Introduction
Pancreatic cancer is a significant worldwide health
problem, with an estimated 277,000 new cases, and a
cancer-associated mortality rate of 266,000 annually worldwide
(1). The majority of patients with
pancreatic cancer are diagnosed at advanced stages of the disease
(2), at which tumor tissues are
surgically unresectable and chemotherapy is mostly ineffective,
leading to a 5-year survival rate of ~ 4% (3). Thus, there is a significant and
urgent requirement to elucidate the molecular mechanisms that
mediate pancreatic cancer development and progression, in order to
effectively control and prevent this life-threatening disease. The
development of pancreatic cancer, as with other types of cancer in
the gastrointestinal tract, involves multiple genetic alterations,
including oncogene activation and tumor suppressor gene
dysfunction. Over the past three decades, a large body of evidence
has accumulated regarding the molecular alterations associated with
the development of pancreatic cancer (4–6).
These disparate observations have coalesced somewhat into an
improved understanding of this disease, however, further
investigation is required to provide early cancer identification
and novel treatment strategies for pancreatic cancer.
The present study focused on Wnt signaling proteins
in pancreatic cancer. Wnt consists of a large family of secreted
lipid-modified glycoproteins, which regulate numerous processes in
animal development (7) and normal
cell processes, including cell growth, differentiation, apoptosis,
migration and cell polarity (8).
Each member of the Wnt family of proteins exhibits unique
expression patterns and has distinct functions in embryonic
development and normal cell homeostasis (9). Based on their ability to induce
transformation of the mouse mammary epithelial cell line, Wnt
proteins are divided into two functional groups, canonical and
noncanonical Wnt (10). Wnt5a is
classified as a member of the noncanonical Wnt family, is crucial
for embryonic development and has been implicated in several human
diseases (11–13). Previous studies have demonstrated
that Wnt5a is important in the progression of various types of
human cancer (14–16); however, the particular signaling
and physiological function of the Wnt5a protein in pancreatic
cancer remains to be elucidated. Our previous study identified that
Wnt5a protein is expressed at high levels in human pancreatic
cancer tissues, and that upregulation of Wnt5a promotes
epithelial-to-mesenchymal transition and metastasis of pancreatic
cancer cells (17). On the basis
of these data, the aim of the present study was to characterize the
role of Wnt5a in the proliferation of pancreatic cancer cells, and
to elucidate the signaling events of the Wnt5a protein.
Material s and methods
Cell lines and culture
Human PANC-1 and BXPC-3 pancreatic cancer cell lines
were obtained from the Institute of Cellular Research, Chinese
Academy of Science (Shanghai, China) and routinely cultured in RPMI
1640 medium supplemented with 10% fetal bovine serum, 100 U/ml
penicillin and 100 U/ml streptomycin (all from Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) at 37°C in 5%
CO2. The medium was replaced every third day.
Gene transfection into pancreatic cancer
cells
Plasmids carrying Wnt5a cDNA were constructed by the
subcloning of Wnt5a cDNA into a pcDNA3.1 vector (Invitrogen; Thermo
Fisher Scientific, Inc.) with double restriction enzyme
(XbaI and BamHI; Invitrogen; Thermo Fisher
Scientific, Inc.) digestion. Subsequently, the pcDNA3.1-WNT5A
vector or empty pcDNA3.1 vector was transfected into the PANC-1 and
BXPC-3 cells (60–80% confluent) using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. To establish cells stably overexpressing
Wnt5a, the cells were selected in G418-containing medium (1,000
µl/ml for PANC-1 and 400 µl/ml for BXPC-3 cells;
Invitrogen; Thermo Fisher Scientific, Inc.). Following 2–3 weeks of
G418 selection (37°C in 5% CO2), single cell clones were
isolated, expanded and screened for the expression of Wnt5a using
immunoblot analysis.
To knock down endogenous expression of Wnt5a in the
pancreatic cancer cells, the present study utilized small
interfering RNA (siRNA) technology, using the target sequence,
5′-GTT TTG GCC ACT GAC TGA-3′. The Wnt5a siRNA expression cassette
(FuNeng Biotech Co., Ltd., Guangzhou, China) was subcloned into a
pcDNA6.2 vector (Invitrogen; Thermo Fisher Scientifc, Inc.),
following which the pcDNA6.2-WNT5A-siRNA vector or the pcDNA6.2
control vector was transfected into the PANC-1 and BXPC-3 cells
(60–80% confluent) using Lipofectamine 2000, according to the
manufacturer's protocol. Blasticidin (Merck Millipore GmbH,
Darmstadt, Germany) was used to screen resistant cell clones (4
µg/ml for PANC-1 and 2 µg/ml for BXPC-3 cells).
Stable clones were confirmed by immunoblot analysis.
In addition, β-catenin and control siRNA were
obtained from GenePharma Company (Shanghai, China) to knock down
the expression of β-catenin. The β-catenin or control siRNA (20 nM)
was then transfected into the pancreatic cancer cells and, after 24
h culture at 37°C in 5% CO2, 200 ng of recombinant human
Wnt5a peptide (Abcam) was added to the cell culture. The expression
levels of nuclear β-catenin were detected by immunoblot
analysis.
Western blot analysis
The cells were lysed in lysis buffer (Invitrogen;
Thermo Fisher Scientific, Inc.), and the proteins were separated on
10% SDS-PAGE (Shenggong Biotech Co., Ltd., Shanghai, China) prior
to being transferred onto 0.45 µm pore polyvi nylidene
difluoride membranes (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The membranes were probed with the following primary
antibodies: Rabbit polyclonal anti-Wnt5a (1:1,000; cat. no.
ab72583; Abcam, Cambridge, MA, USA), mouse monoclonal
anti-glyceral-dehyde-3-phosphate dehydrogenase (GAPDH; 1:3,000;
cat. no. KC-5G4; Kangchen Biotech, Inc., Shanghai, China), rabbit
monoclonal anti-β-catenin (1:1,000; cat. no. ab32572; Abcam) and
rabbit polyclonal anti-lamin A/C (1:1,000; cat. no. sc-20681; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA). Subsequently, the
membrane was washed once in 1X TBS (1X Tris-buffered saline: 50 mM
Tris-HCl, pH 7.4; 150 mM NaCl; Shenggong Biotech Co., Ltd.) and
three times in 1X TBS-T buffer (1X TBS containing 0.1% Tween 20;
Shenggong Biotech Co., Ltd.). The goat polyclonal anti-rabbit
(1:5,000; cat. no. ab6720; Abcam) or goat monoclonal anti mouse
(1:5,000; cat. no.ab197767; Abcam) secondary antibodies were added
in blocking solution (3% nonfat dry milk in TBS-T buffer) and the
membranes were shaken for 2 h at room temperature. Reactive
proteins were detected using enhanced chemiluminescence (GE
Healthcare Bio-Sciences, Pittsburgh, PA, USA). The intensity of
each band was measured by densitometric analysis using the Quantity
One software (Bio-Rad Laboratories, Inc.).
