Introduction
Sepsis, a systemic inflammatory response syndrome,
is a leading cause of mortality in critically ill patients
(1). Sepsis resulting from severe
infection is frequently associated with confusion, metabolic
acidosis, low blood pressure, decreased systemic vascular
resistance and blood coagulation dysfunctions (2). Severe sepsis may lead to organ
dysfunction and septic shock (3).
Although the recommendations of the treatment targeting for sepsis
are updated every four years (4–6), the
mortality rate resulting from sepsis remains high, at between 30
and 50% (7).
To decrease the mortality rate and identify an
effective treatment for sepsis, numerous studies have focused on
identifying the pathogenic mechanisms that underlie sepsis.
Disease-conditioned mesenteric lymph, but not portal blood, carries
high levels of gut-derived factors leading to organ injury
(8,9). As outlined in previous studies,
protein fractions in the lymph are partly responsible for
gut-origin sepsis (10,11). In addition, certain proteins in the
lymph promote the progression of sepsis by acting as carriers of
biologically active molecules or effector molecules. With advances
in modern technology, proteomic analysis in lymph has provided a
number of breakthroughs in examining the mechanisms underlying this
critical disease (12–15).
Previously, investigations concerning sepsis have
primarily focused on the proteome in the plasma (16,17),
heart (1), liver (18) and skeletal muscle (19); however, there is little information
regarding the role of lymph proteins. The experimental sepsis model
induced by cecal ligation and puncture (CLP) has been widely used
to reproduce numerous pathophysiological features of clinical
sepsis (20,21). The significant changes in the
levels of plasma proteins in mice with CLP-induced sepsis were
reported to be during the late stage of septic development (24 h
following surgery) (17).
Therefore, in the present study, a sepsis model was constructed by
CLP and the lymph was collected at 6 h and 24 h following surgery,
which represented the early and the late stages of the acute phase,
respectively. The aim was to examine the changes in rodent proteome
mesenteric lymph in response to sepsis, in order to contribute to
the current understanding of the pathogenesis of sepsis
progression.
Materials and methods
Animals
A total of 28 adult male Sprague-Dawley rats
(Slaccas Laboratory Inc., Shanghai, China), weighing 250–300 g,
were housed at 25°C and subjected to 12 h light/dark cycle
treatment conditions. All of the animals were fed a standard rodent
diet (Slaccas Laboratory Inc.) with access to water ad
libitum. All of the animals received humane care in accordance
with the recommendations of the Guide for the Care and Use of
Laboratory Animals. Approval was obtained from the Institutional
Animal Care and Use Committee at the Second Military Medical
University (Shanghai, China).
Experimental model
The animals were randomly divided into the following
four groups (n=7 per group): CLP-6 h, sham-6 h, CLP-24 h and
sham-24 h groups. In the CLP groups, the sepsis models were
constructed by CLP as previously described (22). Briefly, general anesthetic was
induced by intraperitoneal injection of pentobarbital (40 mg/kg;
Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) and CLP was
performed with a single in-and-out puncture (20 gage needle;
Yangzhou Great Wall Medical Equipment Factory, Yangzhou, China) at
the middle part of the cecum. To ensure successful puncture,
droplets of feces were extruded from the penetration holes. The
rats in the sham groups underwent cecum exenteration under general
anesthetic instead of CLP. After surgery, all the rats were
injected with 100 g/5 ml of normal saline subcutaneously. Following
recovery from the anesthesia, the rats received free access to food
and water ad libitum.
Lymph collection
Prior to the CLP/sham surgery, the thoracic duct was
cannulated with micro-urethane tubing (inner diameter, 0.635 mm;
outer diameter, 1.02 mm; American Health & Medical Supply
International Corp. Co., Ltd., New York, NY, USA) and externalized
through a 14-gage angiocatheter (Hospira, Inc., Lake Forest, IL,
USA) in the left flank of all the experimental animals. At 6 h and
24 h following CLP/sham surgery, the lymph samples were collected
from all groups. Lymph collection was conducted by thoracic duct
drainage according to a modified method that has been previously
described (23). In brief, the
rats were re-anesthetized with intraperitoneal injection of 40
mg/kg pentobarbital. The lymph samples were collected directly into
sterile ice-chilled vacuum EDTA tubes (Shanghai Kehua
Bio-engineering Co., Ltd., Shanghai; China) for 60 min. One sample
from the seven rats within one group was centrifuged at 4°C (1,700
× g) for 15 min and immediately stored at −80°C for further
analysis.
Proteomic analysis
Proteomic analysis, according to a previous
description (24), and
bioinformatic analysis were performed by Sensichip Infotech Co.,
Ltd. (Shanghai, China). The bioinformatics analysis was used to
analyze the molecular functions of differentially expressed genes
of the identified proteins. All of the chemical reagents, software
and equipment were provided by this company. The experimental
procedures were performed according to standardized manufacturer’s
instructions.
Major protein depletion
The lymph samples were defrosted, diluted and
filtered through a 0.22-μm filter membrane (AB SCIEX, Framingham,
MA, USA). Next, three major abundant proteins (albumin, IgG and
transferrin) were depleted by the Multiple Affinity Removal Column
(M-3; Agilent Technologies Co., Ltd., Santa Clara, CA, USA). When
the samples had been individually depleted, quantitative analysis
of the lymph samples was performed with the Dc protein assay
reagent (Bio-Rad, Hercules, CA, USA). The samples were then
desalted and concentrated by ultrafiltration with a Vivaspin 4
centrifugal concentrator and 5-kDa polyethersulfone filter
(Sartorius AG, Göttingen, Germany). The target proteins were
detected by 12.5% sodium dodecyl sulfate-polyacrylamide gel
(SDS-PAGE) electrophoresis.
Capillary high performance liquid
chromatography-tandem mass spectrometry (HPLC-MS/MS)-based
proteomics
The proteins from each sample (150 μg) were
dissolved in 200 μl UA buffer (8 M urea in 150 mM Tris-HCl; pH
8.0). The solution was centrifuged (14,000 × g, 30 min) twice and
the supernate was abandoned. The protein sample was then incubated
with 100 μl iodoacetamide in the dark at room temperature for 30
min. Following centrifugation (14,000 × g, 20 min), the sample was
mixed with 100 μl UA buffer followed by reduplicate centrifugation
(14,000 × g, 20 min). The protein sample underwent reduction
(blending with 100 μl dissolution buffer and reduplicate
centrifugation at 14000 × g for 20 min) and was digested in 40 μl
trypsin buffer (2 μg trypsin in 40 μl dissolution buffer) at 37°C
for 16–18 h. Following centrifugation (14,000 × g, 10 min) in a new
collection tube, the peptides were then eluted and quantified at
optical density (OD)280 in the Unicam SP 600
spectrophotometer (Pye Unicam Ltd., Cambridge, UK)
The sample (50 μg) was labeled with iTRAQ reagents
(iTRAQ Reagent-4 plex Multiplex kit; AB SCIEX) according to the
manufacturer’s instructions. Samples from CLP-6 h, CLP-24 h, sham-6
h and sham-24 h groups were labeled with 114, 115, 116 and 117
iTRAQ isobaric tags, respectively. The labeled materials were then
combined and pre-fractionated on a polysulfoethyl 4.6×100-mm column
(5 μm, 200 Å; PolyLC Inc., Columbia, MD, USA) using an AKTA
Purifier 100 (GE Healthcare, Little Chalfont, UK). Approximately 30
effluent fractions were collected and merged into ten fractions
according to the strong cation exchange chromatogram. The fractions
were freeze-dried and desalted using a C18 cartridge
(Sigma, St. Louis, MO, USA).
Subsequently, HPLC analysis was performed. The
samples were added to the sample column (2 cm × 100 μm, 5 μm-C18;
EASY-Column Capillary Column; AB Sciex, Waltham, MA, USA)
automatically and were then separated through an analytical column
(75 μm × 100 mm, 3 μm-C18; EASY-Column Capillary Column; Thermo
Fisher Scientific). The HPLC Easy nLC gradient between buffer A
(0.1% formic acid in water) and buffer B (0.1% formic acid in
acetonitrile) was formed at 250 μl/min as follows: Increasing to
35% B from 0–100 min, increasing to 100% B by 108 min and held at
100% until 120 min. The sample effluent was directed into the ion
spray source of a Q-Exactive mass spectrometer (Thermo Finnigan,
San Jose, CA, USA) scanning from 300 to 1800 m/z for 120 min.
