Introduction
As one of the most prevalent causes of human
mortality, stroke accounts for hundreds of thousands of mortalities
annually worldwide (1). Rapid
revascularization of the occluded vessels and timely reperfusion is
one of the most effective approaches to salvage cerebral ischemic
damage; however, the restoration of blood flow during reperfusion
may evoke ischemia/reperfusion (I/R) injury that is not present
during ischemia but can be modulated only at reperfusion (2). Efforts have been made to attenuate
the injury occurring during reperfusion, including numerous types
of pretreatment measurements (2,3).
Electroacupuncture (EA) is a novel therapy based on traditional
acupuncture in combination with modern electrotherapy (4). Evidence has shown that following
vessel occlusion, EA not only reduces myocardial injury but also
significantly promotes the recovery of neurological function and
thus improves quality of life for patients (5,6). In
recent years, several studies have shown that EA pretreatment has
beneficial effects, including reperfusion tolerance (7); however, it remains to be elucidated
whether the underlying mechanisms of EA require the involvement of
autophagy.
Autophagy, a cellular process mediating the
lysosomal degradation of long-lived cytoplasmic proteins, is
initiated under different conditions, including differentiation,
starvation, stress (such as, oxidative or endoplasmic reticulum
stress) and protein aggregate accumulation (8–10).
During autophagy, cytoplasmic components are sequestered into
double-membrane vesicles termed autophagosomes, which then fuse
with lysosomes to produce single-membraned autophagosomes and are
eventually degraded by lysosomal hydrolases (11,12).
There are at least two marked proteins in the autophagic cascade:
Microtubule-associated protein 1A light chain 3 (LC3), which exists
in cytosolic form (LC3-I) and membrane-bound form (LC3-II), the
ratio of conversation from LC3-I to LC3-II is closely correlated
with the extent of autophagosome formation (13); and Beclin 1, which is essential for
recruitment of other autophagic proteins during the expansion of
pre-autophagosomal membrane (14,15).
Previous studies have reported the autophagy was activated
following ischemic injury; however, the contribution of autophagy
to neuronal survival/death remains in debate. Autophagy activation
maintains cellular homeostasis and survival either through purging
the cell of dysfunctional organelles, toxic metabolites and
intracellular pathogens, or generating the intracellular building
blocks required to preserve vital functions during nutrient
deprivation (16,17); for example, the autophagy induced
by focal ischemia preconditioning was reported to be beneficial
(18,19). Conversely, autophagy also triggers
non-apoptotic programmed cell death (autophagic cell death) through
excessive self-digestion and degradation of essential cellular
constituents, which was implicated in various physiological and
pathological conditions associated with neurological diseases
(20); for example, autophagy
induced by permanent cerebral ischemia was reported to be
deleterious (21–23). Overall, physiological levels of
autophagy due to mild hypoxia, appear to be protective. By
contrast, increased levels of autophagy due to severe hypoxia
and/or reperfusion may lead to self-digestion and eventual cell
death (24). The present study
aimed to investigate the hypothesis that EA exerted neuroprotective
effects via inhibition of autophagy during reperfusion, using
middle cerebral artery occlusion (MCAO) model rats.
Materials and methods
The experimental protocol used in the present study
was approved by the Ethics Committee for Animal Experimentation of
Nanjing University of Chinese Medicine (Nanjing, China), and was
conducted according to the Guidelines for Animal Experimentation of
Nanjing University of Chinese Medicine. Male Sprague-Dawley rats,
weighing 280–320 g, were provided by the Experimental Animal Center
of the Nanjing University of Chinese Medicine and were housed under
controlled conditions as follows: 12-h light/dark cycle, 21±2°C and
60–70% humidity, for a minimum of one week prior to drug treatment
or surgery. Rats were provided with a standard rodent diet and tap
water ad libitum. The focal ischemia/reperfusion models were
established through MCAO (25).
During the procedure, room temperature was maintained at 25°C. All
rats were anesthetized with an intraperitoneal injection of sodium
pentobarbital (40 mg/kg; Sigma-Aldrich, St. Louis, MO, USA), and an
incision was made on the midline of the neck. The right common
carotid artery (CCA), the external carotid artery (ECA) and
internal carotid artery (ICA) were isolated, and temporarily
ligated at their origin with a 6.0 silk suture. Microaneurysm clips
were placed around the CCA and CEA to prevent bleeding during
insertion of the suture. A hole was subsequently made in the CCA,
between the clips with a needle (2-ml injector) and the blunted tip
of a nylon suture (diameter, 0.205 mm; length, 20±2 mm) was
inserted through the hole until a mild resistance was felt. The two
remaining loose collar sutures were gently tightened, and the
vessel clips were withdrawn. Rats were anesthetized using an
intraperitoneal injection of 40 mg/kg sodium pentobarbital while
undergoing EA stimulation or surgery.
