Introduction
Cisplatin is a chemotherapeutic agent, which is
commonly used in the treatment of cancer (1,2).
However, cisplatin has been shown to induce a number of severe side
effects, including peripheral neuropathy and nephrotoxicity. One of
the most serious adverse effects of cisplatin treatment is
ototoxicity, which is manifested as irreversible sensorineural
hearing loss (3,4). Ototoxicity occurs due to
cisplatin-induced auditory and vestibular sensory cell apoptosis
(5).
Accumulating evidence has indicated that
pro-inflammatory cytokines may be involved in cochlear damage
(6–10). These cytokines were shown to induce
secondary inflammatory responses, including leukocyte infiltration
and scar formation. In addition, one study demonstrated that
cisplatin induced ototoxicity via the apoptosis of outer hair cells
(OHCs) and supporting cells in the organ of Corti, which occurred
in parallel with the production and expression of pro-inflammatory
cytokines (11). Nuclear factor
(NF)-κB was reported to be involved in the regulation of cell
proliferation, survival, angiogenesis, apoptosis and
differentiation (12,13). NF-κB was shown to activate genes
inside the nucleus, which are involved in the regulation of
apoptosis, the inflammatory response and oxidative stress (14,15).
Numerous studies have indicated that NF-κB may have a significant
role in the mediation of hearing loss. Ototoxic stimulants,
including noise exposure and ototoxic drugs, were found to induce
the activation of NF-κB (16,17),
which results in damage to the cochlear lateral wall via the
production of large amounts of reactive oxygen species (ROS)
(18,19). Caspase-1, a member of the caspase
family characterized by large pro-domains (20), has been reported to be involved in
apoptosis and inflammation following activation (21). Caspase-1 activation results in
inflammation through the generation of pro-inflammatory cytokines
(22). In addition, caspase-1 has
a significant role in the cisplatin-induced apoptosis of cochlear
hair cells and spiral ganglion neurons.
Korean red ginseng (KRG) has previously been used as
a supplementary herbal medicine in the treatment of numerous
diseases, including inflammation, diabetes and atherosclerosis
(23,24). However, the underlying protective
mechanisms of KRG in the context of cisplatin have remained to be
elucidated. The present study aimed to further investigate the
mechanism of KRG in cisplatin-induced toxicity in auditory HEI-OC1
cells. Furthermore, the protective effects of KRG on
cisplatin-induced hair cell damage were investigated in the organ
of Corti in a rat model.
Materials and methods
Reagents
Fetal bovine serum (FBS) and high-glucose Dulbecco’s
modified Eagle’s medium (DMEM) were purchased from Gibco-BRL (Grand
Island, NY, USA). cis-Diamineplatinum(II) dichloride (cisplatin;
cat. no. 479306), paraformaldehyde, Tween® 20,
avidinperoxidase, Triton X-100 and liquid nitrogen were purchased
from Sigma-Aldrich (St. Louis, MO, USA). 10X phosphate-buffered
saline was purchased from bioWORLD (Dublin, OH, USA). ABTS was
purchased from BD Bioscience (San Diego, CA, USA). Rabbit
polyclonal NF-κB (cat. no. sc-7151), mouse monoclonal cytochrome
(cyt) c (cat. no. sc-13156), mouse monoclonal α-tubulin
(cat. no. sc-8035), rabbit polyclonal histone H1 (cat. no.
sc-10806) and rabbit polyclonal caspase-3 (cat. no. sc-7148)
antibodies (Abs) were obtained from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). Caspase-1, caspase-3 and caspase-9 colorimetric
kits were supplied by R&D systems, Inc. (Minneapolis, MN, USA).
Purified anti-mouse interleukin (IL)-6 (cat. no. 554400),
biotinylated IL-6 (cat. no. 554402) and recombinant IL-6 (cat. no.
554401) Abs were obtained from BD-Pharmingen (San Diego, CA, USA).
Peroxidase-conjugated AffiniPure goat anti-rabbit immunoglobulin
(Ig)G (cat. no. 111-035-003) and peroxidase-conjugated AffiniPure
goat anti-mouse IgG (cat. no. 115-035-062) Abs were purchased from
Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA, USA).
Nonidet P (NP)-40 was purchased from Abcam (Cambridge, UK).
Preparation of KRG
KRG was manufactured by the Korea Ginseng
Corporation (Seoul, Korea). Extract was prepared by decocting the
KRG with boiling distilled water (100 g/l). The extraction was
decocted for ~3 h, and was subsequently filtered with Labsystem 2
(EMD Millipore, Billerica, MA, USA), lyophilized with a freeze
dryer (Ilshin Bio Base Co., Ltd., Yangju, Korea) and stored at 4°C.
Dilutions were made using saline and filtered through a
0.22-μm Millex-GS Syringe Filter unit (EMD Millipore). The
KRG extracts contained the following seven glycosides, known as
ginsenosides (mg/g): Rg1 (2.481), Rb1
(5.481), Rg3(S) (0.197), Re (2.975), Rc (2.248),
Rb2 (2.175) and Rb (0.566) (25).
Cell culture
The HEI-OC1 cell line was provided by Dr Federico
Kalinec (House Ear Institute, Los Angeles, CA, USA), which was
produced as previously described (26). The passage number of the cells used
was five, and to avoid mycoplasmic infection, the provided cell
stocks were reconstituted with Mycoplasma Removal Agent (cat. no.
9030500) from MP Biochemicals (Santa Ana, CA, USA). HEI-OC1 cells
express multiple molecular markers that are characteristic of the
organ of Corti sensory cells, including α-thyroid hormone,
brain-derived neurotrophic factor, calbindin, calmodulin, Connexin
26, Math 1, Myosin 7a, organ of Corti protein 2, tyrosine kinase
receptors B and C, platelet-derived growth factor receptor and
prestin; in addition, HEI-OC1 cells are particularly sensitive to
ototoxic drugs (26). These cells
were maintained in DMEM supplemented with 10% FBS at 33°C with 5%
CO2. Cells were treated with various concentrations of
KRG (0.5, 1 and 2 mg/ml) for 1 h, and subsequently treated with
cisplatin (20 μM) for 48 h at 33°C.
