Introduction
Alcoholic liver disease (ALD) remains an important
cause of morbidity and mortality, with >75,000 annual deaths
worldwide and incidence rates having increased in the last decade
(1,2). ALD encompasses a spectrum of
pathological states including fatty liver, inflammation, fibrosis,
cirrhosis, and even malignancy (3,4).
Clinical and experimental data have demonstrated that gut-derived
endotoxins and endotoxemia play a major role in the development of
ALD (5,6).
Ethanol can affect the intestinal barrier in
multiple ways, leading to an increased portal blood level of
endotoxin and hepatic exposure, thereby further promoting liver
inflammation and the progression to ALD (7). Numerous studies have indicated that
ethanol induces intestinal permeability in vivo via
oxidative stress (7,8). Antioxidants have been shown to
normalize intestinal permeability in alcoholic patients (9). It is now generally accepted that
impaired intestinal epithelial integrity and intestinal barrier
dysfunctions are the two fundamental causes of increased intestinal
permeability (10,11). The intestinal epithelial barrier is
a complex system composed of cellular, physical, and chemical
components (12). The epithelial
cells form a layer with the paracellular space sealed by tight
junctions (TJs) and adherens junctions (AJs). TJs are essential in
maintaining intestinal mucosal integrity, which can effectively
prevent bacteria, endotoxins and other harmful substances from
entering into the blood stream through the intestinal barrier
(13). Two studies indicated that
altered expression of the TJ-associated proteins zonula occludens-1
(ZO-l) and claudin-1 associates with increased intestinal
permeability and higher susceptibility to ALD (14,15).
Most studies to date have addressed the potential effects of
ethanol on paracellular permeability and disruption of epithelial
TJs in a cell culture model of intestinal epithelium, the Caco-2
cell monolayer (16–18).
The Rho-associated protein kinase (ROCK) is a
serine-threonine kinase that acts as a key downstream effector of
RhoA signaling, and was shown to contribute in the maintenance of
TJ integrity in endothelial cells (19) and in human intestinal epithelial
cells (20). In this study, we
investigated the hypothesis that ROCK plays a key role in the
ethanol-induced increase in permeability of the intestinal
epithelial barrier (IEB), and investigated the molecular links
between TJ-associated proteins, IEB permeability and ROCK.
Materials and methods
Caco-2 cells culture and establishment of
an in vitro IEB model
The human colon adenocarcinoma cell line Caco-2 was
obtained from the American Type Culture Collection (Manassas, VA,
USA). Caco-2 cells were cultured in HyClone™ Dulbecco’s modified
Eagle’s medium (DMEM) purchased from Thermo Fisher Scientific
(Waltham, MA, USA) and supplemented with 4.5 mg/ml glucose, 50 U/ml
penicillin, 50 U/ml streptomycin, 4 mM glutamine, and 10% HyClone™
fetal calf serum (FCS), in a humidified atmosphere (95% air, 5%
CO2) at 37°C. To establish an in vitro intestinal
epithelial barrier model, Caco-2 cells were plated onto Transwell
filters (Corning, New York, NY, USA) and regularly monitored by
visual inspections under an inverted microscope and by measurements
of epithelial resistance.
Assessment of intestinal epithelial
barrier permeability
Measurements of transepithelial electrical
resistance (TEER) and Lucifer yellow (LY) flux were performed to
assess the permeability of IEB. The electrical resistance of Caco-2
cell monolayers cultured on Transwell filters was measured using a
Millicell®-ERS instrument (Millipore, Bedford, MA, USA).
Electrical resistance of Transwell insert was expressed in units of
ohm (Ω)/cm2. The Lucifer yellow fluorescent dye
(Sigma-Aldrich, St. Louis, MO, USA) was added to the upper chamber
of the Transwell system at 40 μg/ml in serum-free DMEM. Following
treatment with ethanol for different time periods, the medium from
the lower chamber was collected. The absorbance of the medium was
measured on a fluorescence spectrophotometer (excitation wavelength
427 nm, emission wavelength 536 nm), and the LY concentration was
calculated based on a standard curve. The LY flux rate (%) = LY
concentration in the lower chamber/LY concentration in the upper
chamber.