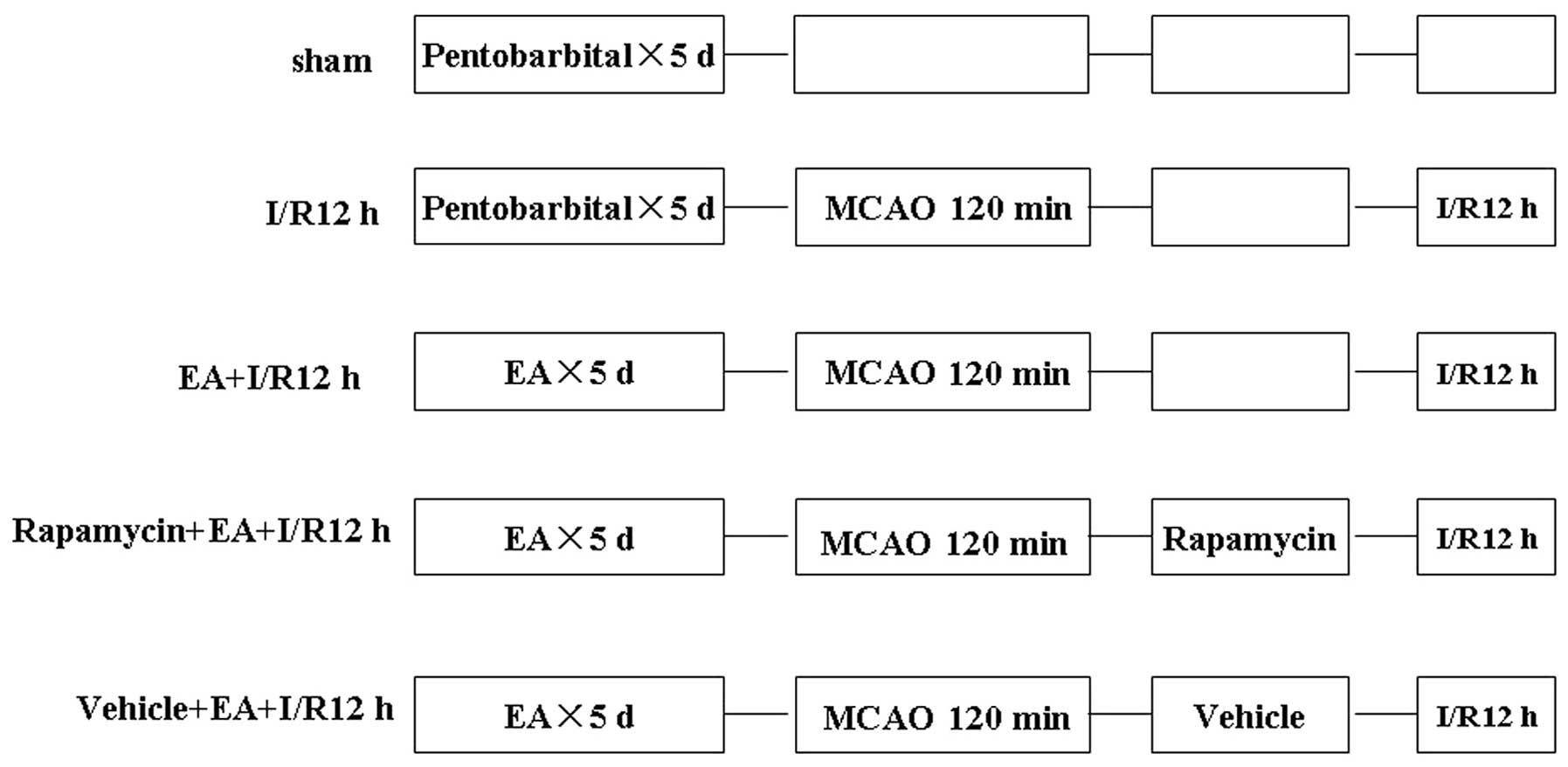
Experiment protocol
In order to determine the time-dependent effects of
I/R on autophagy expression, rats were assigned at random to one of
the following four groups: Sham, 6 h following I/R (I/R 6 h), I/R
12 h, and I/R 24 h. The optimum time-point was determined and rats
were then assigned to one of the following five groups: Sham, I/R
12 h, EA+I/R 12 h, Rapamycin+ EA+I/R 12 h (Sigma-Aldrich) or
Vehicle+EA+I/R 12 h (Fig. 1). The
rats in the sham group underwent an identical surgical procedure
without MCAO. I/R was established through MCAO for 120 min followed
by reperfusion for 12 h; EA pretreatment was achieved through
stimulation of the acupoint ‘Baihui (GV20)’ on five consecutive
days prior to I/R. Levels of autophagy were determined using
transmission electron microscopy and western blot analysis. The
neuroprotective effects of EA were determined using a terminal
deoxynucleotidyl transferase-mediated deoxyuridine-triphosphatase
nick end labeling (TUNEL) assay, infarct volumes, brain water
content and motor deficit. The numbers of rats used for each
experiment are summarized in Table
I.
 | Table IExperimental protocol. |
Table I
Experimental protocol.
| Protocol | Experiment | Time-point | Group | Number | Total |
|---|
| Effect of EA on
autophagy expression | Transmission
electron microscopic | I/R 12 h | sham, I/R 12 h, EA
+I/R 12 h, Rapamycin+EA+ I/R 12 h, Vehicle +EA+I/R 12 h | n=5 | 25 |
| Western
blotting | I/R 12 h | sham, I/R 12 h, EA
+I/R 12 h, Rapamycin+EA +I/R 12 h, Vehicle+EA +I/R 12 h | n=5 | 25 |
| Neuroprotective
effect of EA | TUNEL assay | I/R 12 h | sham, I/R 12 h, EA
+I/R 12 h, Rapamycin+EA+ I/R 12 h, Vehicle +EA +I/R 12 h | n=5 | 25 |
| TTC staining | I/R 12 h | I/R 12 h, EA +I/R
12 h, Rapamycin+EA+I/R 12 h | n=8 | 24 |
| Sum | | | | | 99 |
EA pretreatment
EA pretreatment was performed at the acupoint
‘Baihui (GV20)’ (5). The acupoint
‘Baihui’ is located at the intersection of the sagittal midline and
the line between the two ears (26). Animals were anesthetized and
stimulated at an intensity of 1 mA and a frequency of 2/15 Hz for
30 min/day for five consecutive days prior to MCAO, using the Hwato
Electronic Acupuncture Treatment Instrument (SDZ-V; Suzhou Medical
Appliances Co., Ltd., Jiangsu, China).
Transient focal cerebral ischemia
Rats were subjected to transient MCAO for 120 min
following the final pretreatment as described above. Reperfusion
was achieved by withdrawing the suture following 120 min ischemia.
Cerebral blood flow (CBF) through the middle cerebral artery was
measured using laser Doppler flowmetry (PeriFlux 5000; Perimed,
Järfälla, Sweden). A flexible fiber-optic probe (PeriFlux 5000;
Perimed) was affixed to the skull over the cortex supplied by the
proximal part of the middle cerebral artery (2 mm caudal to bregma
and 6 mm lateral to middle). Animals with <80% reduction in CBF
in the core of the middle cerebral artery area were excluded from
this study.
Treatment with the autophagy inducer
In order to determine whether rapamycin, an
autophagy inducer, attenuated the neuroprotective effects of EA at
the acupoint ‘Baihui (GV20)’, rats were treated with intracerebral
ventricle injection (i.c.v.) of 35 pmol rapamycin (Sigma, St.
Louis, MO, USA) at the onset of reperfusion; the induction of
autophagy using 35 pmol rapamycin was significant according to
previous studies (19). Rapamycin
was dissolved in ethanol and then diluted to the final
concentration using normal saline solution (final ethanol
concentration, <2%). Control animals received the same volume
injection of vehicle.
Transmission electron microscopic
examination
Following reperfusion for 12 h, rats were
anesthetized, as described above, and then perfused with pre-cooled
phosphate-buffered saline (PBS; pH 7.4) followed by PBS containing
4% paraformaldehyde and 0.25% glutaraldehyde. The brains were
removed and kept overnight in a solution of 2% paraformaldehyde and
2.5% glutaraldehyde in 0.1 M PBS (pH 7.4). The following day, the
rat brains were sectioned using a vibratome (myNeuroLab; Leica
Microsystems, Ltd, Milton Keynes, UK) into 0.5-mm slices. The
parietal lobe cortex in the ischemic core area was selected for
analysis, and selected areas were processed by post-fixation in 1%
osmium tetroxide for 1 h, dehydrated in graded ethanol and then
embedded in epoxy resin. Polymerization was performed at 80°C for
24 h. Blocks were sectioned using an PowerTome-PC ultramicrotome
(RMC, Inc., Tucson, AZ, USA) into ultrathin sections (60–70 nm),
which were post-stained with uranyl acetate and lead citrate, and
visualized using a Hitachi 7100 electron microscopy (Nikon, Tokyo,
Japan). For quantitative analysis of the number of autophagosomes,
ten fields of vision for each rat (three rats in each group) were
examined as previously described (27).
