Introduction
Vestibular schwannomas (VS), also known as acoustic
neuromas, are the most frequent benign tumor of the lateral skull
base, and originate from Schwann cells of the vestibular branch of
the eighth cranial nerve. VSs are neoplasms that occur as a result
of the increased proliferation of Schwann cells and are diagnosed
histopathologically by the presence of singular architectural
patterns called Antoni A and Antoni B areas. VS represents 8% of
all primary intracranial tumors (1). Individuals between 30 and 60 years
old are the most frequently affected, and there is no gender
prevalence (2,3). The majority of VSs are sporadic and,
in general, are benign slow-growing neoplasms. They exhibit a wide
variability in growth rate and size. If growth continues it may
result in complex pathological conditions, including brainstem
compression and hydrocephalus. Magnetic resonance imaging has a
central role in the diagnosis of this condition, and complete
surgical resection remains the preferred treatment. The growth of
VS is not directly correlated with tumor size, symptoms, duration
of symptoms or the patient’s age (4). The growth rate of VS is
heterogeneous, at 0.3–1.42 mm/year. VS evolves from an abnormal
growth and proliferation of Schwann cells, at their junction with
glial cells surrounding the vestibular nerve. This neoplasm is
exclusively formed from the hyperproliferation of Schwann cells and
associated neovasculature (5,6).
Schwann cells are principal glial cells of the peripheral nervous
system. During embryogenesis, they migrate along axons and
synthesize a basal lamina, consisting predominantly of laminin,
collagen and proteoglycans. Schwann cells have an important role in
nerve regeneration following laceration, when they replace damaged
Schwann cells and synthesize a new basal lamina and myelin sheath.
Following nerve injury, successful remyelination of damaged axons
by Schwann cells relies upon a combination of signals that Schwann
cells receive from demyelinated axons during in the inflammatory
response. These signals initially prompt Schwann cells to re-enter
the cell cycle and subsequently to differentiate into myelinating
cells (6). Axonal myelination is
important for the functional recovery of injured peripheral nerves.
In particular, it facilitates rapid saltatory impulse conduction by
producing a faster conduction velocity of action potentials
(7,8). In order to better understand the
molecular mechanisms of Schwann cells, including cell
proliferation, migration, survival and apoptosis during peripheral
nerve injury, the present study aimed to invetigate the involvement
of certain inflammatory cytokines and growth factors in VS. The
majority of the pathogenetic mechanisms regulating neoplastic
growth in vestibular nerve cells, remain to be elucidated. A number
of studies have demonstrated that neurotrophins and growth factors
have a role in governing the development of homeostasis, cell
survival and regeneration processes within Schwann cells (9,10).
Although further neoplastic growth appears to depend on cytokines
with angiogenic and mitogenic properties, data concerning the
involvement of growth factors in VS growth are not currently
available. Trophic factors, including transforming growth factor-β1
(TGF-β1) and vascular endothelial growth factor (VEGF), have been
designated as possible key mediators of VS growth. TGFs are a
family of polypeptides involved in wound healing and tumorigenesis
in vivo. TGF-β1 may act as either an inhibitor or stimulator
of cell proliferation, depending on the cell type and growth
conditions. TGF-β1 may be involved in the development of VS,
stimulating the proliferation of Schwann cells. The signalling
pathway of TGF-β1 is activated by two transmembrane
serine/threonine kinases, TGF-βR1 and TGF-βR2. The type 2 receptor
(TGF-βR2) is involved in the antiproliferative activity of TGF-β,
whereas the type 1 receptor (TGF-βR1) appears to cause cellular
proliferation following cell-matrix interactions (11). Interactions between cytokines and
Schwann cells are involved in the development of disorders of the
peripheral nervous system. Tumor necrosis factor-α (TNF-α) is a
pro-inflammatory cytokines produced by activated macrophages in
response to pathogens and other noxious stimuli. TNF-α is released
by Schwann cells as well as by macrophages (12). TNF-α, one of the major initiators
of the inflammatory cascade, activates pleiotropic functions in
physiological and pathological conditions by binding to its
receptors, type I (TNFRI) and type II (TNFRII). Wang et al
(13) described the involvement of
TNF-α-associated signalling molecules, including a baculoviral
inhibitor of apoptosis repeat-containing protein (BIRC) 2, BIRC 3
and TNFRI, in the anti-apoptotic process of injured peripheral
nerves, indicating that a higher level of TNF-α may induce
apoptosis in Schwann cells in vitro, while a lower level of
TNF-α may not act in the same way. Wagner and Myers (12) confirmed that the production of
TNF-α by peripheral nerve glial cells has a pathogenic role in
nerve injury. A number of studies have investigated the possible
role of interleukin (IL)-6 in peripheral nerve regeneration
(14,15). However, the molecular mechanisms
underlying the involvement of IL-6 in the development of Schwann
cells remain to be fully elucidated. The induction of
pro-inflammatory genes by IL-6 in Schwann cells may indicate that
IL-6 is involved in the degeneration of injured neurons, in
cooperation with other inflammatory cytokines, such as TNF-α.
Certain pro-inflammatory cytokines, including TNF-α, IL-1β and IL-6
are known to induce the expression of adhesion molecules. Adhesion
molecule expression, including that of intracellular adhesion
molecule-1 (ICAM-1), is tightly-regulated by cytokines generated
during an inflammatory response (16). ICAM-1 is able to facilitate
leukocyte attachment and interactions with cells from the target
tissue (17). The expansion of any
solid tumor with a volume >2–3 mm is reliant upon angiogenesis
to provide oxygen and nutrients to the enlarging tumor. VEGF
induces angiogenesis through endothelial cell proliferation and
migration. It is considered to be one of the most potent
pro-angiogenic factors, causing vasodilatation, vascular
permeability and angiogenesis. Angiogenesis is defined as the
process of new blood vessel formation from pre-existing vasculature
and involves a cascade of processes, during which the vessel’s
basal membrane and the surrounding extracellular matrix are
modified by endothelial cell proliferation and migration (18). The binding of VEGF to high-affinity
receptors, VEGFR-1 and VEGFR-2, promotes extravasation of plasma
proteins from tumor vessels, thereby forming a temporary
extravascular matrix which favors the migration and proliferation
of endothelial cells, resulting in new blood vessel formation
(19). The aim of the present
study was to investigate the expression of pro-inflammatory
cytokines in VS compared with normal vestibular nerve tissue, using
immunohistochemistry, in order to improve understanding of the
pathogenesis of this disease. An increased knowledge of this
subject may have significant clinical consequences in terms of
improvement in clinical treatment, prevention of the postsurgical
relapse, and a reduction in the severity of inflammation, tissue
invasion, tumor proliferation and angiogenesis.
Materials and methods
Ethical considerations and VS
samples
In accordance with approval of the ethical committee
of Policlinico Umberto, Sapienza University of Rome (Rome, Italy),
tissues were harvested from ten randomly selected patients, four
females and six males, with unilateral, sporadic VS that had been
removed surgically, and ten healthy control samples, which
consisted of three females and seven males with Ménière’s syndrome
following vestibular neurectomy. The patients, aged between 45 and
69 years, consisted of four females and six males. Routine
histopathological examination confirmed the diagnosis of benign VS
in the samples, which included one patient with a tumor
recurrence.
