1. Introduction
The hepatitis B virus (HBV) infection is a prevalent
type of infectious disease that is causing a global concern for
public health (1). Although there
has been considerable improvement in understanding the molecular
virology and pathogenesis of the HBV infection over the past decade
and effective therapeutic measures have been developed for its
treatment, there are currently 240 million individuals globally who
are chronic HBV carriers, and ~620,000 succumb per year due to late
sequelae of liver cirrhosis or hepatocellular carcinoma (HCC)
(1). The treatment of chronic
hepatitis (CH) B is currently limited; predominantly consisting of
interferons (IFNs) and nucleoside analogs (NUCs) (2). NUCs are efficient antiviral agents,
however, NUCs only control rather than cure HBV infections due to
persistent viral covalently closed circular (ccc)DNA. Therefore, a
long-term treatment is required, which is expensive and may lead to
concomitant resistance (2). IFN
therapy is associated with side effects, and treatment with this
cytokine can result in viral clearance in a small proportion of
patients (3). Thus, it is
important to continue conducting research in order to identify
therapeutic targets to stimulate the development of novel antiviral
agents and immunotherapies.
The activation-induced cytidine deaminase
(AID)/apolipoprotein B mRNA editing enzyme, catalytic
polypeptide-like (APOBEC) family, which was first described for
inhibition of human immunodeficiency virus type 1 (HIV-1), is
important in the innate immune system, as it defends against
viruses, including HBV through hypermutation-dependent and
-independent mechanisms. Nine of 11 APOBEC3 family members have
been identified to exert varying levels of activity against HBV
under experimental conditions (Table
I). APOBEC-3A and APOBEC-3B are essential for cccDNA
degradation by IFN-α or the lymphotoxin-β receptor-agonist without
damaging the infected host cells (4). Thus, targeting the formation and
subsequent processing of viral cccDNA may be more rational
approaches.
 | Table IReported hyperediting and restriction
activity of AID/APOBECs against HBV DNA. |
Table I
Reported hyperediting and restriction
activity of AID/APOBECs against HBV DNA.
| Cytidine
deaminases | Hyperediting
activity | Dinucleotide
preference | Restriction
activity against HBV DNA | Reference |
|---|
| AID | Deaminates each
viral RNA and HBV minus-strand DNA | GpC, ApC | Active | 25,27 |
| A1 | Deaminates HBV
minus-strand DNA | TpC | Active | 19,21,25 |
| A2 | NI | NI | Not active | 25 |
| A3A | Deaminates each
strand of HBV DNA | Weak bias | Active/not
activea | 4,17,25 |
| A3B | Deaminates each
strand of HBV DNA | GpC, ApC | Active | 4,16,24–26,28,37 |
| A3C | Deaminates HBV
minus-strand DNA | No bias | Active but weak/not
activea | 16–18,20,26,37 |
| A3DE | Not active | N/A | NI | 17 |
| A3F | Deaminates each
strand of HBV DNA | TpC, GpC | Active | 16–18,20,25,26,37 |
| A3G | Deaminates each
strand of HBV DNA | CpC | Active | 16–18,20,26,37,75 |
| A3H | Deaminates each
strand of HBV DNA | TpC | NI | 17,20 |
The aim of the present study was to review the
rapidly accumulating evidence for the involvement of AID/APOBEC
cytidine deaminase expression and HBV replication, and to summarize
the current knowledge surrounding the role of editing in
HBV-associated liver disease as a consequence of its actions in
host cells.
2. AID/APOBEC family of deaminases
AID/APOBEC family members
All members of the AID/APOBEC family possess one or
two catalytic domains that deaminate cytidine in RNA and DNA. The
deaminases mediate the hydrolytic removal of an amino group at the
C4 position of a cytidine (C) or deoxycytidine (dC) generating a
uridine (U) or deoxyuridine (dU), respectively (5). The presence of the enzyme in cells
producing RNA virus results in C-to-U conversion of minus strand
reverse transcripts and G-to-A in plus strand DNA. The binding to
the target DNA creates a U-G mismatch, which generates a C-to-T
transition in minus strand DNA and a G-to-A transition in plus
strand DNA during general DNA replication without repair pathways
(Fig. 1) (6). In humans, the family comprises 11
members with distinct functions, including AID, APOBEC1 (A1),
APOBEC2 (A2), APOBEC4 (A4) and APOBEC3 (A3) subgroups. The A3 group
consists of seven proteins: A3A, A3B, A3C, A3DE, A3F, A3G and A3H
(7–10). The seven A3 genes are arranged in a
tandem gene cluster on chromosome 22 in humans (11). The presence of the AID/APOBEC
family is restricted to vertebrates. AID and A2 are likely to be
the ancestral members, while A1 and A3 are later evolutionary
arrivals (12), A3s are restricted
to placental mammals, and their gene copy number is
species-specific. For example, mice only possess a single A3 gene,
pigs have two, sheep and cattle have three, cats have four, horses
have six and primates have at least seven A3 genes (13). The rapid expansion of the A3 locus
in humans indicates an important role in the host genome defense
against exogenous viruses and endogenous retroelements (12–14).
The role of the AID/APOBEC family in the inhibition of viral
infection was initially described for HIV-1. Various studies have
shown that the genome of hepadnaviruses is hyperedited by cytidine
deaminases (15–21). Recent reports demonstrated that HBV
DNA replication is restricted by A1, AID, A3A, A3B, A3C, A3G, A3F,
but not A3DE (4,15,17,22–26),
specifically via the degradation of HBV cccDNA (4), which was investigated in an
experimental setting through deaminase-independent and -dependent
mechanisms (27,28).
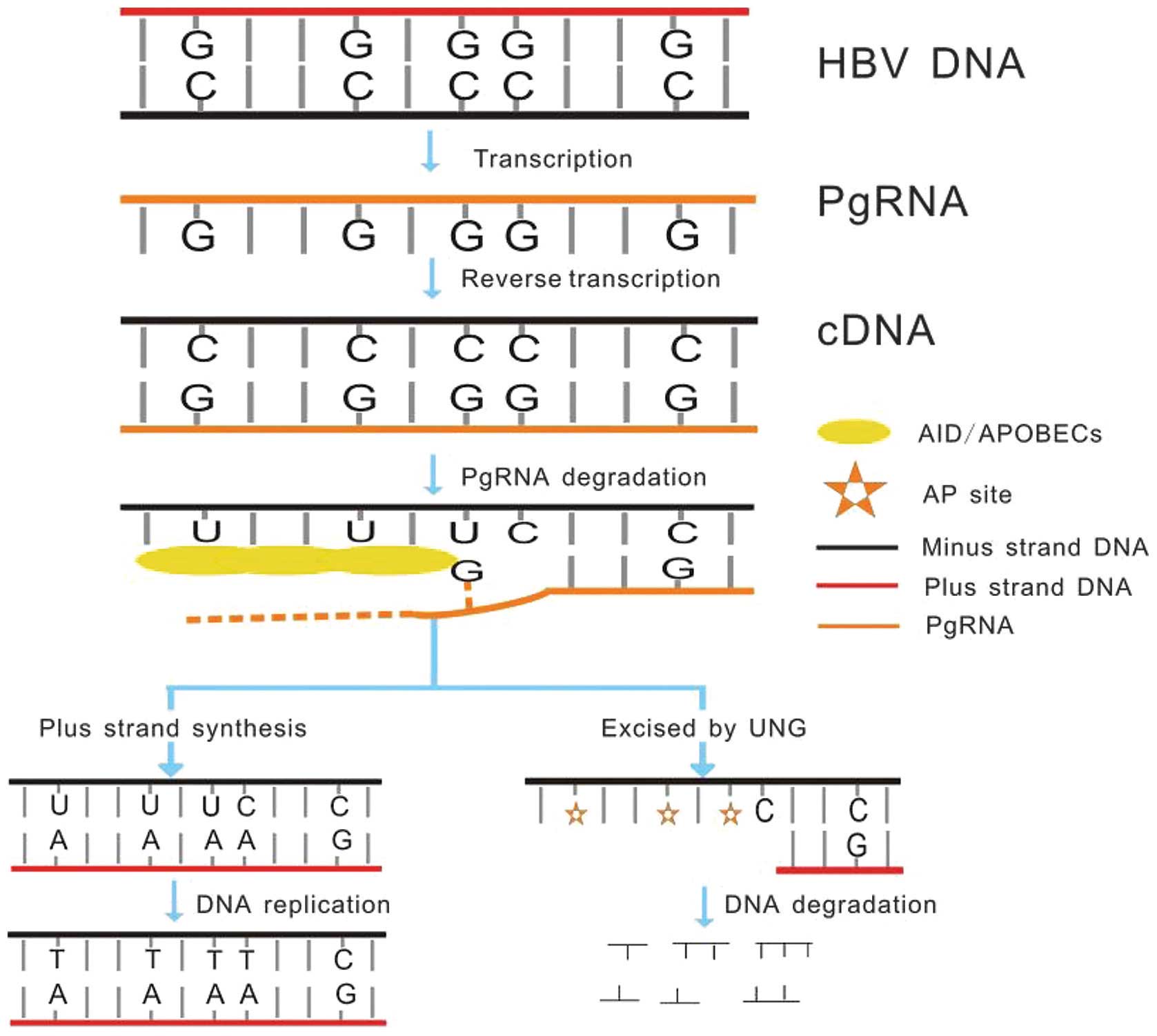 | Figure 1HBV DNA is transcribed to pgRNA as a
replicative RNA intermediate, according to which the minus DNA
strand (cDNA) forms. ssDNA is formed as the pgRNA is degraded.
