated twice, 30 sec each time, with 1-min intervals to
ensure intimal injury. The catheter was removed, the vascular
proximal and distal ends were ligated, and the subcutaneous tissue
and the skin were sutured. The wound was washed with penicillin
sodium, with intramuscular injection of 400,000 units performed for
3 days.
Histological assessment of artery
damage
The artery was transversally sliced and fixed in 10%
formalin for 24 h prior to embedment in paraffin. Sections (3 mm)
were prepared for histological assessment by staining with
hematoxylin and eosin (H&E). The percentage of vessel wall
lumen occlusion was calculated with the following formula, as
previously described (17): 1 - [L
area ÷ (I + L area × 100)], where L denotes the lumen and I the
intima.
Immunohistochemistry
Paraffin sections of the artery were deparaffinized,
and the endogenous peroxidase activity was inactivated with 3%
H2O2 for 10 min. The primary antibody mouse
anti-rabbit PPAR-α (no. NB300–537; Novus Biologicals, Cambridge,
UK) or normal blocking serum was added and incubated overnight.
Biotin-conjugated goat anti-mouse immunoglobulin G (IgG) (Novus
Biologicals) was used as the secondary antibody and incubated for
30 min. An avidin-biotin enzyme reagent (Novus Biologicals) was
sequentially added and incubated for 20 min. A peroxidase substrate
was added and incubated until the desired stain intensity was
reached. Finally, sections were covered with a glass coverslip and
observed under a light microscope. The intensity of positive
staining in the tissues was analyzed by integrated optical density
(IOD) using Image-Pro Plus software (Media Cybernetics, Rockville,
MD, USA). Briefly, four 20X TIF-format images from five individual
rabbits in each group were analyzed. The IOD and area were
calculated, as well as the lumen and the internal elastic lamina
area. The PPAR-α expression was expressed as [(IOD/area) × 100] in
accordance with a previous study (18).
Western blot analysis
These vascular tissue samples were washed with
ice-cold phosphate-buffered saline, and lysed for 20 min on ice
with the lysis buffer. The lysates were centrifuged for 4 min at
12,000 × g, and the supernatant was collected in a fresh tube kept
on ice. Protein concentrations in each sample were determined using
the bicinchoninic acid (BCA) assay. One hundred micrograms of total
protein were mixed with loading buffer with the anionic denaturing
detergent sodium dodecyl sulfate (SDS), were boiled for 5 min, and
then resolved by 10% SDS polyacrylamide gel electrophoresis. The
proteins were transferred onto a polyvinylidene difluoride (PVDF)
membrane. After blocking the membrane in Tris-buffered saline with
Tween 20 (TBST) containing non-fat milk for 1 h at 4°C under
agitation, the membrane was washed three times in TBST and
incubated for 2 h with mouse anti-rabbit PPAR-α antibody (1:200
dilution) or GAPDH monoclonal antibody (1:200 dilution, no. 20028;
Abmart, Shanghai, China). After washing three times in TBST, the
membrane was incubated with HRP-conjugated goat anti-mouse IgG
(1:1,000) for 1 h and then washed three times with TBST.
Immuno-stained proteins were detected using a streptavidin
amplification reagent (no. WBKL SOO 50; Millipore, Billerica, MA,
USA) according to the manufacturer’s instructions.