Prior to signing the consent form, patients were
informed about the study in detail and were given sufficient time
to ask questions. The study was conducted in accordance with the
Declaration of Helsinki. Each clinical unit selected specimens and
assigned a number to each sample, followed by a letter indicating
the participating unit. For each case, a report was prepared,
indicating the age and gender of the patient, as well as their
clinical signs and symptoms. Control morphological sections were
stained with hematoxylin and eosin (H&E), or processed for
immunohistochemistry. The following molecules were investigated in
the tumoral samples and in normal vestibular nerve specimens: VEGF,
TGF-β1, IL-1β, IL-6, ICAM-1 and TNF-α.
Immunohistochemical analysis
For light microscopic immunohistochemical analysis,
small fragments of VS were processed according to the avidin-biotin
complex/horseradish peroxidase technique. These samples were washed
in phosphate-buffered saline (PBS), fixed in 10% formalin and
embedded in paraffin according to a standard procedure (20). Serial 3-μm sections were cut
using a rotative microtome (RM2265; Leica Biosystems, Wetzlar,
Germany), mounted on gelatin-coated slides and processed for
immunohistochemistry. These sections were deparaffinized in xylene
and dehydrated. They were immersed in citrate buffer (pH 6.0;
15M103; BioOptica Milano, S.p.A, Milan, Italy) and subjected to
microwave irradiation twice for 5 min. Subsequently, all sections
were treated for 30 min with 0.3% hydrogen peroxide in methanol in
order to quench endogenous peroxidase activity. To block
non-specific binding, the slides were incubated in 3% normal goat
serum (S-100; Vector Laboratories Burlingame, CA, USA) in PBS
(15M108; BioOptica Milano, S.p.A.) for 30 min at room temperature.
The slides were incubated overnight at 4°C with the following
antibodies all purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA): Rabbit anti-IL-1β polyclonal antibody (1:50;
sc-7884); rabbit anti-IL-6 polyclonal antibody (1:200; sc-7920);
mouse anti-TNF-α monoclonal antibody (1:100; sc-52791); mouse
anti-VEGF monoclonal antibody (1:200; sc-152); mouse anti-ICAM-1
monoclonal antibody (1:50; sc-107); and rabbit anti-TGF-β1
polyclonal antibody (1:200; sc-146). Optimal antisera dilutions and
incubation times were assessed in a series of preliminary
experiments. Following exposure to the primary antibodies, slides
were rinsed twice in PBS and incubated for 1 h at room temperature
with the appropriate secondary biotinylated goat anti-mouse or
anti-rabbit immunoglobulin G (1:200; cat. nos. BA9200 and BA1000;
Vector Laboratories) and with peroxidase-conjugated avidin
(Vectastain Elite ABC kit standard* PK 6-100; Vector Laboratories)
for 35 min. Following a further wash with PBS, slides were treated
with 0,05% 3,3-diaminobenzidine (DAB) and 0,1%
H2O2 (DAB substrate kit for peroxidase,
Vector Laboratories; SK-4100). Finally, sections were
counter-stained with Mayer’s hematoxylin and observed using a light
microscope (Axio Lab.A1; Zeiss, Oberkochen, Germany). Negative
control experiments were performed by omitting the primary
antibody, substituting the primary antibody with an equivalent