AID/APOBECs catalyze cytosine deamination of HBV DNA on the cDNA,
producing uracil during reverse transcription. Uracils in DNA
(including cccDNA) are recognized and excised by UNG leading to
formation of AP sites. These AP sites are recognized by cellular AP
endonucleases leading to DNA digestion. The HBV DNAs that do not
undergo degradation, generate C-to-T transitions in minus strand
DNA and G-to-A transitions in plus strand DNA during general DNA
replication. HBV, hepatitis B virus; pgRNA, pregenomic RNA; ssDNA,
single stranded DNA; AID, activation-induced cytidine deaminase;
APOBEC, apolipoprotein B mRNA editing enzyme, catalytic
polypeptide-like; cccDNA, covalently closed circular DNA; UNG,
uracil DNA glycosylases; AP, apurinic/apyrimidinic. |
AID/APOBEC family gene expression
A1 is primarily expressed in the gastrointestinal
compartment and catalyzes the expression of a truncated form of
apolipoprotein (Apo) B (ApoB48) with the distinct biological
function of ApoB100 (10). AID is
expressed in germinal center B cells and randomly edits dC residues
to dU in the variable region of immunoglobulin gene loci, which is
essential for the process of antibody diversification (29,30).
As expected, AID and A1 were not detected in the HCC cell lines and
liver tissue (20,25). A2 is widely expressed in muscle;
predominantly in cardiac and skeletal muscle, thus, is exclusively
associated with the development of cardiac and skeletal muscle, as
well as early embryogenesis (9,31–33).
A4 was discovered by a computational homology search and was found
to be expressed in the testicles, although its function remains
undetermined (8). A3 (particularly
A3G and A3F) is widely expressed in human tissues, where the mRNA
levels broadly correlate with the lymphoid cell content (with the
exception of gonadal tissues) (34). In the healthy human liver, low to
moderate APOBEC expression levels have been observed (11,20,26,35,36),
with A3G expression levels identified to be the highest, followed
by A3C and A3H expression (18).
Positive transcription and expression has been identified for A3G,
A3C and A3H in HepG2 cells, although not for A3B and A3F.
Amplification experiments with A2- and A3A-specific primers
resulted in particularly faint signals, and A1- and A3DE-specific
sequences were not detected (18).
Onset of liver disease results in changes to the
levels of these deaminases. For example, in hepatitis/cirrhotic
samples, five of seven A3 genes were significantly upregulated in
the following order: HCV ± HBV > HBV > alcoholic cirrhosis.
A3C and A3D were upregulated for all groups, while IFN inducible
A3G was overexpressed in virus-associated cirrhotic patients, and
AID was present in 50% of the HBV/HCV samples (21). The expression levels of A1, A2, and
A4 were not identified to be significantly different between
cirrhotic liver and normal tissue samples (21). Furthermore, the A3B cytidine
deaminase was widely upregulated in HCC tumor tissue samples
(37).
Upregulation by IFNs and other
cytokines
The expression of APOBEC genes is induced by IFNs,
as well as other cytokines. IFN-α has been observed to trigger the
expression of A3 in numerous reports. For example, treatment with
IFN-α in primary human hepatocytes (PHHs) and HCC cell lines
resulted in induction of A3-associated genes, particularly A3G;
however, no changes were triggered in the expression of A1, A2 or
AID (25,26,36).
In addition, A3A was demonstrated to be significantly expressed in
peripheral blood mononuclear cells, HBV-infected differentiated
HepaRG cells and PHHs that were treated with IFN-α (4,36,38).
IFN-α enhances A3 expression levels in human macrophages (39), phagocytes (38) and peripheral plasmacytoid dendritic
cells (40). This indicates that
A3 expression may contribute to the antiviral effects of IFN-α
against HBV DNA. In addition to IFN-α, other IFN family members,
such as IFN-γ and IFN-β, directly upregulate A3G expression in
macrophages (39,41). IFN-γ is known to control HBV in a
dose-dependent reduction manner, which was consistent with the
observation of a dose-dependent increase in A3G and A3F protein
expression following IFN-γ therapy (42). In addition, the expression of AID
is induced in response to tumor necrosis factor (TNF)-α.
Interleukin (IL)-1β stimulation in cultured human hepatocytes and
pro-inflammatory cytokine-mediated expression was meditated by
nuclear factor-κB signaling pathways (43,44)
and by IL-4 and -13 through STAT6 (45). Transforming growth factor-β is a
cytokine that induces AID in B and HepG2 cells (46), whereas TNF-α and IL-1β stimulate
the expression of A2 in human hepatocytes (44).
3. Specific regulation of editing by
AID/APOBECs
Mechanism of regulation
The mutagenic activity resulting from
AID/APOBEC-mediated deamination is toxic for retroviruses and
beneficial to organisms in host cell defense (13). However, its excessive activity and
off-target mutations within the cellular genome are also toxic and
oncogenic to the host. Thus, organisms possess multiple mechanisms
to regulate the expression of AID/APOBECs; for example, controlling
their enzymatic activity and restricting their access to DNA or RNA
substrates (47). A1 is the sole
family member capable of recognizing and using mRNA as a substrate.
Editing only occurs on cytidines 5′ of the mooring sequence
(48). Although RNA editing occurs
in the cytoplasm when A1 is overexpressed (49), cytoplasmic A1 editing activity is
suppressed under normal physiological conditions. In addition, RNA
editing is restricted to the cell nucleus within a temporal and
spatial window that occurs subsequent to pre-mRNA splicing and
prior to mRNA nuclear export, only when interacting with cofactor,
APOBEC-1 complementation factor (50,51).
The sequence requirements for AID/APOBEC deamination are not
stringent, however, deamination generally occurs within transcribed
or single-stranded regions of DNA. Therefore, single-stranded
(ss)DNA is always recognized as a substrate by the AID/APOBEC
family. The deaminase activity of AID is responsible for a variety
of point mutations and DNA breaks. This mutagenic activity leads to
somatic hypermutation (SHM) and class switch recombination
(52). Notably, the expression of
AID is restricted to activated B cells under physiological
conditions, and only targets variable and switch regions of
immunoglobulin genes for mutagenesis. A2 is essential for muscle
development with its expression being largely restricted to
striated muscle. However, in vitro (32,53)
A2 was identified to lack autonomous deaminase activity. The
above-mentioned processes are regulated in a tissue-specific manner
during development and in response to metabolic regulation.
Furthermore, subcellular localization and RNA binding to
AID/APOBECs regulate their deaminase activity. Furthermore, AID is
regulated by nuclear-cytoplasm transport and is predominantly
located in the cytoplasm. Controlling the abundance of AID
exclusion from the nucleus in steady state is one of the major
regulatory mechanisms restricting its contact with genomic material
(54,55). A3G is strongly retained in the
cytoplasm and excluded from chromosomes based on the specialized
property of a novel cytoplasmic retention signal (56). A3G is present in two distinct
molecular forms within the cell: A low molecular mass form and a
high molecular mass (HMM) complex, which contains one or more
inhibitory RNAs that inactivate ssDNA deaminase activity (57–59).