Sequence database searches
The sequence database searches and quantitative
analysis were performed by Mascott 2.2 software (Matrix Science
Ltd., Boston, MA, USA), Proteome Discoverer 1.3 and Xcalibur
softwares (Thermo Fisher Scientific).
The data were searched against the
ipi.RAT.v3.87.fasta database (sequence no. 39925) using Mascot 2.2
software with the search parameters as follows: Type of search,
MS/MS ion search; enzyme, trypsin; mass values, monoisotopic; max
missed cleavages, 2; fixed modifications, carbamidomethyl (C),
iTRAQ4plex (N-term), iTRAQ4plex (K); variable modifications,
oxidation (M), iTRAQ4plex (Y); peptide mass tolerance, ±20 ppm; and
fragment mass tolerance, 0.1 Da. Peptide false discovery rates
≤0.01 and a peptide Mascot score >20 were defined as
significant.
Data analysis
Quantitative analysis was conducted using Proteome
Discoverer 1.3 software according to the peak intensity of labeled
peptides. For different samples labeled with different tags, the
peptide quantification in one sample was calculated as the ratio of
the signal intensity of one tag compared with other the tags. The
median of the calculated ratios was considered as the result of
protein quantitation. Each median of the signal ratio was
normalized to eliminate human sample loading error. Differences in
the relative abundance were calculated as differences in the log
peak areas and reported as fold changes between two groups.
Differences among the groups were analyzed by Student’s t-test.
P<0.05 was considered to indicate a statistically significant
difference. Differentially expressed proteins were screened by the
t-test (P<0.05) and fold change (fold change>1.5) method.
Bioinformatics
In the gene ontology (GO) analysis, differentially
expressed genes (DEGs) encoding the 158 identified proteins were
mapped to the GO database using the GSEABase software package on
the R statistics platform (http://www.r-project.org/). DEGs were classified
according to three ontologies: Biological processes, cellular
components and molecular functions.
In the pathway analysis, DEGs were mapped to the
Kyoto Encyclopedia of Genes and Genomes (KEGG) database (www.bioconductor.org) by GenMAPP v2.1 (Gladstone
Institutes, San Francisco, CA, USA) and the enrichment degree of
each gene was counted in different pathways.
In the network analysis, the interactions between
DEGs were analyzed by downloading the pathway data from the KEGG,
MIPS (http://mips.helmholtz-muenchen.de/proj/ppi/) and
PubMed databases (25) and
KEGGSOAP software package (www.bioconductor.org). Inter-correlations between DEGs
were analyzed using the co-citation algorithm.
Finally, the results of the three types of analyses
were integrated into a network of protein-protein correlations with
comprehensive consideration. The network was illustrated
graphically using Medusa software (Informer Technologies, Inc.,
Bilbao, Spain).
Results
Sepsis model
Following induction of the sepsis model, the rats in
the sham and CLP groups presented physiological changes of
different degrees.
At 6 h following surgery, the rats in the sham-6 h
group presented with normal physical ability. The mental state
(assessed by observation of movements of the animal) and weight
loss of rats was marginally lower than prior to surgery. The rats
were responsive, and actively feeding and drinking. No piloerection
or abdominal distension were observed and the rats had dry granular
feces, soft stools and a clean anus. At 24 h, the rats in the
sham-24 h presented with a similar physiological status.
However, rats in the CLP-6h group exhibited poor
mental state, decreased coat glossiness, piloerection, abdominal
distension, and a fairly sensitive response to stimuli
(stimuloation by a needle in the tail). The rats stayed in groups,
did not eat and only about half of them were actively drinking
water. The abdominal incisions of the rats were red with a small
quantity of exudate and stools were soft or loose. In the CLP-24 h
group, the rats were more sluggish and also stayed in groups. The
rats had evident abdominal distention, decreased eating and
drinking habits and secretion from their eyes. The rats responded
slowly in spite of intense stimulation. Feces were observed on the
anuses and tails of the rats. The perianal hairs of the rats were
wet and the feces were primarily loose stools.
The mortality rate was 60% in the CLP groups
following ten days post-operation.
Sample preparation
After the highly-abundant protein was depleted, the
concentrations of the remaining lymph protein in the CLP-6, CLP-24,
sham-6 and sham-24 groups were 1.52, 1.54, 1.51 and 2.09 μg/μl,
respectively. By quantitative analysis, the relative concentrations
of peptides were estimated by absorbance at 280 nm
(OD2801.1=1 μg/μl). The OD280 of the samples
were 1.31 (CLP-6), 1.63 (CLP-24), 1.44 (sham-6) and 1.39 (sham-24),
respectively. The depletion rate of the highly-abundant protein was
relatively high, while the concentration of the target proteins was
low.
Lymph proteomics
The abundances of 984 proteins identified in the rat
lymph samples were compared between the CLP-6 and sham-6; CLP-24
and sham-24; CLP-6 and CLP-24; and sham-6 and sham-24 groups. A
total of 158 distinct proteins with a fold change ≥1.5; P<0.05
between the different groups were identified in the present study.
Compared with the sham-6 h group, 30 of the 158 proteins were
significantly increased in the CLP-6 h group (Table I). These proteins were listed
according to their molecular function (using the freely available
protein database UniProt; http://www.uniprot.org/) and there were shown to be 13
binding proteins, two structural proteins, three enzymes, two
protease inhibitors, one cytokine, one transcription factor, five
uncharacterized proteins and three proteins with unclear function.
An additional uncharacterized protein, Pfkp (fold=0.28, P=0.0016),
was found to decrease significantly (fold change≤0.3, P<0.05) in
the CLP-6 group.
 | Table IUpregulated proteins in the lymph of
the cecal ligation and puncture-6 h group. |
Table I
Upregulated proteins in the lymph of
the cecal ligation and puncture-6 h group.
| Gene name | Description | Fold change | P-value |
|---|
| Sbp | Uncharacterized
protein | 4.29 | 0.0000 |
| Scgb2a1 | Secretoglobin
family 2A member 2 | 4.19 | 0.0000 |
| S100a9 | S100-A9
protein | 3.73 | 0.0000 |
| Ngp | Uncharacterized
protein | 3.37 | 0.0000 |
| S100a8 | S100-A8
protein | 3.04 | 0.0000 |
| Retnlg | Resistin-like
molecule gamma | 2.83 | 0.0000 |
| Psbpc2 | Prostatic
steroid-binding protein C2 | 2.71 | 0.0000 |
| Psbpc1 | Prostatic
steroid-binding protein C1 | 2.51 | 0.0000 |
| Klk1c9 | Submandibular
glandular kallikrein-9 | 2.48 | 0.0000 |
| Tceb2 | Transcription
elongation factor B polypeptide 2 | 2.38 | 0.0000 |
| LOC298109 | Uncharacterized
protein | 2.07 | 0.0000 |
| Lmnb1 | Lamin-B1 | 2.06 | 0.0000 |
| Igfbp1 | Insulin-like growth
factor-binding protein 1 | 2.00 | 0.0002 |
| Lcn2 | Neutrophil
gelatinase-associated lipocalin | 1.97 | 0.0002 |
| Fh1 | Isoform
Mitochondrial of Fumarate hydratase, mitochondrial | 1.96 | 0.0002 |
| Pigr | Polymeric
immunoglobulin receptor | 1.90 | 0.0002 |
| Clca1 | Uncharacterized
protein | 1.90 | 0.0002 |
| Tsku | Tsukushin | 1.89 | 0.0002 |
| LOC679994 | Histone H3.1 | 1.80 | 0.0004 |
| GF20391-like
isoform 1 | 1.76 | 0.0010 |
| Mpst | 3-Mercaptopyruvate
sulfurtransferase | 1.71 | 0.0041 |
| Andpro | Cystatin-related
protein 1 | 1.69 | 0.0056 |
| Myh6 | Myosin-6 | 1.64 | 0.0074 |
| Mmrn1 | Uncharacterized
protein | 1.63 | 0.0072 |
| Anxa1 | Annexin A1 | 1.61 | 0.0103 |
| Nono | Non-POU
domain-containing octamer-binding protein | 1.59 | 0.0123 |
| ApoE | Apolipoprotein
E | 1.53 | 0.0265 |
| Serpinb1a | Leukocyte elastase
inhibitor A | 1.52 | 0.0301 |
| Chi3l1 | Chitinase-3-like
protein 1 | 1.51 | 0.0310 |
| Ccdc48 | EF-hand
domain-containing protein ENSP00000381169 homolog | 1.50 | 0.0311 |
Compared with the sham-24 group, 20 proteins were
confirmed to increase significantly (fold change≥1.5; P<0.05) in
abundance of the CLP-24 group (Table
II) and included six binding proteins, two enzymes, one
protease inhibitor, one signal transducer protein and ten
uncharacterized proteins. By contrast, two proteins were identified
to decrease significantly (fold change≤0.3; P<0.05) in the
CLP-24h group compared with the sham-24 group: Svs4 (fold=0.25,
P=0.0016) and one uncharacterized protein, Svs6 (fold=0.27;
P=0.0016).