Drug treatment and experimental
groups
First, Caco-2 cells were plated onto the lower side
of the Transwell insert and were grown in DMEM. When 80% of Caco-2
cells were confluent (at ~21–23 days of culture), they were seeded
on the upper chamber of the Transwell insert, and ethanol was
added. The cultured cells were then rinsed with cold
phosphate-buffered saline (PBS). Subsequently, Caco-2 cells from
the upper chamber were harvested by gently scraping with a cell
scraper and were stored at −80°C. We previously showed that ethanol
treatment causes a concentration- and time-dependent decrease in
Caco-2 transepithelial resistance and an increase in the
transepithelial permeability to the paracellular marker LY. We also
demonstrated that IEB permeability reaches a peak at 60 min of
treatment with 5% ethanol (21).
Based on these findings, the present experimental plan included 6
groups (n=4/group): the control (monolayer of Caco-2 cells), the
ethanol (5%) 0-min group, the ethanol (5%) 15-min group, the
ethanol (5%) 30-min group, the ethanol (5%) 60-min group, and the
ethanol (5%) 60-min + Y-27632 group, where Caco-2 cells were
pretreated with 10 μM Y-27632 for 60 min, then treated with ethanol
for 60 min.
Western blot analysis
The Caco-2 cells were washed three times with
Dulbecco’s phosphate-buffered saline (D-PBS) containing 0.1 mM
ethylenediamine tetraacetic acid (EDTA) without calcium and
magnesium; then, they were homogenized in 1 ml lysis buffer A [2 mM
EDTA, 10 mM ethylene glycol tetraacetic acid (EGTA), 0.4% sodium
fluoride (NaF), 20 mM Tris-HCL, protease inhibitor cocktail,
phosphatase inhibitor 1% Triton X-100, pH 7.5] at 4°C. Samples were
centrifuged at 14,000 × g for 30 min, and the supernatant was
transferred to a separate tube and collected as the soluble
fraction (S). Buffer A (150 μl) with 1% sodium dodecyl sulfate
(SDS) at 4°C was then added to the pellet. The pellet was disrupted
with an ultrasonic crusher. Samples were then centrifuged at 14,000
× g for 30 min at 4°C as described above. The supernatant was
collected as the insoluble fraction (IS). Equal amounts of proteins
(40–50 μg) were separated by SDS-polyacrylamide gel electrophoresis
(PAGE), and immunoblotting was performed by incubating with
antibodies targeting ZO-1 or claudin-1 (both at 1:1,000 dilution;
Santa Cruz Biotechnology. Inc., Santa Cruz, CA, USA). The protein
bands were visualized and quantified using the ChemiImager 5500
version 2.03 software (AlPha InnCh, Miami, FL, USA). The integrated
density values (IDV) for each protein were calculated using the
computerized image analysis system Fluor Chen 2.0 (Bio-Rad,
Hercules, CA, USA) and were normalized to the IDV of β-actin, used
as the loading control.
Immunofluorescence
The Caco-2 cell monolayers grown on glass coverslips
were fixed in 4% paraformaldehyde and permeabilized with 0.5%
Triton X-100. Following blocking with 2% BSA in PBS, the cells were
incubated with rabbit anti-ZO-1 (1:50 dilution) and -claudin-1
antibodies (1:100 dilution; both from Santa Cruz Biotechnology,
Inc.) in order to visualize the distribution of ZO-1 and claudin-1
proteins. The glass slides were analyzed using immunofluorescence
microscopy (Olympus, Japan).
Statistical analysis
Experiments were repeated at least three times. Data
are expressed as mean ± standard deviation (SD). Differences
between the groups were analyzed using one-way analysis of variance
(ANOVA) followed by Dunnett’s tests. All statistical tests were
two-sided, with P<0.05 considered to indicate statistically
significant differences.
