Introduction
Formaldehyde (FA) is an environmental agent commonly
found in numerous products, including paint, cloth and exhaust gas,
as well as other medicinal and industrial products. FA exposure has
raised significant concerns due to mounting evidence suggesting its
carcinogenic potential and severe effects on human health (1). Epidemiological and experimental data
have demonstrated that FA may cause leukemia, particularly myeloid
leukemia; however, the mechanism underlying this effect remains
unclear (2–4). In the process of tumor formation, DNA
damage may be the initiating factor (5), while myeloperoxidase (MPO) activity
is associated with carcinogen metabolism, DNA repair inhibition and
DNA damage (6–8). MPO is a specific neutrophil marker
and is able to catalyze the generation of reactive oxygen species
(ROS). ROS comprises a class of ubiquitous molecules that include
superoxide anion (O2), hydrogen peroxide
(H2O2), the hydroxyl radical (OH) and ROOH.
When ROS formation and the levels of protective anti-oxidants are
unbalanced, excess ROS causes toxic effects and ultimately leads to
pathological damage (9–10).
Oxidative stress is considered to be a critical
mediator of damage induced by FA, as has been observed in a number
of studies (11). Peroxiredoxins2
(Prx2), a member of the Prxs family, exhibits an important role in
scavenging reactive oxygen species in the body and protecting cells
from damage (12). A previous
study identified that Prx2 deficiencies are associated with the
onset of oxidative stress and are involved in the removal of
H2O2 (13).
Although toxicity studies of FA have been widely reported, few
studies have examined the changes in Prx2 protein expression.
Therefore, the present study aimed to examine the possible toxic
mechanism induced by FA and investigate whether Prx2 is implicated
in FA-induced bone marrow toxicity.
Materials and methods
Reagents
FA (36.5–38% in water, FW 30.03) was obtained from
Beijing Jinhui Chemical Plant (Beijing, China). GSH-Px and MPO
Assay kits were purchased from Nanjing Jiancheng Biotechnology
Institute (Nanjing, China). The anti-MPO antibody was purchased
from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). The
anti-Prx2 antibody was purchased from SAB (College Park, MD, USA).
Agarose and ethidium bromide were purchased from Ameresco LLC
(Solon, OH, USA). All reagents were of the highest purity
commercially available.
Animals and treatment
Male ICR mice, weighing 13±1 g, were purchased from
the Center for Experimental Animals of the Norman Bethune College
of Jilin University, (Changchun, China; Animal Certificate no.
SCXK-Ji 2007–0003) and maintained under standard housing conditions
(temperature, 20~24°C; relative humidity, 40~70%; light/dark cycle,
12 h:12 h). Food and water were provided ad libitum. In
accordance with the principle of randomization, the mice were
divided into four groups (n=6 per group). According to the results
of the preliminary experiment, one group was exposed to ambient air
and the other three groups were exposed to different concentrations
of FA (20, 40, 80 mg/m3) for 15 days in the respective
inhalation chambers, for 2 h a day. All animal experiments were
treated according to the procedures evaluated and approved by the
Ethical Committee of Norman Bethune Department of Medicine, Jilin
University (Jilin, China). At the end of the 15-day experimental
period, all of the mice were sacrificed by cervical dislocation and
the bone marrow cells were obtained. A number of the bone marrow
cells were used for the determination of pathology, glutathione
peroxidase (GSH-Px) activity, and MPO activity and protein
expression; others were used for the determination of DNA damage
and the changes in Prx2 expression.
Histopathology of bone marrow
The sternums of mice were extracted once the mice
had been sacrificed. The sections were fixed in 10% FA solution and
dehydrated and embedded in paraffin. Finally 5-μm sections were cut
and stained with hematoxylin-eosin. The sections were observed
under a BH-2 light microscope (Olympus, Tokyo, Japan).
Determination of GSH-Px and MPO
activity
The femurs were extracted, and the surrounding
muscles and connective tissues were removed. The two ends of the
femurs were cut and the bone marrow cells were washed out with 0.01
mol/l phosphate-buffered saline using a 1 ml syringe. The bone
marrow cells were filtered through 200 mesh nylon nets to obtain a
single bone marrow cell suspension. GSH-Px and MPO activity was
detected by MPO Activity Assay kits purchased from Nanjing
Jiancheng Biotechnology Institute (Nanjing, China).