Cell viability assay
The stable PANC-1 and BXPC-3 cells exhibiting Wnt5a
overexpression or knockdown were seeded into a 96-well plate at
1×103 cells/well and maintained in G418-conditioned
medium, with eight replicate wells used for each group. After 24,
48, 72, 96, 120, 144 and 168 h of cell culture at 37°C in 5%
CO2, 400 µg/ml of
methylthiazolyldiphenyl-tetrazolium bromide (MTT; Sigma-Aldrich,
St. Louis, MO, USA) was added to each well, and the cells were
incubated for an additional 4 h at 37°C. Subsequently,
solubilization was performed using 150 µl dimethyl sulfoxide
(DMSO; Tiangen Biotech Co., Ltd., Beijing, China) and incubated at
37°C in 5% CO2 for 20 min. The optical density was then
measured using a spectrophotometer (Ultrospec K; Biochrom, Ltd.,
Berlin, Germany) at a wavelength of 570 nm. The optical density
values of each experiment were plotted to determine the cell
viability curve. The experiment was repeated three times.
Tumor cell colony formation assay
For the clonogenic assay, the BXPC-3 and PANC-1
cells exhibiting Wnt5a overexpression or knockdown were plated at a
density of 1,000 viable cells/well in six-well plates and cultured
for 14 days in a 37°C incubator. The cells were then washed with
phosphate buffered saline (PBS; Invitrogen; Thermo Fisher
Scientific, Inc.), fixed with a mixture of methanol (MingRui
Biotechnology Co., Ltd., Shanghai, China) and acetic acid (1:1
ratio; MingRui Biotechnology Co., Ltd.) and then stained with 1%
crystal violet (MingRui Biotechnology Co., Ltd.). The colonies
containing >50 cells were counted under an inverted microscope
(DMI3000B; Leica Microsystems GmbH, Wetzlar, Germany). The assay
was performed in triplicate and repeated once.
Flow cytometric analysis of
apoptosis
Tumor cell apoptosis was detected using flow
cytometry using an Annexin V-fluorescein isothiocyanate (FITC)
apoptosis detection kit (cat. no. MR-KA020) from MingRui
Biotechnology Co. Ltd., according to the manufacturer's protocol.
Briefly, near-80% confluent PANC-1 and BXPC-3 cells were harvested
using 0.25% trypsin (MingRui Biotechnology Co., Ltd.), washed twice
in PBS and resuspended at a density of 1×106 viable
cells/ml in Annexin V binding buffer (MingRui Biotechnology Co.,
Ltd.). Subsequently, 5 ml Annexin V/FITC, at a final concentration
of 50 µg/ml, was added to the cell suspension with 10
µl propidium iodide, at a final concentration of 1
µg/ml. The cell solution was incubated for 15 min at room
temperature in the dark. Following incubation, the stained cells
were immediately analyzed using a flow cytometer (FACSCalibur;
Bio-Rad Laboratories, Inc.). Each experiment was performed in
triplicate and repeated at least three times.
Animal experiments
A total of 12 nude mice (nu/nu-nuBR), aged 4 weeks
old and weighing 14–16 g, were obtained from the Shanghai
Laboratory Animal Center, Chinese Academy of Science. The animals
were housed in a specific pathogen-free environment with a constant
temperature between 25 and 27°C in a 12/12 h light/dark cycle. The
mice were fed with a standard pellet diet and water ad
libitum. The present study was approved by the ethics committee
of The Second Military Medical University (Shanghai, China) and was
performed in compliance with the regulations of the Administration
of Experimental Animals of the State Council in China.
The nude mice were anesthetized using 1% sodium
pentobarbital (Shenggong Biotech Co., Ltd.) and surgery was
performed to expose the pancreas and spleen. A total of
5×107 pancreatic cancer, PANC-1 or BXPC-3 cells,
transfected with Wnt5a-overexpressing or empty vector, were
injected into the pancreatic parenchyma, close to the hilum of the
spleen. Subsequently, the pancreas was relocated into the abdominal
cavity, which was then closed in two layers using 6–0 absorbable
vicryl sutures [Anhui Kangning Industrial (Group) Co., Ltd,
Shanghai, China]. The animals were divided into 4 groups,
PANC-1-Wnt5a-overexpressing, PANC-1-mock control,
BXPC-3-Wnt5a-overexpressing and BXPC-3-mock control, according to
the different cells injected. Each group consisted of three
animals. When the orthotopic tumors reached a size of 1–2 cm in
diameter, the mice were sacrificed by exposure to CO2
followed by cervical dislocation. Images of the pancreatic tissues
with tumor lesions were captured using a DSC-HX50 Sony camera (Sony
Corporation, Tokyo, Japan), and the xenografts were resected, fixed
in formalin (Shenggong Biotech Co., Ltd.), embedded in paraffin
(Shenggong Biotech Co., Ltd.) and cut into 0.4 µm tissue
sections. A streptavidin-peroxidase-biotin immunohistochemical
method was used for immunostaining. Briefly, tissue sections were
deparaffinized and hydrated in xylene (ShengGong Biotechnology Co.,
Ltd.) and serial alcohol solutions (ShengGong Biotechnology Co.,
Ltd.). Endogenous peroxidase was blocked by incubation in 3%
H2O2 (ShengGong Biotechnology Co., Ltd.) for
15 min at room temperature. The antigen retrieval step was
performed in a steam pressure cooker containing preheated 0.01 M
citrate buffer (pH 6.0; Dako, Glostrup, Denmark) at 95°C for 15
min. Rabbit polyclonal anti-Wnt5a antibody (cat. no. ab72583;
Abcam; 1:50) and rabbit polyclonal anti-Ki67 antibody (cat. no.
ab15580; Abcam; 1:100) were incubated with the tissue sections for
2 h at room temperature. The tissue sections were then treated with
biotinylated polyclonal anti-rabbit secondary antibodies (cat. no.
ab6720; Abcam; 1:1,000), followed by incubation with
streptavidin-horseradish peroxidase complex (Santa Cruz
Biotechnology, Inc.) for 30 min at room temperature.
