Introduction
Bronchopulmonary dysplasia (BPD) is the most common
form of chronic lung disorder in babies born prematurely and causes
ongoing morbidity and mortality, characterized by impaired alveolar
development and inflammation (1).
There is currently no effective intervention with which to prevent
or treat bronchopulmonary dysplasia (2). Stem cell therapy is a potential
therapeutic approach for BPD and studies have shown that BMSCs
protect the lung and aid in the repair of injured lung tissues in a
number of animal models (3,4).
Studies have also demonstrated that intravenous or intratracheal
administration of BMSCs protects against hyperoxic lung injury by
reducing inflammation and improving alveolar structure (5,6). To
date, the mechanisms underlying these effects remain elusive. It is
likely that paracrine-mediated mechanisms and immunomodulation
(7) are involved, amongst other
unidentified processes.
Transforming growth factor-β (TGF-β) signaling and
the pathways in which it is involved are known to be important
during lung development (8).
Increased expression of TGF-β inhibits development of the alveoli
and also causes pulmonary fibrosis in lungs with advanced
alveolarization. Overexpression of TGF-β leads to structural
changes associated with BPD, including the proliferation of
α-actin-positive myofibroblasts within the alveolar septal walls,
the development of abnormal alveolar structure and vascular
development in the neonatal mouse lung (9,10).
Vascular endothelial growth factor (VEGF) is also known to be
involved in the regulation of pulmonary vascular growth and
development, the stimulation of angiogenesis and the promotion of
endothelial survival (11).
Inhibition of VEGF receptor-2 (VEGFR2) in neonatal rats causes
rarefaction of pulmonary vessels and impairs alveolar formation
(12), whereas enhancement of VEGF
signaling ameliorates the alveolar disruption induced by hyperemia
(13). The present study
investigated the effect of BMSCs in the treatment of lung injury in
a mouse model of BPD and examined whether the underlying mechanism
was via the abovementioned signaling pathways.
Materials and methods
Animal
Neonatal C57BL/6 mice (24 h old; weight, 1–2 g;
n=15), used in all experiments, were obtained from the Animal
Experiment Center of Shandong University (Shandong, China). The
animals were kept in a temperature-controlled environment (22–24°C)
under a 12-h light/dark cycle with access to food and water ad
libitum. Mice were randomized into various groups. They were
then weighed and blood samples were collected. All animal
procedures were approved by the animal ethics committee of Shandong
University (Jinan, China) and followed the Guide for the Care and
Use of Laboratory Animals published by the US National Institutes
of Health (NIH Publication no. 85-23, revised 1996).
Culture, immunophenotype analysis and
lentiviral transfection with green fluorescent protein (GFP) of
BMSCs
BMSCs were isolated from the femur of six to
eight-week-old C57BL/6 mice, as previously described (14). Briefly, fresh bone marrow cells
were isolated by flushing Dulbecco’s modified Eagle’s medium (DMEM;
Gibco-BRL, Carlsbad, CA, USA) containing 1% penicillin-streptomycin
(Sigma-Aldrich, St Louis, MO, USA) through the medullary cavity of
mouse femurs. Cells were isolated using the Ficoll (1.077;
Sigma-Aldrich) density gradient centrifugation method. The isolated
bone marrow cells were then cultured and expanded in a low-glucose
culture containing DMEM with F-12 (LG-DMEM; Gibco-BRL, Life
Technologies, Carlsbad, CA, USA), supplemented with 10% fetal
bovine serum (Hyclone, Logan, UT, USA), 100 U/ml penicillin and 100
g/ml streptomycin (Sigma-Aldrich). All cultures were maintained at
37°C in 5% humidified CO2. Characteristic
immunoreactivity for cell markers was detected using fluorescent
activated cell sorting analysis (FACS; FACScan BD LSRFortessa; BD
Biosciences, Franklin Lakes, NJ, USA) with rat monoclonal
anti-mouse anitbodies for CD44, CD117, CD34 and CD106 (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). Briefly, cells growing on a
poly-l-lysine-coated
24-well plate (Sigma-Aldrich) were washed three times with
phosphate-buffered saline (PBS; Sigma-Aldrich) and fixed in 4%
paraformaldehyde for 30 min. Cells were then permeabilized with
0.1% Triton X-100 in PBS for 20 min and blocked with 4% goat serum
for 1 h. Cells were incubated with fluorescein
isothiocyanate-conjugated goat-anti-rat secondary antibodies for 1
h at room temperature in the absence of sunlight.
GFP was cloned into a lentiviral vector using the
Ubiquitin C promoter, as described previously (15). In brief, virus particles were
packaged in 293FT cells (Shanghai Genechem Co., Ltd., Shanghai,
China) in vitro and condensed by ultracentrifugation (600 ×
g, 5 min). Green fluorescence was observed after 48 h using a
fluorescence microscope (Nikon Elcipse 90i; Nikon Corp., Tokyo,
Japan), and the medium was replaced every two days with fresh
medium. The cells were transferred when they reached 80%
confluence.