RNA extraction and quantitative PCR
Total RNA was extracted from arterial samples using
the TRIzol® reagent according to the manufacturer’s
instructions (Invitrogen Life Technologies, Carlsbad, CA, USA). One
microgram of total RNA was used as template to synthesize cDNA
using a reverse transcription kit from BioDev-Tech Co., Ltd
(Beijing, China). Quantitative PCR was performed by monitoring the
increase in fluorescence of the SYBR-Green dye using the
GreenMaster mix (Genaxxon BioScience GmbH, Ulm, Germany) according
to the manufacturer’s instructions. The primer sets used to amplify
the PPAR-α gene were: 5′-gttccggtggcgttgat-3′ (forward) and
5′-gcggtcgcatttgtc-3′ (reverse). The primer sets used to amplify
GAPDH were 5′-ccactttgtgaagctcatttcct-3′ (forward) and
5′-tcgtcctcctctggtgctct-3′ (reverse). PCR amplification was
performed for 32 cycles using a Taq polymerase for the
reverse transcription kit, with the following program: 95°C for 45
sec, 62°C for 30 sec, and 72°C for 1 min. The 2−ΔΔCt
method was used to quantify the expression of the PPAR-α and
GAPDH genes. The resulting values were used to express the
relative quantity (RQ) of PPAR-α with regard to that of
GAPDH.
Enzyme-linked immunosorbent assay (ELISA)
for TNF-α, IL-10 and P-selectin
Fresh blood (3 ml) was extracted from all animals
via the femoral vein and centrifuged at 3,000 × g for 10 min at
4°C. The supernatant was transferred in a clean centrifuge tube,
and frozen at −20°C. Concentrations of plasma TNF-α, IL-10 and
P-selectin were assayed with ELISA kits (R&D Systems Inc.,
Minneapolis, MN, USA) according to the manufacturer’s
instructions.
Statistical analysis
Data were analyzed using the SPSS 11.5 software for
Windows (IBM, Armonk, NY, USA). Quantitative data were expressed as
mean ± standard deviation. Comparisons between multiple groups were
conducted with analysis of variance, and pairwise comparisons with
the Student-Newman-Keuls test. P<0.05 was considered to indicate
statistically significant differences.
Results
Histological changes
H&E-stained sections of the aorta were examined
for signs of atheroma. High-fat diet and balloon injury induced a
notable atherosclerotic lesion, whereas animals receiving standard
diet had no visible atheroma. In the high-fat diet + balloon injury
group, the lesion site showed foam cell formation, accumulation of
smooth muscle cells in the intima, and fragmentation of the
internal elastic lamina of the femoral artery. In the high-fat diet
+ balloon injury + placebo group, the lesion site showed smooth
muscle cell migration, foam cell formation, and a lipid core in the
intima. In the high-fat diet + balloon injury +
fenofibrate/WY-14643 groups, the lesion also showed smooth muscle
cell migration, foam cell formation, and a lipid core in the
intima. The percentage of vessel lumen occlusion in the fenofibrate
and WY-14643 groups was significantly decreased compared to the
placebo group (Fig. 1).
The percentage of vessel wall lumen occlusion was
calculated (Fig. 2). No obviously
occluded lumen was observed in the control group, while the
high-fat diet + balloon injury and high-fat diet + balloon injury +
placebo groups showed lumen occlusion. Both fenofibrate and
WY-14643 reduced the percentage of vessel wall lumen occlusion
compared to the placebo group. There was no difference in the
percentage of vessel wall lumen occlusion between the high-fat diet
+ balloon injury + fenofibrate and high-fat diet + balloon injury +
WY-14643 groups.
Localization of PPAR-α protein
Immunohistochemical examination showed that the
PPAR-α protein was rarely detected in the cytoplasm of the control
group (Fig. 3). By contrast,
PPAR-α was found at high staining intensities in the cytoplasm of
macrophages, especially near the intima, in the high-fat diet +
balloon injury, high-fat diet + balloon injury + placebo, high-fat
diet + balloon injury + fenofibrate and high-fat diet +
balloon-injury + WY-14643 groups (Fig.
3). Compared to the placebo group, the staining intensity of
PPAR-α was higher in the fenofibrate and WY-14643 groups.
Expression of PPAR-α protein
Western blot analysis was performed to determine the
protein level of the PPAR-α protein in femoral artery tissues.
Western blot bands were quantified by densitometry. Compared to
standard treatment, high-fat diet and balloon injury increased the
PPAR-α level (Fig. 4).