Results
Effect of Y-27632 on ethanol-induced IEB
permeability
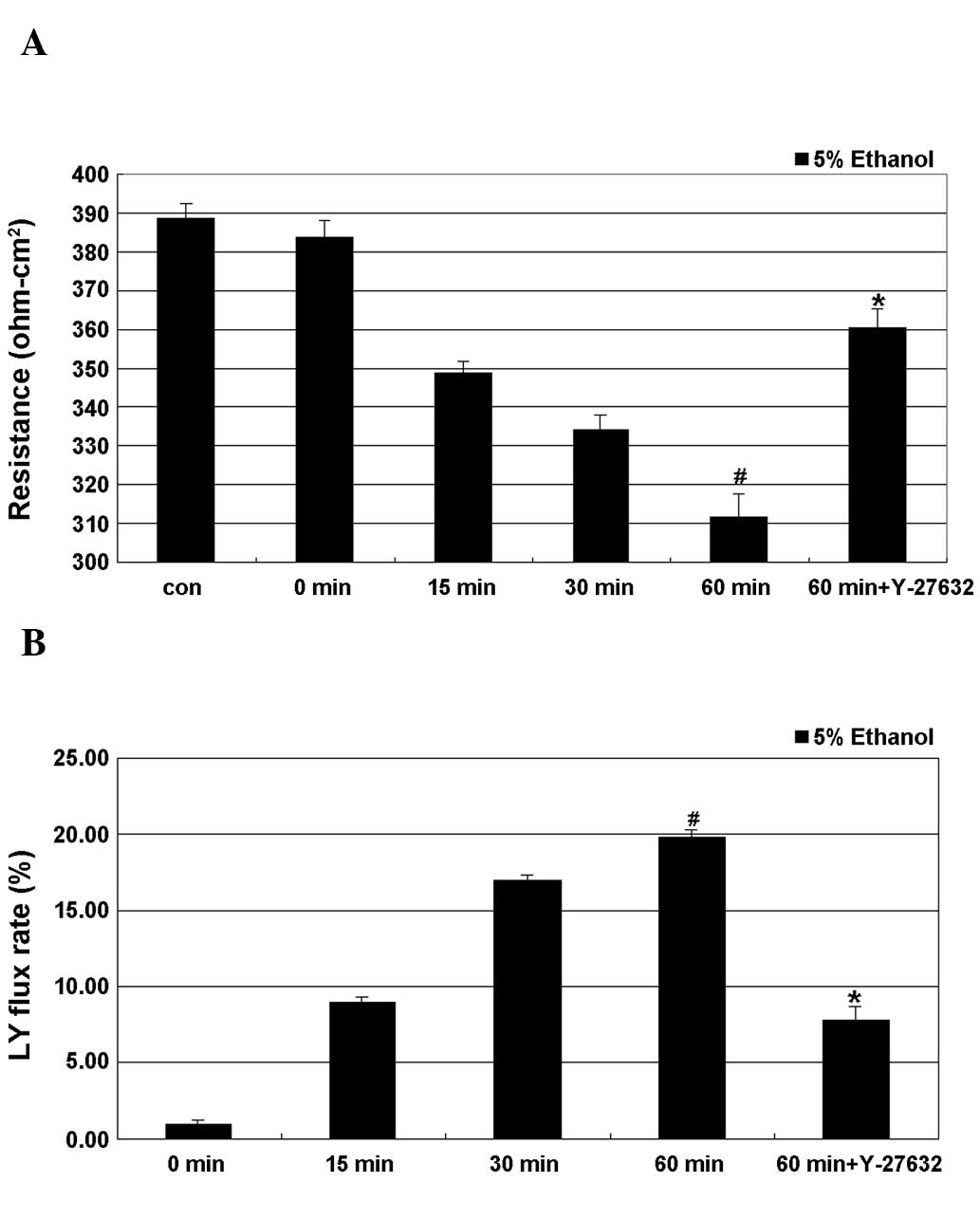
As shown in Fig. 1,
treatment of cells with 5% ethanol induced a time-dependent
decrease in TEER, with the lowest value obtained after 60 min of
treatment, where this decrease was significant. A time-dependent
increase in the LY flux rate as a result of ethanol treatment was
also observed, with the maximum rate observed at 60 min of
treatment, where the difference to the control cells was also
significant. Y-27632 partially and significantly inhibited
ethanol-induced epithelial leakage by restoring the TEER values and
the LY flux rate of the IEB.