A3G-RNA complexes in viral particles have been found to be inactive
until RNase H activity of reverse transcriptase (RT) degrades the
RNA of DNA-RNA hybrids (60). A3C,
A3F and A3H all form intracellular HMM complexes (61–63)
in addition to A3G. AID exerts no measurable deaminase activity on
ssDNA unless the AID is pretreated with RNase to remove the
inhibitory RNA, which is bound to the AID (29).
4. Role of AID/APOBECs in HBV-associated
disease progression
Promotion of the evolution of HBV
virus
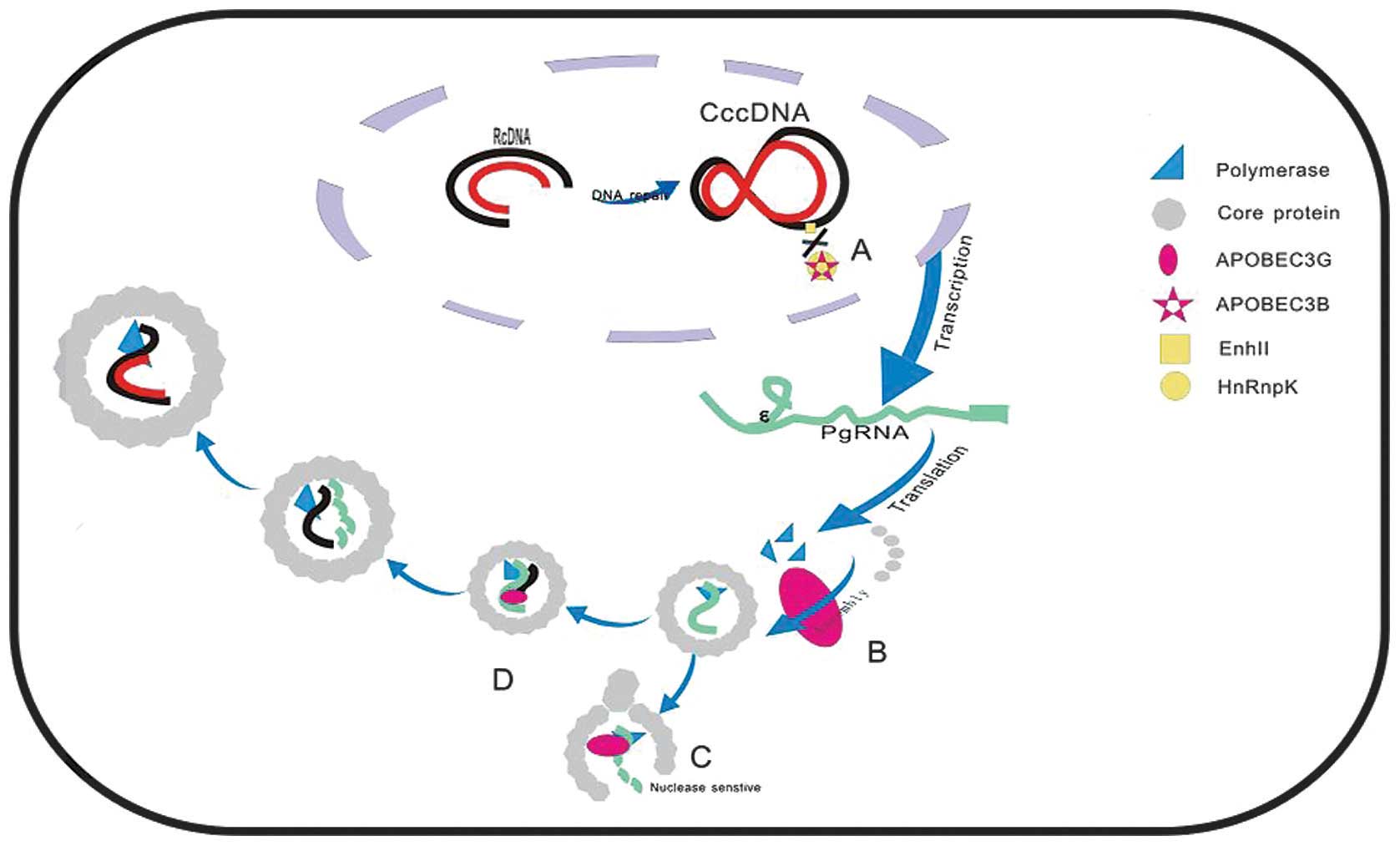
The encapsidated HBV genome consists of a 3.2-kb
partially double-stranded relaxed circular (rc)DNA molecule. Upon
translocation to the nucleus, the rcDNA genome is converted into a
cccDNA by cellular repair factors and remains an episomal
minichromo-some, which transcribes all viral RNAs, including
pregenomic (pg)RNA as a replicative RNA intermediate. The pgRNA is
translated to form the core protein and internal translation
initiation synthesizes the DNA polymerase. The pgRNA, viral core,
and polymerase proteins are assembled into the nucleocapsid in the
cytoplasm (1). Reverse
transcription starts to synthesize the (minus) DNA strand via
reverse transcription activity of the viral polymerase. Following
degradation of the pgRNA by the viral RNase H, the plus strand is
synthesized to form mature core particles, which interact with the
preS domain of the membrane-associated viral surface proteins and
acquire the viral envelope (64).
Approximately 1011 viral particles are released per day
into the circulation of individuals with the chronic HBV infection
and HBV particles are cleared from the plasma with a half-life of
~1.0 day (65,66). In the initial immunotolerant phase
of chronic HBV infection, the immune pressure is weak. With the
progression of chronic infection, especially during Hepatitis B e
antigen (HBeAg) seroconversion, HBV mutations gradually occur
(67). The mutation rate of HBV
DNA caused by AID/APOBECs in chronic HBV infection patients has
been shown to be higher than that of acute HBV infection patients.
In the chronic HBV infection group, the frequency of hypermutated
genomes was found to be higher in HBeAg-negative individuals when
compared with seropositive cases (68,69),
and the degree of mutation of the HBV DNA was significantly
associated with the extent of fibrosis (68). This may have been due to the fact
that HBeAg-negative patients had undergone HBeAg seroconversion,
resulting in an already elevated immune response (70). Although mutations in A3 that result
in altered editing can be highly deleterious, lightly edited
genomes may facilitate the virus to evolve and even escape from the
immune responses (21).
Non-resolving inflammation has been shown to be indispensable for
immune-selection of the AID/APOBECs-dependent HBV mutations
(6).
5. Inhibition of HBV by AID/APOBEC
proteins
A3s are effective inhibitors of HBV DNA
replication
A3s are effective restrictive factors for various
types of virus with ssDNA, including HBV. It has been shown that A3
catalyzes cytosine deamination of HBV DNA produced during reverse
transcription as a template, resulting in the presence of uracil in
cDNA. Uracils in DNA (including cccDNA) are recognized and excised
by cellular uracil DNA glycosylases (UNGs) leading to formation of
apurinic/apyrimidinic (AP) sites, which are then recognized by
cellular AP endonucleases resulting in DNA digestion (Fig. 1) (4,71–73).
Certain studies describe contradictory results and report that the
loss of UNG does not restore the levels of viral reverse
transcripts, which had been decreased by A3G (74). A previous study found that HBV
replication was inhibited by A3G (75). However, the G-to-A hypermutation
was not observed in HBV DNA. Further studies using a 3D-polymerase
chain reaction (PCR) method showed that A3B, A3C, A3F and A3G
extensively deami-nated cytidine residues in minus strand DNA.
Unexpectedly, three of the four enzymes (A3B, A3F and A3G)
deaminated HBV plus strand DNA in human HCC cell lines; however,
the underlying mechanism remained undetermined (16). In another study, Henry et al
(17) compared the HBV editing by
all seven enzymes in a quail cell line, which did not produce any
endogenous DNA cytidine deaminase activity. The study identified
that all of the A3s, not including A3DE, deaminated the HBV DNA
levels from 10−2 to 10−5 in vitro,
with A3A proving to be the most efficient editor (17). The hyperediting ability of A3A was
confirmed by Lucifora et al (4). Although A3C was weaker at inhibiting
HBV replication compared with A3G, its capacity for DNA
hyperediting was higher than A3G and A3H (18,20).