Determination of MPO and Prx2 protein
expression by immunohistochemistry
The sternums of mice were removed. The bone marrow
cells in the sternums were washed out and conventionally smeared.
The slides were naturally dried and fixed with 10% formalin buffer
solution for 15 min, naturally dried again and placed in a −20°C
freezer. Next, the expression of MPO and Prx2 proteins was detected
by immunohistochemistry. Finally, the bone marrow cells were
observed under a light microscope. Cells with brown particles
deposited on their membrane or nucleus were counted as positive
cells and the rate of positive cells was calculated using the
following formula: Rate of positive cells (%) = number of cells
positive for MPO or Prx2/total number of cells × 100.
Determination of micronuclei
formation
First the sternums of the mice were removed, the
bone marrow cells were washed out and then smeared onto the slides.
Next, the slides were fixed and stained with a 1:9 Giemsa phosphate
buffer for 15 min. Excess dye was washed off and the slides were
dried. The slides were observed under a light microscope and 1,000
polychromatic erythrocytes from each mouse were counted.
Determination of DNA damage
Single cell gel electrophoresis (SCGE) assay, also
termed the comet assay was used to determine the DNA damage of bone
marrow cells (14). Following
staining by ethidium bromide (EB), the bone marrow cells were
observed under a BX51 fluorescence microscope (Olympus). According
to the level of damage, the DNA damage was divided into five
grades, 0, I, II, III and IV. The rate of DNA damage (%) = number
of cells with damaged DNA/total number of cells × 100 (15).
Western blot analysis of Prx2
protein
After the single bone marrow cell suspension was
established, the protein was extracted and measured. Next, 40 μg of
protein was separated by 12% SDS-PAGE and electro-transferred onto
nitrocellulose membranes, blocked with western blotting analysis
confining fluidII (1X TBS containing 5% skimmed milk powder and
0.1% Tween-20) for 2 h and incubated with primary polyclonal
antibodies against β-actin at a dilution of 1:3,000 and against
Prx2 at a dilution of 1:100 for 1 h at room temperature. The
membrane was washed and the primary antibodies were incubated for
40 min at room temperature with the appropriate horseradish
peroxidase-conjugated secondary antibody. The bands were visualized
using a Western Luminescent Detection kit (Beijing Vigorous
Biotechnology Institute, Beijing, China) on X-ray film (Fusion FX7,
Vilber Lourmat, Marne La Vallée, France).
Statistical analysis
Statistical analysis was performed by SPSS 14.0
statistical software (SPSS, Inc., Chicago, IL, USA). The difference
between the control and FA-treated groups was tested by one-way
analysis of variance and the DNA (measured by single cell gel
electrophoresis assay) was tested by a χ2-test.
P<0.05 was considered to indicate a statistically signficant
difference.
Results
Bone marrow pathology
It was observed that the numbers of bone marrow
cells in distal areas were decreased, particularly in the 40 and 80
mg/m3 FA-treated groups. In the central areas, the
numbers of bone marrow cells were also decreased and were
simultaneously accompanied by the sinus expansion. (Fig. 1).
Effects of FA on GSH-Px activity
Following FA exposure, the GSH-Px activity was
significantly decreased, and there was a significant difference in
the activty in the 40 and 80 mg/m3 FA-treated groups
compared with the control (P<0.05; Fig. 2).
Effects of FA on MPO activity and protein
expression
MPO activity and protein expression were increased
in a dose-dependent manner compared with the control (P<0.05;
Fig. 3).
