Introduction
Hepatocellular carcinoma (HCC) is the fifth most
frequently diagnosed cancer in men, and the second leading cause of
cancer-associated mortality worldwide (1). In 2008, an estimated 748,300 new HCC
cases and 695,900 HCC fatalities occurred worldwide, half of which
were located in China (1,2). Although substantial progress has been
made in the chemotherapeutic treatment of HCC, the efficacy of
chemotherapeutic drugs is often hampered by various adverse side
effects. Therefore, natural products have become increasingly
important for the discovery of novel pharmaceutical treatment
strategies. Traditional Chinese herbology pioneers the use of
natural products in the biomedical field, and Chinese medicinal
herbs are increasingly being used for their antitumor effects.
Hypericum japonicum, also know as
Tianjihuang, is a Chinese medicinal herb which has been used for
centuries in the treatment of infectious hepatitis, bacterial
infection, and tumors (3–5). Hypericum japonicum has been
shown to exhibit significant anti-carcinogenic effects in clinical
settings, specifically with regards to tumors of the digestive
system, such as HCC. In addition, treatment with Hypericum
japonicum has been shown to increase patient survival and
quality of life, and decrease tumor size (6). Furthermore, previous studies
demonstrated that Hypericum japonicum exhibited
hepatoprotective effects on CCL4- and
acetaminophen-induced acute hepatotoxicity in rat models (3,7).
However, the mechanism underlying the hepatoprotective effects of
Hypericum japonicum on HCC remains to be elucidated.
The present study evaluated the in vivo
efficacy of ethyl acetate extract of Hypericum japonicum
(EAEHJ) against tumor growth in an H22 cell-bearing liver cancer
mouse model, and investigated the molecular mechanisms underlying
the anti-carcinogenic effects of EAEHJ in vitro. It is
hypothesized that the anti-carcinogenic effects of Hypericum
japonicum in vitro likely result from its ability to promote
apoptosis of cancer cells via the mitochondria-dependent
pathway.
Materials and methods
Materials and reagents
Dulbecco's modified Eagle's medium (DMEM),
trypsin-EDTA, and penicillin-streptomycin were purchased from
Thermo Fisher Scientific, Inc. (Waltham, MA, USA). Fetal bovine
serum (FBS), TRIzol® Reagent, JC-1, and Caspase-3 and -9
Colorimetric Protease Assay kits were purchased from Invitrogen
Life Technologies (Carlsbad, CA, USA). SuperScript II reverse
transcriptase was purchased from Thermo Fisher Scientific, Inc. The
B-cell lymphoma 2 (Bcl-2; #2872; 1:1,000), Bcl-2-associated X
protein (Bax; #2772; 1:1,000) and β-actin (#4967; 1:1,000) primary
antibodies, and the horseradish peroxidase (HRP)-conjugated
secondary antibodies (#7075; 1:2,000) were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). The fluorescein
isothiocyanate (FITC)-conjugated Annexin V Apoptosis Detection kit
was purchased from BD Biosciences (San Jose, CA, USA). All the
other reagents, unless otherwise stated, were obtained from
Sigma-Aldrich (St. Louis, MO, USA).
EAEHJ preparation
A total of 200 g Hypericum japonicum was
extracted from Hypericum japonicum Thunb. ex Murray herbs
(Fujian Tongchun Pharmaceutical Co., Ltd., Fuzhou, China) using 1.6
l 95% ethanol, according to the reflux method (8), prior to being filtered. The ethanol
was then evaporated using a RE-2000 Rotary Evaporator (Shanghai
Yarong Biochemical Instrument Factory, Shanghai, China). The
solution obtained following ethanol evaporation was concentrated in
a rotary evaporator at 50°C until 26 g Hypericum japonicum
ethanol extract powder was produced. A total of 20 g of
Hypericum japonicum ethanol extract powder was then
resuspended in 200 ml distilled water. Four organic solvents in
increasing order of polarity: Petroleum ether, chloroform, ethyl
acetate, and butan-1-ol were subsequently partitioned using a
separating funnel. The ethyl acetate extract was used for further
experimentation, and was dissolved in dimethyl sulfoxide (DMSO) in
order to form a 600 mg/ml EAEHJ stock solution, prior to being
stored at -20°C. The experimental concentrations of EAEHJ (0, 0.5,
0.75, 1.5 and 3 mg/ml) were obtained by diluting the stock solution
in culture medium supplemented with <0.5% DMSO.
Animals
Male Kunming mice (10-12 weeks of age, weighing
18-22 g) were obtained from Vital River Laboratory Animal
Technology Co., Ltd. (Beijing, China), and housed in a sterile
humidified environment (50-60%) at 22°C, with a 12 h light/dark
cycle. Food and water were provided ad libitum throughout
the experiment. All animal treatments were conducted in strict
accordance with international ethical guidelines, according to the
National Institute of Health Care and Use of Laboratory Animals
Guide (9), and all experiments
were approved by the Institutional Animal Care and Use Committee of
the Fujian University of Traditional Chinese Medicine (Fuzhou,
China). The H22 liver cancer cells were injected subcutaneously
into the mice, as previously described (10,11).