The Y-27632 inhibitor prevents the
ethanol-induced shift from the insoluble to the soluble fraction of
claudin-1
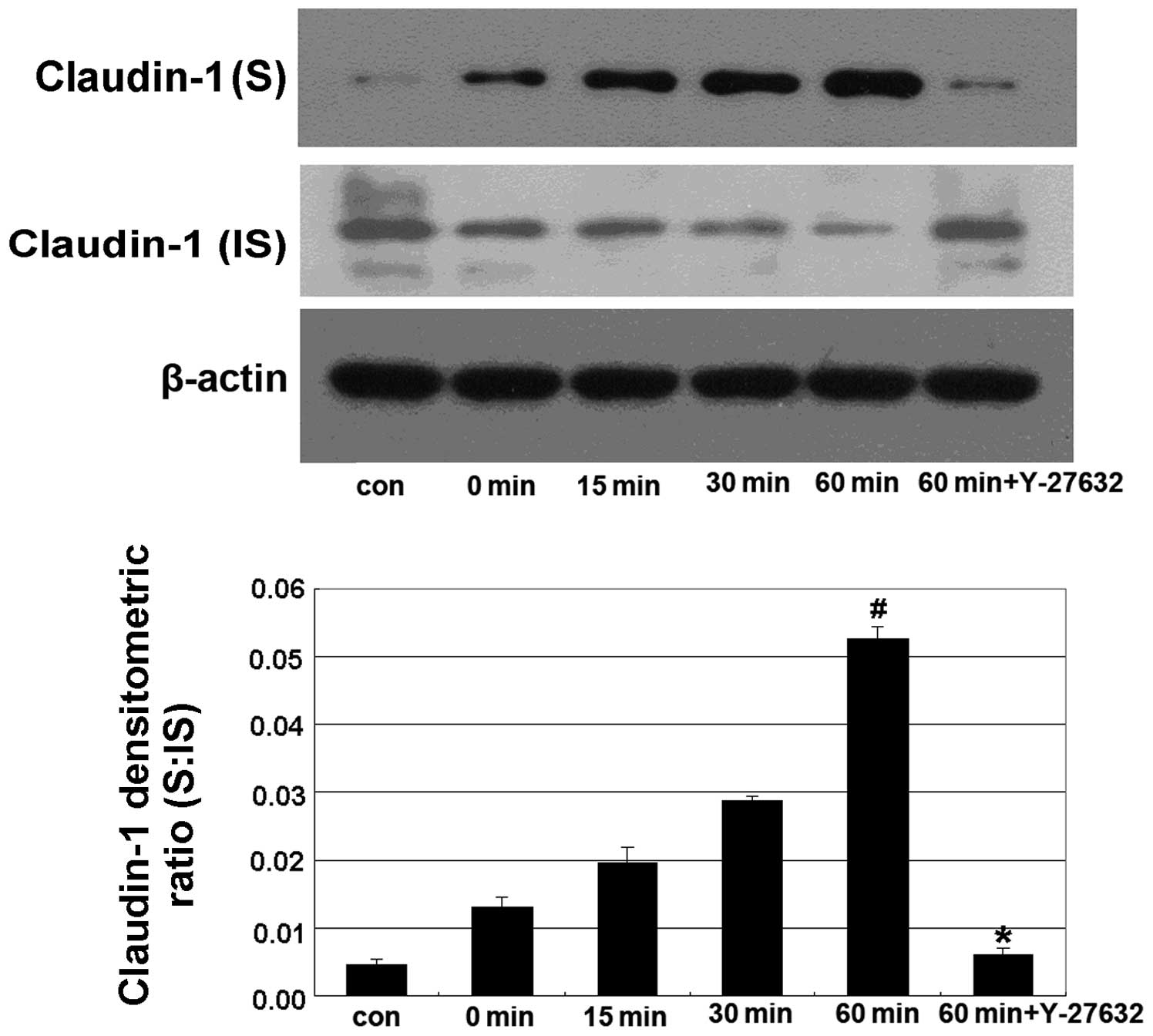
In this experiment, Caco-2 cells were treated with
5% ethanol for 0, 15, 30 and 60 min in the alcohol-treated groups,
and were pretreated with 10 μm Y-27632 for 60 min prior to
treatment with 5% ethanol for 60 min in the Y-27632 group. We found
that treatment with 5% ethanol induced a shift in the distribution
of claudin-1 from the IS to the S fraction, while Y-27632
significantly reverted this shift (Fig. 2).