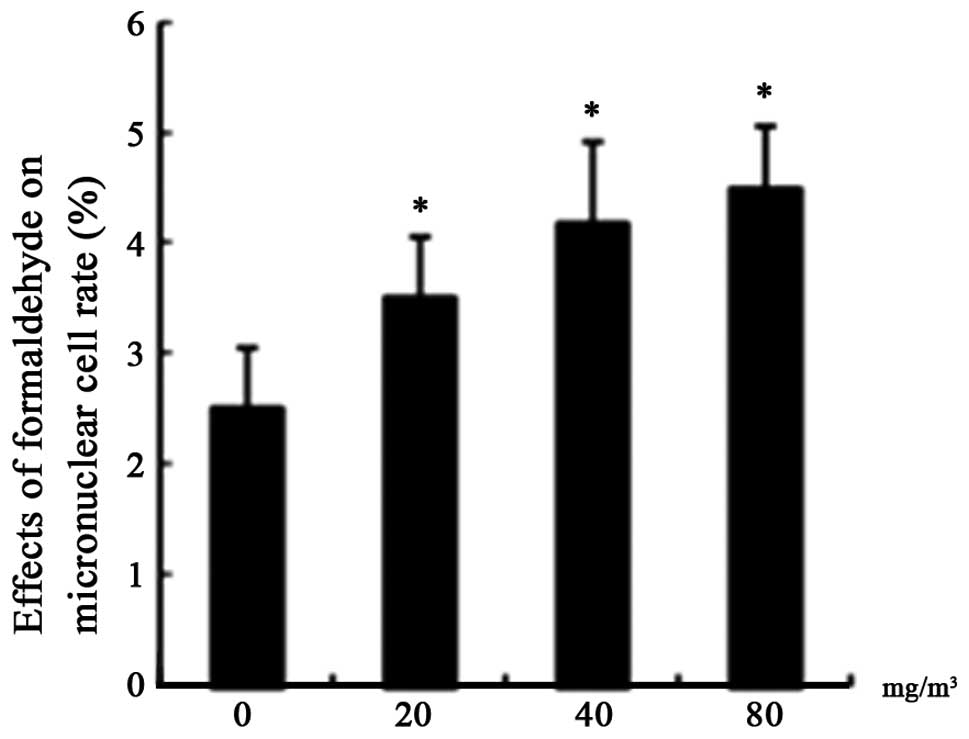
Effects of FA on the rate of micronucleus
formation
The reate of micronucleus formation in each
FA-treated group was increased and the difference was significant
compared with the control (P<0.05; Fig. 4).
Effects of FA on DNA damage
The DNA damage of bone marrow cells was detected by
SCGE, in which the DNA-damaged cells had comet tails and were
divided into five grades according to the level of damage. The
results demonstrated that the DNA damage rate was enhanced as the
FA concentration increased and there was a significant difference
compared with the control (P<0.05; Table I).
 | Table IEffects of formaldehyde on the DNA
damage in the bone marrow of mice. |
Table I
Effects of formaldehyde on the DNA
damage in the bone marrow of mice.
| FA concentration