A3G demonstrated a strong preference for CpC, whereas A3H
demonstrated a preference for TpC (19). These findings indicate that human
A3 enzymes impact HBV replication by cytidine deamination. However,
the induction of hypermutations is not sufficient for full
inhibition of HBV replication.
A cytidine deaminase null mutant of A3B (E255A) was
also found to inhibit HBV RNA production, although it was unable to
edit HBV (28). It was proposed
that only the carboxy (C)-terminal deaminase domain of A3B
catalyzes HBV hypermutations (24,26).
By investigating a series of A3B point mutants, it was found that
the N- and C-terminal cytosine deaminase domains of A3B (24) and A3G (76) exerted inhibitory effects on HBV
replication. By contrast, a truncated splice variant of A3B that
lacked the C-terminal deaminase domain exerted no effect on HBV
replication (26). However, A3G,
which was defective for deaminase activity, was found to inhibit
HBV replication (75). From the
above-mentioned reports, it is hypothesized that the inhibition of
HBV may be mediated by additional, different mechanisms.
Possible editing-independent
mechanisms
In principle, a decrease in HBV DNA may result from
either accelerated DNA degradation or decreased synthesis. A
previous report suggested that A3G decreased the viral DNA levels
by inhibiting pgRNA packaging (75). Subsequently, Rösler et al
(15) revealed that the early
stages of HBV DNA morphogenesis, including RNA and protein
synthesis, binding of pgRNA to the core protein, and self-assembly
of the viral core protein, were unaffected. However, A3G rendered
HBV core protein-associated full-length pgRNA nuclease sensitive.
Nguyen et al (27) analyzed
the mechanisms of deamination-independent suppression of HBV
replication by A3G and did not identify enhanced DNA degradation by
A3G, either in vivo or in vitro. A3G appeared to
inhibit the very early steps in viral reverse transcription and
blocked DNA strand elongation. Furthermore, the
deamination-independent antiviral function of A3G targets DNA-RNA
hybrids (27). The A3B protein
inhibits the binding of heterogeneous nuclear ribonucleoprotein K
to the enhancer II of HBV and represses the activity of HBV. In
addition, A3B directly suppresses HBV S-promoter activity (23). The process is presented
schematically in Fig. 2.
Inhibition of HBV by additional APOBEC
proteins
AID and A1 also inhibit HBV replication. AID was
significantly upregulated in ~50% of HBV and HCV cirrhotic samples
indicating that ectopic expression of AID was also a feature of HBV
liver disease. A1 was markedly expressed in one sample (although
normalization of the expression levels was not conducted), A2
transcripts were detected in certain samples, but A4 was not
detected in any sample. AID was identified to be as effective as
A3G at hyperediting HBV DNA (21).
Mutation matrices and editing frequencies in the HBV minus strand
DNA were comparable, for example, the editing frequency for A1 was
32%, and for A3G was 34% (18).
C-to-T (U) hypermutations were observed in rcDNA and nucleocapsid
viral RNA indicating that AID deaminates viral RNA and ssDNA
(77). A1 exhibited a strong
preference for TpC and a strong aversion to GpC (19) and AID editing of the HBV target was
concentrated at the GpC and ApC sites (21). Furthermore, the low expression
levels of A1 and AID in Huh7 cells was consistently associated with
decreased levels of core-associated HBV DNA by quantitative PCR.
However, A2 did not exert any effect on HBV replication (25).
AID/APOBECs incorporation into viral
particles
Research on HBV reveals that the antiviral activity
of A3G requires incorporation into assembling viral particles to
inhibit reverse transcription. Factors required for incorporation
of the antiviral deaminase protein, A3G into HBV nucleocapsids
continues to be investigated. It has been demonstrated that A3G and
A3C bind to the HBV core protein in immunoprecipitation experiments
(78,79). Such binding is essential for
reverse transcription following infection. The binding of A3G to
the HBV core protein was only indirectly demonstrated with
coexpression of RT and pgRNA, however, not with core protein alone
(15,18). The results are consistent with the
findings of Nguyen et al (80) that A3G was specifically
incorporated into replication-competent HBV nucleocapsids by
interacting with viral RT and RNA packaging signals. However, by
fluorescence resonance energy transfer (FRET) and acceptor
photobleaching experiments, Zhao et al (81) revealed that A3G directly binds to
core proteins. Similarly, direct interaction of HBV core protein
and A3A was confirmed by proximity ligation assay and FRET
analysis. Deletion analysis was used to confirm that the central
region of the HBV core protein (between aa 77 and 149) was involved
in the interaction with A3A (4).
Additionally, the A3B, A3C and A3F enzymes were also found to be
associated with the HBV capsid by interaction with the core protein
(16). Similar to A3G, AID was
co-immunoprecipitated with the nucleocapsid core protein. The
assumption was made that AID formed a ribonucleopro-tein complex
with the HBV core proteins and RNA during nucleocapsid assembly in
which AID deaminated cytosines of the viral RNA, including pgRNA
and ssDNA (77).
6. APOBEC polymorphisms in HBV
infections
The APOBEC family is considered to be significant in
innate cellular immunity, particularly in HBV hyperediting and
infection inhibition. It was of interest to elucidate whether gene
polymorphisms alter host susceptibility to HBV progression of CH.
A3B is an efficient editor and inhibitor in the process of HBV
infection. A 29.5-kb deletion between exon 5 in A3A and exon 8 in
an A3B gene cluster resulted in complete removal of the A3B coding
region (82,83). A hybrid gene containing the coding
region of A3A and the 3′-untranslated region of A3B was initially
discovered by mapping end-sequence pairs from a human fosmid
library against the human genome reference sequence assembly
(82). This was confirmed by dense
single nucleotide polymorphism marker mapping (83). A report on the frequency of the
deletion allele conducted using 1,277 diverse human samples
revealed that its expression is rare in African and European
individuals (frequency of 0.9 and 6%, respectively), more common in
East Asian and Amerindian individuals (36.9 and 57.7%,
respectively) and almost universal in Oceanic populations (92.9%)
(84). Analyzing the association
between this polymorphism and chronic HBV infection may clarify the
effect of A3B on the establishment of chronic HBV carrier state.
Abe et al (85) initially
investigated the association between this polymorphism and chronic
HBV infection. No association was identified between the chronic
HBV carrier state and deletion polymorphisms; however, the extent
of liver fibrosis was found to be associated with insertion
homozygosity (85). Similarly,
another report demonstrated that there was no significant
difference in the frequencies of deleted A3B alleles or genotypes
between patients with CH B and control subjects. By contrast,
subjects bearing a deleted genotype experienced a more rapid
progression of liver disease when compared with those exhibiting an
insertion genotype (86). This
result indicated that in the A3B deletion polymorphism, A3 cytidine
deaminases do not predispose an individual to chronicity, but may
modulate the course of persistent HBV infection. These results are
contradictory to a study, which revealed that a 29.5-kb deletion
obliterated the production of A3B, and was significantly associated
with increased susceptibility to persistent HBV infection in an
ethnic Han Chinese population (28).
Seven haplotypes (designated with Roman numerals,
I-VII) were reported to be present in various human populations
based on the five single amino acid polymorphisms, N15Δ, R18L,
G105R, K121D and E178D (87). The
N15Δ and R105G mutations may independently cause a marked decrease
in A3H expression. Only the haplotypes II, IV and VII were without
these two mutations, and were stably expressed and found to
efficiently inhibit HIV-1 replication (88). In addition, Wang et al
(87) found that E140K, H54R,
E56Q, C85S and C88S, as well as the two mutants, W115A and W115L,
disrupted A3H expression. The A3G H186R polymorphisms in
HBV-infected patients were found to be different to those of
healthy Moroccan individuals, although the difference was not
significant (86). The outcomes of
a selection of studies are summarized in Table II. The association between
AID/APOBEC polymorphisms and liver disease requires further,
in-depth investigation.
| Table IISummary of linkage studies between
clinical indicators and AID/APOBEC. |
Table II
Summary of linkage studies between
clinical indicators and AID/APOBEC.
A, APOBEC3
polymorphism studies
|
|---|
| Study (ref.) | Cohort
description | Association with
APOBEC3 polymorphisms |
|---|
| Abe et al,
2009 (85) | 724 patients with
chronic HBV infection and 469 healthy control subjects. | No significant
association between A3B deletion polymorphism and chronic HBV
carrier state. A3B gene deletion homozygosity was associated with
mild liver fibrosis. |
| Zhang et al,
2013 (28) | 1,124 patients with
HBV-associated HCC, 510 individuals with persistent HBV infection
and 826 healthy control subjects.