The Y-27632 inhibitor prevents the
ethanol-induced decrease in the expression of ZO-1
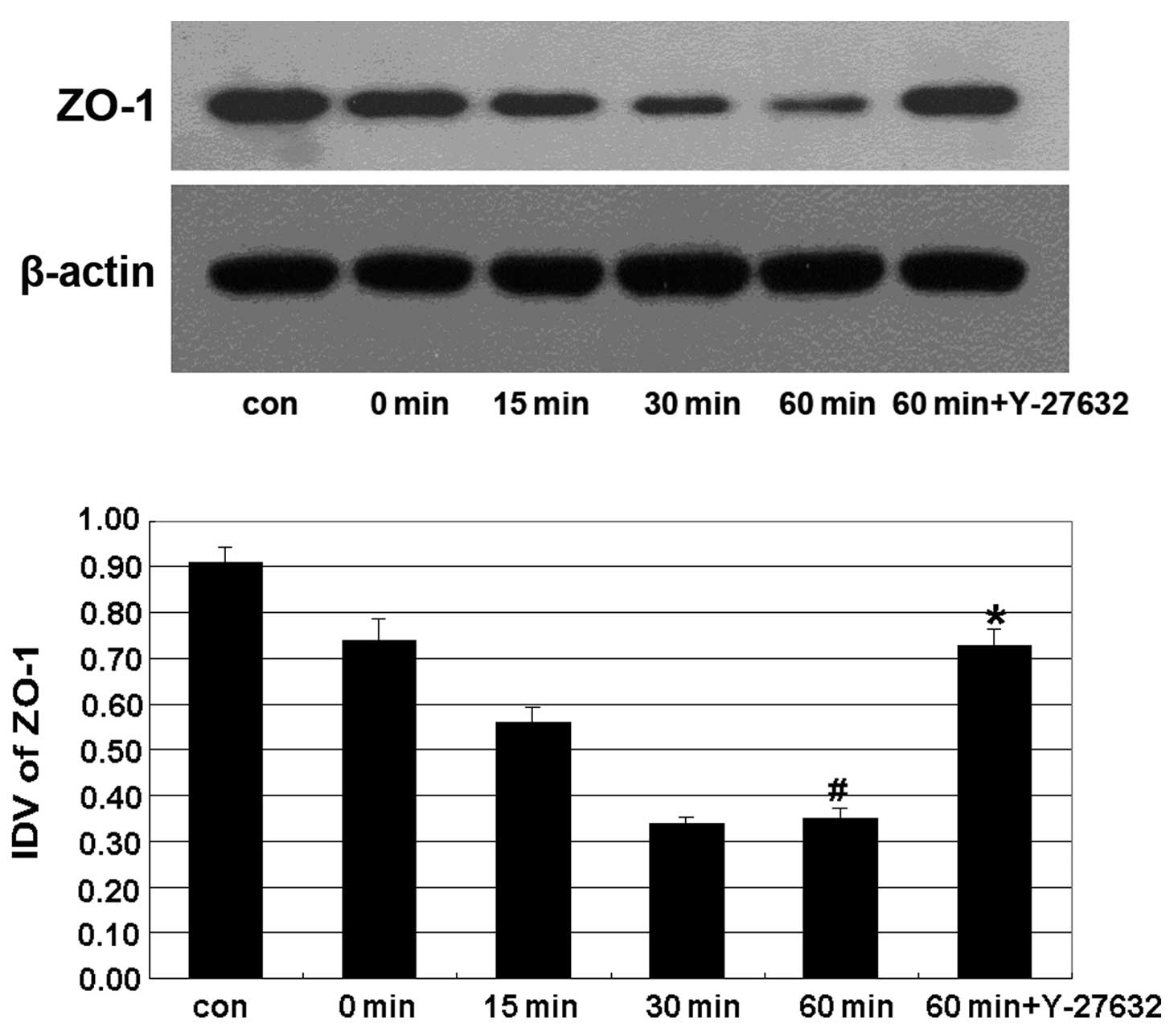
The expression of the TJ-associated protein ZO-1
displayed a progressive decline from 15 min of treatment with
ethanol, and reached its lowest level at 60 min. Y-27632 treatment
significantly reverted this effect (Fig. 3).