(mg/m3) | No. of cells with DNA
damage | Total no. | Damage rate (%) |
|---|
|
|---|
| 0 | I | II | III | IV |
|---|
| 0 | 95 | 4 | 1 | 0 | 0 | 100 | 5.00 |
| 20 | 65 | 6 | 2 | 0 | 0 | 73 | 10.96 |
| 40 | 78 | 10 | 3 | 0 | 0 | 91 | 14.29a |
| 80 | 76 | 18 | 3 | 1 | 0 | 98 | 22.45a |
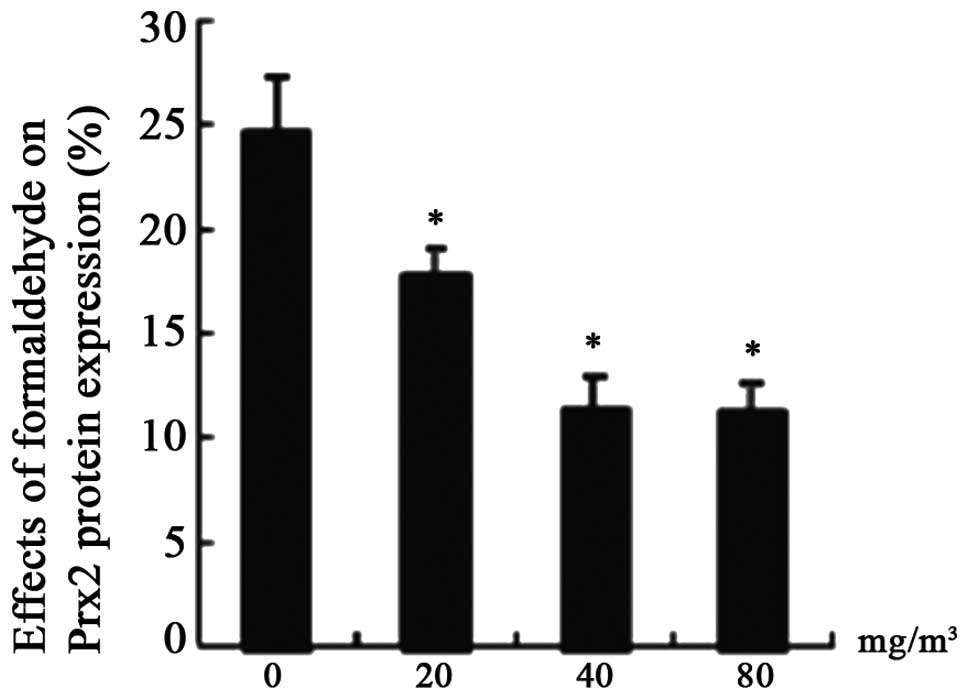
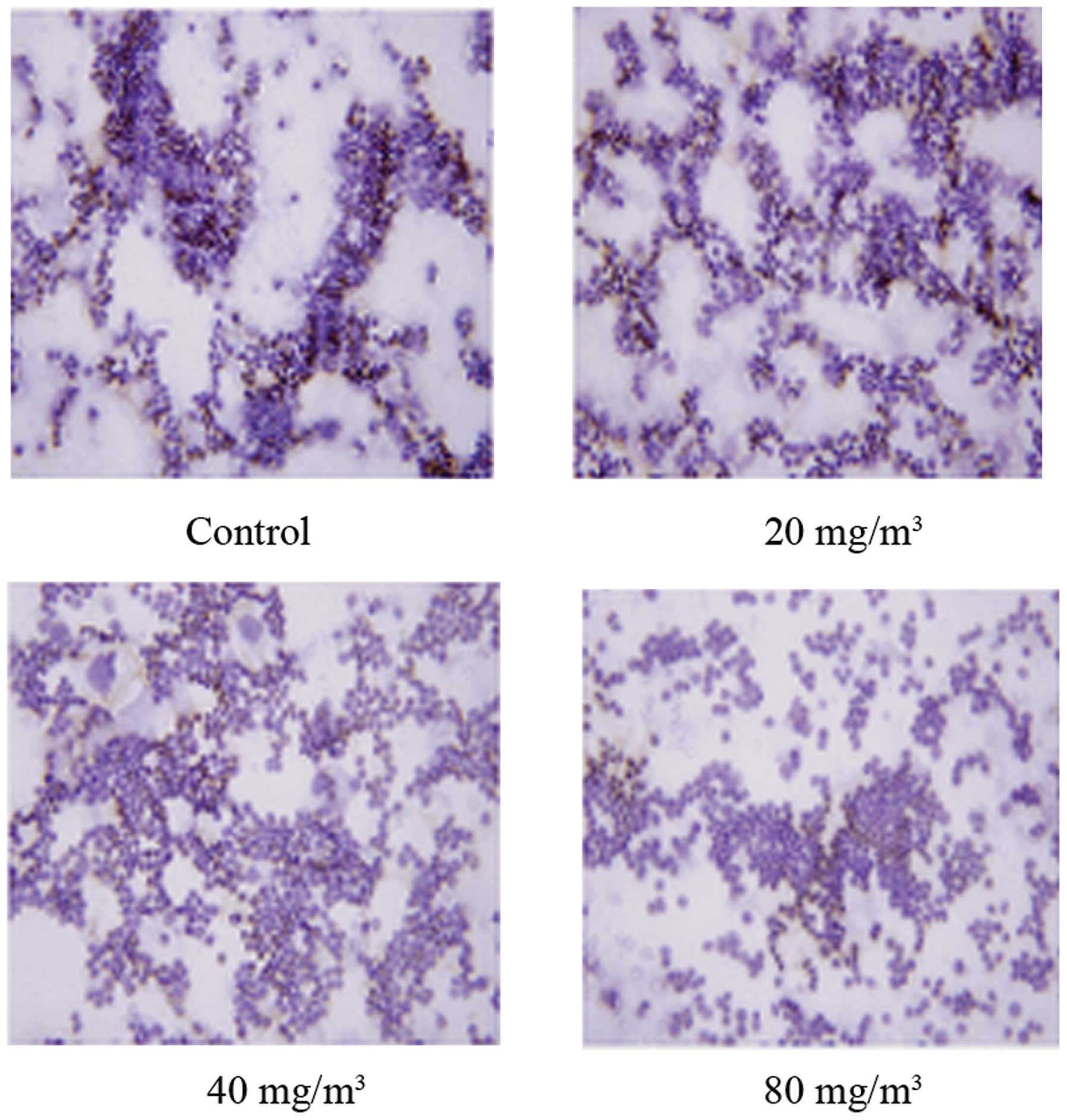
Expression of Prx2 protein
The expression of Prx2 protein was detected by
immunohistochemistry and western blotting (Figs. 5 and 6). The results demonstrated that Prx2
expression decreased as the FA concentration increased, and there
was an evident difference in FA-treated groups compared with the
control (P<0.05). As demonstrated in Fig. 7, the number of brown positive cells
progressively decreased between the FA-treated and control
groups.