Ethics statement
All animal procedures and experiments were approved
by the Animal Care and Use Committee of Wonkwang University
(approval no. WKU14-05; Iskan, Korea). All animals were treated
humanely and with regard for alleviation of suffering.
Organ of Corti explant culture
The organ culturing procedure was similar to that
described previously (27). In
brief, two-day-old Sprague Dawley rats (n=5/group; weighing 6.5±0.5
g) were anesthetized with CO2 and sacrificed by
decapitation. Subsequently, the cochlea was carefully removed by
dissection. The cochlear explants were treated with KRG (2 mg/ml)
for 1 h, and subsequently treated with cisplatin (30 μM) for
24 h at 33°C. When the experimental treatment was completed, the
culture was prepared for histological analysis. The organ of Corti
explants were fixed for 15 min in 2% paraformaldehyde in PBS. The
specimens were rinsed in PBS, incubated in 0.25% Triton X-100 for 2
min and immersed in tetramethylrhodamine (TRITC)-labeled phalloidin
(1:100; Sigma-Aldrich) in PBS for 20 min. Following rinsing with
PBS, the specimens were examined using fluorescence microscopy
(IX71, Olympus Corporation, Tokyo, Japan) with the appropriate
filters for TRITC.
Animal experiment
All experiments were performed using male Balb/c
mice (four-weeks-old; weighing 16–18 g), which were housed in
stainless steel cages in a temperature controlled (25°C) room
equipped to maintain a 12-h light/dark cycle. The animals were
distributed at random into three groups (n=5 per group) and fed a
regular diet. Group 1, regarded as the control group, received
intraperitoneal (i.p.) injection of 200 μl PBS; group 2 was
administered cisplatin (4 mg/kg) via i.p. injection for four
consecutive days; and group 3 was orally administered KRG (200
mg/kg) once a day for four days; and group 4 was orally
administered KRG (200 mg/kg) 1 h prior to i.p injection of
cisplatin (4 mg/kg). An auditory brainstem response (ABR) test was
subsequently performed to investigate the effect of KRG on hearing
function. Following the ABR test, all of the mice were sacrificed
by cervical dislocation.
Assessment of hearing function
ABR tests were conducted under ketamine/xylazine
(equivalent to 172.4 mg/kg ketamine and 5.5 mg/kg xylazine)
sedation. The ABR was used to determine the auditory threshold of
mice. While under sedation, ABR testing was performed on the
animals in response to 4, 8 and 32 kHz tone bursts. A
computer-based signal-averaging system (ABR workstation with SigGen
software) from Tucker Davis Technologies (Gainesville, FL, USA) was
used to compile ABR data. The ABR was recorded using three
platinum-iridium needle electrodes (Ambu® Neuroline
Disposable Subdermal Needle Electrodes; Cadwell, Kennewick, WA,
USA), positioned subdermally over the vertex (positive), mastoid
(negative) and dorsum (reference/ground) areas of the animal. Sound
was generated through an Etymotic ER-2 earphone (Etymotic Research,
Elk Grove Village, IL, USA), which was placed directly in the ear
canal. The ABR threshold began at 90 dB and decreased in 10 dB
steps, and each response was repeated in triplicate.
Cytokine assays
Expression levels of IL-6 in HEI-OCI cells were
examined using a modified ELISA. In brief, the ELISA was set up by
coating 96-well plates with mouse monoclonal Ab against IL-6. The
coated plates were then washed with PBS containing 0.05% Tween 20.
All reagents used in this assay were incubated for 2 h at 37°C.
Recombinant IL-6 was diluted and used as a standard. Standards were
diluted 1:1 with diluents (PBS with 10% FBS) from 10 ng/ml to
0.34125 ng/ml and were used to construct the standard curve.
Following 2 h incubation of standards and cell supernants
(3×105 cells/well) at room temperature, the assay plates
were sequentially exposed to biotinylated mouse IL-6, avidin
peroxidase and [2,2′-Azinobis(3-ethylbenzothiazoline-6-sulfonic
acid)-diammonium salt] substrate solution containing 30%
H2O2. The plates were read at a wavelength of
405 nm using a VersaMax™ microplate reader (Molecular Devices,
Sunnyvale, CA, USA).
Spectrofluorimetric measurement of
intracellular ROS production
Intracellular ROS levels in HEI-OCI cells were
measured using fluorescent dye, 2′,7′-dichlorofluorescein diacetate
(DCFH-DA). In the presence of an oxidant, DCFH is converted into a
highly fluorescent molecule, 2′,7′-dichlorofluorescein (DCF). Cells
(3×105 cells/well) were treated with KRG (0.5, 1 and 2
mg/ml) for 1 h, cultured in the presence or absence of cisplatin
(20 μM) for 48 h and subsequently incubated with 5 μM
DCFH-DA for 30 min. The fluorescence intensity was measured using a
spectrofluorometer (RF-5301PC; Shimadzu Corp., Kyoto, Japan) at
excitation and emission wavelengths of 485 and 538 nm,
respectively.
Preparation of cytoplasmic and nuclear
extracts
Nuclear and cytoplasmic extracts were prepared as
described previously (28). In
brief, following activation of the HEI-COI cells (7×106
cells/well) with cisplatin, they were washed with ice-cold PBS.