quantity of non-specific immunoglobulins or pre-incubating the
primary antibody with the specific blocking peptide
(antigen/antibody=5 according to the manufacturer’s instructions).
The staining assessment was made by two observers experienced in
light microscopy. Immunoreactivity was assessed for IL-1β, IL-6,
TNF-α, VEGF, ICAM-1 and TGF-β1 in Schwann cells, vascular
endothelium and dense connective tissue of neoplastic vestibular
nerve samples, and compared with that of the healthy samples. The
intensity of the immune reaction was assessed
microdensitometrically using an IAS 2000 image analyzer (Delta
Sistemi, Rome, Italy) connected via a TV camera to the microscope.
The system was calibrated using zero as the background obtained in
sections exposed to non-immune serum. A total of ten
100-μm2 areas were delineated in each section
using a measuring diaphragm. The quantitative data regarding the
intensity of immune staining were analyzed statistically using an
analysis of variance followed by Duncan’s multiple range test as a
post hoc test.
Statistical analysis
The comparison of the expression levels of TNF-α,
TGF-β1, IL-1β, IL-6 and VEGF between the VS and normal vestibular
nerve samples was performed using a t-test. Statistical analyses
were performed using the SPSS statistical software package version
12.0 (SPSS, Inc., Chicago, IL, USA). P<0.001 was considered to
indicate a statistically significant difference.
Results
Morphological analysis of vestibular
nerves using H&E staining
A total of 10 patients, ranging in age between 45
and 69 years, with a diagnosis of VS were investigated. A total of
10 samples of healthy vestibular nerves were obtained via
vestibular neurectomy to serve as controls. The control peripheral
nerve samples consisted of a single fascicle surrounded by a dense
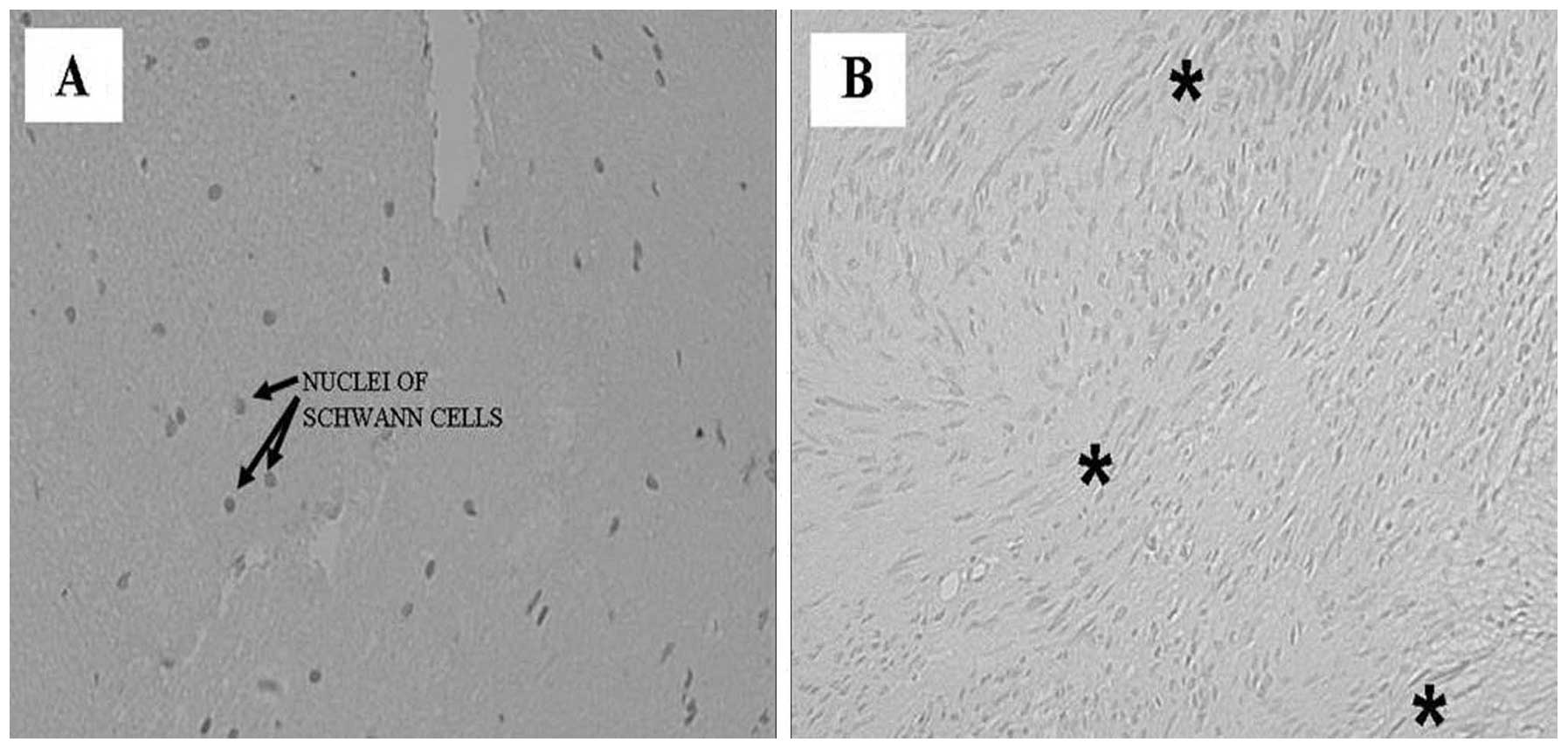
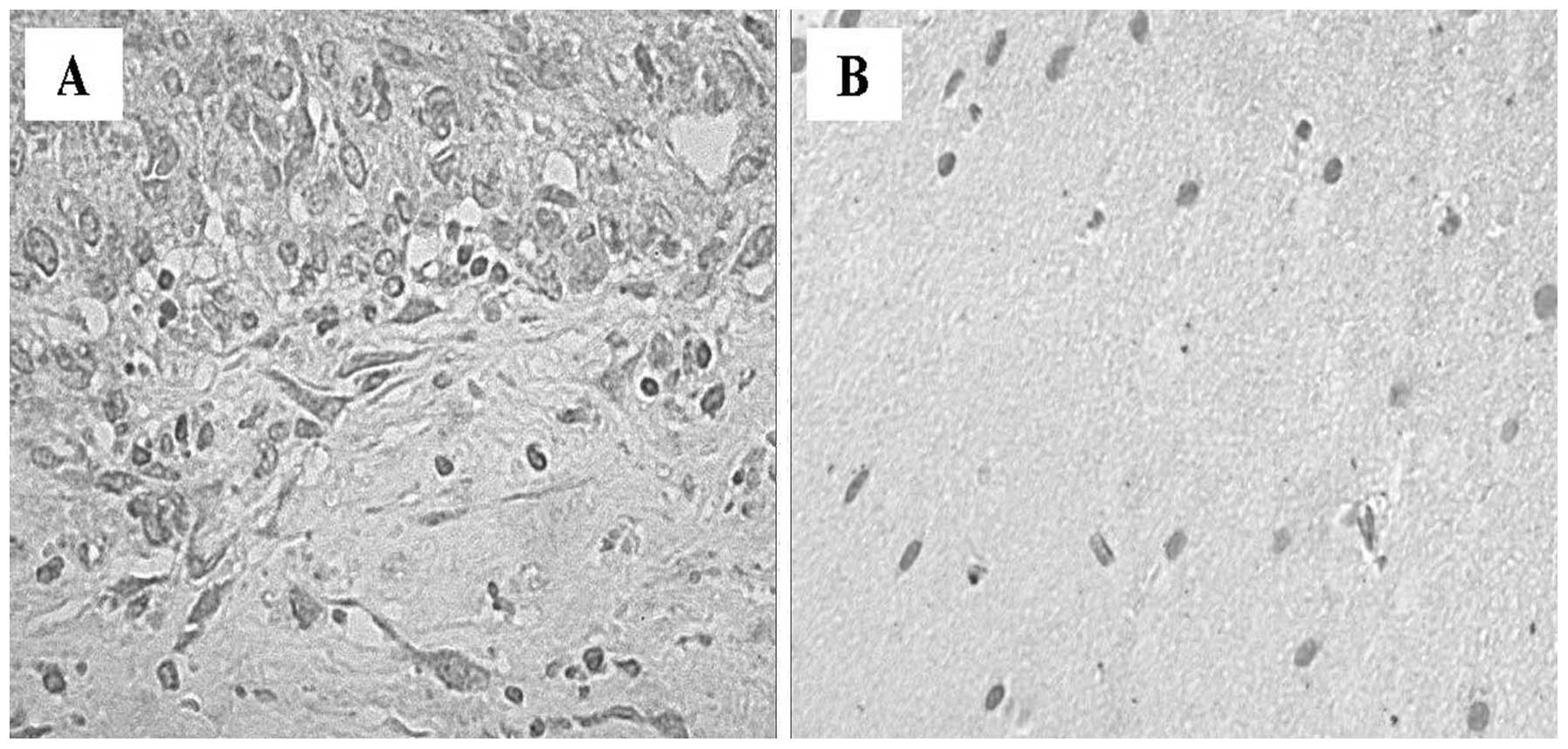
perineurium, containing small vessels (Fig. 1). The greatest number of nuclei
within the fascicle were attributed to Schwann cells. The shape and
the arrangement of these nuclei reflected the course of individual
axons. The fibroblasts of the endoneurium were dispersed amongst
numerous Schwann cells, and exhibited thinner nuclei and increased
cellular condensation compared with the Schwann cells. The nuclei
of Schwann cells were elongated along the major axis of the nerve
(Fig. 1A). Sections of VS samples
exhibited compact spindle cell areas, which were densely populated,
and formed a fascicular, storiform and whorled-growth pattern.
Histologically, schwannomas are composed of spindle cells arranged
in bundles with elongated nuclei that form Verocay’s bodies
(Fig. 1B). Phenomena associated
with cystic degeneration are common. Thickening and hyalinization
of the vessel walls are associated with microhemorrhagic phenomena.
The sections were exposed to primary and secondary antibodies,
resulting in the development of dark-brown (intense), yellow-brown
(slight) or no immune staining. Immunoreactivity was deemed to be
specific as no immunostaining was observed in control sections
incubated with primary antibodies absorbed with the specific
peptide or with the pre-immune serum.