Briefly, the ascites H22 cells were harvested, diluted to a
concentration of 5.0×106/ml using sterilized normal
saline, and injected subcutaneously into the right axillary region
of the mice. The tumor volumes were subsequently calculated using
the following formula: Volume = π/6 × L × W2 (12,13).
When the tumors reached a size of 100 mm3, the animals
were randomly assigned into two groups (n=8), and intragastrically
administered either 60 mg/kg/d EAEHJ or saline for 10 days. At the
end of the experiment, the animals were anaesthetized with
intraperitoneal pelltobarbitalum natricum (300 mg/kg), and the
tumor tissue samples were removed and weighed. The tumor tissue
samples were subsequently fixed with 10% formaldehyde for 12 h, and
placed on paraffin-embedded tumor slides. The inhibitory effects of
EAEHJ on tumor growth were evaluated by measuring the tumor growth
inhibition rate. The tumor growth inhibition rates were calculated
as follows: Tumor inhibition rate = (1 - average tumor weights of
the treatment group/average tumor weight of control group) ×
100.
Cell culture and cell viability
assay
The H22 murine hepatoma and HepG2 human hepatoma
cell lines were obtained from the American Type Culture Collection
(Manassas, VA, USA). Both cell lines were cultured in DMEM
supplemented with 10% (v/v) FBS, 100 U/ml penicillin and 100
μg/ml streptomycin at 37°C in in an atmosphere containing 5%
CO2. The cells were cultured to 80-90% confluence. The
cells used throughout the present study were subjected to <20
cell passages. Cellular morphological changes were observed using a
phase-contrast microscope (IX70; Olympus Corporation, Tokyo,
Japan). The images were captured at a ×100 magnification, and cell
viability was assessed using an MTT colorimetric assay. Briefly,
the HepG2 human hepatoma cells were seeded into 96-well plates at a
density of 1.0×104 cells/well in 0.1 ml medium.
Following a period of 24 h, the cells were treated with various
concentrations (0, 0.5, 0.75, 1.5 and 3 mg/ml) of EAEHJ. Treatment
with 0.5% DMSO was included as a vehicle control. At the end of the
cellular treatment period, the medium was discarded, and 100
μl MTT solution [0.5 mg/ml in phosphate-buffered saline
(PBS)] was added to each well. Following 4 h of incubation at 37°C,
MTT was discarded and 100 μl DMSO was added to each well.
The purple/blue MTT formazan precipitate was dissolved, and the
absorbance was measured at 570 nm using an EL×800 ELISA reader
(BioTek Instruments, Inc., Winooski, VT, USA).
In situ apoptosis detection by TUNEL
staining
The 5 μm tumor tissue sample sections were
analyzed by TUNEL staining using a TumorTACS™ In Situ
Apoptosis Detection kit in accordance with the manufacturer's
instructions (R&D Systems, Inc., Minneapolis, MN, USA). The
apoptotic diaminobenzidine (DAB)-positive cells (brown stained)
were counted in five arbitrarily selected microscopic fields at a
×400 magnification (DM IL LED Microscope; Leica Microsystems GmbH,
Wetzlar, Germany).
Immunohistochemical analysis
The slides containing the tumor tissue samples were
subjected to antigen retrieval with citrate buffer, and the
endogenous peroxidase activity was blocked using 3% hydrogen
peroxide in water. For immunohistochemical staining, the slides
were incubated with rabbit polyclonal antibodies targeting Bax
(sc-6236) and Bcl-2 (sc-783) (1:200; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA). Following washing with PBS, the slides were
incubated with biotinylated HRP-conjugated streptavidin secondary
antibody (E0432; 1:1,000; Dako, Glostrup, Denmark), prior to being
further washed with PBS. The slides were then incubated with DAB,
prior to being counterstained with diluted Harris hematoxylin.
Following staining, five ×400 magnification fields (DM IL LED
Microscope) were randomly selected in each slide, and the average
ratio of positive cells in each field was counted using the
Image-Pro Plus true color multi-functional cell image analysis
management system, version 6.0 (Media Cybernetics, Inc., Rockville,
MD, USA). In order to prevent any non-specific staining, PBS was
used as a replacement to the primary antibody in the negative
control.
Annexin V-FITC/propidium iodine (PI)
staining apoptosis assay
Following incubation with various concentrations (0,
0.5, 0.75, 1.5 and 3 mg/ml) of EAEHJ, the apoptotic levels of the
HepG2 human hepatoma cells were determined by
fluorescence-activated cell sorting (FACS) flow cytometry
(FACSCalibur; BD Biosciences), using an Annexin V-FITC/PI kit (BD
Biosciences). Staining was performed according to the
manufacturer's instructions. The Annexin V/PI-double negative cells
were considered to be viable. The Annexin V-positive/PI-negative
population represents cells undergoing early apoptosis, while the
population of Annexin V/PI double-positive cells represents cells
in the late apoptotic stage.