These cells were resuspended in 60 μl buffer A [10 mM
Hepes/KOH, 2 mM MgCl2, 0.1 mM EDTA, 10 mM KCl, 1 mM
dithiothreitol (DTT) and 0.5 mM phenylmethylsulfonyl fluoride
(PMSF); pH 7.9]. The cells were allowed to swell on ice for 15 min,
lysed gently with 2.5 μl of 10% NP-40 and centrifuged at
2,000 × g for 10 min at 4°C. The supernatant was collected and used
as the cytoplasmic extract. The nuclei pellet was resuspended in 40
μl of buffer B (50 mM HEPES/KOH, 50 mM KCl, 300 mM NaCl, 0.1
mM EDTA, 10% glycerol, 1 mM DTT and 0.5 mM PMSF; pH 7.9), left on
ice for 20 min, inverted and centrifuged at 15,000 × g for 15 min
to remove nuclear debris. The supernatant (nuclear extract) was
collected, frozen in liquid nitrogen and stored at −70°C until
required for further analysis.
Western blot analysis
For analysis of the expresssion levels of cyt
c, the cells (5×106 cells/well) were rinsed twice
with ice-cold PBS and then lysed in ice-cold lysis buffer (1%
Triton, 1% NP-40, 0.1% SDS and 1% deoxycholate in PBS). Samples
were centrifuged at 15,000 × g for 5 min at 4°C and the
supernatants were mixed with an equal volume of 2X SDS sample
buffer (ELIPIS-Biotech, Inc., Dajeon, Korea), boiled for 5 min and
separated through a 10% denaturing protein gel. Following
electrophoresis, the proteins were transferred to nitrocellulose
membranes (GE Healthcare Life Sciences, Uppsala, Sweden) by
electrophoretic transfer. The membranes were blocked in 5% Difco
skim milk (BD Diagnostics, Sparks Glencoe, MD, USA) for 2 h, rinsed
and incubated overnight at 4°C with primary antibodies (1:500).
Following three washes (30 min per wash) in PBS/0.5% Tween 20, the
membranes were incubated for 1 h with horseradish
peroxidase-conjugated anti-rabbit immunoglobulin (secondary
antibodies). Following three washes (30 min per wash) in PBS/0.1%
Tween 20, the protein bands were visualized using an enhanced
chemiluminescence assay (Amersham Pharmacia, Piscataway, NJ, USA)
according to the manufacturer’s instructions and subsequently
exposed to X-ray film (Agfa-Gevaert N.V, Mortsel, Belgium). The
relative protein expression levels were evaluated using Image J
software, version 1.48.
Luciferase assay
Cells were transiently transfected with NF-κB
luciferase reporter plasmid (Stratagene, Cedar Creek, TX, USA)
using the transfection reagent, Lipofectamine 2000 (Invitrogen Life
Technologies, Carlsbad, CA, USA). Following 36-h incubation, the
cells were treated with various concentrations of KRG (1 and 2
mg/ml) for 1 h and then treated with cisplatin (20 μM) for
24 h. The cells were then washed twice with PBS buffer and
subsequently lysed in reporter lysis buffer (Promega Corporation,
Madison, WI, USA). A 20-μl aliquot of the lysate was then
mixed with 100 μl luciferase assay reagent, after which the
emitted light intensity was measured using an AutoLumat LB 953
luminometer (Berthold Technologies GmbH & Co. KG, Bad Wildbad,
Germany). Finally, the luciferase activity was measured in
triplicate, averaged, and then normalized against the
β-galactosidase activity using the galactosidase assay system
(Galacto-Light; Tropix, Inc., Bedford, MA, USA) according to the
manufacturer’s instructions.
Assay of caspase-1, -3 and -9
activity
The enzymatic activity of caspase-1, -3 and -9 was
evaluated using a caspase colorimetric assay kit (R&D Systems,
Inc.) according to the manufacturer’s instructions. Cells
(5×106 cells/well) were treated with various
concentrations of KRG (0.5, 1 and 2 mg/ml) for 1 h and then treated
with cisplatin (20 μM) for 48 h. In brief, the cytoplasmic
protein supernatant was incubated with 50 μl reaction buffer
and 5 μl caspase substrate at 37°C for 2 h. The absorbance
was measured using a plate reader (VersaMax™) at a wavelength of
405 nm. Equal amounts of the total protein from each lysate were
quantified using a bicinchoninic acid (BCA) protein quantification
kit (Sigma-Aldrich) according to the manufacturer’s instructions.
Inhibition rate was calculated as follows: Inhibition rate=[(KRG
OD)−(control OD)]/[(cisplatin-only OD)−(control OD)]×100%; where OD
stands for optical density.
Statistical analysis
Values are presented as the mean ± standard error of
the mean of at least three independent experiments. Statistical
analysis was performed using one-way analysis of variance with
Tukey’s post hoc test (PASW Statistics 18.0.0.; SPSS Inc., Chicago,
IL, USA) P<0.05 was considered to indicate a statistically
significant difference between values.