Immunohistochemical analysis of human
healthy vestibular nerve and schwannoma samples
All VS samples examined in the present study
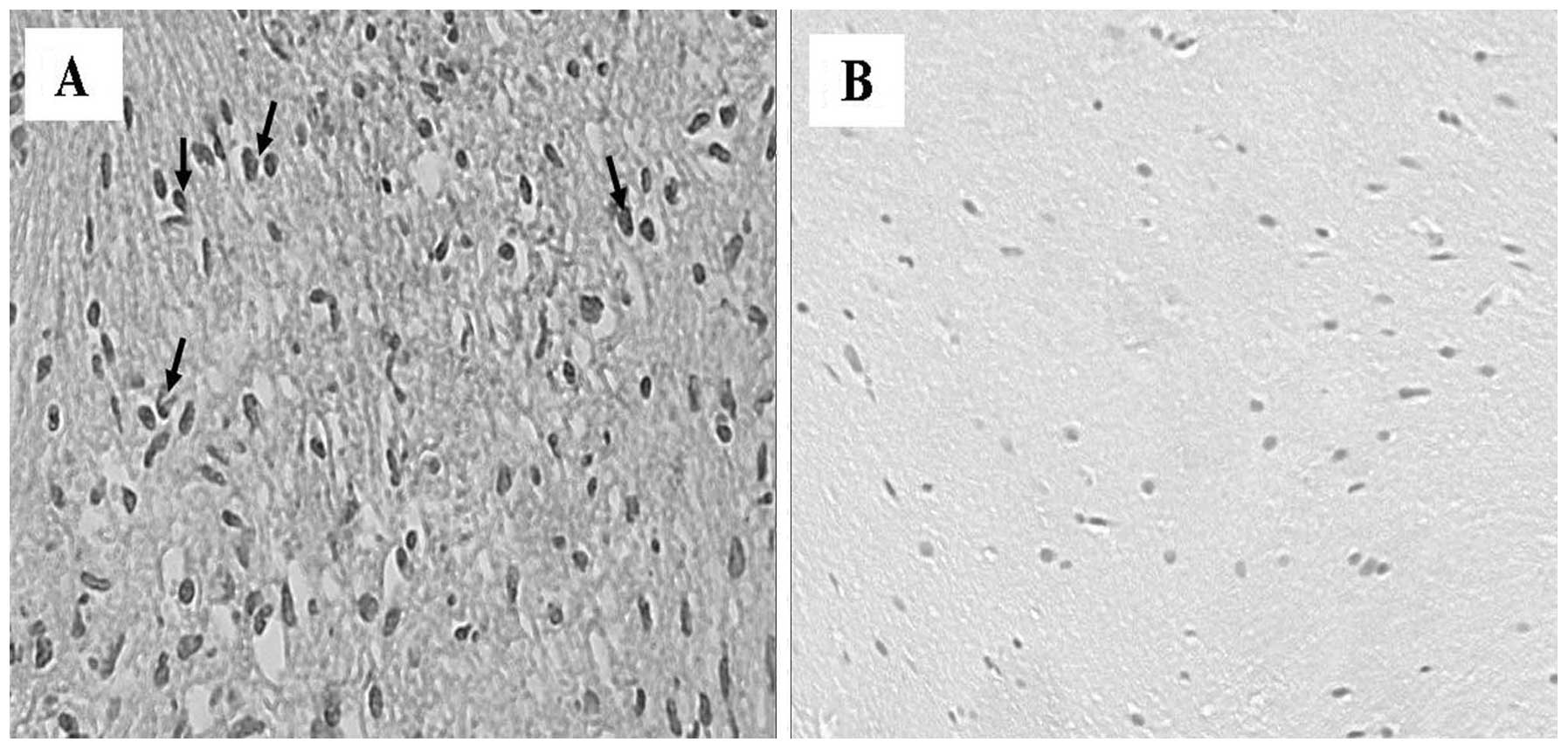
exhibited a marked immunoreactivity for TGF-β1. TGF-β1
immunoreactivity was detected primarily in the cytoplasm of Schwann
cells and revealed differences in the number of immunopositive
cells between Antoni A and Antoni B tissue types. Antoni A and
Antoni B tissue types represent distinct histologic architectural
patterns that aid in the histopathologic diagnosis of schwannoma
(21). Type A tissue is highly
cellular and demonstrates nuclear palisading as well as associated
Verocay bodies, which reflects their prominent extracellular matrix
and secretion of laminin. Type B tissue is loosely organized with
myxomatous and cystic change and may represent degenerated Antoni A
tissue. Antoni A cellular areas expressed more prominent TGF-β1
immunoreactivity than Antoni B areas. The Antoni B regions
exhibited less dense cellular areas than Antoni A regions, in which
there were compactly arranged spindle cells with long and oval
nuclei (Fig. 2A). TGF-β1
reactivity was also demonstrated in the blood vessel walls
distributed in the neoplastic dense connective tissue. No
immunoreactivity for TGF-β1 was observed in vascular endothelial
cells, or Antoni A and Antoni B areas in the control specimens
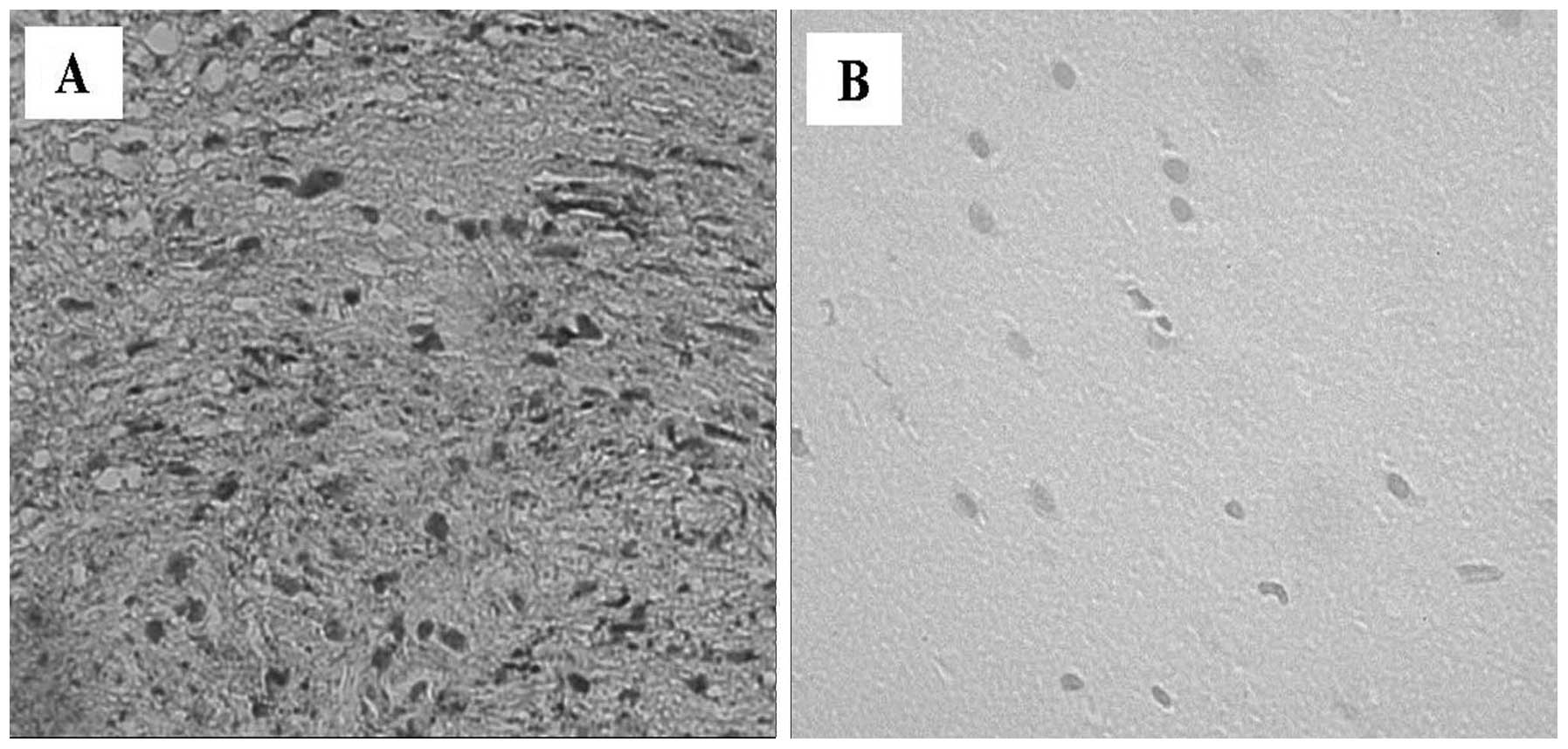
(Fig. 2B). TNF-α was weakly
expressed by the majority of human VS specimens in the endothelial
cells of vessels, and Antoni A and Antoni B regions. Numerous
immunopositive nuclei were detected in the Antoni A regions
compared with the Antoni B regions (Fig. 3A). No immunoreactivity for TNF-α