Immunoreactivity was visualized with diaminobenzidine
(Sigma-Aldrich). The tissue sections were then counter-stained with
hematoxylin (ShengGong Biotechnology Co., Ltd.). Wnt5a staining was
predominantly located in the cell cytoplasm and nuclei, whereas
Ki67 staining in the nuclei. The proportion of positive tumor cells
was scored.
Statistical analysis
Statistical analyses were performed using the SPSS
17 software package (SPSS, Inc., Chicago, IL, USA). Differences in
the means were determined using Student's t-test or one-way
analysis of variance followed by Tukey's post-hoc test. P≤0.05 was
considered to indicate a statistically significant difference.
Results
Wnt5a induces pancreatic cancer cell
viability and colony formation
Stable Wnt5a-overexpressing or Wnt5a
siRNA-expressing pancreatic cancer cell lines were established
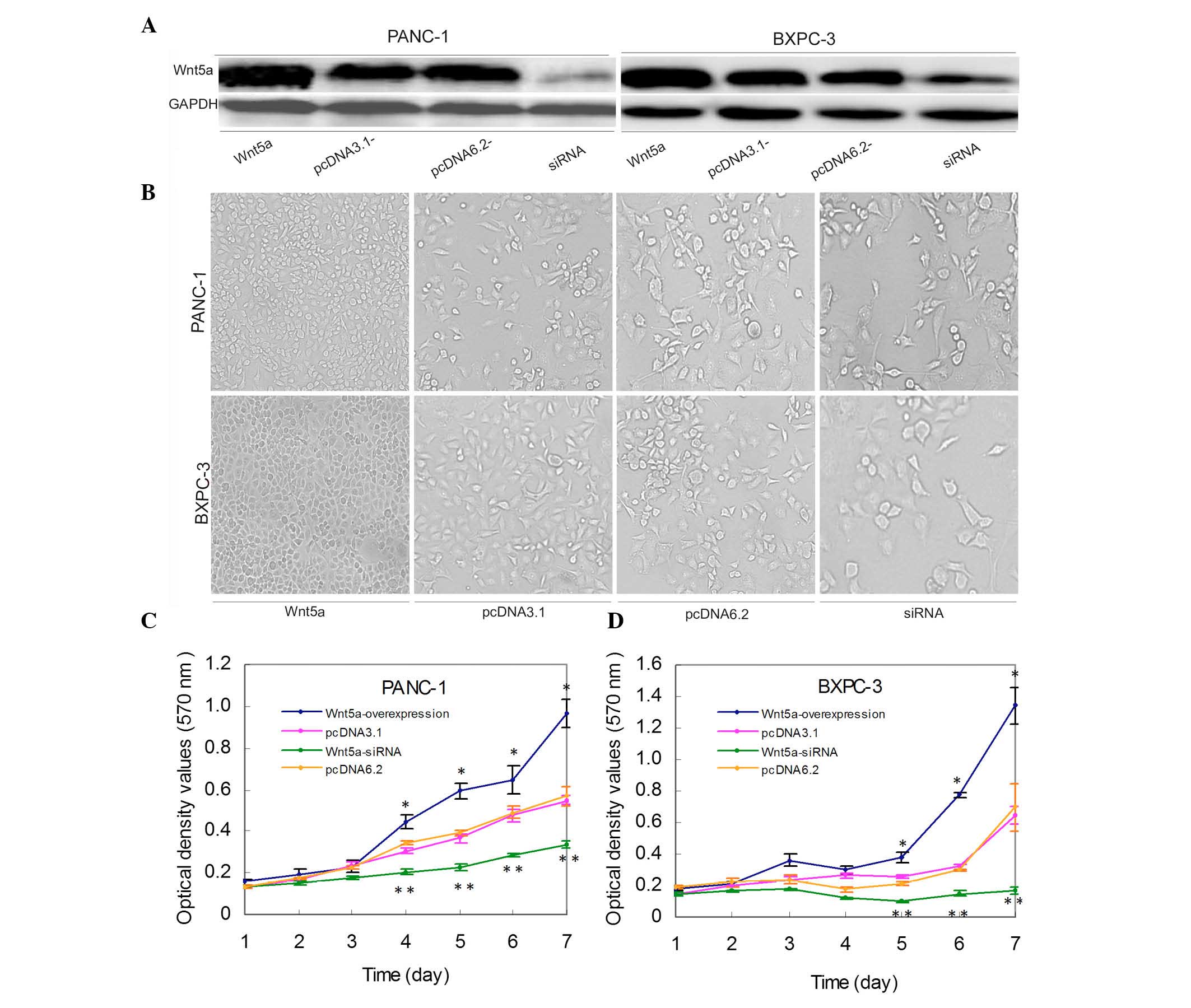
following gene transfection and confirmed using immunoblot analysis
(Fig. 1A). A subsequent cell
viability assay showed that the cell density of PANC-1 and BXPC-3
cells was significantly different among the groups following seven
days of culture (Fig. 1B).
Statistical data confirmed that the numbers of viable cells were
significantly higher in the Wnt5a-overexpressing cells, compared
with that in the control cells (P<0.05; Fig. 1B–D). By contrast, siRNA-mediated
silencing of the expression of Wnt5a significantly reduced cell
viability, compared with the control (P<0.05; Fig. 1B–D).
Furthermore, as shown in Fig. 2A, the Wnt5a-overexpressing PANC-1
and BXPC-3 cells formed an increased number of colonies, compared
with the control cells (211.9 and 180.7%, respectively; P<0.05;
Fig. 2B and C). The siRNA
transfected Wnt5a PANC-1 and BXPC-3 cells exhibited a significant
decrease in colony formation, compared with the control cells
(32.8% for PANC-1 and 35.77% for BXPC-3; P<0.05; Fig. 2B and C).