Population, Han Chinese. | At least one A3B
deletion allele increased the risk for persistent HBV infection and
HCC development. |
| Ezzikouri et
al, 2013 (86) | 179 HBV chronic
carriers and 216 healthy control subjects.
Population, Moroccan. | No significant
difference in the frequencies of deleted A3B alleles or genotypes
between the two groups. Patients with deleted genotypes experienced
a faster progression of liver disease than those with insertion
genotypes.
A3B deletion exhibited significantly lower viral loads than
patients with the wild-type. A3G H186R polymorphism R/R genotype
frequencies were not significantly different in the HBV-infected
patients and healthy subjects. |
B, Hypermutation
studies
|
|---|
| Study (ref.) | Cohort
description | Association with
hypermutation |
|---|
| Beggel et
al, 2013 (68) | 80 treatment-naïve
HBV infection patients (47 HBeAg-positive and 33
HBeAg-negative). | Hypermutation rates
for HBeAg-negative patients were >10-fold higher than those of
HBeAg-positive patients. HBeAg-negative patients higher
hypermutation rates were significantly associated with the degree
of fibrosis. |
| Noguchi et
al, 2005 (69) | 8 Japanese adult
patients with acute HBV infection, 10 patients were chronic
carriers. | Hypermutated HBV
DNA was detected in 1/8 patients with acute HBV infection and 4/10
patients with chronic HBV infection. In the latter group,
hypermutated genomes were found only in eAb-positive patients. |
C, APOBEC3
expression studies
|
|---|
| Study (ref.) | Cohort
description | Association with
APOBEC3 expression |
|---|
| Vartanian et
al, 2010 (21) | 41 cirrhotic
samples (10 alcoholic cirrhosis, 10 HBV+, 11
HBV+HCV+, 10 HCV+, 4 normal
livers). | 5/7 A3 genes were
significantly upregulated in the order: HCV ± HBV > HBV >
alcoholic cirrhosis. A3C and A3D were up regulated for all groups.
Interferon inducible A3G was overexpressed in virus-associated
cirrhosis, as was AID in 50% of these HBV/HCV samples. |
| Xu et al,
2007 (37) | 29 pairs of HCC and
surrounding non-cancerous tissue samples. | A3B transcripts
were significantly elevated in 24/29 HCC. |
| Kou et al,
2007 (46) | 51 HCC patients (14
HBV+, 30 HCV+) with 25 CH and 26 LC patients,
and 6 normal livers. | AID was
significantly upregulated in HCC and surrounding non-cancerous
liver tissues underlying CH or LC. |
7. Emerging role of the AID/APOBEC family in
the development of cancer
The AID/APOBEC family and their associated editing
patterns may be involved in oncogenesis. Recent analyses of the
mutations have implicated APOBEC cytidine deaminases as significant
factors in the mutagenesis of human cancer genomes (89,90).
Within cancer genomes, APOBEC mutagenesis has been found to be
pervasive and correlated with APOBEC mRNA levels (89–91).
Roberts et al (89)
identified that tumor samples, from 14 types of cancer, contained
hundreds of APOBEC-signature mutations, constituting ≤68% of
mutations in the exome. In at least six distinct types of cancer,
similar results have revealed that A3B is upregulated, and its
preferred target sequence was frequently mutated and clustered
(91).
AID is critical in class-switch recombination and
SHM of the immunoglobulin gene in B lymphocytes (92). Animal models have demonstrated that
the aberrant expression of AID contributes to HCC tumorigenesis
(93,94). A positive correlation between
somatic mutation frequencies and AID expression levels was observed
in the p53 gene in liver tissues with AID upregulation, as
downregulation and somatic mutations in the p53 gene have been well
characterized in human hepatocarcinogenesis (46). These data indicate that
AID/APOBEC-catalyzed deamination may be important in generating
somatic mutations during the progression of virus-associated
HCC.
Transgenic overexpression of A1 in rabbit and mouse
livers resulted in liver dysplasia and HCC through excessive
editing of hepatic mRNAs, such as tyrosine kinase (95). Furthermore, hyperediting of the
novel A1 target no 1 mRNA that encoded a tumor suppressor gene,
created stop codons and truncated protein products, which are
linked to liver cancer (96).
Aberrant A2 expression resulted in nucleotide alterations in the
transcripts of a specific target gene and may be involved in the
development of human HCC via hepatic inflammation (97). A3 deaminases generate C-terminally
truncated HBx mutants, which enhance the colony-forming
ability and proliferative capacity of neoplastic cells. Notably,
A3B was observed to be widely upregulated in HCC tissues, and
promoted the growth of neoplastic human HepG2 liver cells and
upregulated heat shock transcription factor 1 expression levels
(37). The expression of A3A may
lead to induction of DNA breaks and activation of damage responses
in a deaminase-dependent manner. Consistent with the
above-mentioned observations, A3A expression was found to affect
genomic integrity by inducing cell-cycle arrest (98). The outcomes of a selection of
studies are summarized in Table
III.
| Table IIISummary of linkage studies between
HCC and AID/APOBECs. |
Table III
Summary of linkage studies between
HCC and AID/APOBECs.
| Protein | Experimental
targets | Indication | Reference |
|---|
| A1 | TM, TR | All of the TM and
one TR had liver dysplasia. 8/35 TM developed HCC. | 95 |
| A1 | TM, TR | The aberrant
editing markedly reduced levels of protein expression by the tumor
suppressor gene, NAT1. | 96 |
| A2 | TM | HCC developed in
2/20 A2 TM at 72 weeks of age. Significantly high frequencies of
nucleotide alterations in EIF4G2 and PTEN genes were
observed in hepatocytes. | 97 |
| A3 | CL, HCCT | C-terminally
truncated HBx mutants generated by A3 enhanced the colony
forming ability and proliferative capacity of neoplastic cells. A3B
upregulated HSF1. | 37 |
| A3A | CL | A3A led to
induction of cellular DNA breaks and activation of damage responses
in a deaminase-dependent manner. A3A expression induced cell cycle
arrest. | 98 |
| AID | CL, HCCT | The majority of
liver tissues with AID upregulation contained multiple genetic
changes in the p53 gene. Aberrant activation of AID in hepatocytes
resulted in accumulation of multiple genetic alterations in the p53
gene. | 46 |
| AID | TM | HCC developed in
27% of tissue-nonspecific alkaline phosphatase-AID TM at the age of
90 weeks. The HCC expressed α-fetoprotein and possessed deleterious
mutations in the tumor suppressor gene, TRP53. | 94 |
8. Conclusion
Innate immunity mechanisms are the first line of
defense against invading viruses. The AID/APOBEC family of cytidine
deaminases is significant in the regulation of tissue development,
responding to metabolic regulation and facilitating innate immunity
by restricting numerous types of virus, including HBV. Unresolved
issues are the mechanisms of AID/APOBEC-dependent specific
recognition of HBV DNA, degradation of cccDNA, and the security and
availability of experimental models, which are required for further
investigation. Recent analyses of the mutations have indicated that
AID/APOBEC cytidine deaminases are significant factors in the
mutagenesis of human cancer genomes. A3B, which is only localized
to the nucleus, is proposed to be responsible for a large
proportion of dispersed and clustered mutations in multiple
distinct cancers, including HCC. It is essential to elucidate the
mutational processes underlying the development of cancer, and its
potential implications on cancer etiology, prevention and
therapy.