was detected in the healthy control vestibular nerve samples
(Fig. 3B). There was increased
expression of IL-1β within the VS tissues (Fig. 4A), which was localized to the
cytoplasm of Schwann cells. Antoni A and B regions exhibited
approximately the same number of immunopositive nuclei for IL-1β. A
moderate expression for IL-1β was also observed in the blood
vessels. No staining was observed in control tissues (Fig. 4B). Immunohistochemistry for IL-6
was positive in the VS cells, with weak expression in the cytoplasm
(Fig. 5A), while in the control
nerves samples it was undetectable (Fig. 5B). The findings confirmed that
these cytokines are involved in the development and progression of
VS via stimulation of Schwann cell proliferation. Pro-inflammatory
cytokines, including TGF-β1, TNF-α, IL-1β and IL-6, may be secreted
by activated leukocytes, fibroblasts and Schwann cells. These
cytokines are known to induce the expression of adhesion molecules,
such us ICAM-1 and VCAM-1, and are able to facilitate leucocyte
attachment and interactions with cells from the trigger tissue, an
area of the neuron that contains a high membrane concentration of
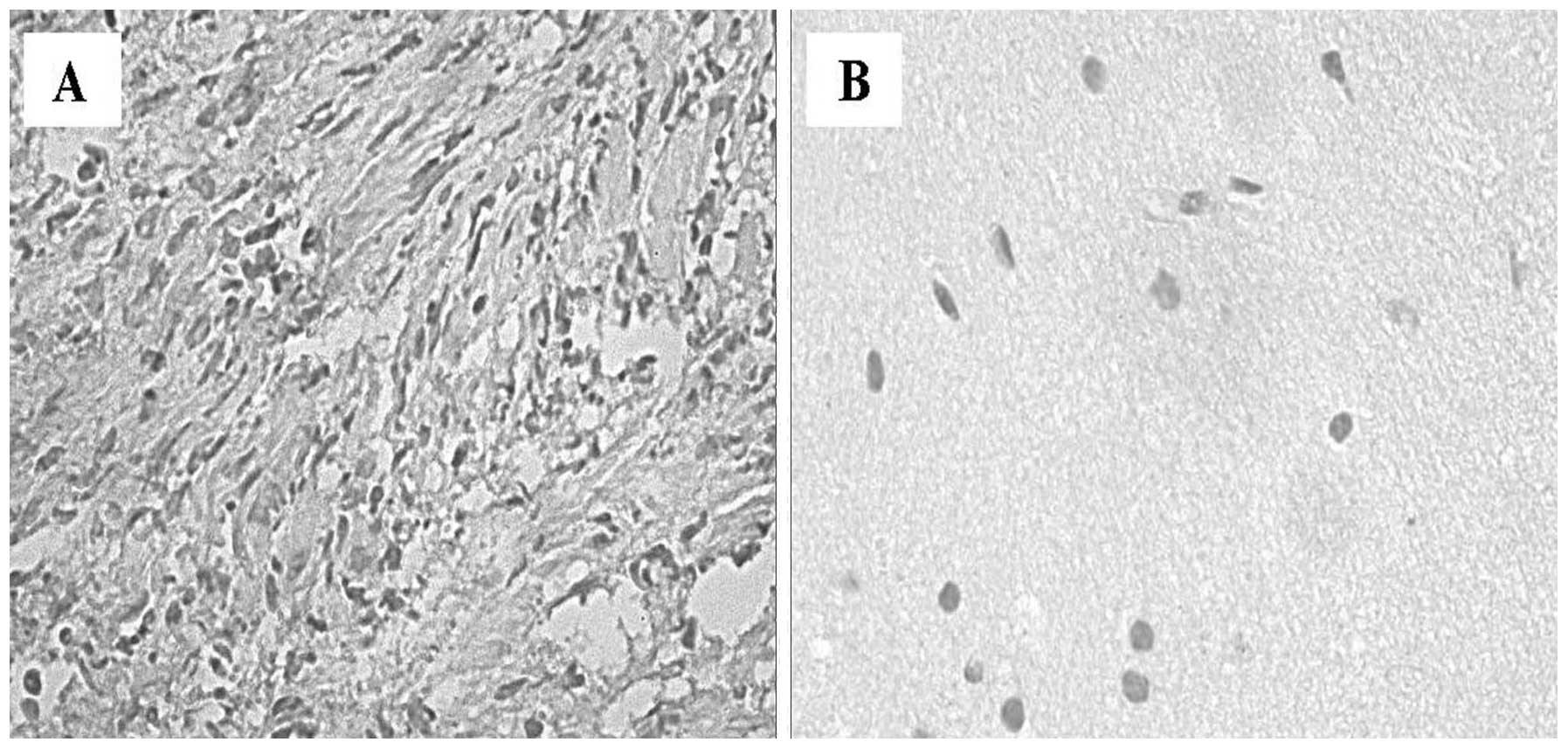
voltage-gated Na+ channels. A significantly higher ICAM-1
expression was observed in the cytoplasm of Schwann cells (Fig. 6A), compared with the control nerve
samples, in which no staining was identified (Fig. 6B).
Immunohistochemical staining for VEGF was positive
in 9/10 of the VS cases (Fig. 7A).
A positive expression of VEGF was observed in the tumor samples.
VEGF exhibited finely granular cytoplasmic staining in the Schwann
cells with intensified focal staining in the perinuclear region.