TUNEL assay
A TUNEL assay (Boster Biological Technology, Ltd,
Wuhan, China) was performed according to the Manufacturer’s
instructions. Brain sections were stained with diaminobenzidine,
counterstained with hematoxylin, dehydrated with gradient alcohol,
cleared with xylene and then mounted with neutral resin. The
reaction mixture was replaced with PBS in the negative control.
Normal nuclei were stained blue and apoptotic nuclei were stained
brownish-black or brown. Four sections were observed from each rat
(magnification, x400) and ten fields of vision (n=5) on each slide
were counted. The percentage of positive apoptotic nuclei in the
total number of cells per field was calculated and the mean value
was taken to be the apoptotic index of cerebral neurons.
Western blotting
Following reperfusion for 12 h, the ischemic
parietal lobe cortex supplied by the right MCAO and the
corresponding area of sham-operated rats were homogenized and
proteins were extracted using a lysis buffer (10 mM Tris-Hcl, pH
7.4; 150 mM NaCl; 1% Triton X-100; 0.1% SDS; 5 mM EDTA; 1 mM PMSF;
0.28 U/ml aprotinin; 50 μg/ml leupeptin; 1 mM benzamidine;
and 7 μg/ml pepstatin A) as previously described (28). Protein concentrations were
determined using a spectrophotometer (UV-2540; SHIMADZH Corp.,
Kyoto, Japan). A 60 μg aliquot of protein from each sample
was separated using 10% SDS-PAGE and subsequently transferred to a
nitrocellulose membrane (Shanghai Haoran Bio-Technology Co., Ltd,
Shanghai, China). The membranes were then incubated with specific
polyclonal antibodies against LC3 (#4108; 1:1,000; Cell Signaling
Technology, Inc., Danvers, MA, USA), or Beclin 1 (#3738; 1:500;
Cell Signaling Technology, Inc.) at 4°C over night, and then
incubated with a horseradish peroxidase-conjugated goat anti-rabbit
secondary antibody (sc2004; 1:5,000; Jackson ImmunoResearch
Laboratories, Inc., West Grove, PA, USA) at room temperature for 1
h. Immunoreactivity was detected using enhanced chemiluminescent
autoradiography (Bioroc Pharmaceutical & Biotech Co., Ltd,
Tianjin, China) in accordance with the manufacturer’s instructions.
The membranes were reprobed with β-actin (1:5,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) following stripping. Films
were then used for final determination of protein expression using
Sigma scan software (scanPDF15; Sigma-Aldrich) and normalized to
the loading control.
Evaluation of infarct volume, brain water
content and motor deficits
Following reperfusion for 12 h, the motor deficits
in rats were evaluated in a blinded manner using a previously
described protocol (29) and were
scored as follows: 0, Rats behave normally; 1, rats cannot fully
stretch their left front leg; 2, rats turn around in a circle; 3,
rats fall down to the left side; 4, rats cannot move by themselves
and lose consciousness. Following scoring, rats with >2 points
were sacrificed by anesthetization with sodium pentobarbital
followed by decapitation, and the brains were dissected and sliced
in a plastic module (3-mm) and then stained with 2%
2,3,5-triphenyltetra-zolium chloride (TTC) for 30 min. The total
wet weight of the TTC-stained brains was quantified using an
electronic scale. The wet red and white brain regions of the
TTC-stained brains were collected separately. Infarct volume was
analyzed using five slices of 3-mm coronal sections from each brain
and calculated with the following formula: Infarct volume (%) =
[(total wet weight - red weight)/total wet weight] × 100. The red
and white sections of these brains were then desiccated at 105°C
for 48 h until the weight was constant. The total weight of the
dried TTC-stained brains was follows: Water content (%) = [(wet