Pretreatment with fenofibrate and WY-14643 significantly increased
the expression of the protein.
Expression of the PPAR-α gene
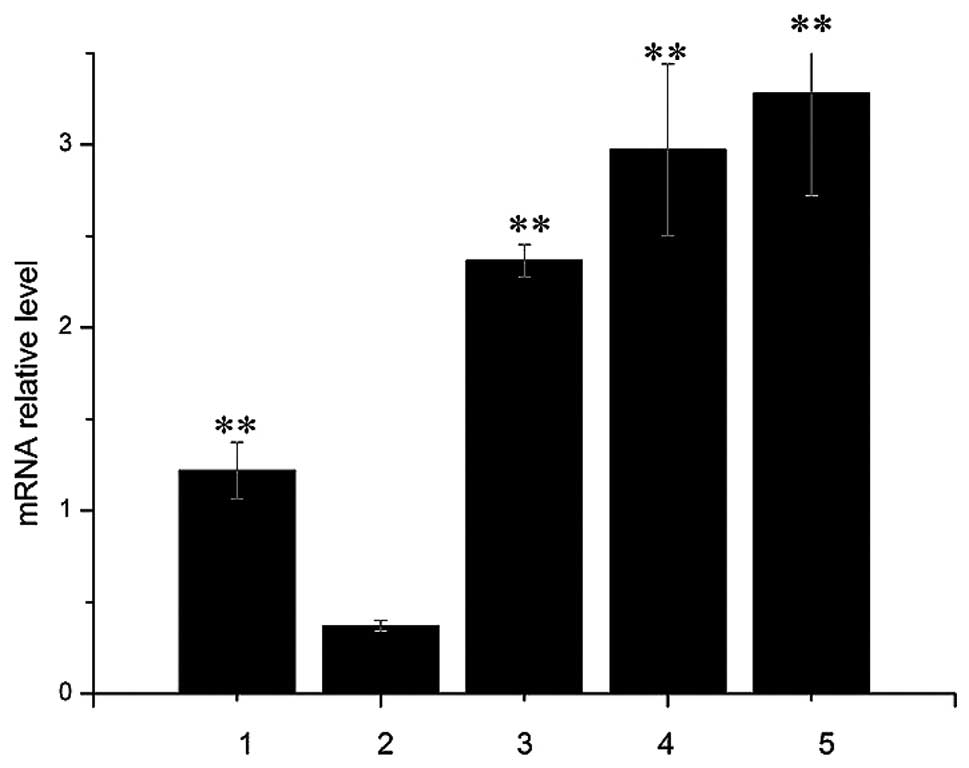
The mRNA level of PPAR-α was analyzed by
quantitative PCR. Compared to the control group, high-fat diet +
balloon injury increased PPAR-α expression (Fig. 5), and pretreatment with fenofibrate
or WY-14643 significantly increased PPAR-α expression.
IL-10, TNF-α and P-selectin levels in the
serum
The serum concentrations of IL-10, TNF-α and
P-selectin were measured in order to assess the degree of
inflammatory response. Compared to the control group, IL-10, TNF-α
and P-selectin levels were all significantly increased in the
high-fat diet + balloon injury group (Fig. 6). Fenofibriate or WY-14643
treatment markedly reduced the TNF-α and P-selectin level, while it
did not change the IL-10 level induced by high-fat diet and balloon
injury.
Discussion
Atherosclerosis is a major pathological process
related to a number of important adverse vascular events, including
coronary artery disease, stroke, and peripheral arterial disease
(19). However, the exact
mechanisms underlying this condition remain elusive. Inflammation
is an important step in the progression of atherosclerosis. The
prevention of inflammation is expected to delay the progression of
atherosclerosis. PPAR-α is considered an inhibitor of inflammation
during atherosclerosis, although the relevant mechanism is still
unclear. In this study, we investigated the role of PPAR-α in
atherosclerosis and in the induction of the expression of
inflammation factors. We investigated the protective effects of
PPAR-α activation by fenofibrate and WY-14643 in a high-fat diet
and balloon injury-induced atherosclerotic model. High-fat diet and
balloon injury induce the formation of vascular lesions, where
inflammatory cytokines are activated. The harmful effects can be
inhibited by fenofibrate or WY-14643 through activation of
PPAR-α.