Abbreviations:
|
AID
|
activation-induced cytidine
deaminase
|
|
APOBEC
|
apolipoprotein B mRNA editing enzyme,
catalytic polypeptide-like
|
|
HCC
|
hepatocellular carcinoma
|
|
cccDNA
|
covalently closed circular DNA
|
|
ApoB48
|
apolipoprotein B
|
|
IFN-α
|
interferon-α
|
|
PHH
|
primary human hepatocytes
|
|
ssDNA
|
single stranded DNA
|
|
SHM
|
somatic hypermutation
|
|
HMM
|
high molecular mass
|
|
rcDNA
|
relaxed circular DNA
|
|
pgRNA
|
pregenomic RNA
|
|
UNG
|
uracil DNA glycosylases
|
|
RT
|
reverse transcriptase
|
|
FRET
|
fluorescence resonance energy
transfer
|
|
CH
|
chronic hepatitis
|
References
|
1
|
Gerlich WH: Medical virology of hepatitis
B: How it began and where we are now. Virol J. 10:2392013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zoulim F: Hepatitis B virus resistance to
antiviral drugs: Where are we going? Liver Int. 31(Suppl 1):
111–116. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wursthorn K, Lutgehetmann M, Dandri M,
Volz T, Buggisch P, Zollner B, Longerich T, Schirmacher P, Metzler
F, Zankel M, et al: Peginterferon alpha-2b plus adefovir induce
strong cccDNA decline and HBsAg reduction in patients with chronic
hepatitis B. Hepatology. 44:675–684. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lucifora J, Xia Y, Reisinger F, Zhang K,
Stadler D, Cheng X, Sprinzl MF, Koppensteiner H, Makowska Z, Volz
T, et al: Specific and nonhepatotoxic degradation of nuclear
hepatitis B virus cccDNA. Science. 343:1221–1228. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wedekind JE, Dance GS, Sowden MP and Smith
HC: Messenger RNA editing in mammals: New members of the APOBEC
family seeking roles in the family business. Trends Genet.
19:207–216. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Deng Y, Du Y, Zhang Q, Han X and Cao G:
Human cytidine deaminases facilitate hepatitis B virus evolution
and link inflammation and hepatocellular carcinoma. Cancer Lett.
343:161–171. 2014. View Article : Google Scholar
|
|
7
|
Dickerson SK, Market E, Besmer E and
Papavasiliou FN: AID mediates hypermutation by deaminating single
stranded DNA. J Exp Med. 197:1291–1296. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rogozin IB, Basu MK, Jordan IK, Pavlov YI
and Koonin EV: APOBEC4, a new member of the AID/APOBEC family of
polynucleotide (deoxy) cytidine deaminases predicted by
computational analysis. Cell Cycle. 4:1281–1285. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liao W, Hong SH, Chan BH, Rudolph FB,
Clark SC and Chan L: APOBEC-2, a cardiac- and skeletal
muscle-specific member of the cytidine deaminase supergene family.
Biochem Biophys Res Commun. 260:398–404. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Teng B, Burant CF and Davidson NO:
Molecular cloning of an apolipoprotein B messenger RNA editing
protein. Science. 260:1816–1819. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jarmuz A, Chester A, Bayliss J, Gisbourne
J, Dunham I, Scott J and Navaratnam N: An anthropoid-specific locus
of orphan C to U RNA-editing enzymes on chromosome 22. Genomics.
79:285–296. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Conticello SG, Thomas CJ, Petersen-Mahrt
SK and Neuberger MS: Evolution of the AID/APOBEC family of
polynucleotide (deoxy) cytidine deaminases. Mol Biol Evol.
22:367–377. 2005. View Article : Google Scholar
|
|
13
|
Vieira VC and Soares MA: The role of
cytidine deaminases on innate immune responses against human viral
infections. Biomed Res Int. 2013:6830952013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Severi F, Chicca A and Conticello SG:
Analysis of reptilian APOBEC1 suggests that RNA editing may not be
its ancestral function. Mol Biol Evol. 28:1125–1129. 2011.
View Article : Google Scholar
|
|
15
|
Rösler C, Köck J, Kann M, Malim MH, Blum
HE, Baumert TF and von Weizsäcker F: APOBEC-mediated interference
with hepadnavirus production. Hepatology. 42:301–309. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suspène R, Guétard D, Henry M, Sommer P,
Wain-Hobson S and Vartanian JP: Extensive editing of both hepatitis
B virus DNA strands by APOBEC3 cytidine deaminases in vitro and in
vivo. Proc Natl Acad Sci USA. 102:8321–8326. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Henry M, Guétard D, Suspène R, Rusniok C,
Wain-Hobson S and Vartanian JP: Genetic editing of HBV DNA by
monodomain human APOBEC3 cytidine deaminases and the recombinant
nature of APOBEC3 G. PLoS One. 4:e42772009. View Article : Google Scholar
|
|
18
|
Baumert TF, Rösler C, Malim MH and von
Weizsäcker F: Hepatitis B virus DNA is subject to extensive editing
by the human deaminase APOBEC3C. Hepatology. 46:682–689. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gonzalez MC, Suspène R, Henry M, Guétard
D, Wain-Hobson S and Vartanian JP: Human APOBEC1 cytidine deaminase
edits HBV DNA. Retrovirology. 6:962009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Köck J and Blum HE: Hypermutation of
hepatitis B virus genomes by APOBEC3 G, APOBEC3C and APOBEC3H. J
Gen Virol. 89:1184–1191. 2008. View Article : Google Scholar
|
|
21
|
Vartanian JP, Henry M, Marchio A, Suspène
R, Aynaud MM, Guétard D, Cervantes-Gonzalez M, Battiston C,
Mazzaferro V, Pineau P, et al: Massive APOBEC3 editing of hepatitis
B viral DNA in cirrhosis. PLoS Pathog. 6:e10009282010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li D, Liu J, Kang F, Guan W, Gao X, Wang Y
and Sun D: Core-APOBEC3C chimerical protein inhibits hepatitis B
virus replication. J Biochem. 150:371–374. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang W, Zhang X, Tian C, Wang T, Sarkis
PT, Fang Y, Zheng S, Yu XF and Xu R: Cytidine deaminase APOBEC3B
interacts with heterogeneous nuclear ribonucleoprotein K and
suppresses hepatitis B virus expression. Cell Microbiol.
10:112–121. 2008.
|
|
24
|
Bonvin M and Greeve J: Effects of point
mutations in the cytidine deaminase domains of APOBEC3B on
replication and hypermu-tation of hepatitis B virus in vitro. J Gen
Virol. 88:3270–3274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jost S, Turelli P, Mangeat B, Protzer U
and Trono D: Induction of antiviral cytidine deaminases does not
explain the inhibition of hepatitis B virus replication by
interferons. J Virol. 81:10588–10596. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bonvin M, Achermann F, Greeve I, Stroka D,
Keogh A, Inderbitzin D, Candinas D, Sommer P, Wain-Hobson S,
Vartanian JP and Greeve J: Interferon-inducible expression of
APOBEC3 editing enzymes in human hepatocytes and inhibition of
hepatitis B virus replication. Hepatology. 43:1364–1374. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nguyen DH, Gummuluru S and Hu J:
Deamination-independent inhibition of hepatitis B virus reverse
transcription by APOBEC3G. J Virol. 81:4465–4472. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang T, Cai J, Chang J, Yu D, Wu C, Yan
T, Zhai K, Bi X, Zhao H, Xu J, et al: Evidence of associations of
APOBEC3B gene deletion with susceptibility to persistent HBV
infection and hepatocellular carcinoma. Hum Mol Genet.
22:1262–1269. 2013. View Article : Google Scholar
|
|
29
|
Bransteitter R, Pham P, Scharff MD and
Goodman MF: Activation-induced cytidine deaminase deaminates
deoxy-cytidine on single-stranded DNA but requires the action of
RNase. Proc Natl Acad Sci USA. 100:4102–4107. 2003. View Article : Google Scholar
|
|
30
|
Muramatsu M, Sankaranand VS, Anant S,
Sugai M, Kinoshita K, Davidson NO and Honjo T: Specific expression
of activation-induced cytidine deaminase (AID), a novel member of
the RNA-editing deaminase family in germinal center B cells. J Biol
Chem. 274:18470–18476. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vonica A, Rosa A, Arduini BL and Brivanlou
AH: APOBEC2, a selective inhibitor of TGFβ signaling, regulates
left-right axis specification during early embryogenesis. Dev Biol.
350:13–23. 2011. View Article : Google Scholar :
|
|
32
|
Sato Y, Probst HC, Tatsumi R, Ikeuchi Y,
Neuberger MS and Rada C: Deficiency in APOBEC2 leads to a shift in
muscle fiber type, diminished body mass and myopathy. J Biol Chem.
285:7111–7118. 2010. View Article : Google Scholar :
|
|
33
|
Etard C, Roostalu U and Strahle U: Lack of
Apobec2-related proteins causes a dystrophic muscle phenotype in
zebrafish embryos. J Cell Biol. 189:527–539. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Koning FA, Newman EN, Kim EY, Kunstman KJ,
Wolinsky SM and Malim MH: Defining APOBEC3 expression patterns in
human tissues and hematopoietic cell subsets. J Virol.