weight-dried weight)/wet weight] × 100 (30).
Statistical analysis
Statistical analysis was performed using a one-way
analysis of variance followed by a Student-Newman-Keul’s post hoc
analysis. Statistical analyses were performed using SPSS 15.0
(SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference between values.
Results
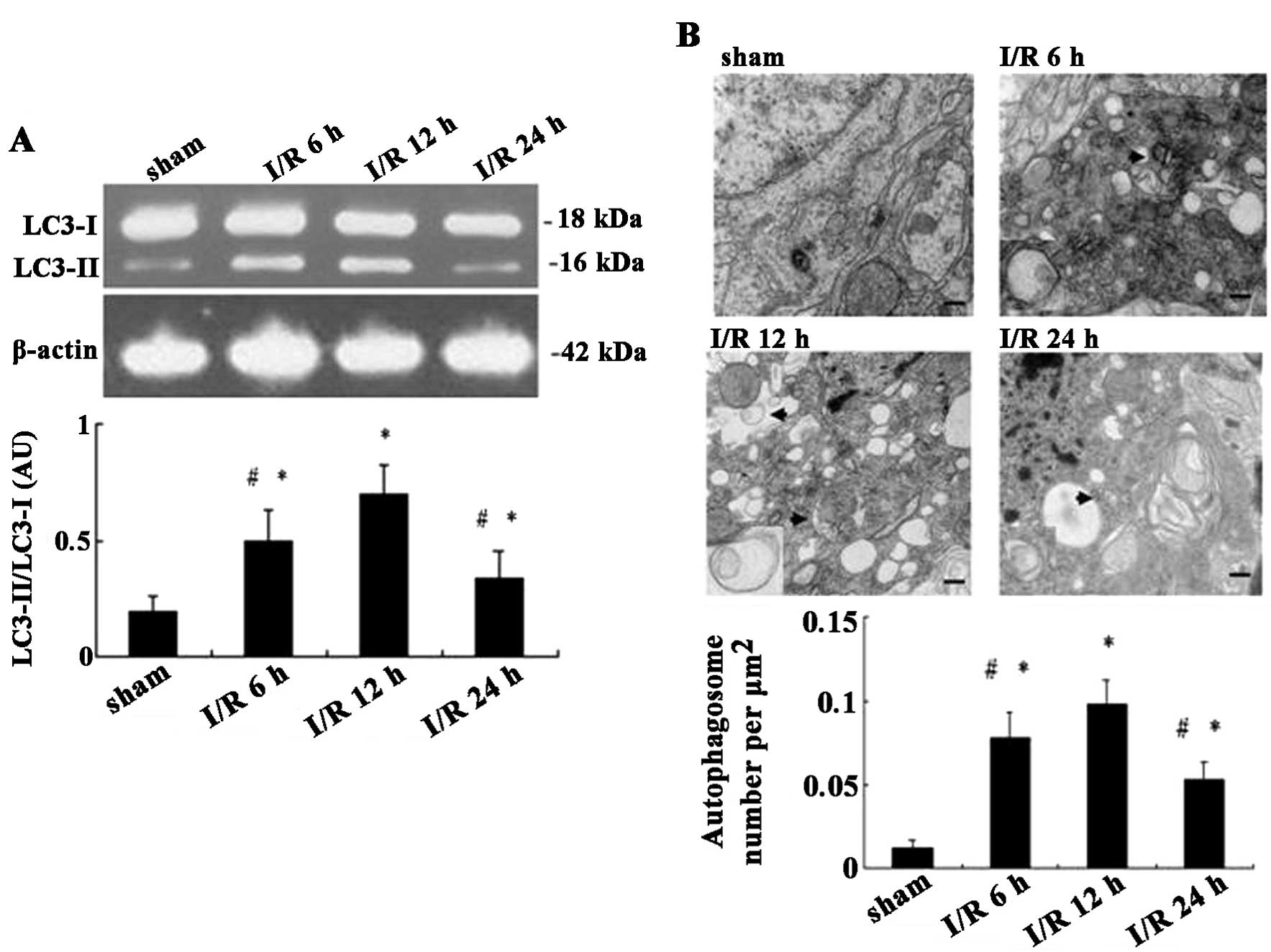
Autophagy is upregulated following
I/R
In order to determine the time-dependent effects of
I/R on autophagy, the expression levels of the activated autophagy
biomarker LC3 were detected using western blot analysis and the
number of autophagosomes present were observed using transmission
electron microscopic at different time points following I/R. In
addition, the ratio of LC3-II/LC3-I was determined, which reflected
the extent of autophagosome formation. As shown in Fig. 2A, the ratio of LC3-II/LC3-I in the
ischemic cortex of adult rats for each group were 0.19, 0.5, 0.7
and 0.34, respectively. The ratio at each time-point (6–24 h)
following I/R was significantly increased compared to that of the
sham group (F=18.63, P=0.000), with a maximal value at 12 h
following I/R. Similarly, the number of autophagosomes
significantly increased at each time-point following I/R (F=48.44,
P=0.000), and peaked at 12 h (Fig.
2B). Therefore 12 h post-I/R was used for all subsequent
experiments to further explore the potential autophagic mechanism
of EA pretreatment.
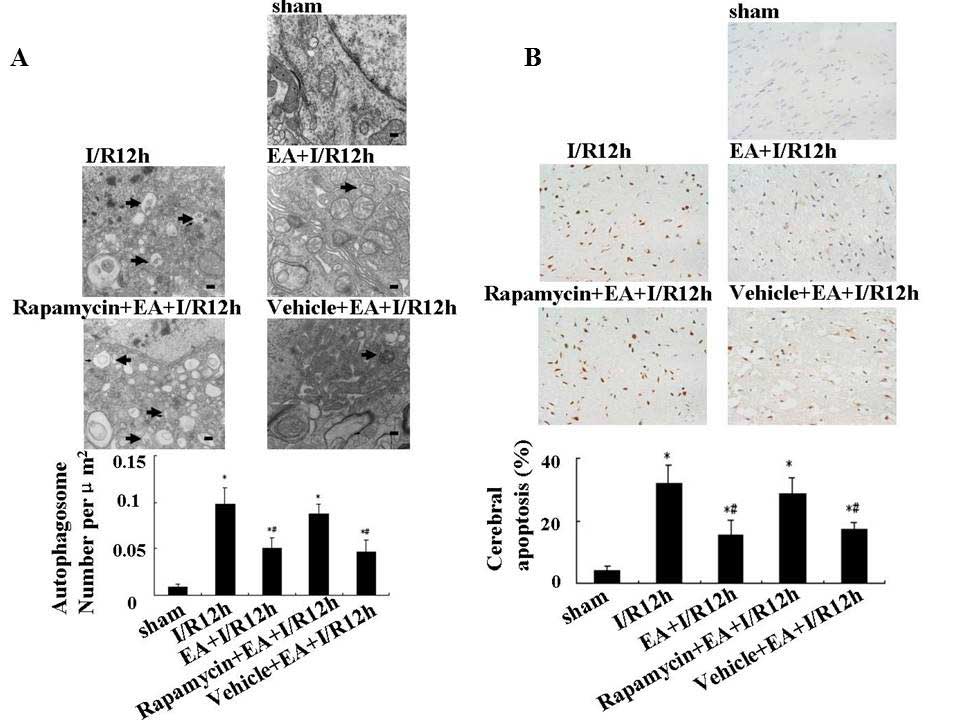
EA pretreatment decreases the
autophagosome and cerebral apoptosis 12 h after I/R
In order to examine the direct effect of EA
pretreatment on autophagy, transmission electron microscopy was
used to observe the morphological changes of neurons in each
treatment group. Neurons in the cortex of the sham group appeared
to be normal with comparatively healthy endoplasmic reticulum,
mitochondria, lysosomes and nuclei (Fig. 3A). By contrast, neurons in the I/R
12 h group displayed an apparent disease morphology; the neurons
showed fragmented endoplasmic reticulum, formation of numerous
vacuoles in the cytoplasm and condensation of chromatin. Numerous
autophagosomes (C-shaped double-membrane structures) and lysosomes
with engulfed cytoplasmic materials were observed. Neurons in the
EA+I/R 12 h group displayed mild injury, with a number of normal
organelles and nuclei observed; however, autophagosomes remained
present. Quantitative analysis of the electron microscopy images
showed that the number of autophagosomes in EA+I/R 12 h group was
significantly decreased compared with that of the I/R 12 h group
(0.098 and 0.05 autophagosomes per μm2,
respectively; P<0.05).
In order to determine the contribution of autophagic
mechanisms to the beneficial effect elicited through EA
pretreatment, a single i.c.v injection of the autophagy inducer
rapamycin (35 pmol) was administered at the onset of reperfusion
and its effects were examined. The results showed that neurons in
the rapamycin+EA+I/R 12 h group exhibited injury comparable to that
of the I/R 12 h group, suggesting that rapamycin suppressed the
beneficial effect of EA pretreatment.
Furthermore, cerebral apoptosis was detected using a
TUNEL assay. As shown in Fig. 3B,
cerebral apoptosis was significantly increased in rats subjected to
I/R 12 h compared to that of the sham group, whereas EA
pretreatment significantly inhibited I/R-induced apoptosis (32.1
and 15.5%, respectively; P<0.05). However, following
administration with 35 pmol rapamycin prior to the onset of
reperfusion, levels of cerebral apoptosis were comparable to that
of the I/R 12 h group, therefore attenuating the effects of EA
pretreatment.