Preparation of a mouse model of BPD
mice
The mice were prepared as previously described
(16). Briefly, C57BL/6 mice (n=5)
were placed in chambers, in which the oxygen concentration was
maintained at FiO2 = 0.60. Exposure to hyperoxia was
continuous, and mice were maintained in a hypoxic environment for
14 d, with a brief interruption for animal care (less than 10
min/day). BMSCs (1×106) were administered intravenously
at 1 h prior to establishment of the BPD model.
Lung histology and morphometric
analysis
Lungs were prepared for histological examination as
previously described (17).
Briefly, mice were sacrificed by intraperitoneal injection of
pentobarbital (100 mg/kg; Sigma-Aldrich) when they were 14 days
old. Lungs were fixed with 4% paraformaldehyde solution overnight,
and the left lower lobe was embedded in paraffin. Sections were cut
with a microtome set at 4–5 μm (Leica RM226, Leica Microsystems,
Wetzlar, Germany). Alveolarization was assessed by performing
radial alveolar counts (RAC) according to standard methods as
previously described (17).
Briefly, from the center of the respiratory bronchiole a
perpendicular line was drawn to the edge of the acinus (as defined
by a connective tissue septum or the pleura). The number of septa
intersected by this line was counted. Five counts were performed
for each animal; the average of the five high-power fields (hpf)
was randomly selected. These experiments were conducted by two
examiners, blinded to treatment assignment.
Immunohistochemical staining
At 14 days following the operation, mice were
anesthetized, lungs were fixed in 4% paraformaldehyde/PBS for 24 h
and then stored in 70% ethanol. The left lower lobe was embedded in
paraffin, cut into sections of 4–5 μm, deparaffinized in xylene and
then rehydrated by serial immersions in 100% ethanol, 95% ethanol,
85% ethanol, 75% ethanol (Sigma-Aldrich) and 100% water. Slides
were blocked with 4% normal goat serum in PBS, then incubated with
primary antibodies overnight at 4°C and a biotinylated secondary
antibody for 20 min. Immunohistochemistry and immunofluorescence
were conducted with rabbit monoclonal anti-mouse TGF-β1 (1:100;
Santa Cruz Biotechnology, Inc.), VEGF (1:150; Abcam, Cambridge, MA,
USA) and vWF (1:150; Abcam) immunoglobulin G. Fluorescein
isothiocyanate-conjugated goat anti-rabbit secondary antibodies
were then used (1:200, Abcam, Cambridge, UK). Staining was
quantified using the image-analysis system Image Pro Plus 6.0
(Media Cybernetics, Rockville, MD, USA) as previously described
(18).
Quantitative reverse
transcription-polymerase chain reaction (qRT-PCR) analysis
Once the lungs had been harvested, total RNA was
extracted from lung tissue homogenates (Invitrogen Life
Technologies). qRT-PCR analysis was conducted as previously
described (18) in order to detect
the relative pulmonary expression levels of TGF-β1 and VEGF. RNA
samples were dissolved in RNase-free water and quantified
spectrophotometrically (BiochromDNA UV-Vis, WPA; Biochrom, Ltd.,
Cambridge, UK). Primers were designed using the Primer Express
software package (Applied Biosystems, Foster City, CA, USA). The
following primers were used: Forward, 5′-AGGTAACGCCAGGAATGTTGCTA-3′
and reverse, 5′-CATTGCTGTCCCGTGCAGA-3′ for TGF-β1; forward,
5′-CTGCTCTCCTGGGTGCATTG-3′ and reverse, 5′-ACTCCTGGAAGATGTCCACCA-3′
for VEGF; and forward, 5′-ACTCTGGCAAAGTGGATATTGTCG-3′ and reverse,
5′-CAGCATCACCCCATTTGATG-3′ for GAPDH. Data were analyzed using the
ABI Prism 7900 sequence detection system software (version 2.2;
Applied Biosystems).