83:9474–9485. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tanaka Y, Marusawa H, Seno H, Matsumoto Y,
Ueda Y, Kodama Y, Endo Y, Yamauchi J, Matsumoto T, Takaori-Kondo A,
et al: Anti-viral protein APOBEC3G is induced by interferon-alpha
stimulation in human hepatocytes. Biochem Biophys Res Commun.
341:314–319. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Refsland EW, Stenglein MD, Shindo K, Albin
JS, Brown WL and Harris RS: Quantitative profiling of the full
APOBEC3 mRNA repertoire in lymphocytes and tissues: Implications
for HIV-1 restriction. Nucleic Acids Res. 38:4274–4284. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu R, Zhang X, Zhang W, Fang Y, Zheng S
and Yu XF: Association of human APOBEC3 cytidine deaminases with
the generation of hepatitis virus B x antigen mutants and
hepatocellular carcinoma. Hepatology. 46:1810–1820. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Stenglein MD, Burns MB, Li M, Lengyel J
and Harris RS: APOBEC3 proteins mediate the clearance of foreign
DNA from human cells. Nat Struct Mol Biol. 17:222–229. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peng G, Lei KJ, Jin W, Greenwell-Wild T
and Wahl SM: Induction of APOBEC3 family proteins, a defensive
maneuver underlying interferon-induced anti-HIV-1 activity. J Exp
Med. 203:41–46. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang FX, Huang J and Zhang H, Ma X and
Zhang H: APOBEC3G upregulation by alpha interferon restricts human
immunodeficiency virus type 1 infection in human peripheral
plasmacytoid dendritic cells. J Gen Virol. 89:722–730. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Stopak KS, Chiu YL, Kropp J, Grant RM and
Greene WC: Distinct patterns of cytokine regulation of APOBEC3G
expression and activity in primary lymphocytes, macrophages and
dendritic cells. J Biol Chem. 282:3539–3546. 2007. View Article : Google Scholar
|
|
42
|
Argyris EG, Acheampong E, Wang F, Huang J,
Chen K, Mukhtar M and Zhang H: The interferon-induced expression of
APOBEC3G in human blood-brain barrier exerts a potent intrinsic
immunity to block HIV-1 entry to central nervous system. Virology.
367:440–451. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Endo Y, Marusawa H, Kinoshita K, Morisawa
T, Sakurai T, Okazaki IM, Watashi K, Shimotohno K, Honjo T and
Chiba T: Expression of activation-induced cytidine deaminase in
human hepatocytes via NF-kappaB signaling. Oncogene. 26:5587–5595.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Matsumoto T, Marusawa H, Endo Y, Ueda Y,
Matsumoto Y and Chiba T: Expression of APOBEC2 is transcriptionally
regulated by NF-kappaB in human hepatocytes. FEBS Lett.
580:731–735. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Endo Y, Marusawa H, Kou T, Nakase H, Fujii
S, Fujimori T, Kinoshita K, Honjo T and Chiba T: Activation-induced
cytidine deaminase links between inflammation and the development
of colitis-associated colorectal cancers. Gastroenterology.
135:889–898. 898 e1–e3. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kou T, Marusawa H, Kinoshita K, Endo Y,
Okazaki IM, Ueda Y, Kodama Y, Haga H, Ikai I and Chiba T:
Expression of activation-induced cytidine deaminase in human
hepatocytes during hepatocarcinogenesis. Int J Cancer. 120:469–476.
2007. View Article : Google Scholar
|
|
47
|
Smith HC, Bennett RP, Kizilyer A,
McDougall WM and Prohaska KM: Functions and regulation of the
APOBEC family of proteins. Semin Cell Dev Biol. 23:258–268. 2012.
View Article : Google Scholar
|
|
48
|
Backus JW, Schock D and Smith HC: Only
cytidines 5′ of the apolipoprotein B mRNA mooring sequence are
edited. Biochim Biophys Acta. 1219:1–14. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yang Y, Sowden MP and Smith HC: Induction
of cytidine to uridine editing on cytoplasmic apolipoprotein B mRNA
by overexpressing APOBEC-1. J Biol Chem. 275:22663–22669. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lau PP, Xiong WJ, Zhu HJ, Chen SH and Chan
L: Apolipoprotein B mRNA editing is an intranuclear event that
occurs posttranscriptionally coincident with splicing and
polyadenylation. J Biol Chem. 266:20550–20554. 1991.PubMed/NCBI
|
|
51
|
Sowden MP and Smith HC: Commitment of
apolipoprotein B RNA to the splicing pathway regulates
cytidine-to-uridine editing-site utilization. Biochem J.
359:697–705. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Papavasiliou FN and Schatz DG: Somatic
hypermutation of immunoglobulin genes: Merging mechanisms for
genetic diversity. Cell. 109(Suppl): S35–S44. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lada AG, Krick CF, Kozmin SG, Mayorov VI,
Karpova TS, Rogozin IB and Pavlov YI: Mutator effects and mutation
signatures of editing deaminases produced in bacteria and yeast.
Biochemistry (Mosc). 76:131–146. 2011. View Article : Google Scholar
|
|
54
|
Stavnezer J: Complex regulation and
function of activation-induced cytidine deaminase. Trends Immunol.
32:194–201. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Patenaude AM and Di Noia JM: The
mechanisms regulating the subcellular localization of AID. Nucleus.
1:325–331. 2010. View Article : Google Scholar
|
|
56
|
Bennett RP, Presnyak V, Wedekind JE and
Smith HC: Nuclear Exclusion of the HIV-1 host defense factor
APOBEC3G requires a novel cytoplasmic retention signal and is not
dependent on RNA binding. J Biol Chem. 283:7320–7327. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Smith HC: APOBEC3G: A double agent in
defense. Trends Biochem Sci. 36:239–244. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chiu YL, Soros VB, Kreisberg JF, Stopak K,
Yonemoto W and Greene WC: Cellular APOBEC3G restricts HIV-1
infection in resting CD4+ T cells. Nature. 435:108–114. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
McDougall WM and Smith HC: Direct evidence
that RNA inhibits APOBEC3G ssDNA cytidine deaminase activity.
Biochem Biophys Res Commun. 412:612–617. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Soros VB, Yonemoto W and Greene WC: Newly
synthesized APOBEC3G is incorporated into HIV virions, inhibited by
HIV RNA and subsequently activated by RNase H. PLoS Pathog.
3:e152007. View Article : Google Scholar
|
|
61
|
Wang X, Dolan PT, Dang Y and Zheng YH:
Biochemical differentiation of APOBEC3F and APOBEC3G proteins
associated with HIV-1 life cycle. J Biol Chem. 282:1585–1594. 2007.
View Article : Google Scholar :
|
|
62
|
Niewiadomska AM, Tian C, Tan L, Wang T,
Sarkis PT and Yu XF: Differential inhibition of long interspersed
element 1 by APOBEC3 does not correlate with
high-molecular-mass-complex formation or P-body association. J
Virol. 81:9577–9583. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Tan L, Sarkis PT, Wang T, Tian C and Yu
XF: Sole copy of Z2-type human cytidine deaminase APOBEC3H has
inhibitory activity against retrotransposons and HIV-1. FASEB J.
23:279–287. 2009. View Article : Google Scholar :
|
|
64
|
Gerelsaikhan T, Tavis JE and Bruss V:
Hepatitis B virus nucleocapsid envelopment does not occur without
genomic DNA synthesis. J Virol. 70:4269–4274. 1996.PubMed/NCBI
|
|
65
|
Lewin SR, Ribeiro RM, Walters T, Lau GK,
Bowden S, Locarnini S and Perelson AS: Analysis of hepatitis B
viral load decline under potent therapy: Complex decay profiles
observed. Hepatology. 34:1012–1020. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Nowak MA, Bonhoeffer S, Hill AM, Boehme R,
Thomas HC and McDade H: Viral dynamics in hepatitis B virus
infection. Proc Natl Acad Sci USA. 93:4398–4402. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Tran A, Kremsdorf D, Capel F, Housset C,
Dauguet C, Petit MA and Brechot C: Emergence of and takeover by
hepatitis B virus (HBV) with rearrangements in the pre-S/S and
pre-C/C genes during chronic HBV infection. J Virol. 65:3566–3574.