The Y-27632 inhibitor affects the
ethanol-induced relocalization of claudin-1 and ZO-1
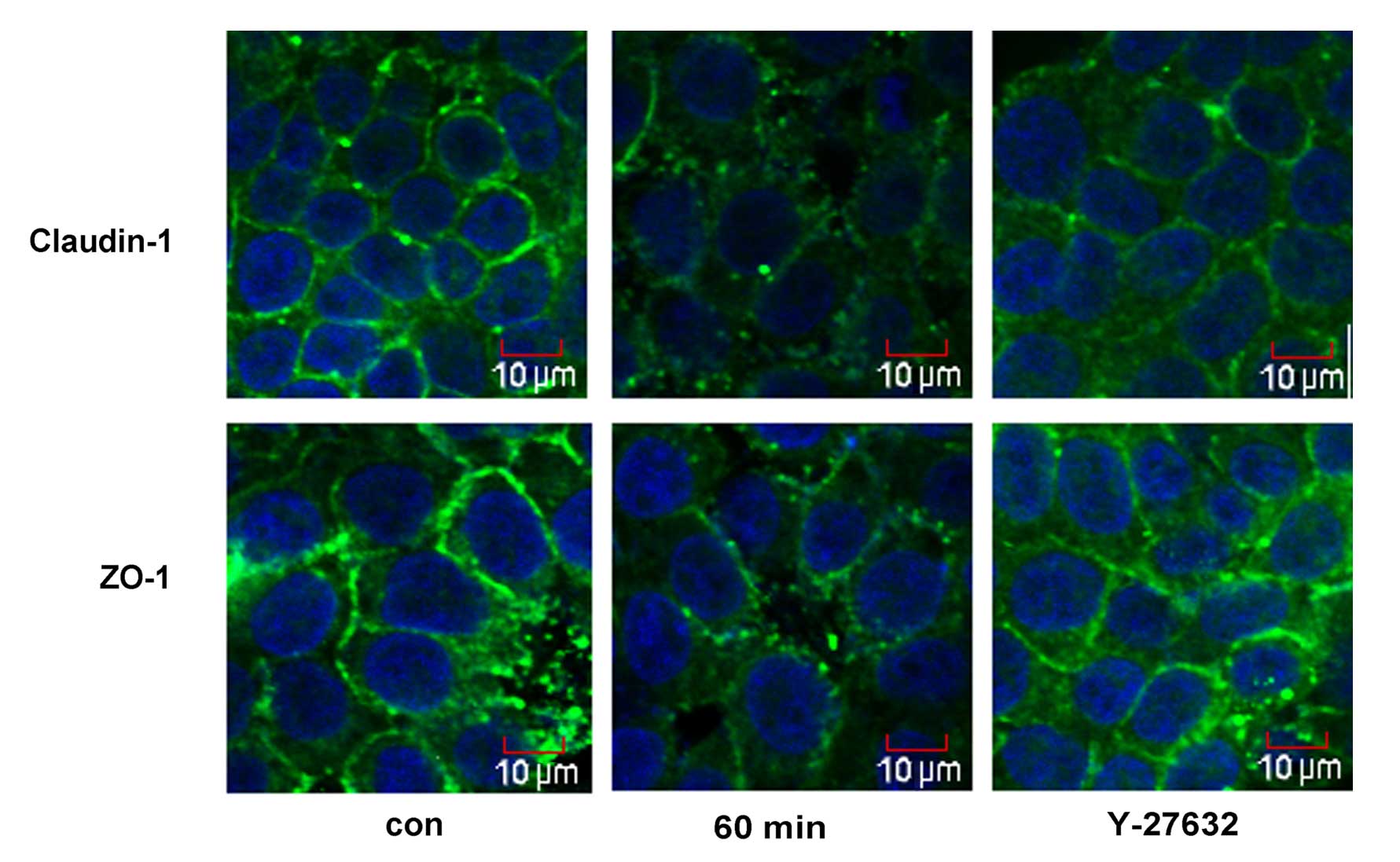
The cellular distribution of ZO-1 and claudin-1 was
assessed by immunofluorescence microscopy (Fig. 4). Treatment with 5% ethanol for 60
min induced the relocalization of claudin-1 from the cellular
membrane to the cytoplasm and the nucleus, and induced a
discontinuous distribution of ZO-1 at the cellular membrane.
Y-27632 strongly enhanced cortical localization of claudin-1 and
ZO-1 in the IEB of Caco-2 cells (Fig.
4).
Discussion
The intestinal mucosal barrier function and
integrity depend on an intact paracellular pathway, which is
largely regulated by intercellular junctions, i.e. TJs, adherens
junctions (AJs) and desmosomes (22). The TJs are multiprotein complexes
composed of transmembrane proteins (the claudin family, occludin,
junction adhesion molecules, etc) that interact with the
cytoplasmic plaque proteins (e.g., ZO-1, ZO-2 and ZO-3) (22–24).
In vitro studies using the conventional two-dimensional (2D)
cell culture model of intestinal cell monolayers grown on filters
have shown that ethanol disrupts the epithelial TJ integrity and
thereby, increases paracellular permeability (25,26).