Results
KRG attenuates cisplatin-induced changes
in the hearing threshold of mice
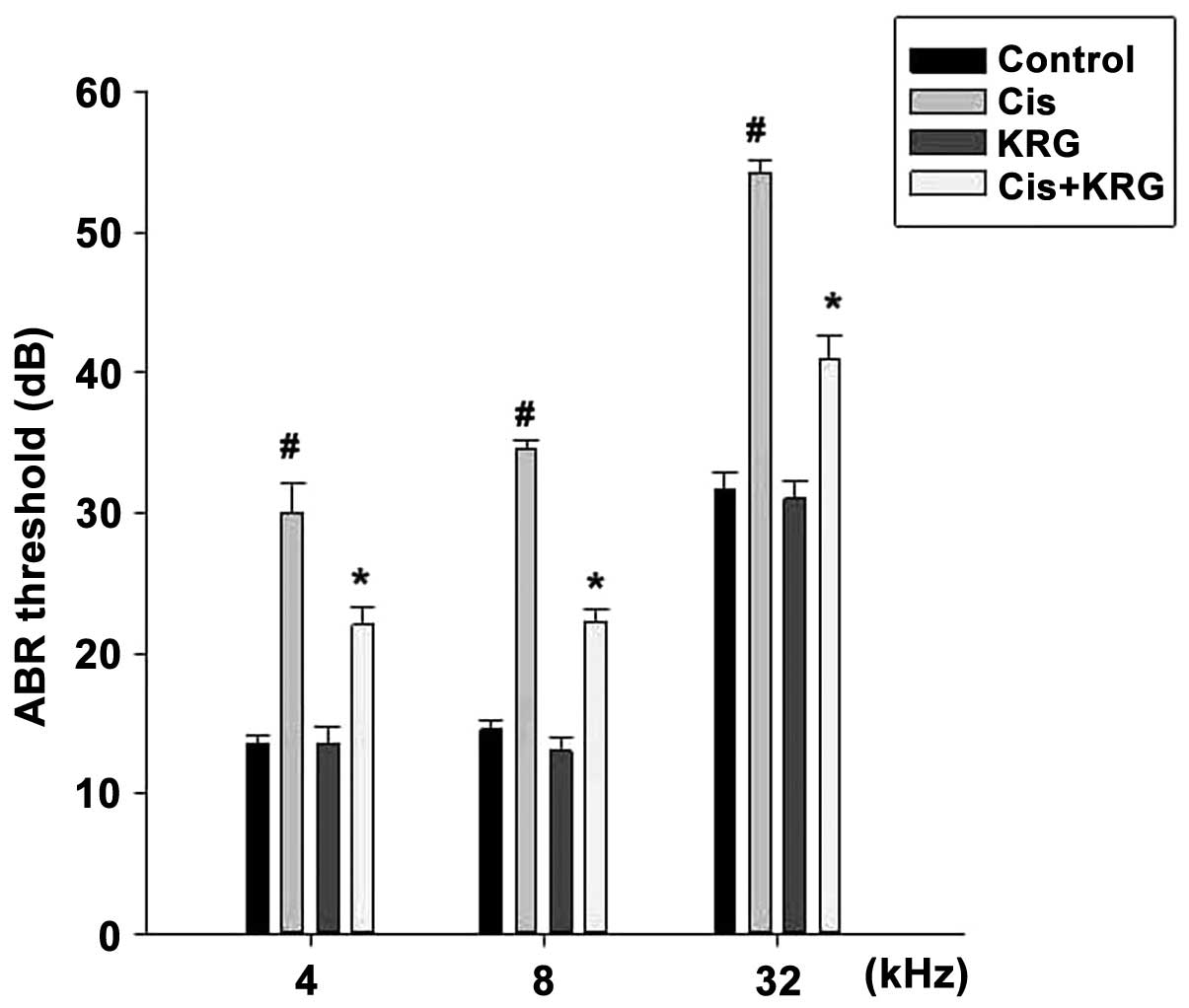
In order to investigate the effect of KRG on hearing
function, ABR was recorded at 4, 8 and 32 kHz. The results
indicated that the cisplatin-only group had a significantly
increased ABR threshold shift at 4, 8 and 32 kHz compared with that
of the control group (P<0.05). However, KRG (200 mg/kg) was
shown to significantly attenuate the cisplatin-induced hearing
threshold shift (Fig. 1).
KRG attenuates cisplatin-induced toxicity
in organ of Corti explants
The organ of Corti was isolated from rats on
postnatal day two and treated with cisplatin (30 μM) in
presence of KRG (2 mg/ml). As indicated in Fig. 2A, cisplatin treatment impaired the
orderly arrangements of the three rows of OHCs and a single row of
inner hair cells (IHCs) in the organ of Corti. However, treatment
with KRG in combination with cisplatin significantly attenuated
this effect (P<0.05). The relative hair cell viability is
presented in Fig. 2B, and
indicates a significant decrease in hair cell viability in the
cisplatin-treated group, compared with that of the control group
(P<0.05) that is attenuated following KRG pretreatment
(P<0.05).
KRG suppresses the cisplatin-induced
increase in IL-6 levels and ROS generation in HEI-OC1 cells
ROS generated in the mitochondria may induce damage
to the mitochondrial membrane, resulting in apoptosis (29). In order to determine the regulatory
effects of KRG on ROS production, cells were treated with DCFH-DA,
which is converted to a fluorescent substance following
intracellular oxidation. The results revealed that KRG effectively
suppressed the significant increase in ROS levels induced by
cisplatin treatment (P<0.05; Fig.
3A).
Previous studies have reported that the expression
of inflammatory cytokines is significantly enhanced in the inner
ear under various damaging conditions (30–33).
In order to determine whether KRG was able to regulate the enhanced
IL-6 levels induced by cisplatin, ELISAs were performed. As shown
in Fig. 3B, IL-6 levels were
significantly increased following treatment with cisplatin compared
with those of the control group (P<0.05); however, this effect
was markedly reduced following treatment with KRG (P<0.05).
KRG inhibits the increased
apoptosis-associated gene levels induced by cisplatin in HEI-OCI
cells
Pro-apoptotic stimuli induce the release of cyt
c into the cytosol, which leads to the activation of
pro-apoptotic factors as well as the maturation of caspase-3
(34,35). As shown in Fig. 4A and B, the effect of KRG on
cisplatin-induced cyt c release was examined. The results
revealed that cisplatin significantly increased the quantity of cyt
c released into the cytosol compared with that of the
control group (P<0.05); however, KRG significantly inhibited
cisplatin-induced cyt c release (P<0.05).
Caspase-3 is known to have a significant role in the
initiation and implementation of apoptosis (36,37).
In the present study, the inhibitory effect of KRG on caspase-9 and
caspase-3 activity was examined. Cisplatin significantly induced
caspase-9 and caspase-3 activation compared with that of the
control group (P<0.05), while KRG effectively inhibited this
induction (P<0.05) (Fig. 4C and
4D).
KRG inhibits cisplatin-induced NF-κB
activation in HEI-OCI cells
Reduced NF-κB activation has previously been
associated with hearing loss (38,39);
therefore, it was hypothesized that the effects of KRG may be
mediated, at least in part, by the suppression of NF-κB activation.