These results therefore supported the hypothesis
that EA pretreatment exerted a protective effect during I/R through
suppression of neuronal autophagy.
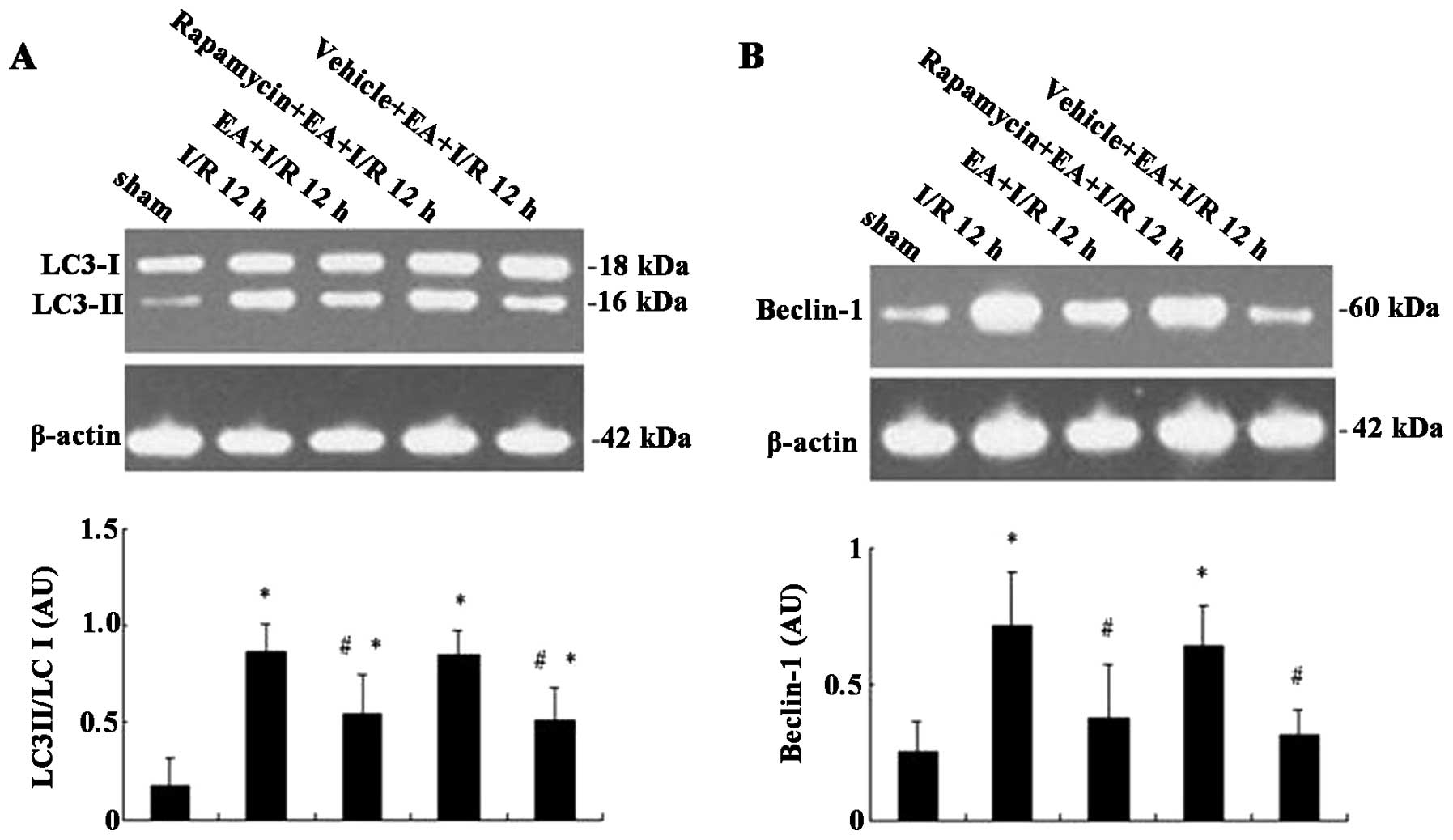
EA pretreatment reduces the expression of
I/R-induced autophagic marker proteins 12 h following IR
In order to further investigate whether EA
pretreatment affects autophagy, western blot analysis was used to
detect the expression levels of autophagic marker proteins, LC3-I,
LC3-II and Beclin-1 in the ischemic cortex of rats. As shown in
Fig. 4A, the expression levels of
LC3-II were low in sham group and high in the I/R 12 h group. EA
pretreatment markedly decreased the expression of LC3-II; however,
rapamycin attenuated the inhibitory effect of EA on LC3-II. In
addition, LC3-I expression only fluctuated marginally among the
treatment groups, therefore the ratio of LC3-II/LC3-I was
quantified. EA pretreatment significantly decreased the ratio of
LC3-II/LC3-I compared with that of the I/R 12 h group (0.55 and
0.87, respectively; P<0.05); however, rapamycin inhibited the
decreased ratio of LC3-II/LC3-I in the EA+I/R 12 h group (0.85 and
0.55, respectively), with comparable results to that of the I/R 12
h group. As shown in Fig. 4B, the
effect of the treatment groups on Beclin-1 expression was
comparable to that on LC3-II expression. Beclin-1 expression was
significantly upregulated following I/R and suppressed by EA
pretreatment; however, rapamycin attenuated the effects of EA
pretreatment on Beclin-1 expression (F=8.86, P=0.000 vs. EA+I/R 12
h group). These results therefore suggested that EA pretreatment
prevented I/R-induced autophagy via downregu-lation of LC3-II and
Beclin-1 expression.