There was only a small variation of staining within the tissues of
individual tumors. Staining for VEGF also occurred in the cytoplasm
of endothelial cells and in polymorphonuclear leukocytes within
vessels. These data confirm that VEGF may have a significant impact
on the growth VSs, stimulating the mitogenic activity of Schwann
cells and angiogenesis in these tumors. VEGF immunoreactivity was
absent in the control nerve samples (Fig. 7B).
Statistical analysis of growth factors
and cytokines expression
The intensity of staining for TGF-β1, TNF-α, IL-1β,
IL-6 and VEGF in human VS and control nerve samples is shown in
Table I. The percentage values of
pro-inflammatory cytokine-positive cells and P-values are also
shown in Fig. 8.
 | Table ILevels of cytokines and growth
factors in vestibular schwannoma tissue samples and in healthy
vestibular nerve samples, and corresponding statistical
significance (t-test). |
Table I
Levels of cytokines and growth
factors in vestibular schwannoma tissue samples and in healthy
vestibular nerve samples, and corresponding statistical
significance (t-test).
| Factor | Vestibular
Schwannoma: 10 patients; 1 specimen for each patient (%) | Control samples: 10
patients; 1 specimen for each patient (%) | P-value |
|---|
| TGF-β1 | 84.3±4.83 | 2.0±0.66 | <0.0001 |
| TNF-α | 72.4±5.14 | 5.7±1.33 | <0.0001 |
| IL-1β | 74.1±6.11 | 6.0±1.24 | <0.0001 |
| IL-6 | 63.3±4.52 | 2.1±0.73 | <0.0001 |
| ICAM-1 | 90.4±4.97 | 6.1±1.19 | <0.0001 |
| VEGF | 77.5±3.30 | 7.2±1.03 | <0.0001 |
Discussion
VSs are rare and slow-growing neoplasms, which occur
as a result of increased proliferation of Schwann cells of the
vestibular branch of the eighth cranial nerve. The tumors generally
originate near the myelin-glial junction, close to the internal
auditory canal (22). Neoplastic
growth appears to rely upon cytokines, which possess angiogenic and
mitogenic properties. Limited data concerning the expression of
growth factors and its implication on VS growth are available. The
aim of the present study was to investigate the role of certain
pro-inflammatory cytokines in sporadic VS, associated with
angiogenesis and tumor growth. The mechanisms underlying schwannoma
development, growth and growth arrest remain to be elucidated. In
order to develop an improved understanding of the mechanisms
responsible for the growth of these neoplasms, associations between
an abnormal proliferation of Schwann cells and the expression of
certain inflammatory cytokines were investigated.
Although VSs are relatively slow-growing neoplasms,
their continued growth depends on a functional vascular system, as
with any other tumor (23). The
positive expression of VEGF in VS specimens suggests that
angiogenesis is involved in facilitating the growth of this tumor.
Indeed, angiogenesis is a prerequisite for the proliferation and
progression of a number of neoplasms (23). Despite the evidence suggesting that
VS are generally slow-growing tumors, and therefore do not require
excessive vascularization, the presence of a functional vascular
system remains paramount for tumor development. The results of the
present study have revealed a marked expression of VEGF in the
cytoplasm of Schwann cells and in vascular endothelial cells from
neoplastic peripheral nerves. The present findings confirm that
VEGF expression may be involved in the development and expansion of
benign tumors as well as malignant ones. In addition to its role as
an angiogenic factor, VEGF also possesses neurotrophic and
neuroprotective properties in the peripheral and central nervous
system, which exert a direct action not only on neurons, but also
on Schwann cells (24). The
mechanisms responsible for an inflammatory reaction in VS required
further elucidation. Inflammation contributes to tumor progression
by stimulating the angiogenic process and providing neoplastic
cells with growth factors.
VEGF and TGF-β1 have are putative key mediators of
VS growth (25). Overexpression of
TGF-β1 increases the invasiveness of neoplastic cells by increasing
their proteolytic activity and promoting their binding to
cell-adhesion molecules (26).
Previous animal studies have identified TGF-β1 as a potent mitogen
for Schwann cells. It has also previously been reported that
Schwann cells secrete and activate the latent form of TGF-β1
(27), thereby stimulating the
proliferation of Schwann cells. TGF-β1 has been hypothesized to be
involved in the regulation of peripheral nerve tumors by modulating
cell proliferation and differentiation, by different mechanisms
from those of glial growth factors and fibroblast growth factor,
which are responsible for the mitogenic activity of Schwann cells
(28,29). The results of the present study
suggested that TGF-β1 may be affect tumor progression by indirectly
stimulating angiogenesis through the upregulation of VEGF
expression in VS. Cytokines are the primary mediators of
communication between cells in the inflammatory tumor
microenvironment. It has been established that neoplastic cells
express pro-inflammatory mediators, including cytokines, such as
TGF-β1, IL-6, IL-1β and TNF-α (30). TNF-α is known to be a major
mediators of inflammation; in addition, TNF-α was reported to be
produced by tumors and to function as an endogenous tumor promoter.
TNF-α has been associated with numerous processes involved in
tumorigenesis, including cellular transformation, promotion,
survival, proliferation, invasion, angiogenesis and metastasis
(31). TNF-α upregulates ICAM-1 on
the Schwann cell surface, suggesting that these cells also carry
functional TNF-α receptors (32).
While TNF-α is toxic to numerous types of cell, it is not injurious
to cultured Schwann cells (33).
However, it does inhibit unstimulated cell proliferation and
connexin 46 expression (34). In
the present study, it was identified that moderate expression of
TNF-α in the cytoplasm of Schwann cells was predominantly
localization in Antoni A regions, while this cytokine was
completely absent in normal vestibular nerve samples. The cytotoxic
or protective effects of TNF-α depend on its receptor, cell type
and the presence of other factors. To date, two types of cell
surface receptors for TNF-α have been identified: TNFRI (p55) and
TNFRII (p75) (35). Activation of
TNFRII receptor results in a complex signalling pathway involving
numerous other TNF-receptor-activated proteins (36,37).
Receptor signaling, via this pathway, triggers the proteolysis of
cytoplasmic protein IκB, which, in turn, allows translocation of
the nuclear transcription factor nuclear factor-κB (38). This has been observed to lead to
apoptosis in specific cell types, whilst it is protective in others
(39). In the present study, TNF-α
appeared to induce a protective effect in Schwann cells, possibly
as a result of the presence on the cellular surface of TNFRII.
IL-1β, secreted by neoplastic cells or infiltrating leukocytes, is
involved in increasing tumor adhesion, invasion, angiogenesis and
immune suppression (40). The
results of the present study demonstrated that VS is associated
with elevated expression of IL-6 and VEGF, indicating that IL-6 is
a possible mediator of the association between VS and systemic
inflammatory responses in patients with this disease. Interleukin-6
is involved in peripheral nerve regeneration (41). However, the molecular mechanisms
underlying IL-6 function in Schwann cell physiology are yet to be
elucidated. The induction of proinflammatory genes by IL-6 in
Schwann cells may indicate that IL-6 is involved in the
degeneration of the injured nerve, in association with other
inflammatory cytokines, including TNF-α. IL-6 may facilitate the
demyelination of peripheral nerves following nerve injury, and
appears to induce degenerative changes in Schwann cells following
nerve injury and to activate proinflammatory signals in Schwann
cells (42). The majority of IL-6
target genes are involved in cell cycle progression and in the
suppression of apoptosis, which emphasizes the importance of IL-6
in tumorigenesis (43).