Statistical analysis
All data are expressed as the mean ± standard error
of the mean. Comparisons of parameters between two groups were made
using unpaired Student’s t-test. Statistical analysis was
conducted using the SPSS 13.0 software (SPSS, Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Characterization of cultured BMSCs and
transfection with GFP
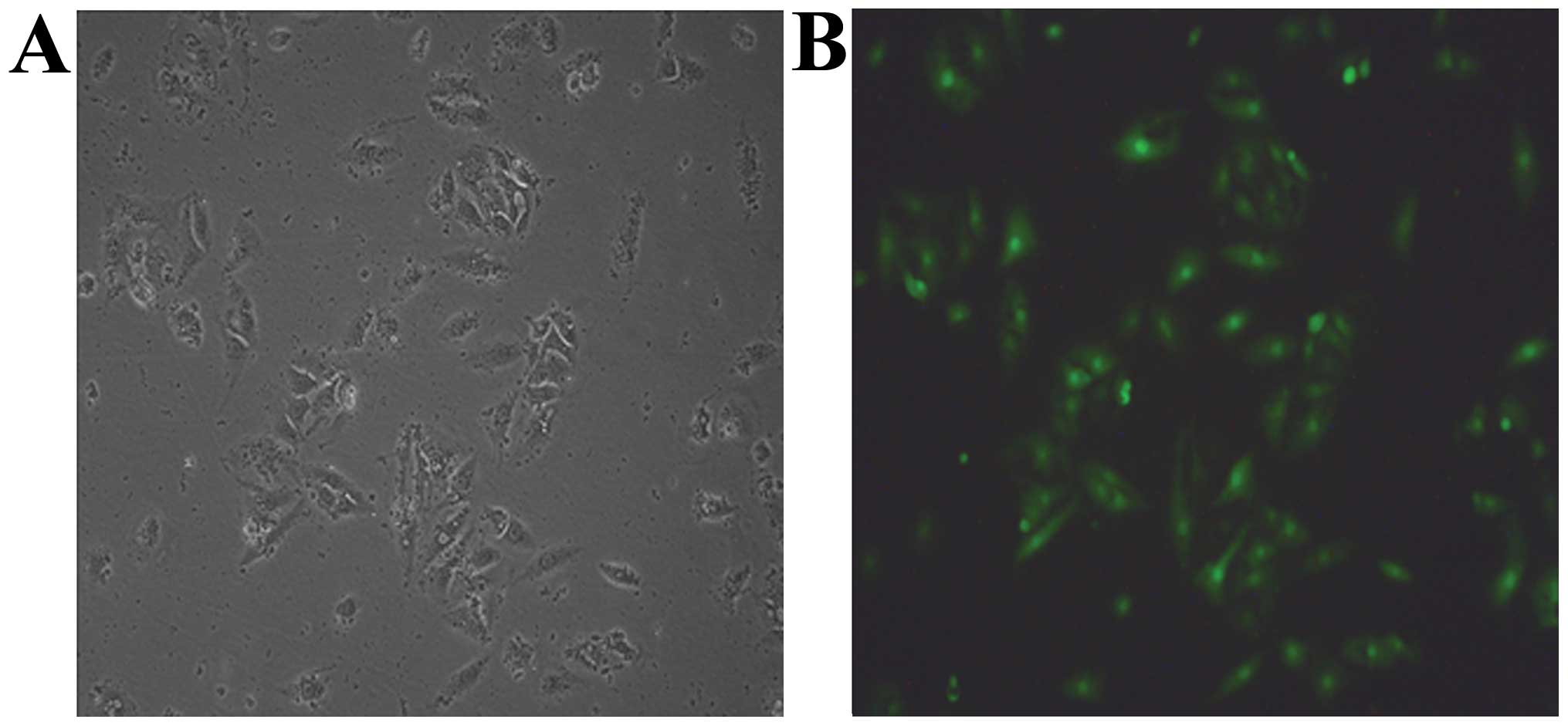
Following primary culture, BMSCs appeared as
spindle-like cells attached to the tissue culture dishes. Three
days after being subcultured, cells were observed to be tightly
adherent to the culture dish and proliferated rapidly in the
culture medium. GFP-transfected cells from C57BL/6 mice expressed
green fluorescence at day 2 (Fig.
1). The surface markers of BMSCs, determined by FAC, showed
that BMSCs were positive for CD44 and CD106, but negative for CD34
and CD117.
Body weight
Each time-point in each study group was represented
by five animals. The average body weight was lower in the BPD group
at 14 days of age (8.73±0.27 g) compared with that of the control
group at the same age (10.96±0.21 g; P<0.05). The group treated
with BMSCs exhibited a higher average body weight (9.18±0.26 g) in