Autophagy inducer rapamycin abolishes the
neuroprotective effects of EA pretreatment
In order to explore the contribution of autophagic
mechanisms to the neuroprotection elicited by EA pretreatment, the
effects of rapamycin on infarct volume, water content and motor
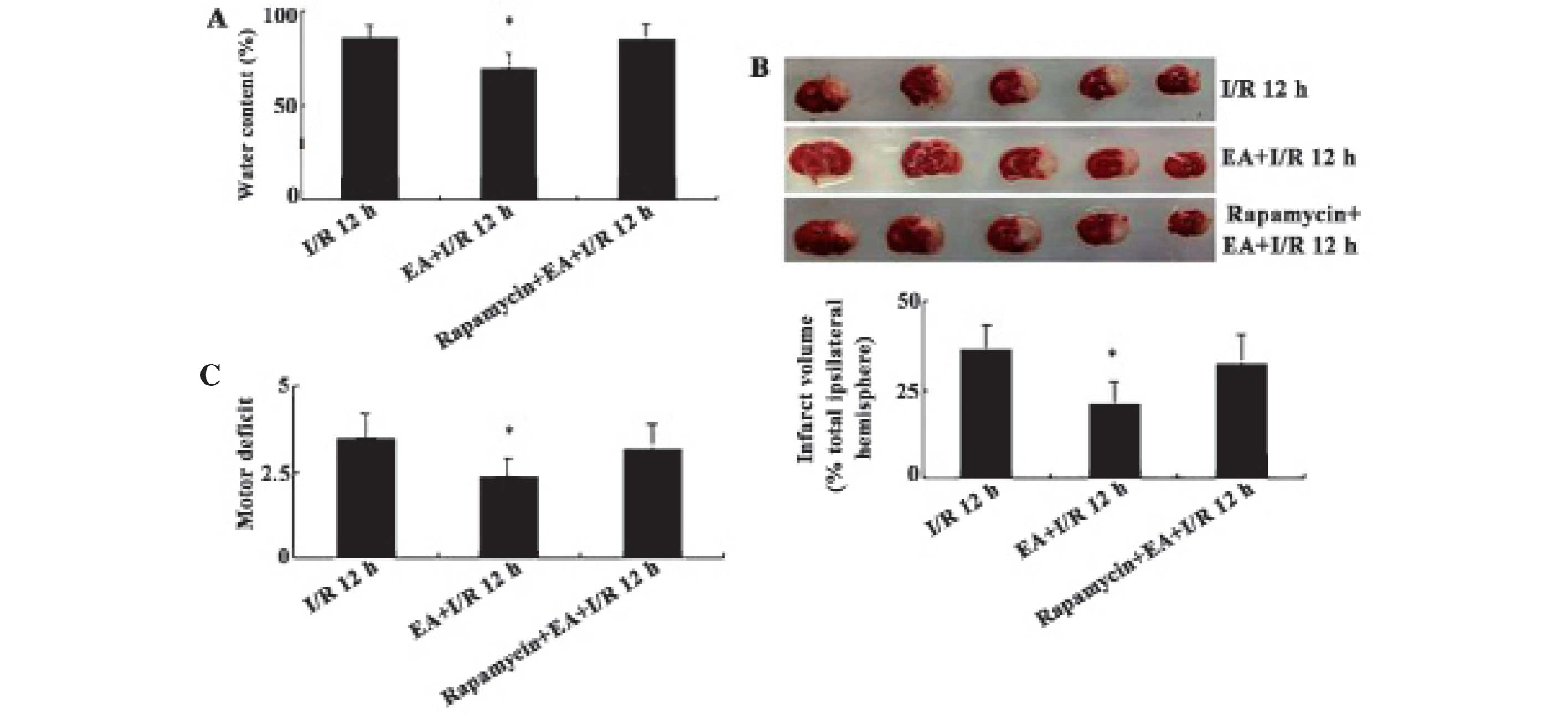
deficits induced by EA pretreatment were examined (Fig. 5). In rats subjected to I/R,
extensive infarction was detected in the ipsilateral cerebral
cortical and subcortical areas over a series of brain sections. EA
pretreatment significantly reduced the infarct volume and water
content of rats in the I/R 12 h group, whereas administration of 35
pmol rapamycin abolished the protective effects of EA pretreatment,
as evidenced by the increased infarct volume and water content
(F=10.24/P= 0.001 and F=11.6/P=0.000, respectively) (Fig. 5A and B). Furthermore, the effects
of EA pretreatment on I/R-induced motor behavior deficits were
evaluated (Fig. 5C); motor
deficits were scored using a five-point scale as described in the
Materials and methods. EA pretreatment (EA+I/R 12 h) resulted in a
significant reduction in motor deficits induced by I/R (F=6.253,
P=0.007 vs. I/R 12 h group), while administration with rapamycin
significantly attenuated the beneficial effects of EA pretreatment
on neurological motor deficits. These results supported the
hypothesis that EA pretreatment protected neurons against I/R
through an autophagic mechanism.
Discussion
EA is a novel combination of traditional
acupuncture-based therapy and modern electrotherapy. EA is
conducted by inserting acupuncture needles into acupoints and then
altering electric stimulation parameters, including stimulation
frequency, current intensity, pulse width and pulse interval
(31). Therefore, the choice of
acupoint and stimulation parameters is important; based on meridian
theory, the ‘Baihui’ acupoint (one of the acupoints of Du merdian)
was selected for use in the present study as it receives
projections from the motor and sensory cortex, as well as from the
anterior cerebral artery. In addition, according to the results of
a previous study, ‘Baihui acupoint (GV20)’-stimulation with a
density-sparse wave of 2/15 Hz and an intensity of 1 mA for 30
min/day over five consecutive days was reported to induce cerebral
I/R tolerance (32). Evidence has
suggested that I/R disturbs energy metabolism, increases oxidative
stress and triggers apoptotic cell death (33). Previous studies have reported that
EA pretreatment exerted neuroprotective effects against I/R injury
via regulation of oxidative stress, maintenance of blood-brain
barrier integrity and inhibition of apoptosis via various receptors
(34,35). In addition, I/R was reported to
induce autophagic cell death (36,37);
therefore, it is possible that EA pretreatment protected neurons
against I/R injury through regulating the levels of autophagy. The
present study provided evidence to suggest that the neuroprotective
effect of EA pretreatment was, at least in part, associated with
the autophagy pathway.
Autophagy is an evolutionarily conserved pathway,
which involves the isolation and transport of cytoplasmic materials
to lysosomes where proteins and organelles are degraded and
recycled. Under physiological conditions, autophagy activation has
been associated with neuroprotection via degradation of misfolded
protein; for example, preconditioning ischemia protects neurons
against subsequent brain damage via upregulated autophagy (38). However, under certain pathological
conditions, for example reperfusion following prolonged ischemia,
autophagy overactivation promotes cell death through
apoptotic-dependent and -independent cascades. A previous study
indicated that excess autophagy activation was involved in several
models of cerebral ischemia, including global cerebral ischemia,
focal ischemia and hypoxia-ischemia, through inducing autophagic
cell death (39). Rami et
al (23) demonstrated that
markedly elevated levels of autophagy contributed to subsequent
neuronal death in the penumbra neurons of adults rats following
focal cerebral I/R. In addition, Liu et al (40) reported that upregulation of LC3-II
was associated with brain injury in post-ischemic brain tissue
following global cerebral I/R. Furthermore, Xu et al
(41) supported the association
between significantly increased levels of the autophagic marker
proteins, LC3-II and Beclin-1, and neuronal death in the ischemic
cortex of mice following focal cerebral I/R. Consistent with these
studies, the results of the present study demonstrated that the
ratio of LC3-II/LC3-I and Beclin-1 expression in the ischemic
cortex of adult rats increased significantly following focal
cerebral ischemia, which may be associated with neuronal damage.