| Table IIUpregulated proteins in the lymph of
the cecal ligation and puncture-24 h group. |
Table II
Upregulated proteins in the lymph of
the cecal ligation and puncture-24 h group.
| Gene name | Description | Fold change | P-value |
|---|
| Siae | Sialate
O-acetylesterase | 3.60 | 0.0000 |
| Lcn2 | Neutrophil
gelatinase-associated lipocalin | 2.30 | 0.0000 |
| Mgp | Matrix Gla
protein | 2.05 | 0.0001 |
| Ldlr | Uncharacterized
protein | 1.81 | 0.0004 |
| Ngp | Uncharacterized
protein | 1.81 | 0.0004 |
| RGD1565682 | Uncharacterized
protein | 1.81 | 0.0004 |
| Ptgr1 | Prostaglandin
reductase 1 | 1.80 | 0.0004 |
| Sbp | Uncharacterized
protein | 1.78 | 0.0014 |
| Tnnc1 | Cardiac troponin
C | 1.70 | 0.0037 |
| LOC299282 | Uncharacterized
protein | 1.70 | 0.0033 |
| Vcan | Uncharacterized
protein (Fragment) | 1.67 | 0.0057 |
| Myh6 | Myosin-6 | 1.64 | 0.0075 |
| Sema6d | Uncharacterized
protein | 1.64 | 0.0074 |
| Il1r2 | Interleukin-1
receptor type 2 | 1.62 | 0.0102 |
| Andpro | Cystatin-related
protein 1 | 1.60 | 0.0114 |
| Slc4a1 | Uncharacterized
protein | 1.57 | 0.0148 |
| Fgl2 | Fibrinogen-like
2 | 1.56 | 0.0190 |
| Mpo | Uncharacterized
protein | 1.54 | 0.0218 |
| Chi3l1 | Chitinase-3-like
protein 1 | 1.52 | 0.0276 |
| Itpa | Uncharacterized
protein | 1.51 | 0.0296 |
Bioinformatics
The 158 proteins identified in the CLP and sham
groups were further analyzed for functional and biological
relevance. Using GO analysis, the proteins were analyzed based of
their involvement in biological processes, cellular components and
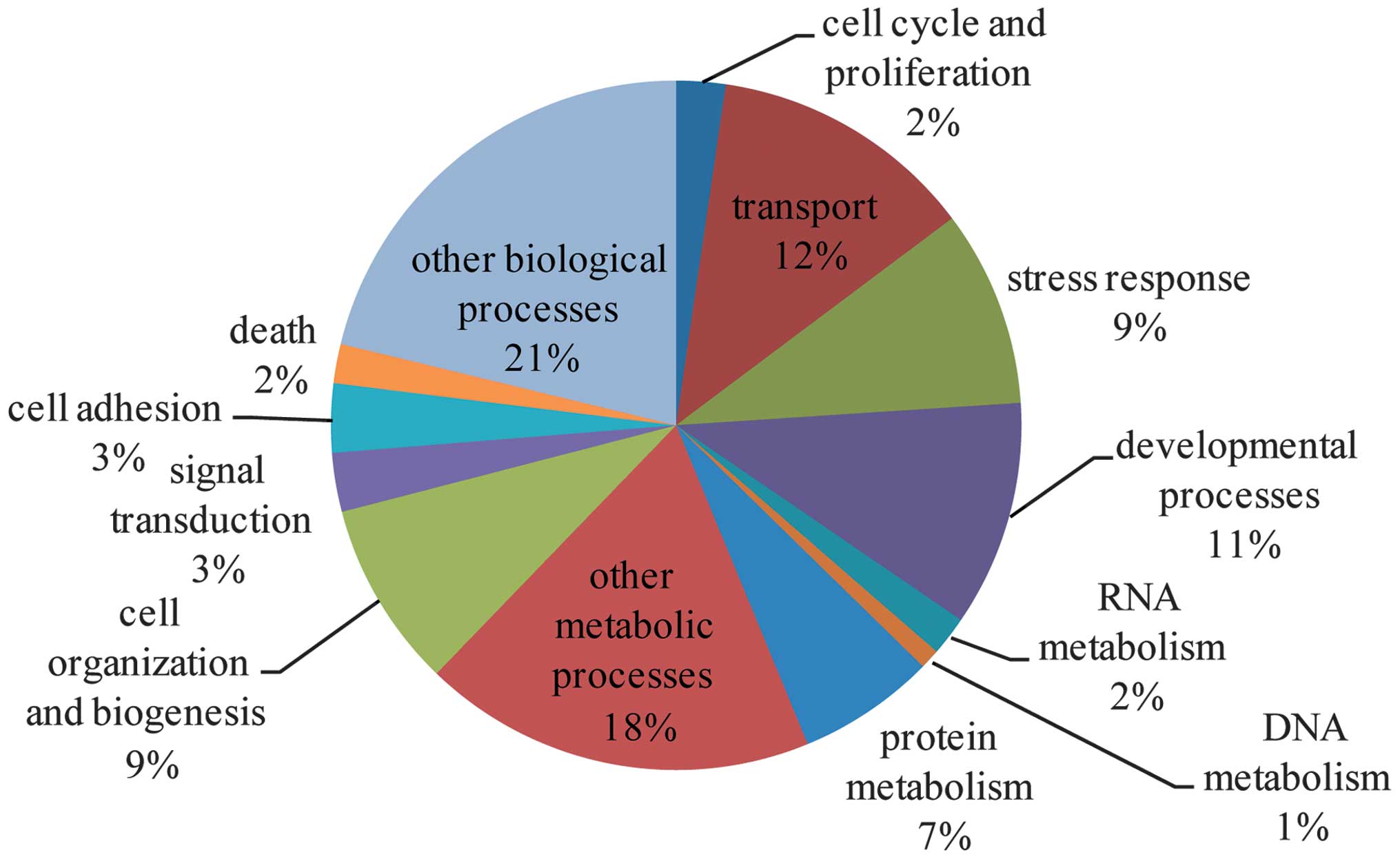
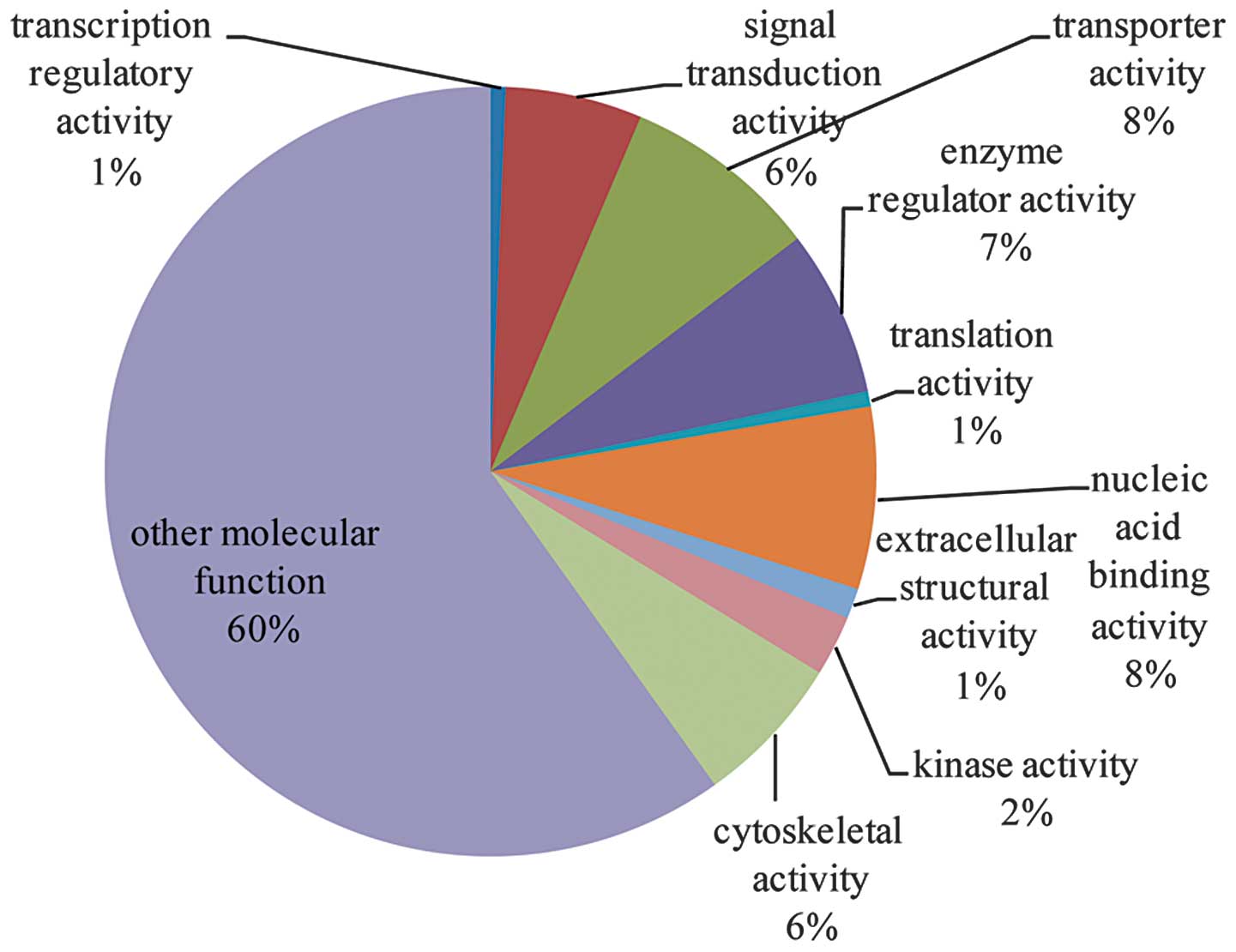
molecular functions (Figs.