In addition, NF-κB activation requires the nuclear translocation of
the RelA/p65 subunit of NF-κB (40); therefore, the present study aimed
to evaluate the effects of KRG on the nuclear pool of RelA/p65
protein using western blot analysis. In cisplatin-treated cells,
the levels of Rel/p65 were markedly increased compared with those
of the untreated group; however, KRG reduced these enhanced nuclear
levels of Rel/p65 (Fig. 5A). The
relative nuclear levels of NF-κB are exhibited in Fig. 5B. The cisplatin-induced increase in
NF-κB expression was significantly attenuated following KRG
pretreatment in a dose-dependent manner.
Furthermore, a luciferase assay was performed in
order to investigate the effects of KRG on NF-κB promoter activity.
As shown in Fig. 5C, cisplatin
treatment enhanced NF-κB promoter activity compared with that of
the control group (P<0.05), while KRG treatment inhibited this
cisplatin-induced increase in NF-κB promoter activity
(P<0.05).
KRG attenuates cisplatin-induced
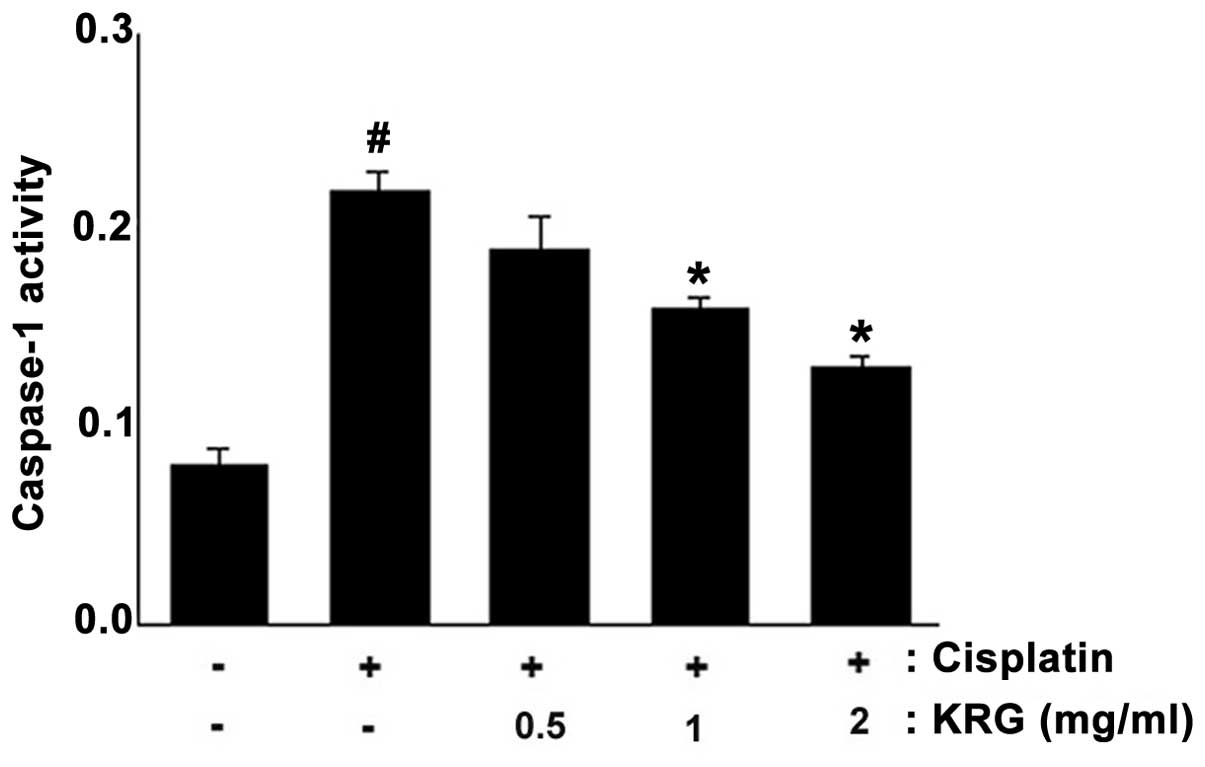
caspase-1 activation in HEI-OCI cells
The activation of caspase-1 induces the activation
of NF-κB and production of inflammatory cytokines (41). In order to elucidate the mechanism
underlying the protective effects of KRG on ototoxicity, the
effects of KRG on caspase-1 activation were evaluated. The results
demonstrated that the enhanced caspase-1 activity induced by
cisplatin was significantly reduced by KRG in a dose-dependent
manner (P<0.05) (Fig. 6). The
maximal inhibition rate of KRG (2 mg/ml) was 42.21±3.1% compared
with that of the cisplatin-only group.
Discussion
KRG has been reported to have numerous beneficial
therapeutic effects including anti-cancer, anti-inflammatory and
anti-oxidant activities (42).
However, the underlying protective mechanisms of KRG in the
auditory system have remained to be fully elucidated. The results
of the present study demonstrated that KRG attenuated the adverse
effects of cisplatin in the organ of Corti as well as apoptosis in
auditory cells.
Cisplatin is an effective, commonly used anticancer
agent; however, its major side effects include hearing damage. A
previous study investigated the mechanisms underlying
cisplatin-induced ototoxicity (43). The results of the present study
demonstrated that KRG pretreatment effectively prevented
cisplatin-induced impairment of hair cell arrays in the rat organ
of Corti primary explants. Cisplatin has been confirmed to induce
the apoptosis of auditory sensory cells (44). The mitochondrial apoptotic pathway
has been demonstrated to be activated by the release of proteins
from the mitochondrial intermembrane space into the cytosol, which
subsequently initiate the activation of apoptotic pathways
(44). Cyt c is a
mitochondrial protein, which induces caspase-dependent cell
apoptotsis; cyt c initiates caspase-9 activation, which in
turn activates caspase-3 (45). It
was reported that cisplatin-mediated ototoxicity was associated
with caspase-3 activity, cyt c release and ROS generation in
auditory cells (46,47). These previous studies therefore
indicated that mitochondrial signaling pathways may represent
potential therapeutic targets for use in the prevention of
cisplatin-mediated ototoxic damage. In the present study, KRG was
demonstrated to inhibit the cyt c release, ROS generation
and caspase-3 activation induced by cisplatin. These results
therefore suggested that the protective effect of KRG in auditory
cells occurred via the regulation of caspase-mediated apoptosis.