comparison with that in the BPD group (P<0.05; Table I).
 | Table IBody weight in each group (n=5). |
Table I
Body weight in each group (n=5).
| Weight (g) |
|---|
|
|
|---|
| Group | Day 0 | Day 14 |
|---|
| Control | 1.93±0.19 | 10.96±0.23 |
| BPD | 1.92±0.16 | 8.73±0.27a |
| BMSCs | 1.92±0.20 | 10.96±0.21a,b |
| P-value | >0.05 | <0.05 |
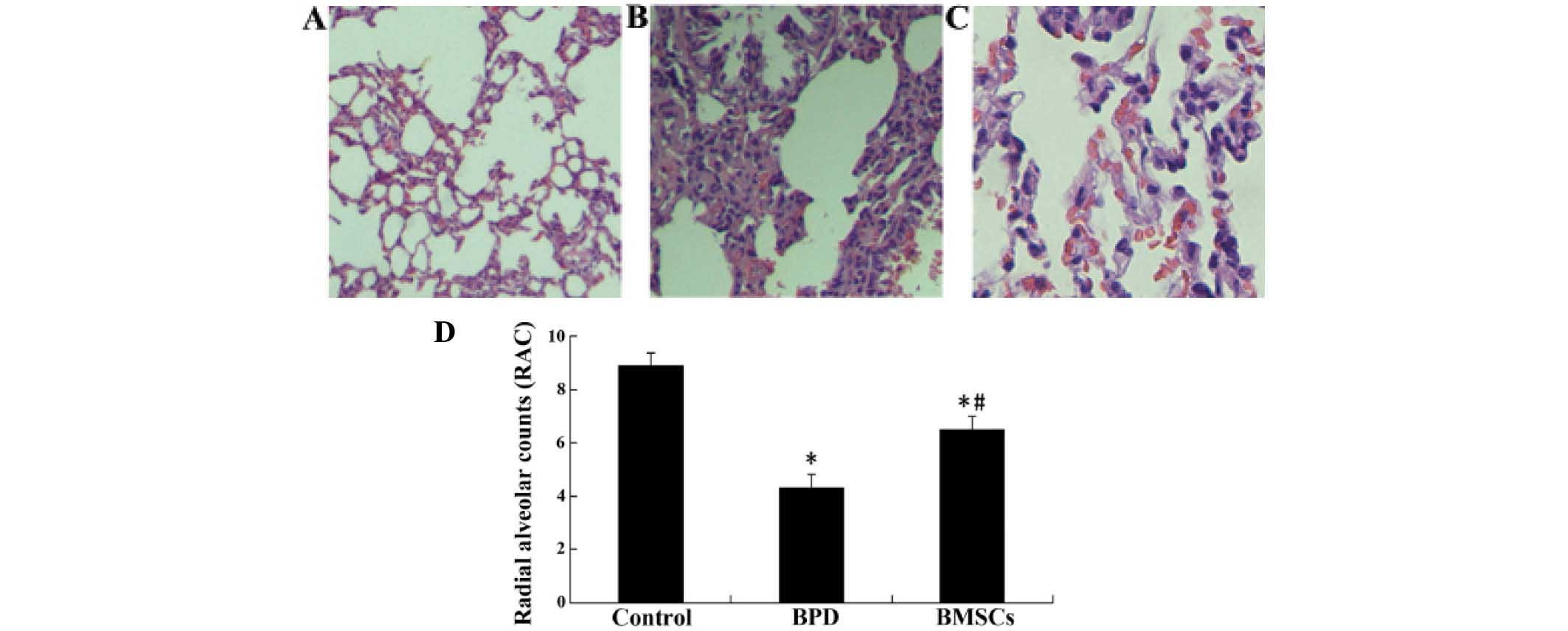
Lung histology and morphometrics
In order to investigate whether BMSCs produce
beneficial effects on hyperoxia-induced impairments in lung
structure, a morphometric analysis was conducted using RAC. The RAC
were significantly lower in the BPD group than they were in the
control group. However, they were significantly higher in the group
treated with BMSCs than those in the BPD group, 14 days of age
(P<0.05; Fig. 2).
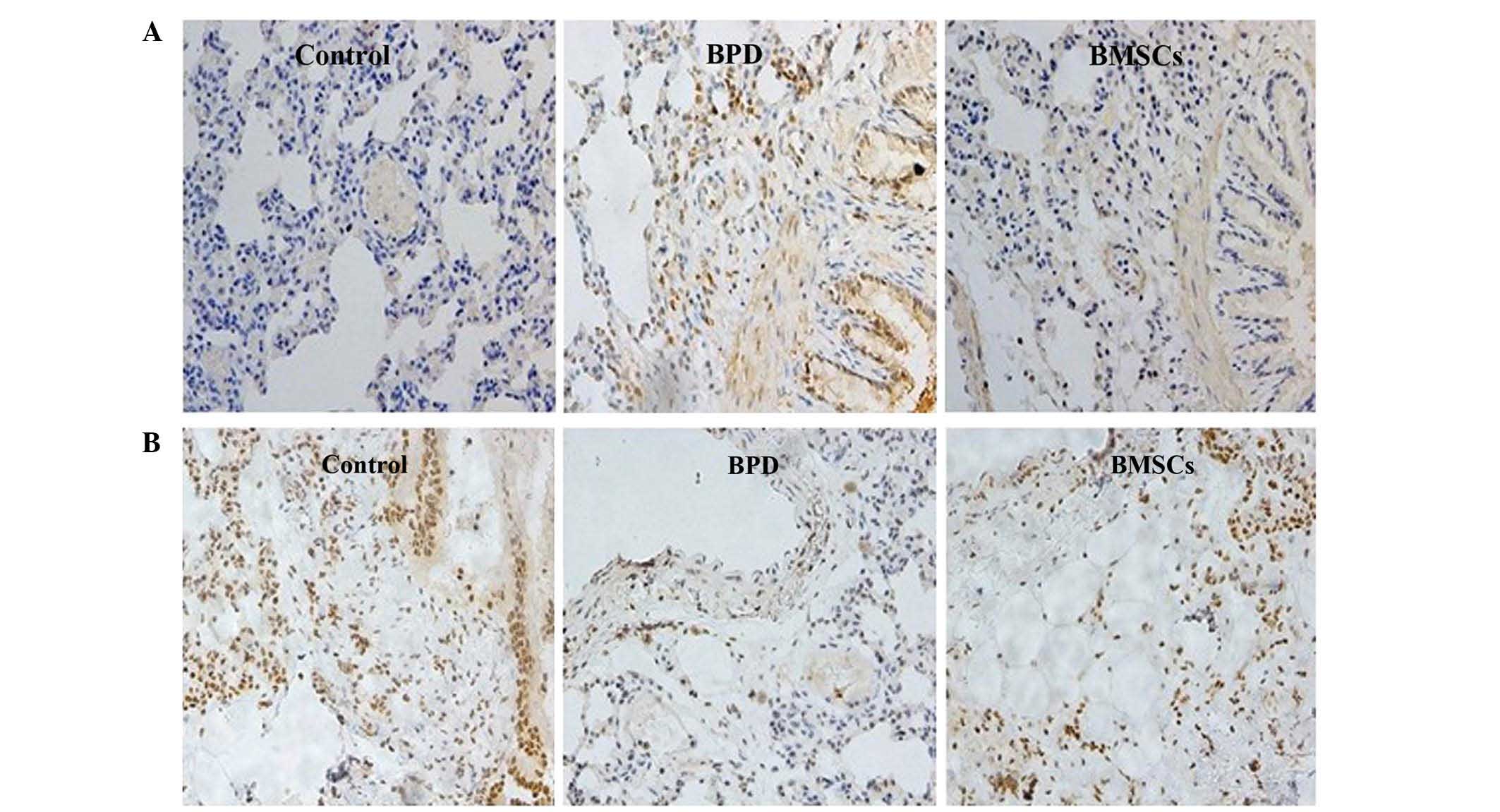
TGF-β1 and VEGF protein expression in
lung tissue
Immunohistochemical staining analysis demonstrated
that exposure of neonatal mice to hyperoxia resulted in a marked
increase in expression of the TGF-β1 protein in the lung, but a
decrease in VEGF expression at 14 days of age. These changes in
protein levels proteins were visibly abrogated at day 14 following
treatment with BMSCs (Fig. 3).