1–3 and cross-referenced to
Tables III–V).
| Table IIIBiological process analysis. |
Table III
Biological process analysis.
| Term | Gene count | P-value | Gene list |
|---|
| Cell cycle and
proliferation | 5 | 0.912015 | Agt, Rcc2, Anxa1,
Hprt1, Ak1 |
| Transport | 27 | 0.002065 | ApoE, Hnrnpa1,
Slc4a1, Agt, Fabp5, Anxa1, Ptx3, Ttr, Apof, Ldlr, Hpx, Fabp4, Mpst,
Abp1, Fabp3, Lcn2, Clca1, Hba-a2, Fabp6, Myh6, Lbp, Tnnc1, Col1a1,
Mb, Atp7a, Orm1, Atp5a1 |
| Stress
response | 20 | 3.25E-05 | F13a1, ApoE, Saa4,
Thbs1, Agt, Ngp, Kng1, Ddb1, Nono, Mpo, Hpx, Fabp4, Fn1, Hspa1l,
Serpina3n, Lbp, Hspa5, Serping1, Mb, Orm1 |
| Developmental
processes | 23 | 0.069737 | ApoE, Neb, Thbs1,
Agt, Hist1h1b, Vcan, Mgp, Acan, Timp1, Hspa1l, Hba-a2, Myh6,
Sema6d, Tpm1, Tpi1, Tagln, Hprt1, Pdlim3, Col1a1, Mb, Atp7a, Hexb,
Atp5a1 |
| RNA metabolism | 4 | 0.999991 | Hnrnpa1, Nono,
Fabp4, Tceb2 |
| DNA metabolism | 2 | 0.74583 | Ddb1, Nono |
| Protein
metabolism | 14 | 0.86033 | F13a1, Pgc, Dpm1,
Ddb1, Serpinb1a, Acy1, Hpx, Rps28, Tceb2, Serping1, Hp, Rpl8,
Atp7a, sHexb |
| Other metabolic
processes | 40 | 2.44E-07 | Aox1, ApoE, Agt,
Fabp5, Dpm1, Ptx3, Acy1, Pgm1, Ttr, Apof, Car3, Ptgr1, Ldlr, Acan,
Chi3l1, Car1, Mpo, Hpx, Fabp4, Abp1, Fabp3, Pfkp, Fabp6, Fh1,
Pgam2, Lbp, Tpi1, Pgls, Pla1a, Ldha, Ckm, Hprt1, Ak1, Itpa, Eno3,
Gstm1, Atp7a, Hexb, S100a9, Atp5a1 |
| Cell organization
and biogenesis | 19 | 0.034714 | ApoE, Neb, Agt,
Hist1h1b, Rcc2, Hist1h1d, Ptx3, Ldlr, Acan, Fn1, Myh6, Lbp, Hprt1,
Hist1h1a, Pdlim3, Atp7a, Hexb, S100a9, Hist1h2ai |
| Signal
transduction | 6 | 1 | Fgl2, Agt, Anxa1,
Fgl1, Hpx, Hspa5 |
| Cell adhesion | 7 | 0.103657 | Thbs1, Agt, Vcan,
Acan, Fn1, Col1a1, Col6a2 |
| Death | 4 | 0.788118 | Fgl2, Agt, Hprt1,
Atp7a |
| Other biological
processes | 46 | 0.332232 | ApoE, Pgc, Mrfap1,
Agt, Ngp, Kng1, Rcc2, Sbp, Gemin4, Ptx3, A2m, Mgp, Apof, Tpm2,
Ldlr, Glrx3, Hpx, Fabp4, Igfbp1, Hspa1l, Retnlg, Serpina3n, Lcn2,
Cald1, Fh1, Myh6, Sema6d, Tpm1, Lbp, Ldha, Myom2, Hprt1, Hist1h1a,
Serping1, Tnnc1, Semg1, Col1a1, Rpl8, Mb, Atp7a, Orm1, S100a8,
Hexb, S100a9, Mybpc1, Prg4 |
| Table VMolecular function analysis. |
Table V
Molecular function analysis.
| Term | Gene count | P-value | Gene list |
|---|
| Transcription
regulatory activity | 1 | 0.999674 | Fabp4 |
| Signal transduction
activity | 9 | 0.999986 | Fgl2, Agt, Ttr,
Ldlr, Fgl1, Lbp, Pigr, Il1r2, Prg4 |
| Transporter
activity | 13 | 0.030194 | ApoE, Slc4a1,
Fabp5, Fabp4, Fabp3, Lcn2, Clca1, Hba-a2, Fabp6, Mb, Atp7a, Orm1,
Atp5a1 |
| Enzyme regulator
activity | 11 | 0.006352 | ApoE, Agt, Kng1,
Anxa1, Serpinb1a, A2m, Timp1, Fn1, Serpina3n, Serping1, Slpi |
| Translation
activity | 1 | 0.285497 | Hspa5 |
| Nucleic acid
binding activity | 12 | 0.962945 | Hnrnpa1, Hist1h1b,
Dpm1, Ddb1, Serbp1, Tsn, Hist1h1d, Nono, Hmgn2, Hist1h1a, Rpl8,
Hist1h2ai |
| Extracellular
structural activity | 2 | 0.013584 | Col1a2, Col1a1 |
| Kinase
activity | 4 | 0.84497 | Pfkp, Ckm, Ckb,
Ak1 |
| Cytoskeletal
activity | 10 | 0.002977 | Tpm2, Tnni2, Tpm3,
Cald1, Myh6, Tpm1, Myom2, Pdlim3, Tnnc1, Mybpc1 |
| Other molecular
function | 94 | 0.222994 | Aox1, F13a1, ApoE,
Pgc, Neb, Hnrnpa1, Mrfap1, Thbs1, Slc4a1, Lmnb1, Ngp, Fabp5,
Hist1h1b, Dpm1, Siae, Vcan, Anxa1, Sbp, Gemin4, Tsn, Hist1h1d,
Ptx3, A2m, Acy1, Mgp, Pgm1, Tnnc2, Apof, Tpm2, Car3, Ptgr1, Ldlr,
Acan, Glrx3, Chi3l1, Nono, Car1, Mpo, Hpx, Fabp4, Mpst, Fn1, Abp1,
Igfbp1, Fabp3, Hspa1l, Col1a2, Retnlg, Rps28, Pfkp, Lcn2, Chad,
Tceb2, S100a11, Hba-a2, Ostf1, Fabp6, Fh1, Myh6, Sema6d, Pgam2,
Lbp, Tpi1, Trim72, Pgls, Hspa5, Pla1a, Ldha, Ckm, Tsku, Hprt1, Ckb,
Pdlim3, Ak1, Itpa, Serping1, Eno3, Tnnc1, Semg1, Gstm1, Hp, Col1a1,
Rpl8, Mb, Atp7a, Orm1, Slpi, S100a8, Hexb, Col6a2, S100a9, Atp5a1,
Limd2, Prg4 |
Following pathway analysis, the 158 proteins were
shown to be mainly involved in extracellular matrix-receptor
interactions (count=7; P<0.01), focal adhesions (count=7;
P<0.05), and complement and coagulation cascades (count=6;
P<0.01; Table VI).