PPAR-α, as a member of a nuclear receptor family,
plays an important role in regulating fatty acid oxidation, lipid
and lipoprotein metabolism, and inflammatory and vascular
responses, all of which are involved in atherosclerosis (5,20).
The expression of inflammatory factors regulated by PPAR-α
correlates to prostate cancer and benign prostatic hyperplasia
(21). Expression of PPAR-α was
also increased in our atherosclerotic model, in both the high-fat
diet and the placebo groups compared to the control. Furthermore,
PPAR-α levels in the high-fat diet group were lower than those in
the control group. It was previously shown that overexpression of
VEGF and TGF-β1 correlates with the progression of cervical
intraepithelial neoplasia to cancer (22,23).
Therefore, in the high-fat diet and balloon injury model, PPAR-α
may be activated by lipoprotein lipolytic products, without
mediation of exogenous agonists. These data indicate that PPAR-α
associates with the progression of atherosclerosis, which could
constitute a self-regulatory mechanism to prevent atherosclerotic
lesion formation. Fenofibrate and WY-14643 are effective agonists
of PPAR-α (24). Our results
showed that fenofibrate and WY-14643 can enhance the
anti-inflammatory response in the atherosclerotic model induced by
high-fat diet and balloon injury. Notably, fenofibrate and WY-14643
minimized the vascular lesions induced by high-fat diet and balloon
injury. The percentage of vessel wall lumen occlusion in the
fenofibrate and the WY-14643 groups was considerably lower compared
to that of the placebo group, and fenofibrate and WY-14643 induced
an increase in the expression of PPAR-α. A previous study
demonstrated that PPAR-α can inhibit vascular wall inflammation
(6). Thus, PPAR-α agonists may
modulate atherosclerosis by inhibiting the inflammatory
response.
The major role played by the inflammatory response
in the progression of atherosclerosis is an emerging concept
(25). Activation of PPAR-α
favorably modulates inflammation markers. IL-10 is an
anti-inflammatory cytokine produced by a variety of immune cells
with anti-atherogenic potential (26). By contrast, TNF-α and P-selectin
are pro-inflammatory cytokines, involved in atherosclerosis and
other metabolic and inflammatory conditions such as obesity and
insulin resistance, which are additional risk factors for the
development of cardiovascular diseases (27,28).
Previous studies indicated that PPAR-α significantly reduces the
level of pro-inflammatory cytokines such as TNF-α and P-selectin in
ischemia-reperfusion injury (28,29),
and enhances the expression of IL-10 in acute pancreatitis
(7). As important inflammatory
cytokines, IL-10, TNF-α and P-selectin are also involved in the
development of atherosclerosis (30,31).
Based on these observations, we aimed to determine whether the
presence and/or the stimulation of PPAR-α can enhance the
anti-inflammatory response in atherosclerosis, by measuring the
serum levels of IL-10, TNF-α and P-selectin. Fenofibrate and
WY-14643 induced the anti-inflammatory response in our experimental
model of atherosclerosis. TNF-α and P-selectin levels were
increased in the placebo group and were reduced by treatment with
fenofibrate or WY-14643, together with reduced vascular lesions.
The IL-10 level was higher in the fenofibrate and WY-14643 groups
compared to the placebo group. These results are in agreement with
previous studies indicating that increased expression of the
IL-10 gene might be relevant to a series of coronary and
cerebrovascular events, and that low capacity to produce IL-10
entails an increased risk for stroke incidents (32,33).
It was also shown that the increased concentration of IL-10 is
associated with an increased risk for cardiovascular events
(34). Taken together, these
results highlight the complexity of roles played by pro- and
anti-inflammatory molecules during the process of atherosclerosis.