Thus, disassembly of tight-junction proteins is likely a causal
factor in the pathogenesis of ethanol-induced intestinal barrier
dysfunctions. However, whether the ROCK inhibitor Y-27632 can
inhibit the ethanol-induced increase in IEB permeability had not
been investigated to date.
RhoA is a member of a subfamily of small GTPases
that is involved in numerous cellular functions, including the
regulation of actin filament reorganization and cell shape. The
Rho-associated protein kinase (ROCK) as a downstream effector of
RhoA, plays important roles in cellular processes that involve
actin cytoskeletal rearrangement, including stress fiber formation,
axonal growth, tumor cell invasion, and activation of platelets
(27,28). However, it is unknown whether this
effector plays a role in Rho-dependent TJ assembly. We previously
showed that ethanol induces a significant time-dependent decrease
in transepithelial electrical resistance (TEER) and a
time-dependent increase in the LY flux rate, with the lowest and
the highest values, respectively, obtained after 60 min of
treatment. In this study, we found that Y-27632 can prevent the
decrease in TEER and the increase in the LY flux rate (Fig. 1). LY flux and TEER assays were
previously used with success to evaluate the IEB permeability
(29). The ROCK inhibitor Y-27632
prevented the ethanol-induced increase in IEB permeability. We used
an ethanol-treated Caco-2 intestinal epithelial cell monolayer
in vitro model along with western blot analysis to
investigate whether ethanol can induce intestinal barrier
dysfunction and affect the expression of the TJ-associated proteins
ZO-1 and claudin-1. Our results demonstrated that ethanol induces a
shift in the distribution of claudin-1, from the insoluble to the
soluble fraction (Fig. 2).
Furthermore, ethanol affected the expression of ZO-1 in Caco-2
cells (Fig. 3), supporting the
conclusion that ethanol can affect the expression of TJ-associated
proteins, disrupt the TJs and increase the permeability of the
intestinal barrier. Moreover, Y-27632 treatment prevented the
disruption of TJ-associated proteins ZO-1 and claudin-1 induced by
ethanol treatment (Figs. 2 and
3). Immunofluorescence microscopy
was further used to study the distribution of claudin-1 and ZO-1 in
the cells (Fig. 4). Ethanol
induced a redistribution of claudin-1 from the plasmalemma to the
cytoplasm and caused a discontinuous distribution of ZO-1 at the
cellular membrane. Treatment with Y-27632 partly prevented the
ethanol-induced relocalization of claudin-1 and ZO-1.
In conclusion, our study showed that the ROCK
inhibitor Y-27632 can prevent the ethanol-induced increase in IEB
permeability and TJ assembly, suggesting that ROCK is required for
the ethanol-mediated increase in IEB permeability. However, Y-27632
only partially reverted the effects of ethanol treatment in our
study. We argue that this suggests the involvement of additional
signaling pathways and molecules in IEB permeability.
References
|
1
|
Tsukamoto H: Conceptual importance of
identifying alcoholic liver disease as a lifestyle disease. J
Gastroenterol. 42:603–609. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gramenzi A, Caputo F, Biselli M, et al:
Review article: alcoholic liver disease - pathophysiological
aspects and risk factors. Aliment Pharmacol Ther. 24:1151–1161.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Adachi M and Brenner DA: Clinical
syndromes of alcoholic liver disease. Dig Dis. 23:255–263. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mann RE, Smart RG and Govoni R: The
epidemiology of alcoholic liver disease. Alcohol Res Health.
27:209–219. 2003.PubMed/NCBI
|
|
5
|
Adachi Y, Moore LE, Bradford BU, Gao W and
Thurman RG: Antibiotics prevent liver injury in rats following
long-term exposure to ethanol. Gastroenterology. 108:218–224. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bode C, Kugler V and Bode JC: Endotoxemia
in patients with alcoholic and non-alcoholic cirrhosis and in
subjects with no evidence of chronic liver disease following acute
alcohol excess. J Hepatol. 4:8–14. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Forsyth CB, Voigt RM, Shaikh M, Tang Y,
Cederbaum AI, Turek FW and Keshavarzian A: Role for intestinal
CYP2E1 in alcohol-induced circadian gene-mediated intestinal
hypermeability. Am J Physiol Gastrointest Liver Physiol.