| Table VIProteins in signaling pathway. |
Table VI
Proteins in signaling pathway.
| Title | Gene count | P-value | Genes |
|---|
| ECM-receptor
interaction | 7 | 5.98E-05 |
Thbs1,Col6a3,Fn1,Col1a2,Chad,Col1a1,Col6a2 |
| Complement and
coagulation cascades | 6 | 0.000165 |
F13a1,Kng1,C4bpb,A2m,C4bpa,Serping1 |
| Focal adhesion | 7 | 0.010068 |
Thbs1,Col6a3,Fn1,Col1a2,Chad,Col1a1,Col6a2 |
| Cardiac muscle
contraction | 5 | 0.002137 |
Tpm2,Tpm3,Myh6,Tpm1,Tnnc1 |
| PPAR signaling
pathway | 4 | 0.009466 |
Fabp5,Fabp4,Fabp3,Fabp6 |
| Renin-angiotensin
system | 1 | 0.187962 | Agt |
| Ribosome | 2 | 0.271977 | Rps28,Rpl8 |
| Antigen processing
and presentation | 2 | 0.290114 | Hspa1l,Hspa5 |
| Nucleotide excision
repair | 1 | 0.417476 | Ddb1 |
| Ubiquitin mediated
proteolysis | 2 | 0.484216 | Ddb1,Tceb2 |
| Calcium signaling
pathway | 2 | 0.643433 | Tnnc2,Tnnc1 |
| TGF-β signaling
pathway | 1 | 0.653817 | Thbs1 |
| Hematopoietic cell
lineage | 1 | 0.658099 | Il1r2 |
| Endocytosis | 2 | 0.664494 | Ldlr,Hspa1l |
| Toll-like receptor
signaling pathway | 1 | 0.712848 | Lbp |
| Vascular smooth
muscle contraction | 1 | 0.755917 | Cald1 |
| Lysosome | 1 | 0.758953 | Hexb |
| Axon guidance | 1 | 0.79776 | Sema6d |
| Tight junction | 1 | 0.805238 | Myh6 |
| Cell adhesion
molecules | 1 | 0.80767 | Vcan |
| MAPK signaling
pathway | 2 | 0.848604 | Hspa1l,Il1r2 |
| Regulation of actin
cytoskeleton | 1 | 0.930221 | Fn1 |
| Cytokine-cytokine
receptor interaction | 1 | 0.96282 | Il1r2 |
| Olfactory
transduction | 1 | 0.993046 | Clca1 |
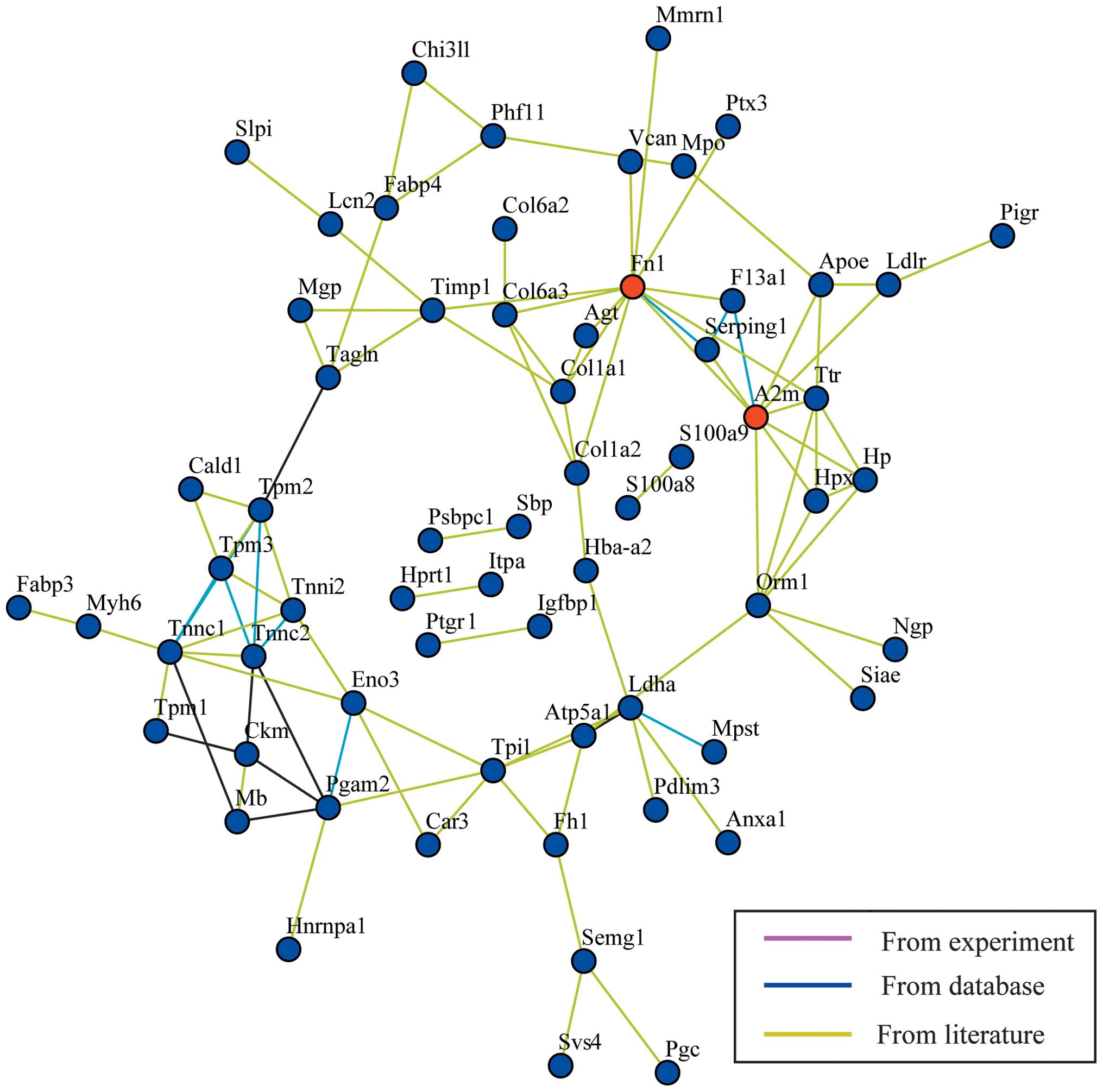
The gene symbols were mapped on a protein-protein
interaction network (Fig. 4). The
closest connection was identified between hub Fn1 and A2m genes.
However, no significant fold-change was found in the Fn1 gene and
A2m-gene associated protein expression. This network only shows the
peripheral interactions between the differentially expressed
proteins, but not the close interactions between individual
proteins. Therefore, further analysis of the time-course data was
performed. Comprehensively considering the fold change (>1.5 or
<0.5) and a P-value <0.05, the biological effects of the
distinct proteins and involvement of proteins in the induction of
inflammation and sepsis, five proteins were selected for further
gene network analysis, including apolipoprotein E (ApoE), annexin
A1 (Anxa1), neutrophil gelatinase-associated lipocalin (NGAL),
S100a8 and S100a9 (Fig. 5).