Overexpression of Wnt5a inhibits tumor
cell apoptosis
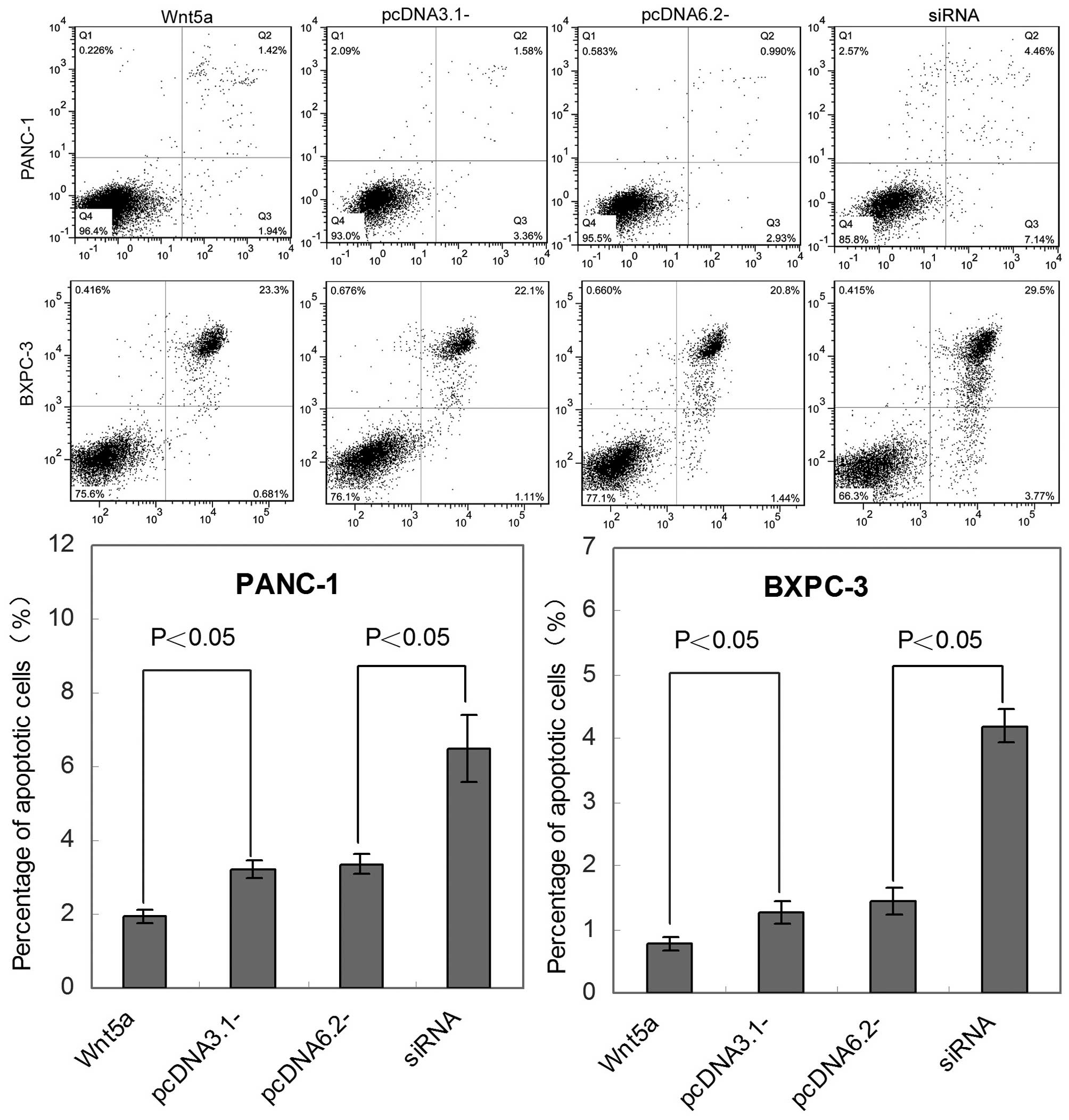
An Annexin V-FITC kit was used to further assess the
effect of Wnt5a on pancreatic cancer apoptosis in vivo.
Cells stained with Annexin V+/PI− (in Q3) are
apoptotic. Compared with the control, the levels of apoptosis were
significantly decreased in the Wnt5a-overexpressing PANC-1 and
BXPC-3 cell lines (P<0.05), whereas an elevated rate of tumor
cell apoptosis occurred in the siRNA-transfected tumor cells
(Fig. 3).
Wnt5a induces pancreatic cancer cell
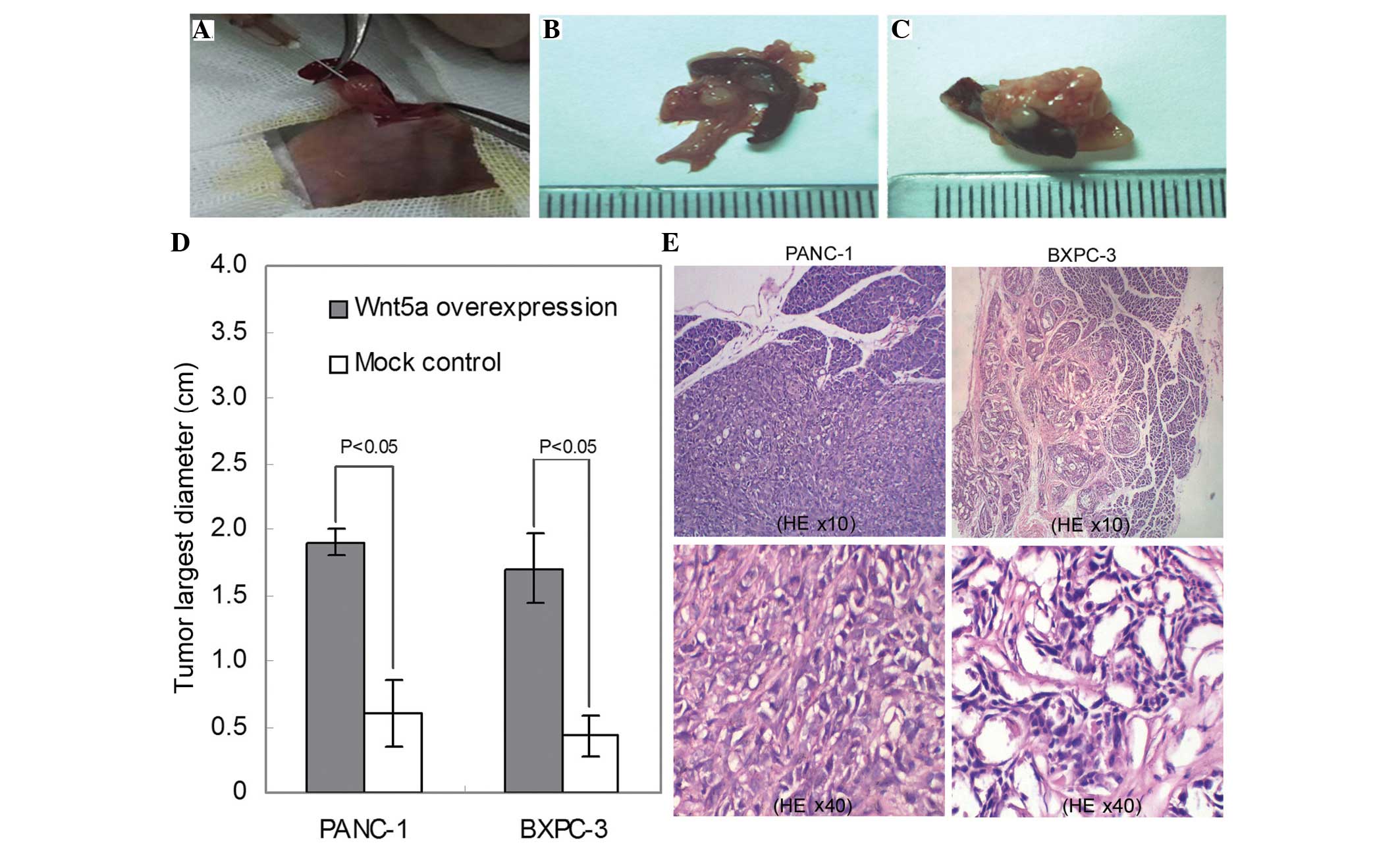
xenograft growth in an orthotopic nude mouse model
Following the in vitro investigations, the
present study investigated the in vivo effects of Wnt5a on
tumor growth using an orthotopic mouse model. The PANC-1 and BXPC-3
cells overexpressing Wnt5a were injected orthotopically into the
pancreatic parenchyma, and empty vector-transfected cells were used
as a control (Fig. 4A). The data
showed that, consistent with the results of the in vitro
assays, the average tumor size of the Wnt5a-overexpressing group
was significantly larger, compared with that of the control group
(P<0.05; Fig. 4B–D).