Detection of ΔΨm by JC-1 staining
JC-1 is a cationic dye that exhibits
potential-dependent accumulation in mitochondria, indicated by a
shift in fluorescence emission from green to red. Therefore, JC-1
may be used as an indicator of mitochondrial potential. The HepG2
human hepatoma cells were plated in 6-well plates at
1×105 cells/well in a volume of 2 ml, and incubated in
DMEM overnight. The cells were then treated with various
concentrations (0, 0.5, 0.75, 1.5 and 3 mg/ml) of EAEHJ for 24 h,
prior to being cultured in fresh media in the absence of FBS for a
further 24 h. The cells were washed twice with PBS, and resuspended
in 1 ml DMEM supplemented with 10% FBS and 10 μg/ml JC-1, at
37°C in an atmosphere containing 5% CO2 for 30 min. Both
red and green fluorescence emissions were analyzed by flow
cytometry following JC-1 staining. The green and red fluorescence
intensities, measured respectively at 520-530 nm (PMT2) and 590 nm
(PMT3) for 1.0×104 individual cells, were analyzed by
FACS analysis (BD Biosciences). The ΔΨm values were expressed as
the ratio of PMT3 to PMT2.
Reverse transcriptase-polymerase chain
reaction (RT-PCR) and western blot analysis
A total of 2×105 HepG2 human hepatoma
cells were seeded into 6-well plates in 2 ml medium, and treated
with various concentrations (0, 0.5, 0.75, 1.5 and 3 mg/ml) of
EAEHJ for 24 h. Total RNA from HepG2 human hepatoma cells was
isolated using TRIzol® Reagent. A total of 1 μg
Oligo(dT)-primed RNA was reverse-transcribed using SuperScript II
reverse transcriptase, according to the manufacturer's
instructions. The obtained cDNA was used to determine the mRNA
expression levels of Bcl-2 or Bax using Taq DNA polymerase
(Thermo Fisher Scientific, Inc.). GAPDH was used as an internal
control. The sequences of the primers used for Bcl-2, Bax, and
GAPDH amplification were as follows: Bcl-2 forward, 5′-CAG CTG CAC
CTG ACG CCC TT-3′, and reverse, 5′-GCC TCC GTT ATC CTG GAT CC-3′;
Bax forward, 5′-TGC TTC AGG GTT TCA TCC AGG-3′, and reverse, 5′-TGG
CAA AGT AGA AAA GGG CGA-3′; and GAPDH forward, 5′-GT CAT CCA TGA
CAA CTT TGG-3′, and reverse, 5′-GA GCT TGA CAA AGT GGT CGT-3′.
Cycling was conducted using the S1000 Thermal Cycler (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) with the following cycling
conditions: Initial dena-turation at 95°C for 5 min, 30 sec rest,
30 cycles of annealing at a temperature 5°C below the melting point
of the primers and extension at 72°C both for ~1 min/kb of expected
product 10 min on the last cycle. PCR products were then analyzed
by electrophoresis on a 2% agarose gel. The DNA bands were examined
using a Gel Doc 2000 system (Bio-Rad Laboratories, Inc.). In order
to carry out the western blot analysis, the HepG2 human hepatoma
cells treated with EAEHJ were lysed using Mammalian Protein
Extraction Reagent cell lysis buffer (Thermo Fisher Scientific,
Inc.) containing protease (EMD Millipore, Billerica, MA, USA) and
phosphatase inhibitor cocktails. The cell lysates were separated by
12% SDS-PAGE prior to immunoblotting to polyvinylidene fluoride
(PVDF) membranes using the iBlot Western Detection Stack/iBlot Dry
Blotting system (Invitrogen Life Technologies). The PVDF membranes
were blocked using SuperBlock T20 Blocking Buffer (Thermo Fisher
Scientific, Inc.) for 30 min, and washed in tris-buffered saline
containing 0.25% Tween® 20 (TBST), prior to being
incubated with the primary antibodies overnight at 4°C. Following
washing with TBST, the membranes were incubated with the
HRP-conjugated secondary antibody for 1 h at room temperature.
Finally, the blots were developed using Super Signal Pico Substrate
(Thermo Fisher Scientific, Inc.), and images were captured using a
Kodak Image Station 400R (Kodak, Rochester, NY, USA).