Discussion
FA is an electrophilic agent. As an oxidant with
weak radicals, it causes chemical processes similar to the Fenton
reaction and produces strong oxidants, including the hydroxyl
radical. Hydroxyl radicals combine with glutathione (GSH), an
important antioxidant in the body, to form thio-hemiacetal. This
process causes GSH depletion damaging the redox balance in the body
and resulting in oxidative stress. The process of lipid
peroxidation is a type of oxidative stress, which can cause cell
dysfunction by producing a variety of free radicals and non-radical
products, particularly lipid free radicals. These lipid free
radicals easily penetrate and diffuse into the nucleus, act on
nucleic acids to induce DNA base modifications and strand breaks,
and thus lead to cell mutations and cancer. Studies have
demonstrated that MPO catalyzes the process of lipid peroxidation
(16).
The present result demonstrated that a certain
concentration of FA may decrease GSH-Px levels and increase MPO
activity and protein expression. GSH-Px is a selenium-containing
protein, GSH is its main substrate and binding results in
conversion of harmful compounds into harmless hydroxyl compounds
and water. Therefore, it is able to protect the structure and
function of the cell membrane from damaging peroxide, and may
effectively remove lipid peroxides from the body. MPO is an
important heme-containing enzyme released by Azure particles of
neutrophils and catalyses the reaction between hydrogen peroxide
and chlorine to produce hypochlorous acid. Therefore, it is
important for the development of host defense and inflammation. In
addition, MPO is also a functional sign and an activation marker of
polymorphonuclear (PMN) leukocytes, the change in MPO activity
represents the function and activity status of PMNs. FA may cause
GSH depletion and lead to lipid peroxidation. This process may
trigger the activation of PMNs. When PMNs are activated, they are
able to release MPO into the extracellular space or into phagosomes
by degranulation, which stimulates H2O2 to
generate a series of ROS with broad biological effects at sites of
inflammation. In addition, MPO may also pass activated signals from
the cytoplasm to the nucleus by a cell-signaling pathway to induce
the expression of inflammation-related genes. The pathology results
demonstrated that the number of bone marrow cells in the 40 and 80
mg/m3 FA-treated groups were evidently decreased and
accompanied by a sinus expansion. This suggested that FA was able
to cause an inflammatory reaction in the bone marrow. MPO may
therefore promote this process.
Increased lipid hydroperoxide production by FA may
be associated with Prx2 activity. Prx2, a member of the peroxidase
family, acts as a key scavenger of ROS. It is able to modulate
cytokine-induced H2O2 levels, which have been
demonstrated to mediate signaling cascades leading to cell
proliferation, differentiation and apoptosis. In addition, it also
has an important role in peroxide detoxification in cells in
cooperation with GSH-Px (17). In
the present study, it was identified that the decrease in Prx2
expression was associated with the bone marrow toxicity induced by
FA. The downregulation of Prx2 increases oxidative stress (18), resulting in subsequent toxic
effects and inflammatory responses. Consistent with these results,
several other studies have observed that, among the six Prx
isoforms, only Prx2 expression decreased in response to FA
(19).
The present study also examined the effects of FA on
DNA damage, using the micronucleus test and SCGE. The results
suggested that certain concentrations of FA may cause chromosome
breakage and the formation of micronuclei during the process of
cell division in vivo. One hypothesis is that FA interferes
with spindle fiber function or exhibits a role in internal
chromosomal bonding, which may cause chromosome breakage to form
micronuclei. FA may also cause lipid peroxidation to produce lipid
free radicals. These lipid free radicals may act on nucleic acids
to induce DNA base modifications and strand breaks, causing an
increase in micronuclei. When detecting DNA damage using SCGE, it
was identified that there was also an evident difference between
the FA-treated groups and control groups.
In conclusion, the results suggest that certain
concentrations of FA may have toxic effects on bone marrow cells.