Microscopic examination showed that PANC-1 and BXPC-3 cells
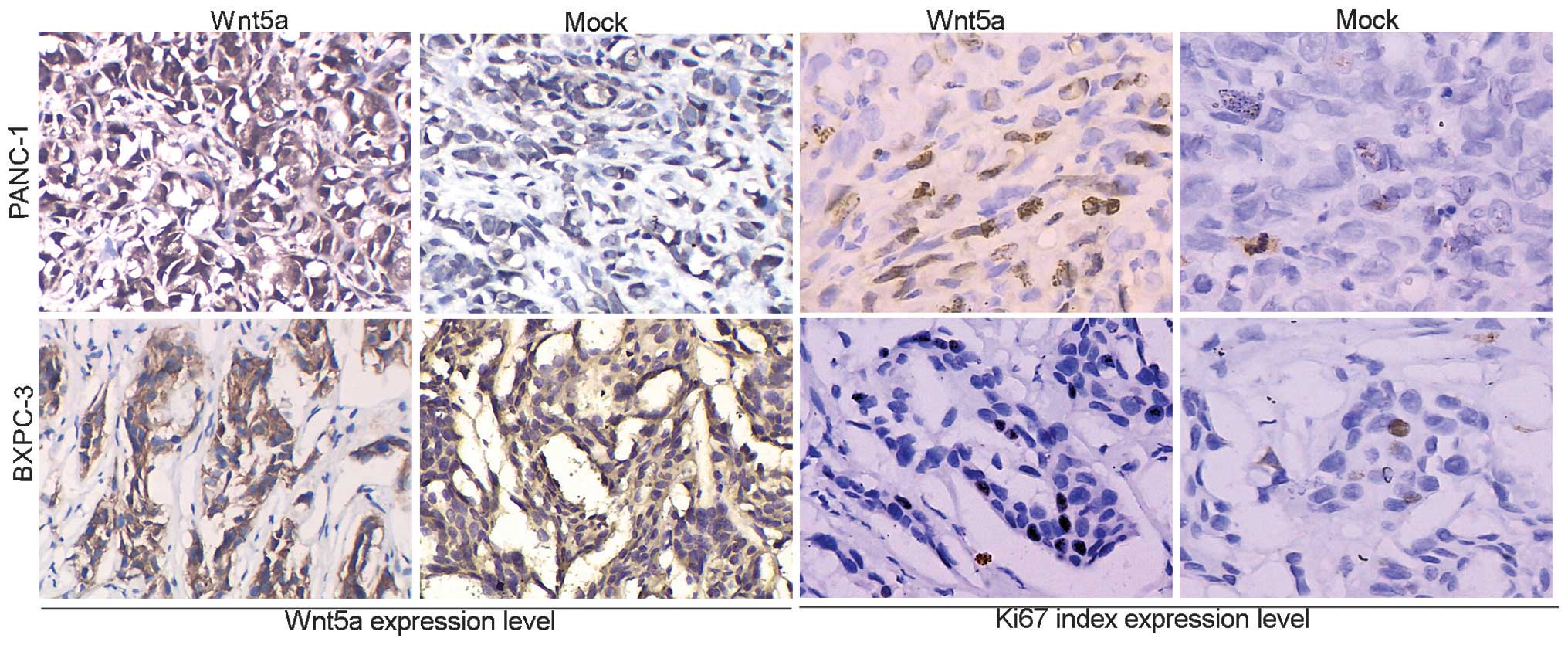
exhibited different histological morphologies (Fig. 4E). In addition, microscopic
examination and immunohistochemistry analysis demonstrated that the
Wnt5a-overexpressing tumors exhibited increased protein expression
levels of Ki67 (Fig. 5).