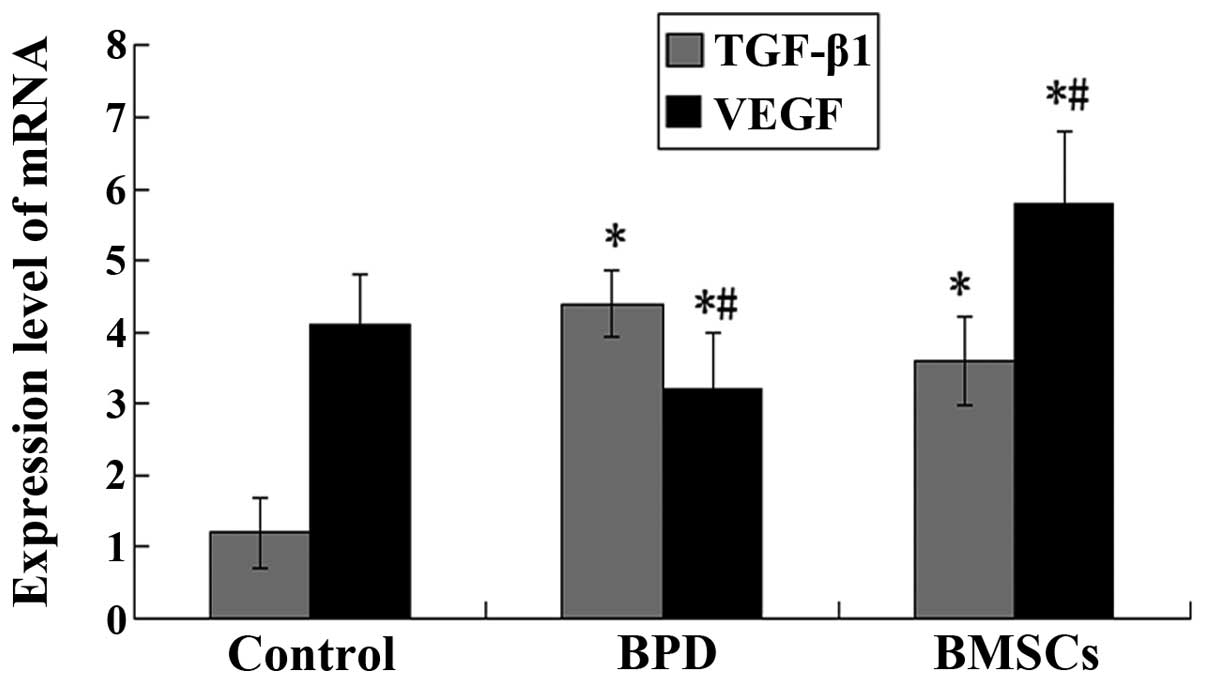
TGF-β1 and VEGF mRNA expression in lung
tissue
The results from the qRT-PCR analysis showed that
the levels of VEGF mRNA in the lungs were significantly lower, and
those of TGF-β1 mRNA were significantly higher in the BPD group
compared with those in the control group at 14 days of age. TGF-β1
levels were significantly decreased in the group treated with BMSCs
compared with those in the BPD group (P<0.05; Fig. 4).