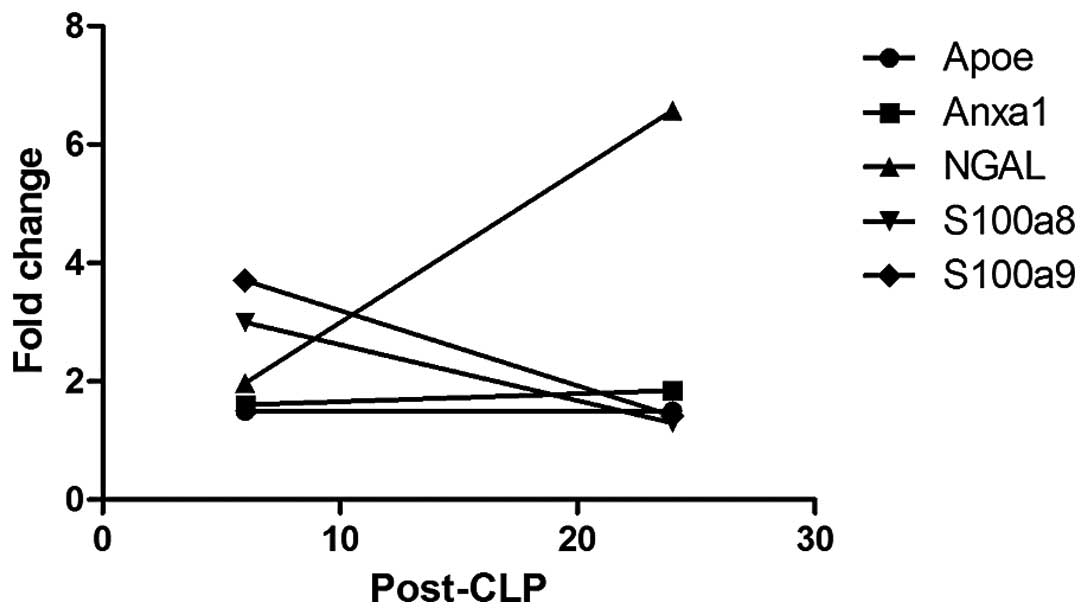
Compared with the sham-6 h group, Anxa1 and ApoE were upregulated
in the CLP-6 h group. There was no significant difference in the
expression of Anxa1 and ApoE between the CLP-24 h and CLP-6 h
groups. S100a8 and S100a9 were overexpressed at 6 h following CLP
compared with the sham-6 h group, while downregulated 24 h after
CLP. In addition, the expression of NGAL was detected to be
elevated in the CLP-6 h (compared with sham-6 h) and CLP-24 h
groups (compared with CLP-6 h group). All of the five proteins were
found to be involved in the same lipid metabolism-associated
network, with amyloidogenic glycoprotein as the hub. The
alterations in the relative abundance of these five proteins
between the CLP-6 h and CLP-24 h groups are demonstrated in
Fig. 6. Among the proteins, NGAL
expression revealed a significant increase from CLP-6 h to CLP-24
h.
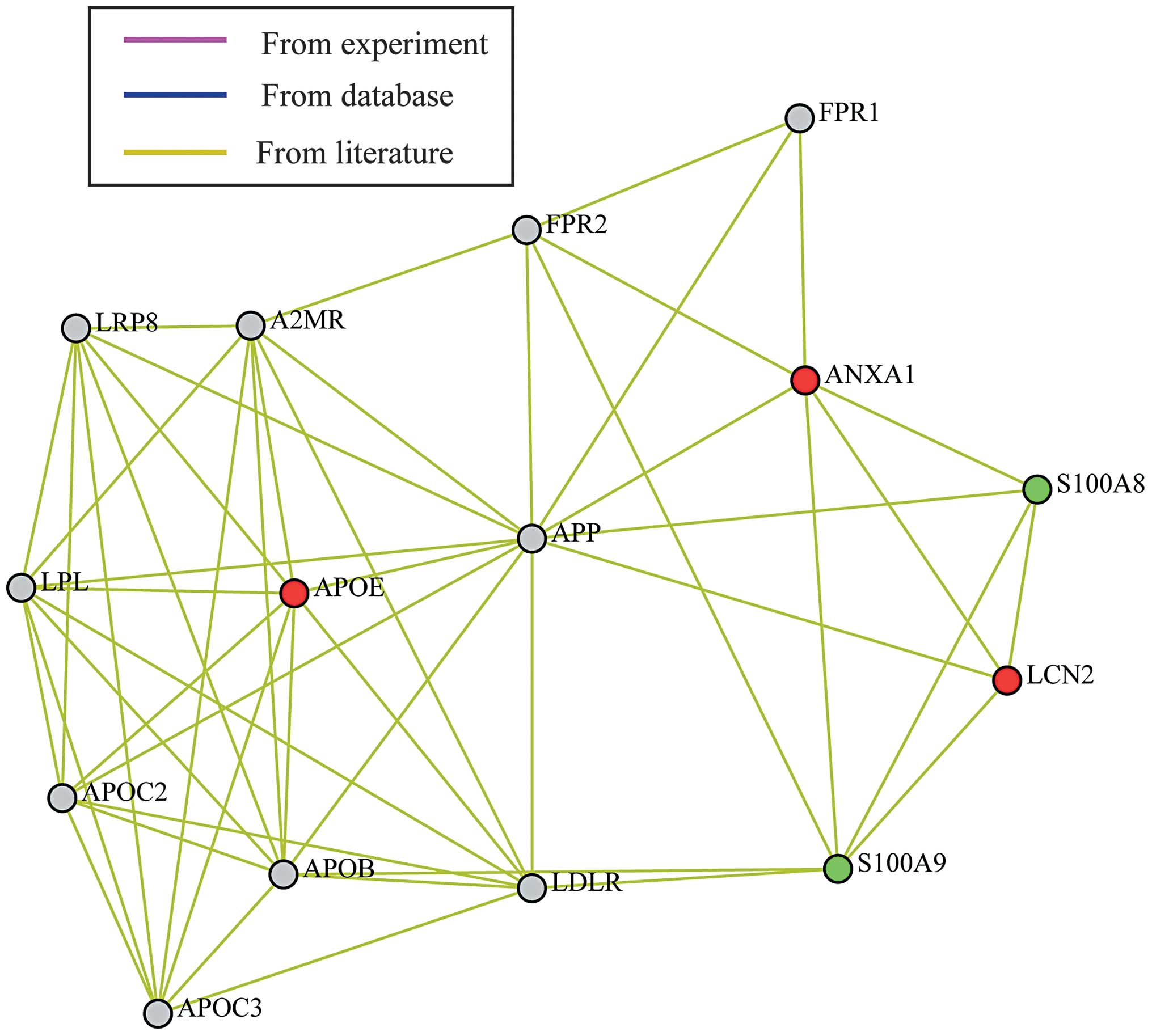 | Figure 5Differentially expressed protein
network analysis (2), using gene
symbols. It is observed that the five selected proteins: ApoE,
Anxa1, NGAL (Lcn2), S100a8 and S100a9 are involved in the same
lipid metabolism-associated network, with amyloidogenic
glycoprotein (APP) as the hub. ApoE, apolipoprotein E; Anxa1,
annexin A1; NGAL, neutrophil gelatinase-associated lipocalin. Red,
upregulated genes (CLP24 vs. CLP-6); green, downregulated genes
(CLP-24 vs. CLP-6). |
Discussion
Although numerous investigations concerning sepsis
have been conducted, the detailed and precise mechanism of sepsis
remains unclear. The present study provided a novel prospective to
aid in the elucidation of the mechanism of sepsis by determining
proteome changes in the mesenteric lymph. The results demonstrated
that a total of 158 proteins in the mesenteric lymph, which were
found to exist in all of the groups, were identified to be
significantly differentially expressed (fold change ≥1.5;
P<0.05). The 158 proteins demonstrated abundant differences
between the CLP and sham groups as well as the CLP-6 h and CLP-24 h
groups. Following analysis of the 158 proteins by bioinformatic
methods, five proteins were investigated as targets: ApoE, Anxa1,
NGAL, S100a8 and S100a9.
ApoE, the ligand of the low density lipoprotein
receptor, has a vital role in lipid metabolism and is involved in
the pathogenesis of critical diseases (26,27).
It has been suggested that ApoE polymorphisms are associated with
severe sepsis in surgical patients (28). In a previous study, the majority of
patients with severe sepsis did not have the APOɛ3
allele. The APOɛ3 allele significantly protected
carriers against the risk of severe sepsis (29). Although the role of ApoE in sepsis
was not clear, ApoE was determined to be associated with a variety
of different factors. In sepsis models induced by CLP, ApoE
reversed the increase of hepatic T cell apoptosis and necrosis, and
promoted Th1 cytokine levels (30). In addition, ApoE−/− mice
demonstrated notably higher levels of proinflammatory cytokines in
the serum, compared with wild-type mice (31,32).