1991.PubMed/NCBI
|
|
68
|
Beggel B, Münk C, Däumer M, Hauck K,
Häussinger D, Lengauer T and Erhardt A: Full genome ultra-deep
pyrosequencing associates G-to-A hypermutation of the hepatitis B
virus genome with the natural progression of hepatitis B. J Viral
Hepat. 20:882–889. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Noguchi C, Ishino H, Tsuge M, Fujimoto Y,
Imamura M, Takahashi S and Chayama K: G to A hypermutation of
hepatitis B virus. Hepatology. 41:626–633. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Hannoun C, Horal P and Lindh M: Long-term
mutation rates in the hepatitis B virus genome. J Gen Virol.
81:75–83. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Mangeat B, Turelli P, Caron G, Friedli M,
Perrin L and Trono D: Broad antiretroviral defence by human
APOBEC3G through lethal editing of nascent reverse transcripts.
Nature. 424:99–103. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Har ris RS, Bishop KN, Sheehy AM, Craig
HM, Petersen-Mahrt SK, Watt IN, Neuberger MS and Malim MH: DNA
deamination mediates innate immunity to retroviral infection. Cell.
113:803–809. 2003. View Article : Google Scholar
|
|
73
|
Schrofelbauer B, Yu Q, Zeitlin SG and
Landau NR: Human immunodeficiency virus type 1 Vpr induces the
degradation of the UNG and SMUG uracil-DNA glycosylases. J Virol.
79:10978–10987. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Kaiser SM and Emerman M: Uracil DNA
glycosylase is dispensable for human immunodeficiency virus type 1
replication and does not contribute to the antiviral effects of the
cytidine deaminase Apobec3G. J Virol. 80:875–882. 2006. View Article : Google Scholar :
|
|
75
|
Turelli P, Mangeat B, Jost S, Vianin S and
Trono D: Inhibition of hepatitis B virus replication by APOBEC3G.
Science. 303:18292004. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lei YC, Tian YJ, Ding HH, Wang BJ, Yang Y,
Hao YH, Zhao XP, Lu MJ, Gong FL and Yang DL: N-terminal and
C-terminal cytosine deaminase domain of APOBEC3G inhibit hepatitis
B virus replication. World J Gastroenterol. 12:7488–7496.
2006.PubMed/NCBI
|
|
77
|
Liang G, Kitamura K, Wang Z, Liu G,
Chowdhury S, Fu W, Koura M, Wakae K, Honjo T and Muramatsu M: RNA
editing of hepatitis B virus transcripts by activation-induced
cytidine deaminase. Proc Natl Acad Sci USA. 110:2246–2251. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Nassal M: The arginine-rich domain of the
hepatitis B virus core protein is required for pregenome
encapsidation and productive viral positive-strand DNA synthesis
but not for virus assembly. J Virol. 66:4107–4116. 1992.PubMed/NCBI
|
|
79
|
Schlicht HJ, Bartenschlager R and Schaller
H: The duck hepatitis B virus core protein contains a highly
phosphorylated C terminus that is essential for replication but not
for RNA packaging. J Virol. 63:2995–3000. 1989.PubMed/NCBI
|
|
80
|
Nguyen DH and Hu J: Reverse transcriptase-
and RNA packaging signal-dependent incorporation of APOBEC3G into
hepatitis B virus nucleocapsids. J Virol. 82:6852–6861. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhao D, Wang X, Lou G, Peng G, Li J, Zhu
H, Chen F, Li S, Liu D, Chen Z and Yang Z: APOBEC3G directly binds
Hepatitis B virus core protein in cell and cell free systems. Virus
Res. 151:213–219. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Tuzun E, Sharp AJ, Bailey JA, Kaul R,
Morrison VA, Pertz LM, Haugen E, Hayden H, Albertson D, Pinkel D,
et al: Fine-scale structural variation of the human genome. Nat
Genet. 37:727–732. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
83
|
McCarroll SA, Hadnott TN, Perry GH, Sabeti
PC, Zody MC, Barrett JC, Dallaire S, Gabriel SB, Lee C, Daly MJ, et
al: Common deletion polymorphisms in the human genome. Nat Genet.
38:86–92. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
84
|
Kidd JM, Newman TL, Tuzun E, Kaul R and
Eichler EE: Population stratification of a common APOBEC gene
deletion polymorphism. PLoS Genet. 3:e632007. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Abe H, Ochi H, Maekawa T, Hatakeyama T,
Tsuge M, Kitamura S, Kimura T, Miki D, Mitsui F, Hiraga N, et al:
Effects of structural variations of APOBEC3A and APOBEC3B genes in
chronic hepatitis B virus infection. Hepatol Res. 39:1159–1168.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Ezzikouri S, Kitab B, Rebbani K, Marchio
A, Wain-Hobson S, Dejean A, Vartanian JP, Pineau P and Benjelloun
S: Polymorphic APOBEC3 modulates chronic hepatitis B in Moroccan
population. J Viral Hepat. 20:678–686. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wang X, Abudu A, Son S, Dang Y, Venta PJ
and Zheng YH: Analysis of human APOBEC3H haplotypes and anti-human
immunodeficiency virus type 1 activity. J Virol. 85:3142–3152.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Harari A, Ooms M, Mulder LC and Simon V:
Polymorphisms and splice variants influence the antiretroviral
activity of human APOBEC3H. J Virol. 83:295–303. 2009. View Article : Google Scholar :
|
|
89
|
Roberts SA, Lawrence MS, Klimczak LJ,
Grimm SA, Fargo D, Stojanov P, Kiezun A, Kryukov GV, Carter SL,
Saksena G, et al: An APOBEC cytidine deaminase mutagenesis pattern
is widespread in human cancers. Nat Genet. 45:970–976. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Alexandrov LB, Nik-Zainal S, Wedge DC,
Aparicio SA, Behjati S, Biankin AV, Bignell GR, Bolli N, Borg A,
Børresen-Dale AL, et al: Signatures of mutational processes in
human cancer. Nature. 500:415–421. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Burns MB, Temiz NA and Harris RS: Evidence
for APOBEC3B mutagenesis in multiple human cancers. Nat Genet.
45:977–983. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Pasqualucci L, Bhagat G, Jankovic M,
Compagno M, Smith P, Muramatsu M, Honjo T, Morse HC III,
Nussenzweig MC and Dalla-Favera R: AID is required for germinal
center-derived lymphomagenesis. Nat Genet. 40:108–112. 2008.
View Article : Google Scholar
|
|
93
|
Chiba T and Marusawa H: A novel mechanism
for inflammation-associated carcinogenesis; an important role of
activation-induced cytidine deaminase (AID) in mutation induction.
J Mol Med (Berl). 87:1023–1027. 2009. View Article : Google Scholar
|
|
94
|
Takai A, Toyoshima T, Uemura M, Kitawaki
Y, Marusawa H, Hiai H, Yamada S, Okazaki IM, Honjo T, Chiba T and
Kinoshita K: A novel mouse model of hepatocarcinogenesis triggered
by AID causing deleterious p53 mutations. Oncogene. 28:469–478.
2009. View Article : Google Scholar
|
|
95
|
Yamanaka S, Balestra ME, Ferrell LD, Fan
J, Arnold KS, Taylor S, Taylor JM and Innerarity TL: Apolipoprotein
B mRNA-editing protein induces hepatocellular carcinoma and
dysplasia in transgenic animals. Proc Natl Acad Sci USA.
92:8483–8487. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yamanaka S, Poksay KS, Arnold KS and
Innerarity TL: A novel translational repressor mRNA is edited
extensively in livers containing tumors caused by the transgene
expression of the apoB mRNA-editing enzyme. Genes Dev. 11:321–333.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Okuyama S, Marusawa H, Matsumoto T, Ueda
Y, Matsumoto Y, Endo Y, Takai A and Chiba T: Excessive activity of
apolipoprotein B mRNA editing enzyme catalytic polypeptide 2
(APOBEC2) contributes to liver and lung tumorigenesis. Int J
Cancer. 130:1294–1301. 2012. View Article : Google Scholar
|
|
98
|
Landry S, Narvaiza I, Linfesty DC and
Weitzman MD: APOBEC3A can activate the DNA damage response and
cause cell-cycle arrest. EMBO Rep. 12:444–450. 2011. View Article : Google Scholar : PubMed/NCBI
|