In parallel with elevated levels of PPAR-α in the high-fat diet and
placebo groups, the increased expression of IL-10 may control the
inflammatory response as a self-protective mechanism.
In conclusion, activated PPAR-α can prevent vascular
lesions induced by high-fat diet and balloon injury in rabbits. Our
findings suggest that the anti-atherosclerotic and
anti-inflammatory effects of PPAR-α are associated with the
increased expression of IL-10 and the reduced expression of TNF-α
and P-selectin. Notably, IL-10, TNF-α and P-selectin, as well as
PPAR-α expression were increased in the high-fat diet and the
placebo groups compared with the control group. These results
suggest that a mild activation of PPAR-α signaling may occur in
atherosclerosis without the presence of exogenous PPAR-α agonists,
but it may not be sufficient to prevent the progression of
atherosclerosis. To fully address the role(s) of PPAR-α in
atherosclerosis, experiments using a PPAR-α inhibitor need to be
carried out in the future.
Acknowledgements
The study was supported by the Shanghai Rising-Star
Program (08QA1404100 grant) and the National Natural Science
Foundation of China (30971265).
References
|
1
|
Ostrom MP, Gopal A, Ahmadi N, Nasir K,
Yang E, Kakadiaris I, Flores F, Mao SS and Budoff MJ: Mortality
incidence and the severity of coronary atherosclerosis assessed by
computed tomography angiography. J Am Coll Cardiol. 52:1335–1343.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mizuno Y, Jacob RF and Mason R:
Inflammation and the development of atherosclerosis. J Atheroscler
Thromb. 18:351–358. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Motojima K: Peroxisome
proliferator-activated receptor (PPAR): structure, mechanisms of
activation and diverse functions. Cell Struct Funct. 18:267–277.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hiukka A, Maranghi M, Matikainen N and
Taskinen MR: PPARα: an emerging therapeutic target in diabetic
microvascular damage. Nat Rev Endocrinol. 6:454–463. 2010.
|
|
5
|
Wilson JL, Duan R, El-Marakby A, Alhashim
A and Lee DL: Peroxisome proliferator activated receptor-α agonist
slows the progression of hypertension, attenuates plasma
interleukin-6 levels and renal inflammatory markers in angiotensin
II infused mice. PPAR Res. 2012:6459692012.
|
|
6
|
Fruchart JC, Duriez P and Staels B:
Peroxisome proliferator-activated receptor-alpha activators
regulate genes governing lipoprotein metabolism, vascular
inflammation and atherosclerosis. Curr Opin Lipidol. 10:245–257.
1999. View Article : Google Scholar
|
|
7
|
Ding JL, Zhou ZG, Zhou XY, Zhou B, Wang L,
Wang R, Zhan L, Sun XF and Li Y: Attenuation of acute pancreatitis
by peroxisome proliferator-activated receptor-α in rats: the effect
on Toll-like receptor signaling pathways. Pancreas. 42:114–122.
2013.
|
|
8
|
Desvergne B and Wahli W: Peroxisome
proliferator-activated receptors: nuclear control of metabolism.
Endocr Rev. 20:649–688. 1999.PubMed/NCBI
|
|
9
|
Devchand PR, Keller H, Peters JM, Vazquez
M, Gonzalez FJ and Wahli W: The PPARalpha-leukotriene B4 pathway to
inflammation control. Nature. 384:39–43. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Heun R, Kölsch H, Ibrahim-Verbaas CA,
Combarros O, Aulchenko YS, Breteler M, Schuur M, van Duijn CM,
Hammond N, Belbin O, Cortina-Borja M, Wilcock GK, Brown K, Barber
R, Kehoe PG, Coto E, Alvarez V, Lehmann MG, Deloukas P, Mateo I,
Morgan K, Warden DR, Smith AD and Lehmann DJ: Interactions between
PPAR-α and inflammation-related cytokine genes on the development
of Alzheimer’s disease, observed by the epistasis project. Int J
Mol Epidemiol Genet. 3:39–47. 2012.
|
|
11
|
Lee JW, Bajwa PJ, Carson MJ, Jeske DR,
Cong Y, Elson CO, Lytle C and Straus DS: Fenofibrate represses
interleukin-17 and interferon-γ expression and improves colitis in
interleukin-10-deficient mice. Gastroenterology. 133:108–123.