Although KRG attenuated the caspase-mediated pathway, the present
study did not investigate the effects of KRG on non-mitochondrial
signaling pathways. Therefore, further studies are required in
order to evaluate the potential role of KRG on non-mitochondrial
signaling pathways in the auditory system.
Numerous previous studies have reported that
inflammatory cytokines are associated with hearing loss (7–10,32).
These cytokines were reported to induce secondary inflammatory
responses, including leukocyte infiltration and scar formation. In
addition, it was reported that cisplatin-induced cochlear injury
occured via pro-inflammatory cytokine expression (11). Flunarizine, a T-type
Ca2+ channel antagonist, was demonstrated to have
cyto-protective effects against cisplatin cytotoxicity, the
mechanism of which was shown to proceed via the inhibition of
pro-inflammatory cytokine expression in HEI-OC1 cells (46). It was therefore hypothesized that
the protective effect of KRG may occur via the regulation of
pro-inflammatory cytokines. In the present study, it was
demonstrated that KRG inhibited IL-6 expression levels, which
indicated that the protective effect of KRG against cisplatin
cytotoxicity occurred due to the suppression of inflammatory
cytokine levels in auditory cells. Although it was demonstrated
that KRG attenuated IL-6 expression levels, the effects of KRG on
other cytokine pathways, or any pathways that may be involved, were
not evaluated. Therefore, further studies are required in order to
clarify the role of KRG in the cytokine-associated pathways in the
auditory system.
The association between NF-κB activation and hearing
loss has been investigated in numerous previous studies (38,39,48).
Certain studies have reported that NF-κB family proteins identified
in the inner ear were essential for the regular function of hair
cells (49), while other studies
have demonstrated that signal-transduction pathways respond rapidly
to ototoxic stimulants, including noise exposure (38) and ototoxic drugs (16,50,51).
NF-κB activation was reported to induce damage to the cochlear
lateral wall through the generation of large quantities of ROS
(52). In addition, acoustic
overstimulation enhances inflammatory factor expression levels via
NF-κB activation in the inner ear (39). The results of these studies
suggested that NF-κB activation may be the primary target for the
prevention of cisplatin-induced ototoxic damage. Therefore, in the
present study, whether the protective mechanism of KRG proceeded
via NF-κB pathway regulation was investigated. The results revealed
that KRG attenuated the cisplatin-induced activation of NF-κB in
auditory cells.
The activation of caspase-1 is known to be involved
in the mediation of apoptosis and inflammation (21). Activated caspase-1 induces
inhibitor of κB (IκB) kinase (IKK) phosphorylation by RIP2
dependent pathway. Caspase-1 interacts with RIP2 through the
caspase recruitment domain, then RIP2 recruits the IKK complex via
interaction with IKK-γ. This is independent of the enzymatic
activity of caspase-1, and results in IκB-α degradation and NF-κB
activation (41). Therefore,
released NF-κB is translocated to the nucleus, where it is involved
in the induction of gene transcription (53). The addition of pro-inflammatory
cytokines, including IL-1β, has previously been shown to increase
the nuclear translocation of NF-κB in HEI-OC1 auditory cells
(11). Caspase-1 may contribute to
NF-κB activation via the autocrine action of IL-1β. Therefore, it
was hypothesized that the NF-κB pathway may interact with caspase-1
in auditory cells. Zhang et al (54) reported that caspase-1 initiated cyt
c release and caspase-3 activation in
ischemia/hypoxia-mediated neuronal cell death. In addition, it was
demonstrated that cisplatin induced caspase-1 activation in
cochlear hair cells and spiral ganglion neurons (55). In the present study, KRG was found
to inhibit cisplatin-induced caspase-1 activation. These results
suggested that the protective mechanism of KRG may be mediated via
the modulation of signaling molecules involved cisplatin-induced
apoptosis.
In conclusion, KRG was demonstrated to attenuate
ototoxicity through prevention of the impairment of the hair cell
arrays in organ of Corti rat primary explants. In addition, KRG
inhibited cisplatin-induced ROS production, enhanced IL-6 levels,
cyt c release, caspase-3 activation and NF-κB translocation
to the nucleus in auditory cells. Therefore, these results have
contributed to the elucidation of the pharmacological mechanism
underlying KRG function and have provided evidence for potential
therapeutic approaches for the treatment of ototoxicity.
Acknowledgments
The present study was supported by the National
Research Foundation of Korea grants funded by the Korean government
(no. 2011-0006220).
References
|
1
|
Cohen SM and Lippard SJ: Cisplatin: from
DNA damage to cancer chemotherapy. Prog Nucleic Acid Res Mol Biol.
67:93–130. 2001.PubMed/NCBI
|
|
2
|
Santos NA, Catão CS, Martins NM, Curti C,
Bianchi ML and Santos AC: Cisplatin-induced nephrotoxicity is
associated with oxidative stress, redox state unbalance, impairment
of energetic metabolism and apoptosis in rat kidney mitochondria.
Arch Toxicol. 81:495–504. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yao X, Panichpisal K, Kurtzman N and
Nugent K: Cisplatin nephrotoxicity: a review. Am J Med Sci.
334:115–124. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu YN, Chen H and Li Yi: Protect effect of
bicyclol on cisplatin-induced nephrotoxicity in mice. Arch Toxicol.