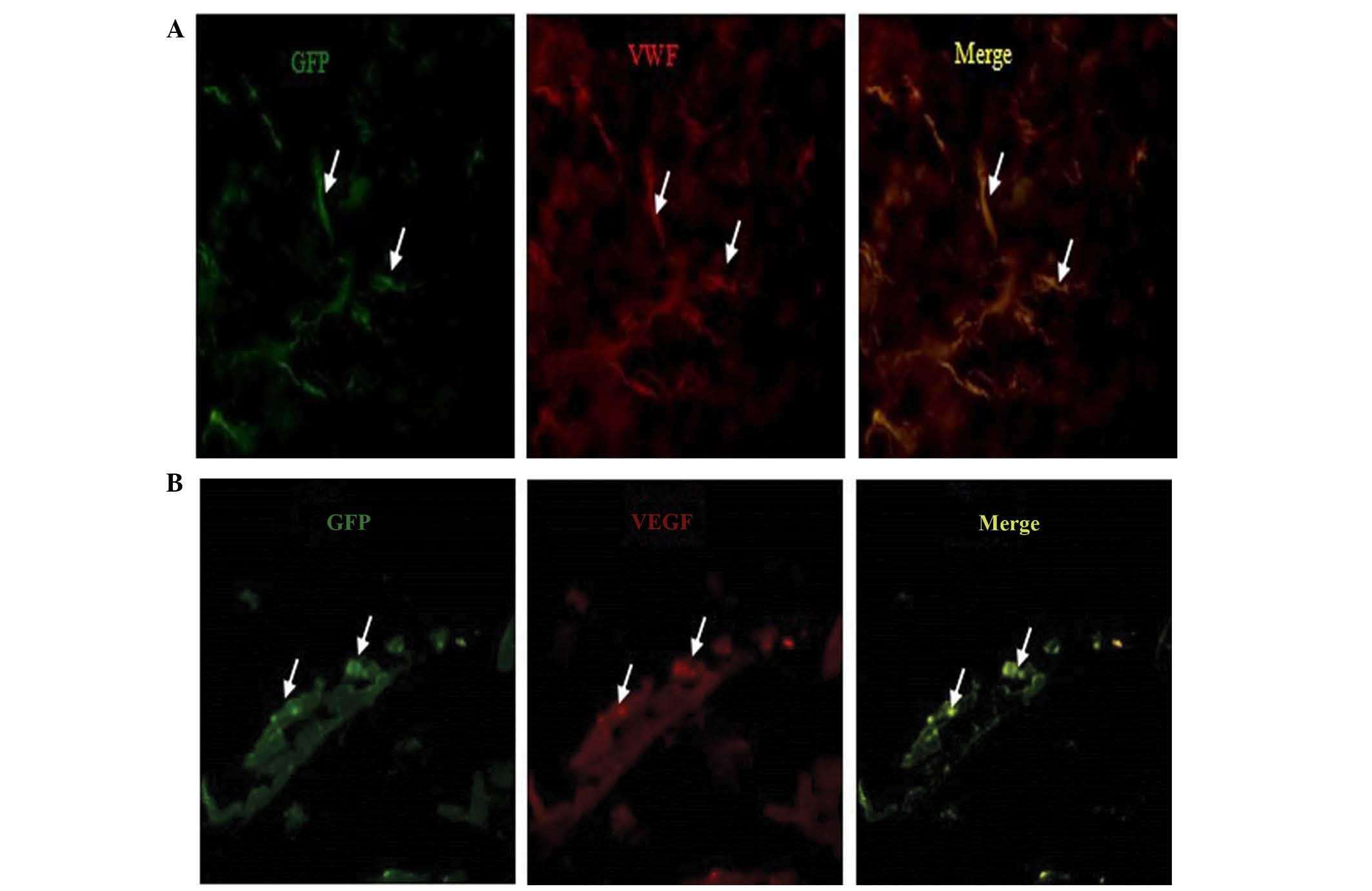
Identification of the injected BMSCs
At 14 days following injection of the cells into
C57BL/6 mice, immunocytochemistry staining for vessel markers, vWF
(red) and VEGF (red), in lung tissue was conducted. The images were
captured with a camera system connected to a fluorescence
microscope. The results indicated that the injected BMSCs were able
differentiate into vascular endothelial cells in vivo
(Fig. 5).
Discussion
The results of the present study showed that
intravenous implantation of BMSCs (×106) improved the
lung impairment caused by exposure to hyperoxia. Treatment with
BMSCs in a mouse model of BPD significantly increased the average
body weight and improved the airway structure in hyperoxia-damaged
lungs. Administration of BMSCs also resulted in a reduced
expression of TGF-β1 and an increased expression of VEGF compared
with those in the BPD group. Immunofluorescence staining indicated
that intravenously implanted BMSCs were capable of differentiating
into cells expressing vWF and VEGF, which are markers of vascular
tissues, at 2 weeks in vivo post-injection. These results
may provide novel therapeutic avenues for lung diseases that are
currently lacking efficient treatments.
Intravenous or intra-alveolar administration of
BMSCs attenuates the severity of lung damage following bleomycin
and endotoxin-induced lung injury in adult rats (19,20).
Recent studies have indicated that BMSCs may prevent arrested
alveolar and vascular growth, in part through paracrine activity,
in experimental neonatal lung injury models in rats (5). Potential mechanisms by which
treatment with BMSCs improves lung structure include engraftment,
anti-inflammatory and immunomodulatory functions, and antiapoptotic
effects. However, there are a number of questions remaining
(21), which require further
investigation, and little is currently known regarding the
potential role of BMSCs in the setting of neonatal lung injury
(22). BMSCs migrate to, or
participate in the development of, lung tissue (23–25).
A number of studies have demonstrated that stem/progenitor cells
have the potential to be used as cellular therapies to contribute
to lung repair mechanisms following acute lung injury.