Analysis of caspase activation
Quantitative measurement of caspase activity was
conducted using caspase-3 and -9 Colorimetric Protease Assay kits,
according to the manufacturer's instructions. Briefly, following
treatment with various concentrations (0, 0.5, 0.75, 1.5 and 3
mg/ml) of EAEHJ for 24 h, the HepG2 human hepatoma cells were lysed
with lysis buffer on ice for 30 min. The lysed cells were
centrifuged at 16,000 × g for 10 min, and 100 μg protein
samples were incubated with the following 50 μl colorimetric
tetrapeptides: Asp-Glu-Val-Asp (DEVD)-p-nitroaniline (pNA)
(specific substrate of caspase-3), or Leu-Glu-His-Asp (LEHD)-pNA
(specific substrate of caspase-9), in total darkness at 37°C, for 2
h. Sample absorbance was measured at 405 nm using an EL×800 ELISA
reader. The data were normalized to the activity levels of
caspase-3 and -9 in the control cells treated with 0.5% DMSO
vehicle, and represented as the “fold of control”.
Statistical analysis
The data are expressed as the mean ± standard
deviation and were analyzed with SPSS software, version 13.0 (SPSS,
Inc., Chicago, IL, USA). The statistical significance of each
experimental test condition was assessed using an unpaired
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Inhibitory effects of EAEHJ in H22
cell-bearing mice, and HepG2 human hepatoma cells
Following the sacrifice of the mice, the tumors were
harvested and weighed. The tumor inhibition rates are shown in
Table I. The mean tumor weight in
the EAEHJ treatment group was significantly lower than that of the
control group (P<0.05). The inhibitory effects of EAEHJ
treatment on tumor growth were evaluated from the tumor inhibition
rates, the results of which indicated that EAEHJ suppressed tumor
growth by 39.2% (Table I).
However, administration of EAEHJ had no apparent effect on the body
weight of the experimental animals, demonstrating that EAEHJ is
able to inhibit liver cancer growth in vivo in the absence
of apparent adverse effects. These results suggest that EAEHJ may
significantly inhibit tumor growth in H22 tumor cell-bearing mouse
model in vivo.
 | Table IInhibitory effects of Hypericum
japonicum ethyl acetate extract (EAEHJ) in H22 cell-bearing
liver cancer mice. |
Table I
Inhibitory effects of Hypericum
japonicum ethyl acetate extract (EAEHJ) in H22 cell-bearing
liver cancer mice.
| Group | Body weight (g)
| Tumor weight
(g) | Tumor inhibition
rate (%) |
|---|
| Pre-treatment | Post-treatment |
|---|
| Control | 20.6±0.99 | 27.4±1.29 | 0.378±0.156 | – |
| EAEHJ | 21.7±0.97 | 26.5±1.12 | 0.230±0.098a | 39.2 |
In order to examine the cytotoxicity and biological
effects of EAEHJ, HepG2 human hepatoma cells were treated with
various doses of EAEHJ (0, 0.5, 0.75, 1.5 and 3.0 mg/ml) for 24 h.
Any cellular morphological changes were subsequently evaluated by
phase-contrast microscopy. As shown in Fig. 1, the control cells exhibited a
typical polygonal intact shape, whereas the cells treated with
various concentrations of EAEHJ for 24 h became rounded, shrunken,
exhibited membrane blebbing, or became non-confluent. Furthermore,
the effects of the various concentrations of EAEHJ on HepG2 human
hepatoma cell viability were examined using an MTT assay. The
results were concordant with the microscopy results indicating that
EAEHJ inhibited cell growth in a dose-dependent manner. At a
concentration of 0.5 mg/ml, a significant loss of cell viability
was observed. The cell growth inhibition rate was ~50% at 0.75
mg/ml EAEHJ (P<0.01). Upon incubation with 3.0 mg/ml EAEHJ, the
cells were only 40% viable, (P<0.01, Fig. 1). These results indicate that EAEHJ
is able to inhibit the growth of HepG2 human hepatoma cells in
vitro.
Effects of EAEHJ on cell apoptosis in the
H22-bearing mice and HepG2 human hepatoma cells
In order to determine whether the inhibitory effects
of EAEHJ on tumor growth were due to apoptosis, the present study
examined the EAEHJ pro-apoptotic activities in tumor-bearing mice
via IHC TUNEL staining. The percentage of TUNEL-positive cells in
the control and EAEHJ-treated mouse groups was 31.5±11 and
57.3±15%, respectively (P<0.01, Fig. 2A and B). These results indicate
that EAEHJ may promote cell apoptosis in vivo.
The induction of cell apoptosis was evidenced by
phosphatidylserine externalization in vitro. Annexin V-/PI
staining was performed in order to examine the occurrence of
phosphatidylserine externalization on the cell surface. The number
of Annexin V-stained cells, indicating both early and late
apoptosis, increased with the concentration of EAEHJ treatment
(Fig. 2C). At a low concentration
of EAEHJ (0.5 mg/ml), only a small number of cells exhibited
early-phase and late-phase apoptosis (9.1%, P<0.01). However,
following incubation with higher concentrations of EAEHJ (3.0
mg/ml), the number of both early- and late-phase (necrotic stage)
apoptotic cells increased (75%, P<0.01). These results indicate
that cell treatment with EAEHJ may induce cell apoptosis in a
dose-dependent manner.