83:381–387. 2009. View Article : Google Scholar
|
|
5
|
Jordan P and Carmo-Fonseca M: Molecular
mechanisms involved in cisplatin cytotoxicity. Cell Mol Life Sci.
57:1229–1235. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Satoh H, Firestein GS, Billings PB, Harris
JP and Keithley EM: Proinflammatory cytokine expression in the
endolymphatic sac during inner ear inflammation. J Assoc Res
Otolaryngol. 4:139–147. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Keithley EM, Wang X and Barkdull GC: Tumor
necrosis factor alpha can induce recruitment of inflammatory cells
to the cochlea. Otol Neurotol. 29:854–859. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wakabayashi K, Fujioka M, Kanzaki S, et
al: Blockade of interleukin-6 signaling suppressed cochlear
inflammatory response and improved hearing impairment in
noise-damaged mice cochlea. Neurosci Res. 66:345–352. 2010.
View Article : Google Scholar
|
|
9
|
Oh GS, Kim HJ, Choi JH, et al: Activation
of lipopolysaccharide-TLR4 signaling accelerates the ototoxic
potential of cisplatin in mice. J Immunol. 186:1140–1150. 2011.
View Article : Google Scholar
|
|
10
|
Kim HJ, Oh GS, Lee JH, et al: Cisplatin
ototoxicity involves cytokines and STAT6 signaling network. Cell
Res. 21:944–956. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
So H, Kim H, Lee JH, et al: Cisplatin
cytotoxicity of auditory cells requires secretions of
proinflammatory cytokines via activation of ERK and NF-kappaB. J
Assoc Res Otolaryngol. 8:338–355. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bharti AC, Donato N, Singh S and Aggarwal
BB: Curcumin (diferuloylmethane) down-regulates the constitutive
activation of nuclear factor-kappaB and IkappaBalpha kinase in
human multiple myeloma cells, leading to suppression of
proliferation and induction of apoptosis. Blood. 101:1053–1062.
2003. View Article : Google Scholar
|
|
13
|
Estrov Z, Shishodia S, Faderl S, et al:
Resveratrol blocks interleukin-1β-induced activation of the nuclear
transcription factor NF-κB, inhibits proliferation, causes S-phase
arrest and induces apoptosis of acute myeloid leukemia cells.
Blood. 102:987–995. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Christman JW, Sadikot RT and Blackwell TS:
The role of nuclear factor-κB in pulmonary diseases. Chest.
117:1482–1487. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Janssen-Heininger YM, Poynter ME and
Baeuerle PA: Recent advances towards understanding redox mechanisms
in the activation of nuclear factor kappaB. Free Radic Biol Med.
28:1317–1327. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang H, Sha SH and Schacht J: NF-κB
pathway protects cochlear hair cells from aminoglycoside-induced
ototoxicity. J Neurosci Res. 79:644–651. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nagashima R, Sugiyama C, Yoneyama M,
Kuramoto N, Kawada K and Ogita K: Acoustic overstimulation
facilitates the expression of glutamate-cysteine ligase catalytic
subunit probably through enhanced DNA binding of activator
protein-1 and/or NF-κB in the murine cochlea. Neurochem Int.
51:209–215. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi X and Nuttall AL: Expression of
adhesion molecular proteins in the cochlear lateral wall of normal
and PARP-1 mutant mice. Hear Res. 224:1–14. 2007. View Article : Google Scholar
|
|
19
|
Masuda M, Nagashima R, Kanzaki S, Fujioka
M, Ogita K and Ogawa K: Nuclear factor-kappaB nuclear translocation
in the cochlea of mice following acoustic overstimulation. Brain
Res. 1068:237–247. 2006. View Article : Google Scholar
|
|
20
|
Lee SH, Stehlik C and Reed JC: Cop, a
caspase recruitment domain-containing protein and inhibitor of
caspase-1 activation processing. J Biol Chem. 276:34495–34500.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang X, Wang H, Figueroa BE, et al:
Dysregulation of receptor interacting protein-2 and caspase
recruitment domain only protein mediates aberrant caspase-1
activation in Huntington’s disease. J Neurosci. 25:11645–11654.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Faubel S, Lewis EC, Reznikov L, et al:
Cisplatin-induced acute renal failure is associated with an
increase in the cytokines interleukin (IL)-1beta, IL-18, IL-6 and
neutrophil infiltration in the kidney. J Pharmacol Exp Ther.
322:8–15. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sung J, Han KH, Zo JH, Park HJ, Kim CH and
Oh BH: Effects of red ginseng upon vascular endothelial function in
patients with essential hypertension. Am J Chin Med. 28:205–216.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ryu JK, Lee T, Kim DJ, et al: Free
radical-scavenging activity of Korean red ginseng for erectile
dysfunction in non-insulin-dependent diabetes mellitus rats.
Urology. 65:611–615. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hong SH, Suk KT, Choi SH, et al:
Anti-oxidant and natural killer cell activity of Korean red ginseng
(Panax ginseng) and urushiol (Rhus vernicifera Stokes) on
non-alcoholic fatty liver disease of rat. Food Chem Toxicol.
55:586–591. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kalinec GM, Webster P, Lim DJ and Kalinec
F: A cochlear cell line as an in vitro system for drug ototoxicity
screening. Audiol Neurootol. 8:177–189. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng JL and Gao WQ: Differential damage
to auditory neurons and hair cells by ototoxins and neuroprotection
by specific neuro-trophins in rat cochlear organotypic cultures.