2007.PubMed/NCBI
|
|
12
|
Liu Y, Jiang H, Liu W, Shang H, Tang Y,
Zhu R and Li B: Effects of fluvastatin therapy on serum
interleukin-18 and interleukin-10 levels in patients with acute
coronary syndrome. Acta Cardiol. 65:285–289. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Miller AM, McPhaden AR, Preston A,
Wadsworth RM and Wainwright CL: TNFα increases the inflammatory
response to vascular balloon injury without accelerating neointimal
formation. Atherosclerosis. 179:51–59. 2005.
|
|
14
|
Mobarrez F, He S, Bröijersen A, Wiklund B,
Antovic A, Antovic J, Egberg N, Jörneskog G and Wallén H:
Atorvastatin reduces thrombin generation and expression of tissue
factor, P-selectin and GPIIIa on platelet-derived microparticles in
patients with peripheral arterial occlusive disease. Thromb
Haemost. 106:344–352. 2011. View Article : Google Scholar
|
|
15
|
Jeanpierre E, Le Tourneau T, Zawadzki C,
Van Belle E, Mouquet F, Susen S, Ezekowitz MD, Staels B, Jude B and
Corseaux D: Beneficial effects of fenofibrate on plaque
thrombogenicity and plaque stability in atherosclerotic rabbits.
Cardiovasc Pathol. 18:140–147. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yeh CH, Chen TP, Lee CH, Wu YC, Lin YM and
Lin PJ: Cardiomyocytic apoptosis following global cardiac ischemia
and reperfusion can be attenuated by peroxisome
proliferator-activated receptor alpha but not gamma activators.
Shock. 26:262–270. 2006. View Article : Google Scholar
|
|
17
|
Buchanan MR and Brister SJ: Inhibition of
chronic vessel wall intimal hyperplasia following acute
anticoagulant treatment: Relative effects of heparin and dermatan
sulphate. Thromb Res. 91:157–167. 1998. View Article : Google Scholar
|
|
18
|
Jia XL, Li SY, Dang SS, Cheng YA, Zhang X,
Wang WJ, Hughes CE and Caterson B: Increased expression of
chondroitin sulphate proteoglycans in rat hepatocellular carcinoma
tissues. World J Gastroenterol. 18:3962–3976. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Libby P, Ridker PM and Hansson GK:
Progress and challenges in translating the biology of
atherosclerosis. Nature. 473:317–325. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chamouton J and Latruffe N: PPARα/HNF4α
interplay on diversified responsive elements. Relevance in the
regulation of liver peroxisomal fatty acid catabolism. Curr Drug
Metab. 13:1436–1453. 2012.
|
|
21
|
Soulitzis N, Karyotis I, Delakas D and
Spandidos DA: Expression analysis of peptide growth factors VEGF,
FGF2, TGFB1, EGF and IGF1 in prostate cancer and benign prostatic
hyperplasia. Int J Oncol. 29:305–314. 2006.PubMed/NCBI
|
|
22
|
Baritaki S, Sifakis S, Huerta-Yepez S,
Neonakis IK, Soufla G, Bonavida B and Spandidos DA: Overexpression
of VEGF and TGF-β1 mRNA in Pap smears correlates with progression
of cervical intraepithelial neoplasia to cancer: implication of YY1
in cervical tumorigenesis and HPV infection. Int J Oncol. 31:69–79.