These results therefore indicated that autophagy levels may be
associated with neuronal fate.
The results of the present study demonstrated that
EA pretreatment exerted significant protective roles in a rat model
of focal cerebral ischemic model (MCAO), including the reduction of
cerebral apoptosis, infarct volumes and brain edema, which in turn
improved motor deficits. These results were concurrent a previous
report (7). Notably, the
protective effects of EA pretreatment were associated with
inhibition of the autophagy pathway, as evidenced by a reduced
number of autophagosomes in the ischemic parietal lobe and the
attenuated expression of the autophagy marker proteins LC3-II and
Beclin-1 at 12 h following I/R. These results demonstrated that the
neuroprotective effects of EA pretreatment may be, in part,
associated with the inhibition of autophagy. Furthermore, the
effects of the autophagy inducer rapamycin (42) on the neuroprotective effects of EA
pretreatment were investigated. The results showed that the
neuroprotective effects induced by EA pretreatment were partially
reversed by rapamycin, which therefore suggested that the
inhibition of autophagic pathways may partially underlie the
mechanisms by which EA pretreatment induced tolerance to cerebral
I/R injury.
The role of autophagy in neuronal death during
reperfusion remains to be fully elucidated. Accumulating evidence
has suggested that autophagy activation contributed to ischemic
neuronal injury following cerebral I/R (25,43);
however, one study supported the opposite conclusion, that
autophagy activation was associated with protective effects against
neuronal injury (44). For
example, Zhang et al (44)
evaluated the expression of the autophagic marker LC3-II in the
penumbral region 0, 1, 12, and 24 h post-reperfusion and found that
levels of LC3-II/GAPDH increased with reperfusion time, and
persisted until 24 h; however, the inhibition of autophagy during
reperfusion enhanced I/R injury (44). The inconsistency may be due to
different ischemic models, durations of the ischemic insults and
ages of animals. It was reported that autophagy was more pronounced
in adult mice than neonatal mice subjected to hypoxia-ischemia
injury, indicating that autophagic cell death may be more
significant in mature animals (45). Furthermore, the different durations
of the ischemic insults may affect the role of autophagy during
reperfusion.
In conclusion, the results of the present study
suggested that the neuroprotection induced by EA pretreatment
against I/R injury was mediated, at least in part, through
inhibition of the autophagy pathway. Therefore, EA may provide a
promising novel strategy for the clinical treatment of I/R
injury.
Acknowledgments
The present study was supported by Jiangsu Province
Hospital of Traditional Chinese Medicine, The Affiliated Hospital
of Nanjing University of Chinese Medicine (project nos. Y12016,
CJ20140038 and 81202802).
References
|
1
|
Donnan GA, Fisher M, Macleod M and Davis
SM: Stroke. Lancet. 371:1612–1623. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhao H, Sapolsky RM and Steinberg GK:
Interrupting reperfusion as a stroke therapy: ischemic
postconditioning reduces infarct size after focal ischemia in rats.
J Cereb Blood Flow Metab. 26:1114–1121. 2006.PubMed/NCBI
|
|
3
|
Wang JK, Yu LN, Zhang FJ, Yang MJ, Yu J,
Yan M and Chen G: Postconditioning with sevoflurane protects
against focal cerebral ischemia and reperfusion injury via PI3
K/Akt pathway. Brain Res. 1357:142–151. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chou P, Chu H and Lin JG: Effects of
electroacupuncture treatment on impaired cognition and quality of
life in Taiwanese stroke patients. J Altern Complement Med.
15:1067–1073. 2009.
|
|
5
|
Wang Q, Xiong L, Chen S, Liu Y and Zhu X:
Rapid tolerance to focal cerebral ischemia in rats is induced by
preconditioning with electroacupuncture: window of protection and
the role of adenosine. Neurosci Lett. 381:158–162. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Q, Peng Y, Chen S, Gou X, Hu B, Du J,
Lu Y and Xiong L: Pretreatment with electroacupuncture induces
rapid tolerance to focal cerebral ischemia through regulation of
endocannabinoid system. Stroke. 40:2157–2164. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Q, Wang F, Li X, Yang Q, Xu N, Huang
Y, Zhang Q, Gou X, Chen S and Xiong L: Electroacupuncture
pretreatment attenuates cerebral ischemic injury through alpha7
nicotinic acetylcholine receptor-mediated inhibition of
high-mobility group box 1 release in rats. J Neuroinflammation.
9:242012. View Article : Google Scholar
|
|
8
|
Mizushima N: Autophagy: process and
function. Genes Dev. 21:2861–2873. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yorimitsu T, Nair U, Yang Z and Klionsky
DJ: Endoplasmic reticulum stress triggers autophagy. J Biol Chem.
281:30299–30304. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sarkar S, Perlstein EO, Imarisio S, Pineau
S, Cordenier A, Maglathlin RL, Webster JA, Lewis TA, et al: Small
molecules enhance autophagy and reduce toxicity in Huntington’s
disease models. Nat Chem Biol. 3:331–338. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shintani T and Klionsky DJ: Autophagy in
health and disease: a double-edged sword. Science. 306:990–995.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mizushima N: The pleiotropic role of
autophagy: from protein metabolism to bactericide. Cell Death
Differ 12 Suppl. 2:1535–1541. 2005. View Article : Google Scholar
|
|
13
|
Kabeya Y, Mizushima N, Ueno T, Yamamoto A,
Kirisako T, Noda T, Kominami E, Ohsumi Y and Yoshimori T: LC3, a
mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liang XH, Jackson S, Seaman M, Brown K,
Kempkes B, Hibshoosh H and Levine B: Induction of autophagy and
inhibition of tumorigenesis by beclin 1. Nature. 402:672–676. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pattingre S, Tassa A, Qu X, Garuti R,
Liang XH, Mizushima N, Packer M, Schneider MD and Levine B: Bcl-2
antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell.
122:927–939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qin ZH, Wang Y, Kegel KB, Kazantsev A,
Apostol BL, Thompson LM, Yoder J, Aronin N and Di Figlia M:
Autophagy regulates the processing of amino terminal huntingtin
fragments. Hum Mol Genet. 12:3231–3244. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fujita E, Kouroku Y, Isoai A, Kumagai H,
Misutani A, Matsuda C, Hayashi YK and Momoi T: Two endoplasmic
reticulum-associated degradation (ERAD) systems for the novel
variant of the mutant dysferlin: ubiquitin/proteasome ERAD(I) and
autophagy/lysosome ERAD(II). Hum Mol Genet. 16:618–629. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park HK, Chu K, Jung KH, Lee ST, Bahn JJ,
Kim M, Lee SK and Roh JK: Autophagy is involved in the ischemic
preconditioning. Neurosci Lett. 451:16–19. 2009. View Article : Google Scholar
|
|
19
|
Sheng R, Zhang LS, Han R, Liu XQ, Gao B
and Qin ZH: Autophagy activation is associated with neuroprotection
in a rat model of focal cerebral ischemic preconditioning.