305:G185–G195. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zima T and Kalousova M: Oxidative stress
and signal transduction pathways in alcoholic liver disease.
Alcohol Clin Exp Res. 29(Suppl 11): 110S–115S. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Farhadi A, Keshavarzian A, Ranjbaran Z,
Fields JZ and Banan A: The role of protein kinase C isoforms in
modulating injury and repair of the intestinal barrier. J Pharmacol
Exp Ther. 316:1–7. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Turner JR: Intestinal mucosal barrier
function in health and disease. Nat Rev Immunol. 9:799–809. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu J, Liu F, Yin P, et al: Involvement of
oxidative stress and mitogen-activated protein kinase signaling
pathways in heat stress-induced injury in the rat small intestine.
Stress. 16:99–113. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rescigno M: The intestinal epithelial
barrier in the control of homeostasis and immunity. Trends Immunol.
32:256–264. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ulluwishewa D, Anderson RC, McNabb WC,
Moughan PJ, Wells JM and Roy NC: Regulation of tight junction
permeability by intestinal bacteria and dietary components. J Nutr.
141:769–776. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Blaskewicz CD, Pudney J and Anderson DJ:
Structure and function of intercellular junctions in human cervical
and vaginal mucosal epithelia. Biol Reprod. 85:97–104. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Groschwitz KR and Hogan SP: Intestinal
barrier function: molecular regulation and disease pathogenesis. J
Allergy Clin Immunol. 124:3–20. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Atkinson KJ and Rao RK: Role of protein
tyrosine phosphorylation in acetaldehyde-induced disruption of
epithelial tight junctions. Am J Physiol Gastrointest Liver
Physiol. 280:G1280–G1288. 2001.PubMed/NCBI
|
|
17
|
Ma TY, Nguyen D, Bui V, Nguyen H and Hoa
N: Ethanol modulation of intestinal epithelial tight junction
barrier. Am J Physiol. 276:G965–G974. 1999.PubMed/NCBI
|
|
18
|
Rao RK: Acetaldehyde-induced increase in
paracellular permeability in Caco-2 cell monolayer. Alcohol Clin
Exp Res. 22:1724–1730. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wojciak-Stothard B and Ridley AJ: Rho
GTPases and the regulation of endothelial permeability. Vascul
Pharmacol. 39:187–199. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Walsh SV, Hopkins AM, Chen J, Narumiya S,
Parkos CA and Nusrat A: Rho kinase regulates tight junction
function and is necessary for tight junction assembly in polarized
intestinal epithelia. Gastroenterology. 121:566–579. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tong J, Wang Y, Chang B, Zhang D and Wang
B: Evidence for the involvement of RhoA signaling in the
ethanol-induced increase in intestinal epithelial barrier
permeability. Int J Mol Sci. 14:3946–3960. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tsukita S, Furuse M and Itoh M:
Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol.
2:285–293. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Laukoetter MG, Bruewer M and Nusrat A:
Regulation of the intestinal epithelial barrier by the apical
junctional complex. Curr Opin Gastroenterol. 22:85–89. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schulzke JD and Fromm M: Tight junctions:
molecular structure meets function. Ann NY Acad Sci. 1165:1–6.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rao RK: Acetaldehyde-induced barrier
disruption and paracellular permeability in Caco-2 cell monolayer.
Methods Mol Biol. 447:171–183. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sheth P, Seth A, Atkinson KJ, et al:
Acetaldehyde dissociates the PTP1B-E-cadherin-beta-catenin complex
in Caco-2 cell monolayers by a phosphorylation-dependent mechanism.
Biochem J. 402:291–300. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Leung T, Chen XQ, Manser E and Lim L: The
p160 RhoA-binding kinase ROK alpha is a member of a kinase family
and is involved in the reorganization of the cytoskeleton. Mol Cell
Biol. 16:5313–5327. 1996.PubMed/NCBI
|
|
28
|
Bito H, Furuyashiki T, Ishihara H, et al:
A critical role for a Rho-associated kinase, p160ROCK, in
determining axon outgrowth in mammalian CNS neurons. Neuron.
26:431–441. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cui W, Li LX, Sun CM, et al: Tumor
necrosis factor alpha increases epithelial barrier permeability by
disrupting tight junctions in Caco-2 cells. Braz J Med Biol Res.
43:330–337. 2010. View Article : Google Scholar : PubMed/NCBI
|