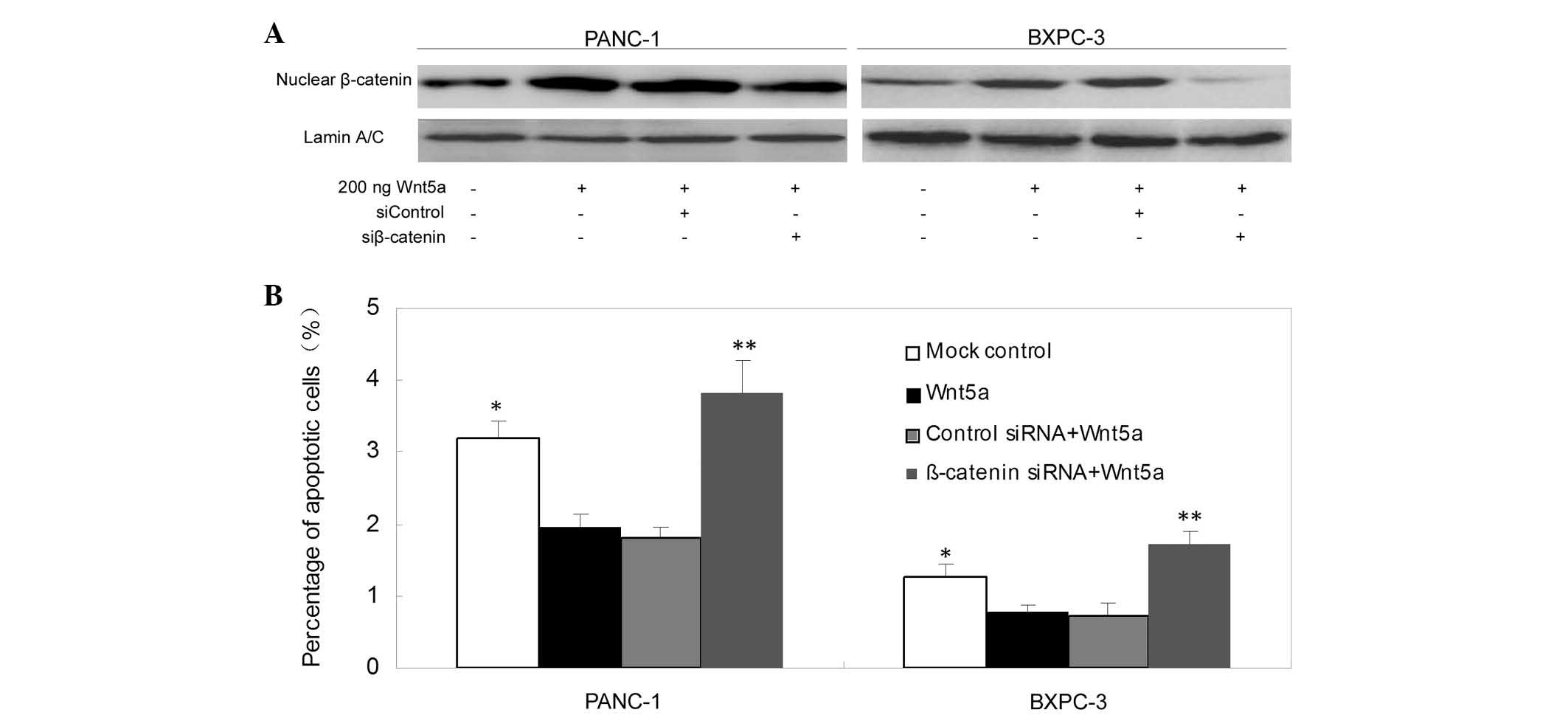
β-catenin mediates the effects of Wnt5a
on pancreatic cancer cells
To examine the molecular mechanisms mediating the
effects of Wnt5a overexpression in pancreatic cancer cells, the
present study pre-transfected β-catenin siRNA into pancreatic
cancer cells, and then treated the cells with a recombinant Wnt5a
peptide (200 ng/well). The data showed that the depletion of
β-catenin reversed the effects of Wnt5a overexpression on the
levels of apoptosis in the PANC-1 and BXPC-3 cells (Fig. 6A and B).
Discussion
Previous studies have demonstrated that Wnt5a can
modulate various cell activities, including cell migration
(18), adhesion (19), invasion (20) and differentiation (21). Specifically, Wnt5a promotes the
progression of prostate and lung cancer cells (22,23),
however it is able to suppress tumor progression in thyroid
carcinoma cells (24). In the
present study, the effects of Wnt5a overexpression and knockdown on
the proliferation of pancreatic cancer cells were examined. The
results demonstrated that overexpression of the Wnt5a protein
induced tumor cell viability and clonogenicity, but inhibited
apoptosis in the two pancreatic cancer cell lines in vitro,
and increased tumor growth in an orthotopic nude mouse model. By
contrast, knockdown of the expression of Wnt5a reduced tumor cell
viability and promoted apoptosis. At a molecular level, β-catenin
mediated the effects of Wnt5a on the pancreatic cancer cells. The
data from the present study indicated that Wnt5a is involved in the
modulation of pancreatic cancer cell proliferation, and that Wnt5a
may be a potential target for pancreatic cancer therapy.
PANC-1 and BXPC-3 cells are moderately to poorly
differentiated pancreatic cancer cell lines with different
morphological phenotypes and proliferative potential. When cultured
in vitro, the PANC-1 cells were spindle-shaped and scattered
in growth, whereas the BXPC-3 cells were arranged in nets with a
polygonal shape. Despite their differences, the present study
obtained consistent results following Wnt5a manipulation in
vitro and in the orthotopic nude mouse model. A previous study
by Ripka et al (25)
demonstrated that transient knockdown of Wnt5a using hWNT5A-siRNA
reduces the viability of the PANC-1 and MiaPaca2 pancreatic cancer
cells, consistent with the data obtained in the present study. In
addition, a clonogenicity assay was used in the present study to
assess the group dependency of individual tumor cells and single
cell proliferation capability in vitro. Several studies have
indicated that Wnt5a has an effect on clonogenicity in different
cancer cell lines. For example, Li et al showed that the
ectopic expression of Wnt5a results in a significant inhibition of
clonogenicity in esophageal squamous cell carcinoma cells (26), whereas Ying et al showed
that transfection of a dominant-negative Wnt5a, a nonfunctional
short isoform with the WNT domain deleted, results in substantial
inhibition of colorectal cancer cell clonogenicity (27) and K562 tumor cell clonogenicity
(28). However, Kremenevskaja
et al showed that transfection of Wnt5a in a FTC-133 thyroid
tumor cell line reduces cell clonogenicity (24). The results of these previous
studies indicate that Wnt5a is tumor type-specific and/or dependent
on the whole Wnt5a gene pathway, and may not be just a Wnt5a
protein. In the present study, the data showed that pancreatic
cancer cell lines overexpressing Wnt5a protein enhanced
clonogenicity, whereas suppression of Wnt5a reduced
clonogenicity.