2007.
|
|
23
|
Arvanitis DA and Spandidos DA:
Deregulation of the G1/S phase transition in cancer and squamous
intraepithelial lesions of the uterine cervix: A case control
study. Oncol Rep. 20:751–760. 2008.PubMed/NCBI
|
|
24
|
Gervois P, Chopin-Delannoy S, Fadel A,
Dubois G, Kosykh V, Fruchart JC, Najïb J, Laudet V and Staels B:
Fibrates increase human REV-ERBα expression in liver via a novel
peroxisome proliferator-activated receptor response element. Mol
Endocrinol. 13:400–409. 1999.
|
|
25
|
Stoll G and Bendszus M: Inflammation and
atherosclerosis novel insights into plaque formation and
destabilization. Stroke. 37:1923–1932. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu Y, Li D, Chen J, Xie J, Bandyopadhyay
S, Zhang D, Nemarkommula AR, Liu H, Mehta JL and Hermonat PL:
Inhibition of atherogenesis in LDLR knockout mice by systemic
delivery of adeno-associated virus type 2-hIL-10. Atherosclerosis.
188:19–27. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kleemann R, Zadelaar S and Kooistra T:
Cytokines and atherosclerosis: a comprehensive review of studies in
mice. Cardiovasc Res. 79:360–376. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Di Paola R, Impellizzeri D, Torre A,
Mazzon E, Cappellani A, Faggio C, Esposito E, Trischitta F and
Cuzzocrea S: Effects of palmitoylethanolamide on intestinal injury
and inflammation caused by ischemia-reperfusion in mice. J Leukoc
Biol. 91:911–920. 2012.PubMed/NCBI
|
|
29
|
Standage SW, Caldwell CC, Zingarelli B and
Wong HR: Reduced peroxisome proliferator-activated receptor α
expression is associated with decreased survival and increased
tissue bacterial load in sepsis. Shock. 37:164–169. 2012.
|
|
30
|
Salagianni M, Galani IE, Lundberg AM,
Davos CH, Varela A, Gavriil A, Lyytikäinen LP, Lehtimäki T, Sigala
F, Folkersen L, Gorgoulis V, Lenglet S, Montecucco F, Mach F, Hedin
U, Hansson GK, Monaco C and Andreakos E: Toll-like receptor 7
protects from atherosclerosis by constraining ‘inflammatory’
macrophage activation. Circulation. 126:952–962. 2012.PubMed/NCBI
|
|
31
|
Zhou B, Pan Y, Hu Z, Wang X, Han J, Zhou
Q, Zhai Z and Wang Y: All-trans-retinoic acid ameliorated high fat
diet-induced atherosclerosis in rabbits by inhibiting platelet
activation and inflammation. J Biomed Biotechnol. 2012:2596932012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Trompet S, Pons D, De Craen AJ, Slagboom
P, Shepherd J, Blauw GJ, Murphy MB, Cobbe SM, Bollen EL, Buckley
BM, Ford I, Hyland M, Gaw A, Macfarlane PW, Packard CJ, Norrie J,
Perry IJ, Stott DJ, Sweeney BJ, Twomey C, Westendorp RG and Jukema
JW: Genetic variation in the interleukin-10 gene promoter and risk
of coronary and cerebrovascular events: the PROSPER study. Ann NY
Acad Sci. 1100:189–198. 2007. View Article : Google Scholar
|
|
33
|
van Exel E, Gussekloo J, de Craen A,
Bootsma-van der Wiel A, Frölich M and Westendorp R: Inflammation
and stroke: the Leiden 85-Plus Study. Stroke. 33:1135–1138.
2002.PubMed/NCBI
|
|
34
|
Lakoski SG, Liu Y, Brosnihan KB and
Herrington DM: Interleukin-10 concentration and coronary heart
disease (CHD) event risk in the estrogen replacement and
atherosclerosis (ERA) study. Atherosclerosis. 197:443–447. 2008.
View Article : Google Scholar : PubMed/NCBI
|