ApoE inhibited the lipopolysaccharide (LPS)-induced increase of
tumor necrosis factor-α, interleukin (IL)-1β and IL-6 in serum and
decreased the mortality rates of mice with sepsis (32). By contrast, another study suggested
that ApoE increased the mortality rates of rats with sepsis
(30). Seven days following CLP,
rats injected with recombinant ApoE3 had a mortality rate of 82.14
and 100% at doses of 114 μg/kg or 1.6 mg/kg, respectively, while
none of the saline-injected rats died (30). The results demonstrated that ApoE
was involved in multiple biological processes, including transport,
stress responses, cell organization and biogenesis. ApoE was also
involved in transporter and enzyme regulator activity. At 6 h
following CLP, ApoE was indicated to be significantly upregulated
in the lymph, but not in the CLP-24 h group. Therefore, it is
suggested that ApoE demonstrated protective effects in the early
stages of sepsis, but that this effect was reversed in the later
stages.
Anxa1, an annexin, is known to be an endogenous
anti-inflammatory mediator (33).
The expression of annexins (AnxA1–AnxA7, AnxA9 and AnxA11) has been
demonstrated to be regulated by pro- and anti-inflammatory
stimulation (34). Furthermore,
pro- and anti-inflammatory responses were revealed to be present
throughout the entire process of sepsis progression (35). Anxa1 decreased the degree of the
acute inflammatory response (36)
and exerted active effects on the suppression of acute inflammation
by preventing the adhesion and migration of neutrophils, adjusting
the production of pro-inflammatory and anti-inflammatory cytokines
and promoting neutrophil apoptosis (37,38).
There was evidence that endogenous Anxa1 exhibited a protective
role in the cerebral microcirculation of sepsis (39). However, it was reported that the
Anxa1 plasma level decreased in patients with sepsis, as compared
with control patients (35). In
the present study, Anxa1 was overexpressed in the lymph of the
CLP-6 h group (fold=1.61, P=0.0103) compared with that in the
sham-6 h group. It indicated that Anxa1 had a key role in sepsis
progression and that the expression of Anxa1 may be different in
the lymph and plasma of patients. Bioinformatics analysis revealed
that Anxa1 was mainly involved in enzyme regulator activity, signal
transduction and cell adhesion, which corresponded with previous
studies.
S100a8 and S100a9 as members of the S100 protein
family, are released from neutrophils and activate phagocytes
during sepsis (40). S100a8 and
S100a9 most commonly emerge as heterodimers and have a prominent
role in the pathogenesis of various inflammatory diseases (41).
S100A8 and/or S100A9 have been reported to possess
multiple important biological activities, including mediating
neutrophil adhesion to fibronectin (42), inducing neutrophil chemotaxis
(43) and mediating apoptosis
(44). It has also been
demonstrated that S100A8 and S100A9 enhanced inflammatory responses
by mediating nuclear factor-κB activation and inducing cytokine
secretion, including IL-6, IL-8 and IL-1β (41). During the early phase of sepsis,
excessive release of proinflammatory cytokines and chemokines
reflect hyperreactive immune responses and result in hyporeactive
immune responses in the later phase along with intractable shock,
refractory infection, multiple organ failure and even mortality
(45). As outlined in a previous
study, the S100a8/S100a9 plasma levels were significantly elevated
in patients with severe sepsis (46). In abdominal sepsis patients, the
S100a8/S100a9 levels in the abdominal fluid were >15-fold that
in the plasma, indicating that S100a8/S100a9 expression was induced
primarily at the site of infection during sepsis (46). The present study indicated that the
expression of S100a8 and S100a9 were significantly elevated in the
lymph during early stages of sepsis (CLP-6h) but decreased in later
stages (CLP-24h). As S100 proteins have been defined as clinical
markers of inflammation in a previous study (47), S100a8 and S100a9 levels in the
lymph may be a marker for the diagnosis of early stage sepsis.
NGAL, a lipocalin, is overexpressed in acute kidney
injury due to ischemia, toxic factors and sepsis (48,49).
In patients with sepsis, the overexpression of NGAL attributed to
the response of the kidneys to sepsis. NGAL expression was found to
be elevated in the serum of patients with sepsis (50). The concentration of urinary NGAL
was also considered to be an indicator of early acute kidney injury
in patients with sepsis (49).
Furthermore, NGAL levels in the urine were reported to be
correlated with the plasma or serum levels (51). In the present study, NGAL levels in
the lymph increased progressively and reached a peak at 24 h
following CLP. The NGAL levels in the plasma and urine have been
widely adopted as an early predictor of acute kidney injury
(52–54). NGAL had a critical role in sepsis
progression and further studies are required to determine its
clinical utility as a diagnostic marker for sepsis.
In conclusion, the present study demonstrated that
the expression of five proteins (ApoE, Anxa1, NGAL, protein S100a8
and S100a9) was significantly elevated in the progression of
sepsis. All five proteins appeared to have vital roles in critical
disease development and may therefore be potential targets for the
treatment and diagnosis of sepsis. The present study provides a
novel perspective to aid in the understanding of the pathological
mechanism of sepsis.
Acknowledgements
This study was supported by the Shanghai Key
Discipline Construction of Public Health Funds (Shanghai, China)
(grant no. O8GWZX1103). The authors would like to thank Shanghai
Sensichip Co., Ltd. (Shanghai, China) for the proteomic and
bioinformatics analysis and Dr. Karen Bysouth who provided editing
services on behalf of Elixigen Co. (Shanghai, China).
References
|
1
|
Hinkelbein J, Kalenka A, Schubert C,
Peterka A, Feldmann J and Robert E: Proteome and metabolome
alterations in heart and liver indicate compromised energy
production during sepsis. Protein Pept Lett. 17:18–31. 2010.
View Article : Google Scholar
|
|
2
|
Mckean SC, Ross JJ, Dressler DD, Brotman
DJ and Ginsberg J: Principles and practice of hospital medicine.
McGraw-Hill; 2012
|
|
3
|
Angus DC and van der Poll T: Severe sepsis
and septic shock. N Engl J Med. 369:840–851. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dellinger RP, Carlet JM, Masur H, et al:
Surviving Sepsis Campaign guidelines for management of severe
sepsis and septic shock. Intensive Care Med. 30:536–555. 2004.
View Article : Google Scholar
|
|
5
|
Dellinger RP, Levy MM, Carlet JM, et al:
Surviving Sepsis Campaign: international guidelines for management
of severe sepsis and septic shock: 2008. Intensive Care Med.
34:17–60. 2008. View Article : Google Scholar
|
|
6
|
Dellinger RP, Levy MM, Rhodes A, et al:
Surviving Sepsis Campaign: international guidelines for management
of severe sepsis and septic shock, 2012. Intensive Care Med.
39:165–228. 2013. View Article : Google Scholar
|
|
7
|
Okazaki Y and Matsukawa A: Pathophysiology
of sepsis and recent patents on the diagnosis, treatment and
prophylaxis for sepsis. Recent Pat Inflamm Allergy Drug. 3:26–32.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Magnotti LJ, Upperman JS, Xu DZ, Lu Q and
Deitch EA: Gut-derived mesenteric lymph but not portal blood
increases endothelial cell permeability and promotes lung injury
after hemorrhagic shock. Ann Surg. 228:518–527. 1998. View Article : Google Scholar
|
|
9
|
Mittal A1, Phillips AR, Middleditch M, et
al: The proteome of mesenteric lymph during acute pancreatitis and
implications for treatment. JOP. 10:130–142. 2009.PubMed/NCBI
|
|
10
|
Dayal SD, Haskó G, Lu Q, et al:
Trauma/hemorrhagic shock mesenteric lymph upregulates adhesion
molecule expression and IL-6 production in human umbilical vein
endothelial cells. Shock. 17:491–495. 2002. View Article : Google Scholar
|
|
11
|
Moore EE: Claude H. Organ, Jr. memorial
lecture: splanchnic hypoperfusion provokes acute lung injury via a
5-lipoxygenase-dependent mechanism. Am J Surg. 200:681–689. 2010.