TGF-β and its downstream mediators have been
implicated in the development of BPD (26,27),
and TGF-β signaling is important in normal lung development and the
lung injury/repair response. TGF-β signaling is initiated by
ligand-induced serine/threonine receptor kinases and
phosphorylation of the cytoplasmic signaling molecules, the Smad
family. BPD may result from abnormal differentiation of alveolar
mesenchymal progenitor cells into myofibroblasts under the
influence of TGF-β. The findings of the present study support the
hypothesis that TGF-β may be involved in BPD. VEGF is also known to
be important in the regulation of pulmonary vascular growth and
development, and for stimulating angiogenesis and promoting
endothelial survival (28). The
expression levels of VEGF were reduced in animal models of BDP and
in the lungs of premature infants, in whom BPD was the cause of
mortality (29–31). Inhibition of VEGF receptor-2
(VEGFR2) has been shown to cause rarefaction of pulmonary vessels
and impairment in alveolar formation (32) in neonatal rats, whereas enhancement
of VEGF signaling ameliorated the alveolar disruption induced by
hyperemia (33). Furthermore,
blocking postnatal angiogenesis impaired alveolarization, and
decreased pulmonary capillary density was observed in animal models
and patients with BPD (34).
The results of the present study are in accordance
with these previous studies. Expression levels of TGF-β1 protein
and mRNA were significantly decreased, and those of VEGF were
significantly increased, at 14 days after intravenous injection of
BMSCs. Furthermore, the results of the immunofluorescence
experiments showed that BMSCs were able to differentiate into cells
expressing vWF and VEGF at day 14. These results suggested that
intravenously implanted BMSCs had the ability to differentiate into
vascular endothelial cells in vivo. Therefore, the
protective mechanisms underlying the effects of treatment with
BMSCs in a mouse model of BPD included homing of these cells to the
lung, where they differentiated into vascular endothelial cells, or
through paracrine effects.
In conclusion, the present study demonstrated that
intravenous injection of BMSCs significantly improved the damaged
airway structure and the levels of TGF-β1 and VEGF in the lungs of
mice with BPD.
Acknowledgements
This study was supported by a grant from the Youth
Foundation of the Second Hospital of Shandong University (grant no.
Y2013010068).
References
|
1
|
Rezvani M, Wilde J, Vitt P, et al:
Association of a FGFR-4 gene polymorphism with bronchopulmonary
dysplasia and neonatal respiratory distress. Dis Markers.
35:633–640. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sakurai R, Li Y, Torday JS and Rehan VK:
Curcumin augments lung maturation, preventing neonatal lung injury
by inhibiting TGF-β signaling. Am J Physiol Lung Cell Mol Physiol.
301:L721–L730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xu J, Woods CR, Mora AL, et al: Prevention
of endotoxin-induced systemic response by bone marrow-derived
mesenchymal stem cells in mice. Am J Physiol Lung Cell Mol Physiol.
293:L131–L141. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ortiz LA, Gambelli F, McBride C, et al:
Mesenchymal stem cell engraftment in lung is enhanced in response
to bleomycin exposure and ameliorates its fibrotic effects. Proc
Natl Acad Sci USA. 100:8407–8411. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
van Haaften T, Byrne R, Bonnet S, et al:
Airway delivery of mesenchymal stem cells prevents arrested
alveolar growth in neonatal lung injury in rats. Am J Respir Crit
Care Med. 180:1131–1142. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aslam M, Baveja R, Liang OD, et al: Bone
marrow stromal cells attenuate lung injury in a murine model of
neonatal chronic lung disease. Am J Respir Crit Care Med.
180:1122–1130. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee JW, Fang X, Krasnodembskaya A, Howard
JP and Matthay MA: Concise review: Mesenchymal stem cells for acute
lung injury: role of paracrine soluble factors. Stem Cells.
29:913–919. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gauldie J, Galt T, Bonniaud P, et al:
Transfer of the active form of transforming growth factor-beta 1
gene to newborn rat lung induces changes consistent with
bronchopulmonary dysplasia. Am J Pathol. 163:2575–2584. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vicencio AG, Lee CG, Cho SJ, et al:
Conditional overexpression of bioactive transforming growth
factor-beta1 in neonatal mouse lung: a new model for
bronchopulmonary dysplasia? Am J Respir Cell Mol Biol. 31:650–656.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Monz D, Tutdibi E, Mildau C, et al: Human
umbilical cord blood mononuclear cells in a double-hit model of
bronchopulmonary dysplasia in neonatal mice. PLoS One.
8:e747402013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Abman SH: Impaired vascular endothelial
growth factor signaling in the pathogenesis of neonatal pulmonary
vascular disease. Adv Exp Med Biol. 661:323–335. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jakkula M, Le Cras TD, Gebb S, et al:
Inhibition of angiogenesis decreases alveolarization in the
developing rat lung. Am J Physiol Lung Cell Mol Physiol.
279:L600–L607. 2000.PubMed/NCBI
|
|
13
|
Acarregui MJ, Penisten ST, Goss KL,
Ramirez K and Snyder JM: Vascular endothelial growth factor gene
expression in human fetal lung in vitro. Am J Respir Cell Mol Biol.