Lipid peroxidation had an important effect and the MPO level
promoted this process. It was also identified that decreased Prx2
expression was implicated as an important mechanism underlying
FA-induced bone marrow toxicity.
References
|
1
|
National Toxicology Program. Formaldehyde.
Rep Carcinog. 12:195–205. 2011.
|
|
2
|
Cole P and Axten C: Formaldehyde and
leukemia: an improbable caused relationship. Regul Toxicol
Pharmacol. 40:107–112. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schwilk E, Zhang L, Smith MT, Smith AH and
Steinmaus C: Formaldehyde and leukemia: an updated meta-analysis
and evaluation of bias. J Occup Environ Med. 52:878–886. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Speit G, Gelbke HP, Pallapies D and
Morfeld P: Occupational exposure to formaldehyde, hematotoxicity
and leukemia-specific chromosome change in cultured myeloid
progenitor cells. Cancer Epidemiol Biomarkers Prev. 19:1882–1884.
2010. View Article : Google Scholar
|
|
5
|
Xie Yingcheng: The progress of DNA damage
and tumor. Chin J Cancer. 16:313–317. 2006.
|
|
6
|
Neuss S, Holzmann K and Speit G: Gene
expression changes in primary human nasal epithelial cells exposed
to formaldehyde in vitro. Toxicol Lett. 198:289–295. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ladeira C, Viegas S, Carolino E, et al:
Genotoxicity biomarkers in occupational exposure to formaldehyde -
the case of histopathology laboratories. Mutat Res. 721:15–20.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Klein MD, Sinha BK and Subramaniam RP:
Statistical inferences from formaldehyde DNA-protein cross-link
data: improving methods for characterization of uncertainty. J
Biopharm Stat. 21:42–55. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou DX, Qiu SD, Zhang J, Tian H and Wang
HX: The protective effect of vitamin E against oxidative damage
caused by formaldehyde in the testes of adult rats. Asian J Androl.
8:584–588. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kovacic P and Somanathan R: Dermal
toxicity and environmental contamination: Electron transfer,
reactive oxygen species, oxidative stress, cell signaling, and
protection by antioxidants. Rev Environ Contam Toxicol.
203:119–138. 2010.
|
|
11
|
Zhang Y, Liu X, McHale C, et al: Bone
marrow injury induced via oxidative stress in mice by inhalation
exposure to formaldehyde. PLoS One. 8:e749742013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cox AG, Winterbourn CC and Hampton MB:
Mitochondrial peroxiredoxin involvement in antioxidant defence and
redox signalling. Biochem J. 425:313–325. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Smith-Pearson PS, Kooshki M, Spitz DR, et
al: Decreasing peroxiredoxin II expression decreases glutathione,
alters cell cycle distribution, and sensitizes glioma cells to
ionizing radiation and H(2)O(2). Free Radic Biol Med. 45:1178–1189.
2008. View Article : Google Scholar
|
|
14
|
Singh NP, McCoy MT, Tice RR and Schneider
EL: A simple technique for quantitation of low levels of DNA damage
in individual cells. Exp Cell Res. 175:184–191. 1988. View Article : Google Scholar
|
|
15
|
Anderson D, Yu TW, Phillips BJ and
Schmezer P: The effect of various antioxidants and other modifying
agents on oxygen-radical-generated DNA damage in human lymphocytes
in the COMET assay. Mutat Res. 307:261–271. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Podrez EA, Abu-Soud HM and Hazen SL:
Myeloperoxidase-generated oxidants and atherosclerosis. Free Radic
Biol Med. 15:1717–1725. 2000. View Article : Google Scholar
|
|
17
|
Wood ZA, Schroder E, Robin Harris J and
Poole LB: Structure, mechanism and regulation of peroxiredoxins.
Trends Biochem Sci. 28:32–40. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qu D, Rashidian J, Mount MP, et al: Role
of Cdk5-mediated phosphorylation of Prx2 in MPTP toxicity and
Parkinson’s disease. Neuron. 55:37–52. 2007.PubMed/NCBI
|
|
19
|
Lim SK, Kim JC, Moon CJ, et al:
Formaldehyde induces apoptosis through decreased Prx 2 via p38 MAPK
in lung epithelial cells. Toxicology. 271:100–106. 2010. View Article : Google Scholar : PubMed/NCBI
|