Autophagy. 6:482–494. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lockshin RA and Zakeri Z: Apoptosis,
autophagy, and more. Int J Biochem Cell Biol. 36:2405–2419. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wen YD, Sheng R, Zhang LS, Han R, Zhang X,
Zhang XD, Han F, Fukunaga K and Qin ZH: Neuronal injury in rat
model of permanent focal cerebral ischemia is associated with
activation of autophagic and lysosomal pathways. Autophagy.
4:762–769. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Koike M, Shibata M, Tadakoshi M, Gotoh K,
Komatsu M, Waguri S, Kawahara N, Kuida K, Nagata S, et al:
Inhibition of autophagy prevents hippocampal pyramidal neuron death
after hypoxic-ischemic injury. Am J Pathol. 172:454–469. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rami A, Langhagen A and Steiger S: Focal
cerebral ischemia induces upregulation of Beclin 1 and
autophagy-like cell death. Neurobiol Dis. 29:132–141. 2008.
View Article : Google Scholar
|
|
24
|
Sadoshima J: The role of autophagy during
ischemia/reperfusion. Autophagy. 4:402–403. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zheng YQ, Liu JX, Li XZ, Xu L and Xu YG:
RNA interference-mediated downregulation of Beclin1 attenuates
cerebral ischemic injury in rats. Acta Pharmacol Sin. 30:919–927.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hou X, Zhang R, Lv H, Cai X, Xie G and
Song X: Acupuncture at Baihui and Dazhui reduces brain cell
apoptosis in heroin readdicts. Neural Regen Res. 9:164–70. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hirai K, Aliev G, Nunomura A, Fujioka H,
Russell RL, Atwood CS, Johnson AB, Kress Y, Vinters HV, et al:
Mitochondrial abnormalities in Alzheimer’s disease. J Neurosci.
21:3017–3023. 2001.PubMed/NCBI
|
|
28
|
Qin ZH, Chen RW, Wang Y, Nakai M, Chuang
DM and Chase TN: Nuclear factor κB nuclear translocation
upregulates c-Myc and p53 expression during NMDA receptor-mediated
apoptosis in rat striatum. J Neurosci. 19:4023–4033.
1999.PubMed/NCBI
|
|
29
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rosen GD and Harry JD: Brain volume
estimation from serial section measurements: a comparison of
methodologies. J Neurosci Methods. 35:115–124. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Napadow V, Makris N, Liu J, Kettner NW,
Kwong KK and Hui KK: Effects of electroacupuncture versus manual
acupuncture on the human brain as measured by fMRI. Hum Brain Mapp.
24:193–205. 2005. View Article : Google Scholar
|
|
32
|
Xiong LZ, Yang J, Wang Q and Lu ZH:
Involvement of delta- and mu-opioid receptors in the delayed
cerebral ischemic tolerance induced by repeated electroacupuncture
preconditioning in rats. Chin Med J (Engl). 120:394–399. 2007.
|
|
33
|
Talha S, Bouitbir J, Charles AL, Zoll J,
Goette-Di Marco P, Meziani F, Piquard F and Geny B: Pretreatment
with brain natriuretic peptide reduces skeletal muscle
mitochondrial dysfunction and oxidative stress after
ischemia-reperfusion. J Appl Physiol (1985). 114:172–179. 2013.
View Article : Google Scholar
|
|
34
|
Wu XD, Du LN, Wu GC and Cao XD: Effects of
electroacupuncture on blood-brain barrier after cerebral
ischemia-reperfusion in rat. Acupunct Electrother Res. 26:1–9.
2001.PubMed/NCBI
|
|
35
|
Zhao J, Xu H, Tian Y, Hu M and Xiao H:
Effect of electroacu-puncture on brain-derived neurotrophic factor
mRNA expression in mouse hippocampus following cerebral
ischemia-reperfusion injury. J Tradit Chin Med. 33:253–257. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cui D, Wang L, Qi A, Zhou Q, Zhang X and
Jiang W: Propofol prevents autophagic cell death following oxygen
and glucose deprivation in PC12 cells and cerebral
ischemia-reperfusion injury in rats. PLoS One. 7:e353242012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu L, Fang YQ, Xue ZF, He YP, Fang RM and
Li L: β-asarone attenuates ischemia-reperfusion-induced autophagy
in rat brains via modulating JNK, p-JNK, Bcl-2 and Beclin 1. Eur J
Pharmacol. 680:34–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xia DY, Li W, Qian HR, Yao S, Liu JG and
Qi XK: Ischemia preconditioning is neuroprotective in a rat
cerebral ischemic injury model through autophagy activation and
apoptosis inhibition. Braz J Med Biol Res. 46:580–588. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang JY, Xia Q, Chu KT, Pan J, Sun LN,
Zeng B, Zhu YJ, Wang Q, Wang K and Luo BY: Severe global cerebral
ischemia-induced programmed necrosis of hippocampal CA1 neurons in
rat is prevented by 3-methyladenine: a widely used inhibitor of
autophagy. J Neuropathol Exp Neurol. 70:314–22. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu C, Gao Y, Barrett J and Hu B:
Autophagy and protein aggregation after brain ischemia. J
Neurochem. 115:68–78. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu F, Li J, Ni W, Shen YW and Zhang XP:
Peroxisome proliferator-activated receptor-gamma agonist
15d-prostaglandin J2 mediates neuronal autophagy after cerebral
ischemia-reperfusion injury. PLoS One. 8:e550802013. View Article : Google Scholar
|
|
42
|
Xie R, Li X, Ling Y, Shen C, Wu X, Xu W
and Gao X: α-lipoic acid pre- and post-treatments provide
protection against in vitro ischemia-reperfusion injury in cerebral
endothelial cells via Akt/mTOR signaling. Brain Res. 1482:81–90.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Puyal J, Vaslin A, Mottier V and Clarke
PG: Postischemic treatment of neonatal cerebral ischemia should
target autophagy. Ann Neurol. 66:378–389. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang X, Yan H, Yuan Y, Gao J, Shen Z,
Cheng Y, Shen Y, Wang RR, Wang X, et al: Cerebral
ischemia-reperfusion-induced autophagy protects against neuronal
injury by mitochondrial clearance. Autophagy. 9:1321–1333. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhu C, Wang X, Xu F, Bahr BA, Shibata M,
Uchiyama Y, Hagberg H and Blomgren K: The influence of age on
apoptotic and other mechanisms of cell death after cerebral
hypoxia-ischemia. Cell Death Differ. 12:162–176. 2005. View Article : Google Scholar
|