View Article : Google Scholar
|
|
12
|
Leak LV, Liotta LA, Krutzsch H, et al:
Proteomic analysis of lymph. Proteomics. 4:753–765. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mittal A, Middleditch M, Ruggiero K, et
al: The proteome of rodent mesenteric lymph. Am J Physiol
Gastrointest Liver Physiol. 295:G895–G903. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mittal A, Phillips A, Middleditch M, et
al: The proteome of mesenteric lymph during acute pancreatitis and
implications for treatment. JOP. 10:130–142. 2009.PubMed/NCBI
|
|
15
|
Clement CC, Aphkhazava D, Nieves E, et al:
Protein expression profiles of human lymph and plasma mapped by
2D-DIGE and 1D SDS-PAGE coupled with nanoLC-ESI-MS/MS bottom-up
proteomics. J Proteomics. 78:172–187. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mcdunn JE, Townsend RR and Cobb JP: The
murine plasma protein response to polymicrobial intra-abdominal
sepsis. Proteomics Clin Appl. 1:373–386. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren Y, Wang J, Xia J, et al: The
alterations of mouse plasma proteins during septic development. J
Proteome Res. 6:2812–2821. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dear JW, Leelahavanichkul A, Aponte A, et
al: Liver proteomics for therapeutic drug discovery: inhibition of
the cyclophilin receptor CD147 attenuates sepsis-induced acute
renal failure. Crit Care Med. 35:2319–2328. 2007. View Article : Google Scholar
|
|
19
|
Duan X, Berthiaume F, Yarmush D and
Yarmush ML: Proteomic analysis of altered protein expression in
skeletal muscle of rats in a hypermetabolic state induced by burn
sepsis. Biochem J. 397:149–158. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Otero-Antón E, González-Quintela A,
López-Soto A, López-Ben S, Llovo J and Pérez L: Cecal ligation and
puncture as a model of sepsis in the rat: influence of the puncture
size on mortality, bacteremia, endotoxemia and tumor necrosis
factor alpha levels. Eur Surg Res. 33:77–79. 2001.PubMed/NCBI
|
|
21
|
Freise H, Brückner U and Spiegel H: Animal
models of sepsis. J Invest Surg. 14:195–212. 2001. View Article : Google Scholar
|
|
22
|
Rittirsch D, Huber-Lang MS, Flierl MA and
Ward PA: Immunodesign of experimental sepsis by cecal ligation and
puncture. Nat Protoc. 4:31–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Y, Wang J, Li Y and Lin ZF: A modified
rat model for cannulation and collection of thoracic duct lymph.
Lymphology. 44:82–84. 2011.PubMed/NCBI
|
|
24
|
Mittal A, Middleditch M, Ruggiero K, et
al: Changes in the mesenteric lymph proteome induced by hemorrhagic
shock. Shock. 34:140–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Neil AL and Christensen H: Australian
school-based prevention and early intervention programs for anxiety
and depression: a systematic review. Med J Aust. 186:305–308.
2007.PubMed/NCBI
|
|
26
|
Bonomini F, Filippini F, Hayek T, et al:
Apolipoprotein E and its role in aging and survival. Exp Gerontol.
45:149–157. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Seshasai RK, Katz R, de Boer IH, et al:
Apolipoprotein E and kidney function in older adults. Clin Nephrol.
78:174–180. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moretti EW, Morris RW, Podgoreanu M, et
al: APOE polymorphism is associated with risk of severe sepsis in
surgical patients. Crit Care Med. 33:2521–2526. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moretti EW, Morris RW, Podgoreanu M, et
al: Perioperative Genetics and Safety Outcomes Study (PEGASUS)
Investigative Team: APOE polymorphism is associated with risk of
severe sepsis in surgical patients. Crit Care Med. 33:2521–2526.
2005. View Article : Google Scholar
|
|
30
|
Kattan OM, Kasravi FB, Elford EL, Schell
MT and Harris HW: Apolipoprotein E-mediated immune regulation in
sepsis. J Immunol. 181:1399–1408. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
de Bont N, Netea MG, Demacker PN, et al:
Apolipoprotein E knock-out mice are highly susceptible to
endotoxemia and Klebsiella pneumoniae infection. J Lipid
Res. 40:680–685. 1999.PubMed/NCBI
|
|
32
|
Van Oosten M, Rensen PC, Van Amersfoort
ES, et al: Apolipoprotein E protects against bacterial
lipopolysaccharide-induced lethality. A new therapeutic approach to
treat gram-negative sepsis. J Biol Chem. 276:8820–8824. 2001.
|
|
33
|
Kamal AM, Flower RJ and Perretti M: An
overview of the effects of annexin 1 on cells involved in the
inflammatory process. Mem Instit Oswaldo Cruz. 100(Suppl 1): 39–47.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kreft S: Annexinabhängige modulation der
immunantwort und hämostase. Universität zu Köln; 2013
|
|
35
|
Tsai WH, Shih CH, Yu YB and Hsu HC: Plasma
levels in sepsis patients of annexin A1, lipoxin A4, macrophage
inflammatory protein-3a, and neutrophil gelatinase-associated
lipocalin. J Chin Med Assoc. 76:486–490. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Flower RJ: Eleventh Gaddum memorial
lecture. Lipocortin and the mechanism of action of the
glucocorticoids. Br J Pharmacol. 94:987–1015. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vago JP, Nogueira CR, Tavares LP, et al:
Annexin A1 modulates natural and glucocorticoid-induced resolution
of inflammation by enhancing neutrophil apoptosis. J Leukoc Biol.
92:249–258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Perretti M: Editorial: to resolve or not
to resolve: Annexin A1 pushes resolution on track. J Leukoc Biol.
92:245–247. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gavins FN, Hughes EL, Buss NA, et al:
Leukocyte recruitment in the brain in sepsis: involvement of the
annexin 1-FPR2/ALX anti-inflammatory system. FASEB J. 26:4977–4989.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nacken W, Roth J, Sorg C and Kerkhoff C:
S100A9/S100A8: Myeloid representatives of the S100 protein family
as prominent players in innate immunity. Microsc Res Tech.
60:569–580. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Simard J-C, Cesaro A, Chapeton-Montes J,
et al: S100A8 and S100A9 induce cytokine expression and regulate
the NLRP3 inflammasome via ROS-dependent activation of NF-κB(1.).
PLoS One. 8:e721382013.PubMed/NCBI
|
|
42
|
Anceriz N, Vandal K and Tessier PA: S100A9
mediates neutrophil adhesion to fibronectin through activation of
β2 integrins. Biochem Biophys Res Commun. 354:84–89.
2007.PubMed/NCBI
|
|
43
|
Ryckman C, Vandal K, Rouleau P, et al:
Proinflammatory activities of S100: proteins S100A8, S100A9, and
S100A8/A9 induce neutrophil chemotaxis and adhesion. J Immunol.
170:3233–3242. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yui S, Nakatani Y and Mikami M:
Calprotectin (S100A8/S100A9), an inflammatory protein complex from
neutrophils with a broad apoptosis-inducing activity. Biol Pharm
Bull. 26:753–760. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Riedemann NC, Guo RF and Ward PA: Novel
strategies for the treatment of sepsis. Nat Med. 9:517–524. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
van Zoelen MA, Vogl T, Foell D, et al:
Expression and role of myeloid-related protein-14 in clinical and
experimental sepsis. Am J Respir Crit Care Med. 180:1098–1106.
2009.PubMed/NCBI
|
|
47
|
Foell D, Frosch M, Sorg C and Roth J:
Phagocyte-specific calcium-binding S100 proteins as clinical
laboratory markers of inflammation. Clin Chim Acta. 344:37–51.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mishra J, Dent C, Tarabishi R, et al:
Neutrophil gelatinase-associated lipocalin (NGAL) as a biomarker
for acute renal injury after cardiac surgery. Lancet.
365:1231–1238. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bagshaw SM, Bennett M, Haase M, et al:
Plasma and urine neutrophil gelatinase-associated lipocalin in
septic versus non-septic acute kidney injury in critical illness.
Intensive Care Med. 36:452–461. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Parravicini E, Nemerofsky SL, Michelson
KA, et al: Urinary neutrophil gelatinase-associated lipocalin is a
promising biomarker for late onset culture-positive sepsis in very
low birth weight infants. Pediatr Res. 67:636–640. 2010. View Article : Google Scholar
|
|
51
|
Uttenthal L: NGAL: a marker molecule for
the distressed kidney. Clin Lab Int. 29:39–41. 2005.
|
|
52
|
Gupta A, Berg DT, Gerlitz B, et al: Role
of protein C in renal dysfunction after polymicrobial sepsis. J Am
Soc Nephrol. 18:860–867. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Peng ZY, Wang H-Z, Srisawat N, et al:
Bactericidal antibiotics temporarily increase inflammation and
worsen acute kidney injury in experimental sepsis. Crit Care Med.
40:538–543. 2012. View Article : Google Scholar
|
|
54
|
Seija M, Baccino C, Nin N, et al: Role of
peroxynitrite in sepsis-induced acute kidney injury in an
experimental model of sepsis in rats. Shock. 38:403–410. 2012.
View Article : Google Scholar : PubMed/NCBI
|