Accordingly, cytokines, including TNF-α and IL-1β, are emerging as
putative targets for anticancer therapies (44). Specific inhibition of
pro-inflammatory mediators, including TNF-α, IL-6, TGF-β1 and
IL-1β, may lead to a reduction in tumor development and inhibition
of transcription associated with the inflammatory process.
TNF-α, IL-6, TGF-β1 and IL-1β have been shown to
upregulate adhesion molecules, such as ICAM-1, in human Schwann
cells. In the present study, ICAM-1 was observed to be highly
expressed in VS samples and its expression was associated with
tumor size and the inflammatory process. Cellular immunity against
tumor cells requires the presence of adhesion molecules, such as
ICAM-1 on the endothelial surface, which mediate the arrest of
leukocytes (16). The induction of
the expression of ICAM-1 in Schwann cells by pro-inflammatory
cytokines suggests a possible role for adhesion molecules in the
pathogenesis of inflammation in the peripheral nerve. The absence
of immunoreactivity observed in the control specimens indicates
that the hyperexpression of growth factors and cytokines is
associated with tumor development or inflammatory conditions.
The current findings suggested that chronic
inflammation, through its promotion of angiogenesis, is involved in
tumor progression. Angiogenesis appears to be important for the
induction of growth of VS as well as the growth of other neoplasms,
including glioblastoma in children (45) and primary or metastatic breast
cancer (46). Numerous previous
studies have reported a significant correlation between the
concentration of VEGF and VEGFR-1 expression in VS, and tumor
growth rate, but did not describe symptom duration or tumor size
(47–49). The present findings confirmed the
expression of VEGF, with cytoplasmic localization, in VS samples.
Previous experiments in nude mice injected with malignant tumor
cells have demonstrated that intravenous infusion of anti-VEGF
monoclonal antibodies reduces the growth of tumors by up to 96%
(50). The anti-VEGF therapy was
directed toward the suppression of VEGF or its receptors. This
treatment, based on the inhibition of VEGF and its receptors, may
be a potential option with which to counteract the development of
VS.
Additional studies on the involvement of chronic
inflammatory processes in the development of VS are required. The
first stage may be to further identify the inflammatory cells
present in VS. Furthermore, their activation and association with
angiogenic growth factors should be examined. VS cells produce and
secrete pro-inflammatory cytokines, which may act in an autocrine
manner, stimulating cellular proliferation.
The potential use of novel therapeutic approaches
based on the combined administration of inhibitors of
proinflammatory cytokines and VEGF may hold promise for the
development of therapies for neoplastic diseases involving the
peripheral nervous system. In conclusion, the present study
demonstrated that the development of synthetic inhibitors of growth
factors may potentially reduce the recurrence rate of VS and enable
non-surgical management of this disease. Additional clinical and
experimental investigations are necessary to clarify the biological
role of these molecules in the development and progression of this
type of neoplasm.
References
|
1
|
McDonald R: Acoustic neuroma: what the
evidence says about evaluation and treatment. J Fam Pract.
60:E1–E4. 2011.PubMed/NCBI
|
|
2
|
Celis-Aguilar E, Lassalletta L, Torres
Martin M, Rodrigues FY, Nistal M, Castresana JS, et al: The
molecular biology of vestibular schwannomas and its association
with hearing loss: a review. Genet Res Int.
2012:8561572012.PubMed/NCBI
|
|
3
|
Bondi S, Limardo P, Toma S and Bussi M:
Non-vestibular head and neck schwannomas: a 10-year experience. Eur
Arch Otorhinolaryngol. 270:2365–2369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
van Leeuwen JP, Cremers CW, Thewissen NP,
Harhangi BS and Meijer E: Acoustic neuroma: correlation among tumor
size, symptoms, and patient age. Laryngoscope. 105:701–707. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carroll SL: Molecular mechanisms promoting
the pathogenesis of Schwann cell neoplasms. Acta Neuropathol.
123:321–348. 2012. View Article : Google Scholar :
|
|
6
|
Klenke C, Widera D, Sepehrnia A, Moffat
DA, Kaltschmidt C, Kaltschmidt B, Ebmeyer J and Sudhoff H: Clinical
and biological behaviour of vestibular schwannomas: signalling
cascades involved in vestibular Schwannoma resemble molecular and
cellular mechanisms of injury-induced Schwann cell
dedifferentiation. Head Neck Oncol. 16:202013.
|
|
7
|
Mirsky R, Parmantier E, McMahon AP and
Jessen KR: Schwann cell-derived desert Hedgehog signals nerve
sheath formation. Ann NY Acad Sci. 883:196–202. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen ZL and Strickland S: Laminin gamma1
is critical for Schwann cell differentiation, axon myelination, and
regeneration in the peripheral nerve. J Cell Biol. 163:889–899.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Weerda H, Gamberger TI, Siegner A, Gjuric
M and Tamm ER: Effects of transforming growth factor-β1 and basic
fibroblast growth factor on proliferation of cell cultures derived
from human vestibular nerve schwannoma. Acta Otolaryngol.
118:337–343. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Diensthuber M, Brandis A, Lenarz T and
Stover T: Co-expression of transforming growth factor-β1 and glial
cell line-derived neurotrophic factor in vestibular schwannoma.
Otol Neurotol. 25:359–365. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Löttrich M, Mawrin C, Chamaon K, Kirches
E, Dietzmann K and Freigang B: Expression of transforming growth
factor-beta receptor type 1 and type 2 in human sporadic vestibular
Schwannoma. Pathol Res Pract. 203:245–249. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wagner R and Myers RR: Schwann cells
produce tumor necrosis factor alpha: expression in injured and
non-injured nerves. Neuroscience. 73:625–629. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Y, Tang X, Yu B, Gu Y, Yuan Y, Yao D,
Ding F and Gu X: Gene network revealed involvements of Birc2, Birc3
and Tnfrsf1a in anti-apoptosis of injured peripheral nerves. PloS
One. 7:e434362012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee HK, Seo IA, Suh DJ, Hong JI, Yoo YH
and Park HT: Interleukin-6 is required for the early induction of
glial fibrillary acidic protein in Schwann cells during Wallerian
degeneration. J Neurochem. 108:776–786. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bolin LM, Verity AN, Silver JE, Shooter EM
and Abrams JS: Interleukin-6 production by Schwann cells and
induction in sciatic nerve injury. J Neurochem. 64:850–858. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Constantin G, Piccio L, Bussini S, Pizzuti
A, Scarpini E, Baron P, Conti G, Pizzul S and Scarlato G: Induction
of adhesion molecules on human Schwann cells by proinflammatory
cytokines, an immunofluorescence study. J Neurol Sci. 170:124–130.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bevilacqua MP: Endothelial-leukocyte
adhesion molecules. Annu Rev Immunol. 11:767–804. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Møller MN, Werther K, Nalla A, Stangerup
SE, Thomsen J, Bøg-Hansen TC, Nielsen HJ and Cayé-Thomasen P:
Angiogenesis in vestibular schwannomas: expression of extracellular
matrix factors MMP-2, MMP-9 and TIMP-1. Laryngoscope. 120:657–662.