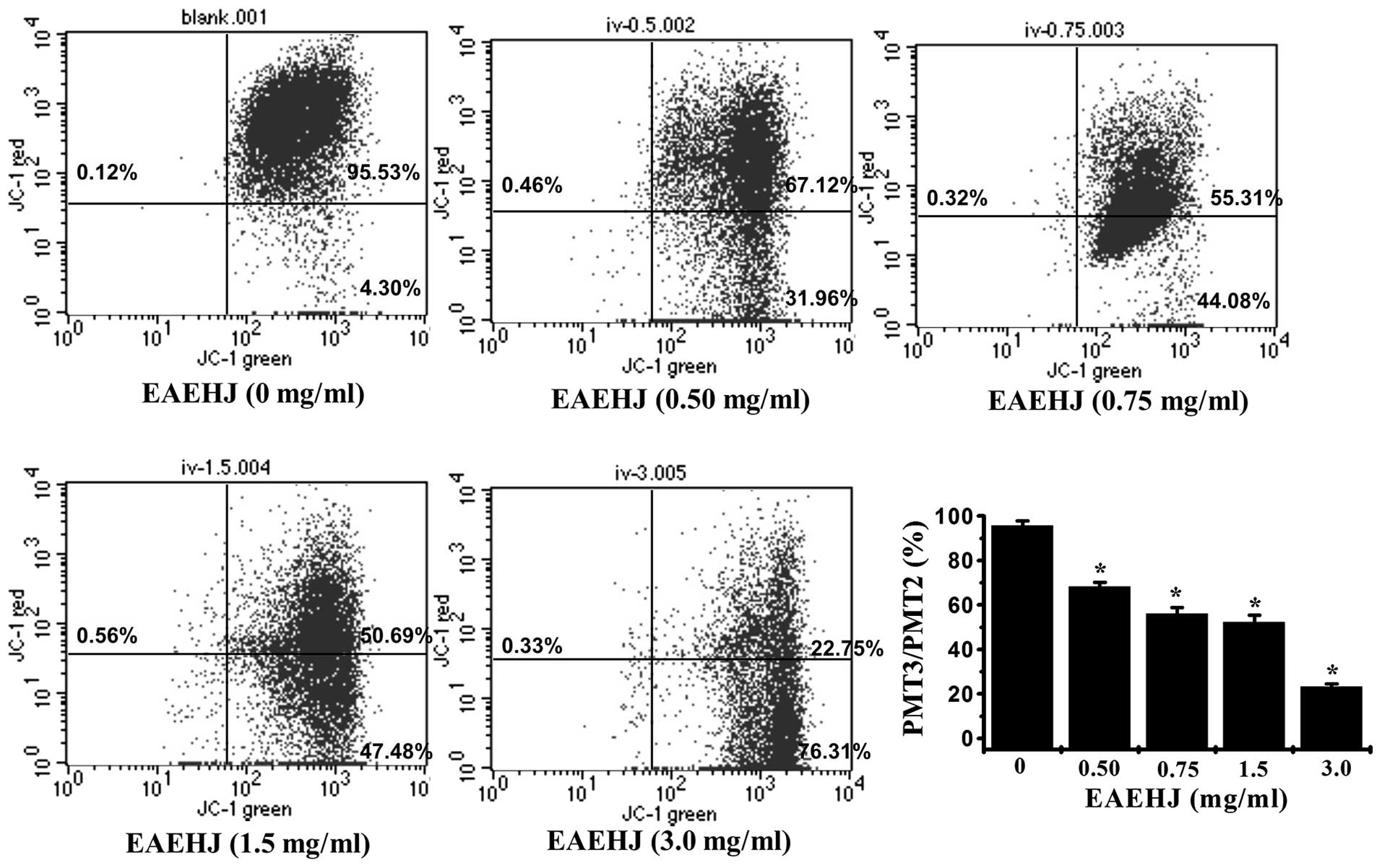
Effects of EAEHJ on the loss of ΔΨm
Early-stage cellular apoptosis is accompanied by
disruption of the mitochondrial membrane, which leads to a rapid
collapse of the electrochemical gradient. In the present study, the
effects of EAEHJ on ΔΨm were investigated using JC-1, a
mitochondria-specific dye. JC-1 is a lipophilic, cationic dye that
selectively enters mitochondria. In healthy cells with high ΔΨm,
JC-1 forms J-aggregates that exhibit intense red fluorescence
(detected in PMT3, 590 nm), whereas under apoptotic conditions, the
ΔΨm collapses, and JC-1 does not accumulate within the
mitochondria, but remains instead in the cytoplasm in its monomeric
form, emitting green fluorescence (detected in PMT2, 525 nm). Flow
cytometric analysis demonstrated the loss of ΔΨm in cells treated
with EAEHJ for 24 h in a dose-dependent manner. As shown in
Fig. 3, following treatment with
0, 0.5, 0.75, 1.5, and 3.0 mg/ml of EAEHJ, the ratio of PMT3 to
PMT2 was 95.8%, 68.2%, 56%, 52.2%, and 23.3%, respectively
(P<0.01). This dose-dependent change in ΔΨm indicates that EAEHJ
promotes the loss of ΔΨm in HepG2 human hepatoma cells.