20:14–23. 1999. View Article : Google Scholar
|
|
14
|
Okabe M, Ikawa M, Kominami K, et al:
‘Green mice’ as a source of ubiquitous green cells. FEBS Lett.
407:313–319. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peng C, Yang K, Xiang P, et al: Effect of
transplantation with autologous bone marrow stem cells on acute
myocardial infarction. Int J Cardiol. 162:158–165. 2013. View Article : Google Scholar
|
|
16
|
Balasubramaniam V, Mervis CF, Maxey AM,
Markham NE and Abman SH: Hyperoxia reduces bone marrow, circulating
and lung endothelial progenitor cells in the developing lung:
implications for the pathogenesis of bronchopulmonary dysplasia. Am
J Physiol Lung Cell Mol Physiol. 292:L1073–L1084. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kunig AM, Balasubramaniam V, Markham NE,
Seedorf G, Gien J and Abman SH: Recombinant human VEGF treatment
transiently increases lung edema but enhances lung structure after
neonatal hyperoxia. Am J Physiol Lung Cell Mol Physiol.
291:L1068–L1078. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Luan Y, Zhang ZH, Wei DE, Lu Y and Wang
YB: Effects of autologous bone marrow mononuclear cells
implantation in canine model of pulmonary hypertension. Circ J.
76:977–985. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rojas M, Xu J, Woods CR, et al: Bone
marrow derived mesenchymal stem cells in repair of the injured
lung. Am J Respir Cell Mol Biol. 33:145–152. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ortiz LA, Gambelli F, McBride C, et al:
MSC engraftment in lung is enhanced in response to bleomycin
exposure and ameliorates its fibroproliferative effects. Proc Natl
Acad Sci USA. 100:8407–8011. 2003. View Article : Google Scholar
|
|
21
|
Abman SH and Matthay MA: Mesenchymal stem
cells for the prevention of bronchopulmonary dysplasia: delivering
the secretome. Am J Respir Crit Care Med. 180:1039–1041. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hennrick KT, Keeton AG, Nanua S, et al:
Lung cells from neonates show a mesenchymal stem cell phenotype. Am
J Respir Crit Care Med. 175:1158–1164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang Y, Jahagirdar BN, Reinhardt RL, et
al: Pluripotency of mesenchymal stem cells derived from adult
marrow. Nature. 418:41–49. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Krause DS, Theise ND, Collector MI, et al:
Multi-organ, multi-lineage engraftment by a single bone
marrow-derived stem cell. Cell. 105:369–377. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wen ST, Chen W, Chen HL, et al: Amniotic
fluid stem cells from EGFP transgenic mice attenuate
hyperoxia-induced acute lung injury. PLoS One. 8:e753832013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Collins JJ, Kallapur SG, Knox CL, et al:
Repeated intrauterine exposures to inflammatory stimuli attenuated
transforming growth factor-β signaling in the ovine fetal lung.
Neonatology. 104:49–55. 2013. View Article : Google Scholar
|
|
27
|
Collins JJ, Kunzmann S, Kuypers E, et al:
Antenatal glucocorticoids counteract LPS changes in TGF-β pathway
and caveolin-1 in ovine fetal lung. Am J Physiol Lung Cell Mol
Physiol. 304:L438–L444. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Abman SH: Impaired vascular endothelial
growth factor signaling in the pathogenesis of neonatal pulmonary
vascular disease. Adv Exp Med Biol. 661:323–335. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
De Paepe ME, Mao Q, Powell J, et al:
Growth of pulmonary microvasculature in ventilated preterm infants.
Am J Respir Crit Care Med. 173:204–211. 2006. View Article : Google Scholar
|
|
30
|
Maniscalco WM, Watkins RH, Pryhuber GS,
Bhatt A, Shea C and Huyck H: Angiogenic factors and alveolar
vasculature: development and alterations by injury in very
premature baboons. Am J Physiol Lung Cell Mol Physiol.
282:L811–L823. 2002.PubMed/NCBI
|
|
31
|
Acarregui MJ, Penisten ST, Goss KL,
Ramirez K and Snyder JM: Vascular endothelial growth factor gene
expression in human fetal lung in vitro. Am J Respir Cell Mol Biol.
20:14–23. 1999. View Article : Google Scholar
|
|
32
|
Jakkula M, Le Cras TD, Gebb S, et al:
Inhibition of angiogenesis decreases alveolarization in the
developing rat lung. Am J Physiol Lung Cell Mol Physiol.
279:L600–L607. 2000.PubMed/NCBI
|
|
33
|
Hosford GE and Olson DM: Effects of
hyperoxia on VEGF, its receptors and HIF-2alpha in the newborn rat
lung. Am J Physiol Lung Cell Mol Physiol. 285:L161–L168.
2003.PubMed/NCBI
|
|
34
|
Le Cras TD, Markham NE, Tuder RM, Voelkel
NF and Abman SH: Treatment of newborn rats with a VEGF receptor
inhibitor causes pulmonary hypertension and abnormal lung
structure. Am J Physiol Lung Cell Mol Physiol. 283:L555–L562.
2002.PubMed/NCBI
|