2010. View Article : Google Scholar
|
|
19
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bancroft JD and Gamble M: Theory and
Practice of Histological Techniques. 6th. Churchill Livingstone;
Elsevier, London: 2008
|
|
21
|
Joshi R: Learning from eponyms: Jose
Verocay and Verocay bodies, Antoni A and B areas, Nils Antoni and
Schwannomas. Indian Dermatol Online J. 3:215–219. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Charabi S: Acoustic neuroma/vestibular
schwannoma in vivo and in vitro growth models. A clinical and
experimental study. Acta Otolaryngol Suppl. 530:1–27.
1997.PubMed/NCBI
|
|
23
|
Folkman J: Tumor angiogenesis: therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Namiecińska M, Marciniak K and Nowak JZ:
VEGF as an angiogenic neurotrophic, and neuroprotective factor.
Postepy Hig Med Dosw (Online). 59:573–583. 2005.In Polish.
|
|
25
|
Kramer F, Stöver T, Wamecke A, Diensthuber
M, Lenarz T and Wissel K: BDNF mRNA expression is significantly
upregulated in vestibular schwannomas and correlates with
proliferative activity. J Neurooncol. 98:31–39. 2010. View Article : Google Scholar
|
|
26
|
Maehara Y, Kakeji Y, Kabashima A, Emi Y,
Watanabe A, Alkazawa K, Baba H, Kohnoe S and Sugimachi K: Role of
transforming growth factor-beta 1 in invasion and metastasis in
gastric carcinoma. J Clin Oncol. 17:607–614. 1999.PubMed/NCBI
|
|
27
|
Ridley AJ, Davis JB, Stroobant P and Land
H: Transforming growth factors-beta 1 and beta 2 are mitogens for
rat Schwann cells. J Cell Biol. 109:3419–3424. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lemke GE and Brockes JP: Identification
and purification of glial growth factor. J Neurosci. 4:74–83.
1984.
|
|
29
|
Ratner N, Bunge RP and Glaser L: Schwann
cell proliferation in vitro. An overview. Ann NY Acad Sci.
486:170–181. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Candido J and Hagemann T: Cancer-related
inflammation. J Clin Immunol. 33:S79–S84. 2013. View Article : Google Scholar
|
|
31
|
Sethi G, Sung B and Aggaewal BB: TNF: a
master switch for inflammation to cancer. Front Biosci.
13:5094–5107. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lisak RP and Bealmear B: Upregulation of
intercellular adhesion molecule-1 (ICAM-1) on rat Schwann cells in
vitro: comparison of interferon-gamma, tumor necrosis factor-alpha
and interleukin-1. J Peripher Nerv Syst. 2:233–243. 1997.PubMed/NCBI
|
|
33
|
Mithen F, Colburn S and Birchem R: Human
alpha tumor necrosis factor does not damage cultures containing rat
Schwann cells and sensory neurons. Neurosci Res. 9:59–63. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chandross KJ, Spray DC, Cohen RI, Kumar
NM, Kremer M, Dermietzel R and Kessler JA: TNF-alpha inhibits
Schwann cell proliferation, connexin46 expression and gap
junctional communication. Mol Cell Neurosci. 7:479–500. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Uren AG and Vaux DL: Molecular and
clinical aspects of apoptosis. Pharmacol Ther. 72:37–50. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nagano K, Alles N, Mian AH, et al: The
tumor necrosis factor type 2 receptor plays a protective role in
tumor necrosis factor-α-induced bone resorption lacunae on mouse
calvariae. J Bone Miner Metab. 29:671–681. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tracey D, Klareskog L, Sasso EH, Salfeld
JG and Tak PP: Tumor necrosis factor antagonist mechanism of
action: a comprehensive review. Pharmacol Ther. 117:244–279. 2008.
View Article : Google Scholar
|
|
38
|
Baker SJ and Reddy EP: Transducers of life
and death: TNF receptor superfamily and associated proteins.
Oncogene. 12:1–9. 1996.PubMed/NCBI
|
|
39
|
Beg AA and Baltimore D: An essential role
for NF-kappaB in preventing TNF-alpha-induced cell death. Science.
274:782–784. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Colotta F, Allavena P, Sica A, Garlanda C
and Mantovani A: Cancer-related inflammation, the seventh hallmark
of cancer: links to genetic instability. Carcinogenesis.
30:1073–1081. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhong J, Dietzel ID, Wahle P, Kopf M and
Heumann R: Sensory impairments and delayed regeneration of sensory
axons in interleukin-6-deficient mice. J Neurosci. 19:4305–4313.
1999.PubMed/NCBI
|
|
42
|
Lee HK, Wang L, Shin YK, Lee KY, Suh DJ
and Park HT: Interleukin-6 induces proinflammatory signaling in
Schwann cells: a high-throughput analysis. Biochem Biophy Res
Commun. 382:410–414. 2009. View Article : Google Scholar
|
|
43
|
Haura EB, Turkson J and Jove R: Mechanisms
of disease: insights into the emerging role of signal transducers
and activators of transcription in cancer. Nat Clin Pract Oncol.
2:315–324. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Klampfer L: Cytokines, inflammation and
colon cancer. Curr Cancer Drug Targets. 11:451–464. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Artico M, Cervoni L, Celli P, Salvati M
and Palma L: Supratentorial glioblastoma in children a series of 27
surgically treated cases. Childs Nerv Syst. 9:7–9. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gobbi G, Mirandola P, Micheloni C, et al:
Expression of HLA class I antigen and proteasome subunits LMP-2 and
LMP-10 in primary vs. metastatic breast carcinoma lesions. Int J
Oncol. 25:1625–1629. 2004.PubMed/NCBI
|
|
47
|
Cayé-Thomasen P, Werther K, Nalla A, et
al: VEGF and VEGF receptor-1 concentration in vestibular schwannoma
homogenates correlates to tumor growth rate. Otol Neurotol.
26:98–101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Brieger J, Bedavanija A, Lehr HA, Maurer J
and Mann WJ: Expression of angiogenic growth factors in acoustic
neurinoma. Acta Otolaryngol. 123:1040–1045. 2003. View Article : Google Scholar
|
|
49
|
Koutsimpelas D, Bjelopavlovic M, Yetis R,
et al: The VEGF/VEGFR axis in sporadic vestibular schwannoma
correlates with irradiation and disease recurrence. ORL J
Otorinilaryngol Relat Spec. 74:330–338. 2012. View Article : Google Scholar
|
|
50
|
Kim KJ, Li B, Winer J, et al: Inhibition
of vascular endothelial growth factor-induced angiogenesis
suppresses tumour growth in vivo. Nature. 362:841–844. 1993.
View Article : Google Scholar : PubMed/NCBI
|