Effects of EAEHJ on the expression levels
of Bax and Bcl-2 in H22 cell-bearing mice and HepG2 human hepatoma
cell line
Pro-apoptotic Bax and anti-apoptotic Bcl-2 are two
members of the Bcl-2 family that regulate the mitochondrial
signaling pathway during apoptosis. Mitochondria-dependent
apoptosis is primarily regulated by Bcl-2 family proteins. It has
been suggested that mitochondrial outer membrane permeabilization
(MOMP) occurs through the formation of mitochondrial pores by
pro-apoptotic Bax-like proteins, which may be inhibited by
anti-apoptotic Bcl-2 like members (14,15).
Therefore, the ratio of Bax to Bcl-2 is crucial for determining
cell fate. Higher Bcl-2 to Bax ratios, caused by the overexpression
of Bcl-2 and/or downregulation of Bax, are commonly detected in
various types of cancer (16),
which confers a survival advantage to the cancer cells, and is
associated with both chemotherapy and radiotherapy resistance. The
results of the IHC assay indicated that treatment with EAEHJ
reduced the expression levels of anti-apoptotic Bcl-2 in tumors,
whereas the expression of pro-apoptotic Bax was increased. As shown
in Fig. 4A, the percentage of Bax-
and Bcl-2-positive cells in the control group was 7.8±2.5 and
38.2±6.9%, respectively, whereas those of the EAEHJ-treated mice
were 16.4±3.5 (P<0.01, vs. controls) and 28±9.4% (P<0.05, vs.
controls), respectively. These results indicate that treatment with
EAEHJ may increase the pro-apoptotic Bax/Bcl-2 ratio. To our
knowledge, this is the first time that EAEHJ has been reported to
inhibit hepatoma cancer growth in vivo via the promotion of
cancer cell apoptosis.
In order to further study the mechanism underlying
the anti-carcinogenic effects of EAEHJ, RT-PCR and western blotting
were performed to examine the mRNA and protein expression levels of
Bax and Bcl-2 in the EAEHJ-treated HepG2 human hepatoma cells. The
results of the RT-PCR assay demonstrated that treatment with EAEHJ
significantly increased the mRNA expression levels of Bax and
reduced the mRNA expression levels of Bcl-2 in the HepG2 human
hepatoma cells (Fig. 4B). The
results from the western blot analysis showed that the protein
expression levels of Bax and Bcl-2 were similar to their respective
mRNA expression levels, suggesting that EAEHJ induces
mitochondria-dependent apoptosis of HepG2 human hepatoma cells
through regulation of the expression levels of Bcl-2 family
proteins (Fig. 4C).
Effects of EAEHJ on the activation of
caspases-3 and 9
Caspases form part of the cysteine protease family,
and are important proteins in the modulation of the apoptotic
response. Caspase-3 is a key regulator of apoptosis, and is
activated by an initiator caspase, such as caspase-9, during
mitochondria-mediated apoptosis (17). In order to identify the downstream
effectors of the apoptotic signaling pathway, the activation of
caspase-3 and 9 was examined using a colorimetric assay with the
following caspase-specific chromophores: DEVD-pNA, a specific
substrate of caspase-3, and LEHD-pNA, a specific substrate of
caspase-9. As shown in Figs. 5A and
B, treatment with EAEHJ significantly and dose-dependently
induced activation of both caspase-3 and 9 in the HepG2 human
hepatoma cells (P<0.01), as compared with the untreated control
cells. These results suggest that EAEHJ is able to promote HepG2
human hepatoma cell apoptosis via the mitochondria-dependent
pathway.
Discussion
Natural products, specifically Chinese herbal
medicines, have become increasingly important for their potency
against various types of cancer, whilst having the advantage of
fewer side effects and reduced drug resistance, as compared with
conventional chemotherapy and radiotherapy. In the present study,
using the H22 cell-bearing mouse model, it was demonstrated that
Hypericum japonicum may inhibit cancer growth in
vivo, with no apparent effect on murine body weight, indicating
that it had no apparent toxicity.
Apoptotic mechanisms are commonly exploited for
tumor therapy. Apoptotic cell death is recognizable by cellular
morphological characteristics, including membrane blebbing and
chromatin condensation (18). As
demonstrated by the current study, the control cells exhibited a
typical polygonal intact shape, whereas the cells treated with
various concentrations of Hypericum japonicum, became
rounded, shrunken, exhibited membrane blebbing, or became
non-confluent.