Furthermore, the role of Wnt5a in cell apoptosis has
been shown to be controversial in previous reports and appears to
be cell type-specific. Several studies have indicated that Wnt5a
has anti-apoptotic actions in different cell lines. For example,
the expression of Wnt5a inhibits serum starvation-induced apoptosis
in primarily cultured human dermal fibroblasts (29), HEK293 cells (30), MC3T3-E1 osteoblast cells (31) and isolated germinal center B cells
(32). In addition, Vuga et
al showed that the expression of Wnt5a induces resistance to
H2O2-induced apoptosis in fibroblasts
isolated from lung tissues with interstitial pneumonia (33). However, additional studies have
reported that Wnt5a has a pro-apoptotic action. Liang et al
showed that loss of Wnt5a inhibits the apoptosis of
CD4+/CD8+ thymocytes, whereas exogenous Wnt5a
increases the apoptosis of fetal thymocytes in vitro
(34). Peng et al (35) showed that Wnt5a markedly increases
serum starvation-induced apoptosis in placental choriocarcinoma
cells, compared with untransfected control cells. In the present
study, the results revealed that overexpression of the Wnt5a
protein promoted tumor cell survival/resistance to apoptosis in the
pancreatic cancer cell lines, whereas downregulation of the Wnt5a
protein promoted cell apoptosis. These data indicated a possible
effect of Wnt5a on the regulation of survival and resistance to
apoptosis in pancreatic cancer cells. Furthermore, the data
obtained using the orthotopic nude mouse model showed that the
overexpression of Wnt5a promoted tumor growth and increased the
protein expression of Ki67 in tumor xenografts. Taken together,
these data indicate that Wnt5a exerted distinct effects on the
pancreatic cancer cells through the promotion of tumor growth and
the inhibition of tumor cell apoptosis. As such, it acts as an
oncogene in pancreatic cancer.
Although Wnt5a, as a non-canonical Wnt, is involved
in almost all aspects of the non-canonical Wnt signaling pathway,
previous studies have shown that Wnt5a is able to activate
β-catenin signaling, based on specific receptor availability
(36). Wnt5a is able to switch
between two considerably diverse forms of signaling (37,38).
Increasing evidence indicates a critical role for the
Wnt5a-β-catenin signaling pathway in the pathogenesis of pancreatic
cancer (39,40). It has been reported that the
addition of recombinant Wnt5a leads to increased nuclear β-catenin
in PANC-1 cells, as well as a significant increase in T-cell
factor/lymphoid enhancer factor-dependent reporter activity
(25), indicating the activation
of the canonical signaling pathway. Wang et al (39) also documented that expression of
the ataxia-telangiectasia group D complementing gene increases the
levels of β-catenin in pancreatic cancer, consequently facilitating
tumor growth and metastasis. In our previous study, data revealed
the promotion of β-catenin nuclear translocation in PANC-1 and
BXPC-3 cells by exposure to recombinant Wnt5a (17). In the present study, it was
observed that the depletion of β-catenin reversed the effects of
the recombinant Wnt5a protein on the apoptosis of pancreatic cancer
cells. This indicated that the Wnt5a-β-catenin canonical pathway
was involved in pancreatic cancer cell behavior and in the
progression of pancreatic cancer.
The data presented in the present study provide
significant support for a role for Wnt5a in the development of
pancreatic cancer, and are consistent with our previous findings,
which showed altered expression of Wnt5a in pancreatic cancer
tissues and an association of Wnt5a with certain
clinicopathological data (17).
However, the results of the present study can be considered as
proof-of-principle, and further investigation is required to
clarify the role of Wnt5a in pancreatic cancer. Future
investigations aim to further elucidate the molecular mechanism
underlying the effects of Wnt5a in pancreatic cancer.
Acknowledgments
This study was supported by grants from the National
Key Project of Scientific and Technical Supporting Programs of
China (grant no, 2006BAI02A14) and the National Natural Science
Foundation of China (grant nos. 30770996 and 81172310) to Dr
Minghua Zhu.
References
|
1
|
Pandol S, Gukovskaya A, Edderkaoui M,
Dawson D, Eibl G and Lugea A: Epidemiology, risk factors and the
promotion of pancreatic cancer: Role of the stellate cell. J
Gastroenterol Hepatol. 27(Suppl 2): S127–S134. 2012. View Article : Google Scholar
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Maitra A, Kern SE and Hruban RH: Molecular
pathogenesis of pancreatic cancer. Best Pract Res Clin
Gastroenterol. 20:211–226. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moore PS, Sipos B, Orlandini S, Sorio C,
Real FX, Lemoine NR, Gress T, Bassi C, Klöppel G, Kalthoff H, et
al: Genetic profile of 22 pancreatic carcinoma cell lines. Analysis
of K-ras, p53, p16 and DPC4/Smad4. Virchows Arch. 439:798–802.
2001. View Article : Google Scholar
|
|
6
|
Li D, Xie K, Wolff R and Abbruzzese JL:
Pancreatic cancer. Lancet. 363:1049–1057. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Logan CY and Nusse R: The Wnt signaling
pathway in development and disease. Annu Rev Cell Dev Biol.
20:781–810. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qian D, Jones C, Rzadzinska A, Mark S,
Zhang X, Steel K, Dai X and Chen P: Wnt5a functions in planar cell
polarity regulation in mice. Dev Biol. 306:121–133. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wong GT, Gavin BJ and McMahon AP:
Differential transformation of mammary epithelial cells by Wnt
genes. Mol Cell Biol. 14:6278–6286. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fang DR, Lv ZF and Qiao G: Dynamic Wnt5a
expression in murine hair follicle cycle and its inhibitory effects
on follicular. Asian Pac J Trop Med. 7:285–288. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maeda K, Kobayashi Y, Udagawa N, Uehara S,
Ishihara A, Mizoguchi T, Kikuchi Y, Takada I, Kato S, Kani S, et
al: Wnt5a-Ror2 signaling between osteoblast-lineage cells and
osteoclast precursors enhances osteoclastogenesis. Nat Med.
18:405–412. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Christman MA 2nd, Goetz DJ, Dickerson E,
McCall KD, Lewis CJ, Benencia F, Silver MJ, Kohn LD and Malgor R:
Wnt5a is expressed in murine and human atherosclerotic lesions. Am
J Physiol Heart Circ Physiol. 294:2864–2870. 2008. View Article : Google Scholar
|
|
14
|
Bakker ER, Das AM, Helvensteijn W, Franken
PF, Swagemakers S, van der Valk MA, ten Hagen TL, Kuipers EJ, van
Veelen W and Smits R: Wnt5a promotes human colon cancer cell
migration and invasion but does not augment intestinal
tumorigenesis in Apc1638N mice. Carcinogenesis. 34:2629–2638. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Trifa F, Karray-Chouayekh S, Jmal E, Jmaa
ZB, Khabir A, Sellami-Boudawara T, Frikha M, Daoud J and
Mokdad-Gargouri R: Loss of WIF-1 and Wnt5a expression is related to
aggressiveness of sporadic breast cancer in Tunisian patients.
Tumour Biol. 34:1625–1633. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee SE, Lim SD, Kang SY, Suh SB and Suh
YL: Prognostic significance of Ror2 and Wnt5a expression in
medulloblastoma. Brain Pathology. 23:445–453. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bo H, Zhang S, Gao L, Chen Y, Zhang J,
Chang X and Zhu M: Upregulation of Wnt5a promotes epithelial- to
mesenchymal transition and metastasis of pancreatic cancer cells.