Eur J Neurosci. 8:1897–1905. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schoonbroodt S, Legrand-Poels S,
Best-Belpomme M and Piette J: Activation of the NF-kappaB
transcription factor in a T-lymphocytic cell line by hypochlorous
acid. Biochem J. 321:777–785. 1997.PubMed/NCBI
|
|
29
|
Orrenius S: Reactive oxygen species in
mitochondria-mediated cell death. Drug Metab Rev. 39:443–455. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fujioka M, Kanzaki S, Okano HJ, Masuda M,
Ogawa K and Okano H: Proinflammatory cytokines expression in
noise-induced damaged cochlea. J Neurosci Res. 83:575–583. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ghaheri BA, Kempton JB, Pillers DA and
Trune DR: Cochlear cytokine gene expression in murine acute otitis
media. Laryngoscope. 117:22–29. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim HJ, So HS, Lee JH, et al: Role of
proinflammatory cytokines in cisplatin-induced vestibular hair cell
damage. Head Neck. 30:1445–1456. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ladrech S, Mathieu M, Puel JL and Lenoir
M: Supporting cells regulate the remodelling of
aminoglycoside-injured organ of Corti, through the release of high
mobility group box 1. Eur J Neurosci. 38:2962–2972. 2013.PubMed/NCBI
|
|
34
|
Gorman AM, Bonfoco E, Zhivotovsky B,
Orrenius S and Ceccatelli S: Cytochrome c release and caspase-3
activation during colchicine-induced apoptosis of cerebellar
granule cells. Eur J Neurosci. 11:1067–1072. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Abu-Qare AW and Abou-Donia MB: Biomarkers
of apoptosis: release of cytochrome c, activation of caspase-3,
induction of 8-hydroxy-2′-deoxyguanosine, increased
3-nitrotyrosine, and alteration of p53 gene. J Toxicol Environ
Health B Crit Rev. 4:313–332. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Brentnall M, Rodriguez-Menocal L, De
Guevara RL, Cepero E and Boise LH: Caspase-9, caspase-3 and
caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell
Biol. 14:322013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lang H, Schulte BA, Zhou D, Smythe N,
Spicer SS and Schmiedt RA: Nuclear factor kappaB deficiency is
associated with auditory nerve degeneration and increased
noise-induced hearing loss. J Neurosci. 26:3541–3550. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yamamoto H, Omelchenko I, Shi X and
Nuttall AL: The influence of NF-κB signal-transduction pathways on
the murine inner ear by acoustic overstimulation. J Neurosci Res.
87:1832–1840. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Stark LA and Dunlop MG: Nucleolar
sequestration of RelA (p65) regulates NF-kappaB-driven
transcription and apoptosis. Mol Cell Biol. 25:5985–6004. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lamkanfi M, Kalai M, Saelens X, Declerq W
and Vandenabeele P: Caspase-1 activates nuclear factor of the
kappa-enhancer in B cells independently of its enzymatic activity.
J Biol Chem. 279:24785–24793. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park K-S, Hwang D-S, Lee J-M, Jang J-B,
Lee K-S and Lee C-H: Inhibitory effect of Panax ginseng C. A. Meyer
on gynecological cancer. Orient Pharm Exp Med. 13:217–223. 2013.
View Article : Google Scholar
|
|
43
|
Rybak LP, Whitworth CA, Mukherjea D and
Ramkumar V: Mechanisms of cisplatin-induced ototoxicity and
prevention. Hear Res. 226:157–167. 2007. View Article : Google Scholar
|
|
44
|
Devarajan P, Savoca M, Castaneda MP, et
al: Cisplatin-induced apoptosis in auditory cells: role of death
receptor and mitochondrial pathways. Hear Res. 174:45–54. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Roy S, Banerjee B and Vedasiromoni J:
Anti-tumor activity of Swietenia mahagoni (L.) Jacq. leaf extract
against Ehrlich’s ascites carcinoma in mice. Orient Pharm Exp Med.
13:205–216. 2013. View Article : Google Scholar
|
|
46
|
So HS, Park C, Kim HJ, et al: Protective
effect of T-type calcium channel blocker flunarizine on
cisplatin-induced death of auditory cells. Hear Res. 204:127–139.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee S, Moon SO, Kim W, et al: Protective
role of L-2-oxothiazolidine-4-carboxylic acid in cisplatin-induced
renal injury. Nephrol Dial Transplant. 21:2085–2095. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tahera Y, Meltser I, Johansson P, et al:
NF-kappaB mediated glucocorticoid response in the inner ear after
acoustic trauma. J Neurosci Res. 83:1066–1076. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shishodia S and Aggarwal BB: Nuclear
factor-kappaB: a friend or a foe in cancer? Biochem Pharmacol.
68:1071–1080. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Nagy I, Monge A, Albinger-Hegyi A, Schmid
S and Bodmer D: NF-kappaB is required for survival of immature
auditory hair cells in vitro. J Assoc Res Otolaryngol. 6:260–268.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kim SJ, Jeong HJ, Myung NY, et al: The
protective mechanism of antioxidants in cadmium-induced ototoxicity
in vitro and in vivo. Environ Health Perspect. 116:854–862. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Watanabe K, Inai S, Jinnouchi K, et al:
Nuclear-factor kappaB (NF-kappaB)-inducible nitric oxide synthase
(iNOS/NOS II) pathway damages the stria vascularis in
cisplatin-treated mice. Anticancer Res. 22:4081–4085. 2002.
|
|
53
|
Chu SH, Lim JW, Kim DG, Lee ES, Kim KH and
Kim H: Down-regulation of Bcl-2 is mediated by NF-κB activation in
Helicobacter pylori-induced apoptosis of gastric epithelial cells.
Scand J Gastroenterol. 46:148–155. 2011. View Article : Google Scholar
|
|
54
|
Zhang WH, Wang X, Narayanan M, et al:
Fundamental role of the Rip2/caspase-1 pathway in hypoxia and
ischemia-induced neuronal cell death. Proc Natl Acad Sci USA.
100:16012–16017. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Liu W, Staecker H, Stupak H, Malgrange B,
Lefebvre P and Van De Water TR: Caspase inhibitors prevent
cisplatin-induced apoptosis of auditory sensory cells. Neuroreport.
9:2609–2614. 1998. View Article : Google Scholar : PubMed/NCBI
|