The initiation of apoptosis involves two major
signaling pathways: Extrinsic (death receptor) and intrinsic
(mitochondrial). The intrinsic apoptotic pathway is regulated by
mitochondria which has an central role in apoptosis (19). In the early stages of apoptosis,
the membrane phosphatidylserine translocates from the inner side of
the plasma membrane to the outer layer, and becomes exposed at the
external surface of the cell (20). Annexin V is a Ca2+
dependent phospholipid-binding protein with high affinity for
phosphatidylserine (19). Using
Annexin-V/PI staining followed by FACS analysis it was demonstrated
that Hypericum japonicum treatment was able to significantly
induce the apoptosis of HepG2 human hepatoma cells. MOMP is the
point of convergence for a large variety of intracellular apoptotic
signals leading to the release of numerous apoptogenic proteins
from the mitochondrial intermembrane space (21). Loss of mitochondrial membrane
potential (ΔΨm) has been suggested to contribute to cell death by
disruption of normal mitochondrial function (22,23).
The current study investigated whether Hypericum japonicum
is able to promote the deregulation of ΔΨm in HepG2 human hepatoma
cells.
The Bcl-2 family of proteins are important
regulators of apoptosis, including pro-apoptotic members such as
Bax and anti-apoptotic members such as Bcl-2 (24,25).
The ratio of active anti-apoptotic Bax to pro-apoptotic Bcl-2
determines cellular fate, and variations in this ratio results in
impaired apoptosis, leading to various diseases, including cancer
(26–28). RT-PCR, western blotting and IHC
were conducted to examine the mRNA and protein expression levels of
the pro-apoptotic protein Bax and the anti-apoptotic protein Bcl-2
in the Hypericum japonicum-treated human hepatoma cells. It
was demonstrated that Hypericum japonicum treatment
dose-dependently enhanced Bax expression and reduces Bcl-2
expression in at mRNA and protein levels.
Caspases, a family of cysteine proteases, are key
proteins that modulate the apoptotic response, and caspase-3 is a
key executioner of apoptosis (29,30).
In the current study, it was demonstrated that the induction of
apoptosis may involve the loss of mitochondrial transmembrane
potential, and caspase-9 and caspase-3 activation.
In conclusion, to the best of our knowledge for the
first time, it is reported that Hypericum japonicum is able
to significantly inhibit the growth of H22-cell tumors without
affecting murine weight, indicating that Hypericum japonicum
is non-toxic with few side effects. The results of the present
study additionally demonstrated that Hypericum japonicum
promotes cancer cell apoptosis via the mitochondria-dependent
pathway in vitro. These results may help establish a
scientific foundation for future research and therapeutic
development of EAEHJ as an effective anti-carcinogenic agent.
Acknowledgments
The authors of the present study are grateful to Dr
Da Wo (Tongji University School of Medicine, Shanghai, China) for
reviewing the manuscript of the present study, and to Dr Zhenyu
Zhang (Tongji University School of Medicine) for editing the
figures of the present study. The present study was supported by
grants from the Open Foundation of Fujian Key Laboratory of
Integrative Medicine on Geriatrics (grant no. ZXY2008003), the
Development Foundation of Chen Keji Integrative Medicine (grant no.
CKJ2008007), the Education Department of Fujian Province (grant no.
JA10169), the Health Department of Fujian Province (grant no.
2010-1-39), and the Open Foundation of Fujian Health College (grant
no. 2010-2-10).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
3
|
Wang N, Li P, Wang Y, Peng W, Wu Z, Tan S,
Liang S, Shen X and Su W: Hepatoprotective effect of Hypericum
japonicum extract and its fractions. J Ethnopharmacol. 116:1–6.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang XW, Zhang DW, Wei XL, et al: Advances
in research on Hypericum japonicum. Chin J Mod Drug Appl.
3:183–185. 2009.In Chinese.
|
|
5
|
Shen YZ, Zhang XC and Tang WZ: Advances in
research on Hypericum japonicum of its chemical components and
pharmacological activities. Shangdong Pharm Ind. 22:28–29. 2003.In
Chinese.
|
|
6
|
Sun ZY, Jin GJ, Xu T, et al: Analysis of
30 patients of Primary Liver Cancer treated by Hypericum japonicum
thunb. Chin J Integr Trad Western Med Liver Dis. 5:29–30. 1995.In
Chinese.
|
|
7
|
Li QX, Peng RX and Gao P: Hepatoprotective
effect of Tianjihuang injection against APAP-induced hepatic
toxicity in mice. Chin Pharm J. 27:472–474. 1992.In Chinese.
|
|
8
|
Dong J, Liu Y, Liang Z and Wang W:
Investigation on ultrasound-assisted extraction of salvianolic acid
B from Salvia miltiorrhiza root. Ultrason Sonochem. 17:61–65. 2010.