BMC Cancer. 13:4962013. View Article : Google Scholar
|
|
18
|
Nishita M, Yoo SK, Nomachi A, Kani S,
Sougawa N, Ohta Y, Takada S, Kikuchi A and Minami Y: Filopodia
formation mediated by receptor tyrosine kinase Ror2 is required for
Wnt5a-induced cell migration. J Cell Biol. 175:555–562. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Matsumoto S, Fumoto K, Okamoto T, Kaibuchi
K and Kikuchi A: Binding of APC and dishevelled mediates
Wnt5a-regulated focal adhesion dynamics in migrating cells. EMBO J.
29:1192–1204. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yamamoto H, Kitadai Y, Yamamoto H, Oue N,
Ohdan H, Yasui W and Kikuchi A: Laminin gamma2 mediates
Wnt5a-induced invasion of gastric cancer cells. Gastroenterology.
137:242–252. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lehtonen A, Ahlfors H, Veckman V,
Miettinen M, Lahesmaa R and Julkunen I: Gene expression profiling
during differentiation of human monocytes to macrophages or
dendritic cells. J Leukoc Biol. 82:710–720. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamamoto H, Oue N, Sato A, Hasegawa Y,
Yamamoto H, Matsubara A, Yasui W and Kikuchi A: Wnt5a signaling is
involved in the aggressiveness of prostate cancer and expression of
metalloproteinase. Oncogene. 29:2036–2046. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang CL, Liu D, Nakano J, Ishikawa S,
Kontani K, Yokomise H and Ueno M: Wnt5a expression is associated
with the tumor proliferation and the stromal vascular endothelial
growth factor-an expression in non-small-cell lung cancer. J Clin
Oncol. 23:8765–8773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kremenevskaja N, von Wasielewski R, Rao
AS, Schöfl C, Andersson T and Brabant G: Wnt-5a has tumor
suppressor activity in thyroid carcinoma. Oncogene. 24:2144–2154.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ripka S, Konig A, Buchholz M, Wagner M,
Sipos B, Kloppel G, Downward J, Gress T and Michl P: WNT5A-target
of CUTL1 and potent modulator of tumor cell migration and invasion
in pancreatic cancer. Carcinogenesis. 28:1178–1187. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li J, Ying J, Fan Y, Wu L, Ying Y, Chan
AT, Srivastava G and Tao Q: WNT5A antagonizes WNT/β-catenin
signaling and is frequently silenced by promoter CpG methylation in
esophageal squamous cell carcinoma. Cancer Biol Ther. 10:617–624.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ying J, Li H, Yu J, Ng KM, Poon FF, Wong
SC, Chan AT, Sung JJ and Tao Q: WNT5A exhibits tumor-suppressive
activity through antagonizing the Wnt/beta-catenin signaling, and
is frequently methylated in colorectal cancer. Clin Cancer Res.
14:55–61. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ying J, Li H, Chen YW, Srivastava G, Gao Z
and Tao Q: WNT5A is epigenetically silenced in hematologic
malignancies and inhibits leukemia cell growth as a tumor
suppressor. Blood. 110:4130–4131. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Torii K, Nishizawa K, Kawasaki A,
Yamashita Y, Katada M, Ito M, Nishimoto I, Terashita K, Aiso S and
Matsuoka M: Anti-apoptotic action of Wnt5a in dermal fibroblasts is
mediated by the PKA signaling pathways. Cell Signal. 20:1256–1266.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jia L, Miao C, Cao Y and Duan EK: Effects
of Wnt proteins on cell proliferation and apoptosis in HEK293
cells. Cell Biol Int. 32:807–813. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Almeida M, Han L, Bellido T, Manolagas SC
and Kousteni S: Wnt proteins prevent apoptosis of both uncommitted
osteoblast progenitors and differentiated osteoblasts by
beta-catenin-dependent and -independent signaling cascades
involving Src/ERK and phosphatidylinositol 3-kinase/AKT. J Biol
Chem. 280:41342–41351. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim J, Kim DW, Chang W, Choe J, Kim J,
Park CS, Song K and Lee I: Wnt5a is secreted by follicular
dendritic cells to protect germinal center B cells via
Wnt/Ca2+/NFAT/NF-κB-B cell lymphoma 6 signaling. J Immunol.
188:182–189. 2012. View Article : Google Scholar
|
|
33
|
Vuga LJ, Ben-Yehudah A,
Kovkarova-Naumovski E, Oriss T, Gibson KF, Feghali-Bostwick C and
Kaminski N: WNT5A is a regulator of fibroblast proliferation and
resistance to apoptosis. Am J Respir Cell Mol Biol. 41:583–589.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liang H, Coles AH, Zhu Z, Zayas J, Jurecic
R, Kang J and Jones SN: Noncanonical Wnt signaling promotes
apoptosis in thymocyte development. J Exp Med. 204:3077–3084. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Peng S, Zhang J, Chen J and Wang H:
Effects of Wnt5a protein on proliferation and apoptosis in JAR
choriocarcinoma cells. Mol Med Report. 4:99–104. 2011.
|
|
36
|
Umbhauer M, Djiane A, Goisset C,
Penzo-Méndez A, Riou JF, Boucaut JC and Shi DL: The C-terminal
cytoplasmic Lys-thr-X-X-X-Trp motif in frizzled receptors mediates
Wnt/beta-catenin signaling. EMBO J. 19:4944–4954. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mikels AJ and Nusse R: Purified Wnt5a
protein activates or inhibits beta-catenin-TCF signaling depending
on receptor context. PLoS Biol. 4:e1152006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van Amerongen R, Fuerer C, Mizutani M and
Nusse R: Wnt5a can both activate and repress Wnt/β-catenin
signaling during mouse embryonic development. Dev Biol.
369:101–114. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang L, Heidt DG, Lee CJ, Yang H, Logsdon
CD, Zhang L, Fearon ER, Ljungman M and Simeone DM: Oncogenic
function of ATDC in pancreatic cancer through Wnt pathway
activation and beta-catenin stabilization. Cancer Cell. 15:207–219.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kobayashi T, Shimura T, Yajima T, Kubo N,
Araki K, Tsutsumi S, Suzuki H, Kuwano H and Raz A: Transient gene
silencing of galectin-3 suppresses pancreatic cancer cell migration
and invasion through degradation of β-catenin. Int J Cancer.
129:2775–2786. 2011. View Article : Google Scholar : PubMed/NCBI
|