View Article : Google Scholar
|
|
9
|
Guide for the Care and Use of Laboratory
Animals; National Research Council (US) Institute of Animal
Research: National Academies Press (US); Washington (DC), USA:
1996
|
|
10
|
Chang A, Cai Z, Wang Z and Sun S:
Extraction and isolation of alkaloids of Sophora alopecuroides and
their anti-tumor effects in H22 tumor-bearing mice. Afr J Tradit
Complement Altern Med. 11:245–248. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gao L, Chen L, Fei XH, Qiu HY, Zhou H and
Wang JM: STI571 combined with vincristine greatly suppressed the
tumor formation of multidrug-resistant K562 cells in a human-nude
mice xenograft model. Chin Med J (Engl). 119:911–918. 2006.
|
|
12
|
Jin Y, Li J, Rong LF, Li YH, Guo L and Xu
SY: Anti-hepatocarcinoma effects of 5-fluorouracil encapsulated by
galactosylceramide liposomes in vivo and in vitro. World J
Gastroenterol. 11:2643–2646. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen H, Takahashi S, Imamura M, Okutani E,
Zhang ZG, Chayama K and Chen BA: Earthworm fibrinolytic enzyme:
Anti-tumor activity on human hepatoma cells in vitro and in vivo.
Chin Med J (Engl). 120:898–904. 2007.
|
|
14
|
Chipuk JE and Green DR: How do BCL-2
proteins induce mitochondrial outer membrane permeabilization?
Trends Cell Biol. 18:157–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Arnoult D: Apoptosis-associated
mitochondrial outer membrane permeabilization assays. Methods.
44:229–234. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Souers AJ, Leverson JD, Boghaert ER,
Ackler SL, Catron ND, Chen J, Dayton BD, Ding H, Enschede SH,
Fairbrother WJ, et al: ABT-199, a potent and selective BCL-2
inhibitor, achieves antitumor activity while sparing platelets. Nat
Med. 19:202–208. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma Y, Zhang A, Shi Z, He C, Ding J, Wang
X, Ma J and Zhang H: A mitochondria-mediated apoptotic pathway
induced by deoxynivalenol in human colon cancer cells. Toxicol In
Vitro. 26:414–420. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lütticken C, Wegenka UM, Yuan J, Buschmann
J, Schindler C, Ziemiecki A, Harpur AG, Wilks AF, Yasukawa K, Taga
T, et al: Association of transcription factor APRF and protein
kinase Jak1 with the interleukin-6 signal transducer gp130.
Science. 263:89–92. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kotwicka M, Filipiak K, Jedrzejczak P and
Warchol JB: Caspase-3 activation and phosphatidylserine membrane
trans-location in human spermatozoa: is there a relationship?
Reprod Biomed Online. 16:657–663. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
David R: Apoptosis: A lipid trigger of
MOMP. Nat Rev Mol Cell Biol. 13:208–209. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ly JD, Grubb DR and Lawen A: The
mitochondrial membrane potential (deltapsi(m)) in apoptosis; an
update. Apoptosis. 8:115–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sesso A: Pitfalls in the use of electron
microscopy to study the mitochondrial membrane permeability
transition in apoptotic cells and pellets: Where do we stand in
relation to the incidence of mitochondrial swelling in apoptosis?
Braz J Morphol Sci. 23:57–74. 2006.
|
|
24
|
Adams JM and Cory S: The Bcl-2 apoptotic
switch in cancer development and therapy. Oncogene. 26:1324–1337.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shamas-Din A, Kale J, Leber B and Andrews
DW: Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb
Perspect Biol. 5:a0087142013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bishayee K, Ghosh S, Mukherjee A,
Sadhukhan R, Mondal J and Khuda-Bukhsh AR: Quercetin induces
cytochrome-c release and ROS accumulation to promote apoptosis and
arrest the cell cycle in G2/M, in cervical carcinoma: Signal
cascade and drug-DNA interaction. Cell Prolif. 46:153–163. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Smerage JB, Budd GT, Doyle GV, Brown M,
Paoletti C, Muniz M, Miller MC, Repollet MI, Chianese DA, Connelly
MC, et al: Monitoring apoptosis and Bcl-2 on circulating tumor
cells in patients with metastatic breast cancer. Mol Oncol.
7:680–692. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Thomas S, Quinn BA, Das SK, Dash R, Emdad
L, Dasgupta S, Wang XY, Dent P, Reed JC, Pellecchia M, et al:
Targeting the Bcl-2 family for cancer therapy. Expert Opin Ther
Targets. 17:61–75. 2013. View Article : Google Scholar
|
|
29
|
Brentnall M, Rodriguez-Menocal L, De
Guevara RL, Cepero E and Boise LH: Caspase-9, caspase-3 and
caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell
Bio. 14:322013. View Article : Google Scholar
|
|
30
|
Cohen GM: Caspases: The executioners of
apoptosis. Biochem J. 326:1–16. 1997. View Article : Google Scholar : PubMed/NCBI
|