Introduction
Ovarian cancer is one of the most common
gynecological malignancies and the seventh most common type of
cancer in women. More than 14,000 women succumb to ovarian cancer
annually and there appears to be a global increase in the incidence
rates (1–3). The clinical application of
chemotherapeutic drugs to treat ovarian cancer is extended,
however, the side effects of these drugs limit their usefulness
(4,5). It is therefore essential that novel
drugs are developed for the treatment of ovarian cancer.
Flavonoids are composed of >4,000 polyphenolic
compounds present in vegetables, fruits, tea and certain medicinal
herbs (6,7). Flavonoids are divided into flavones,
flavanonols, flavanols, isoflavones and flavonols, according to
their saturation levels (8,9).
Myricetin (2,5,7,3,4,5-pentahydroxylflavonol) is a ubiquitous
flavonol (10). Previous studies
have shown that myricetin is effective against certain types of
cancer (11–13), however, its mechanisms of action in
human ovarian cancer remain to be elucidated. The present study
examined the effects of myricetin on SKOV3 ovarian cancer cells and
explored the apoptotic pathways involved in myricetin-induced SKOV3
cell death.
The endoplasmic reticulum (ER) is an organelle with
various important biological functions in eukaryotic cells. Several
physiological and pathological factors can trigger ER stress.
Glucose-regulated protein (GRP)-78, an ER stress chaperone
molecule, is upregulated when various stimuli trigger ER stress.
Moderate ER stress can act as a protective mechanism (14), however, prolonged and excessive ER
stress eventually leads to apoptosis (14,15).
Previous studies have suggested that DNA doubled-strand breaks
(DSBs) can also lead to apoptosis (16–18).
Phosphorylation of H2AX (γ-H2AX) is a widely
recognized marker of DNA DSBs, and increases in DNA DSBs are
reflected by the upregulation of γ-H2AX levels (17).
In the present study, myricetin treatment inhibited
SKOV3 cell viability and induced subsequent apoptosis. The present
study identified both ER stress and DNA DSBs as factors involved in
myricetin-induced SKOV3 cell apoptosis.
Materials and methods
Cell culture and treatment
Human ovarian cancer SKOV3 cells were purchased from
the Chinese Academy of Medical Sciences (Beijing, China) and were
maintained in RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.,
Carlsbad, CA, USA), supplemented with 10% (v/v) fetal calf serum
(Gibco; Thermo Fisher Scientific, Inc.), 100 mg/ml streptomycin and
100 U/ml penicillin (each from Genview, Galveston, TX, USA). The
cells were incubated at 37°C in an atmosphere containing 5%
CO2. Myricetin was purchased from Sigma-Aldrich (St.
Louis, MO, USA) and was dissolved in dimethylsulfoxide (DMSO) for
storage at −20°C.
Cell viability assay
Cell viability was determined using a
3-(4,5-dimetrylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay (Beyotime Institute of Biotechnology, Haimen, China). SKOV3
cells, during the exponential growth phase, were seeded into
96-well culture plates in 100 µl RPMI-1640 at a density of
8×103 cells/well. Following 24 h incubation, the
indicated dose of myricetin were added for a further 24 h
incubation in four parallel wells. The MTT assays were performed as
follows: 20 µl MTT solution [(5 mg/ml in phosphate-buffered
saline (PBS)] was added to each well and the cells were incubated
at 37°C for 4 h, following which 150 µl DMSO (Beijing
Chemical Industry Co., Ltd., Beijing, China) was added to each
well. The cells were agitated for 10 min prior to measuring the
absorbance at 570 nm using a microplate reader (680; Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The growth inhibition rate
was calculated as follows: Inhibition (%) = [1 − (absorbance of
experimental group / absorbance of control group)] × 100. The mean
value of four replicate wells was calculated for each treatment
group.
Western blotting
Whole-cell protein extracts from SKOV3 cells were
prepared using cell lysis buffer [50 mM Tris-hydrochloride (HCl; pH
7.5); 150 mM NaCl; 1 mM Na2 EDTA; 1 mM EDTA; 1% Triton;
2.5 mM sodium pyrophosphate; 1 mM β-glycerophosphate; 1 mM
Na3VO4; 1 mM NaF; 1 µg/ml leupeptin; 1
mM PMSF]. The protein extracts were quantified using a Bio-Rad
Protein Assay kit (Bio-Rad Laboratories, Inc.). For western blot
analysis, protein lysates (30–50 µg) were separated by 12%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis and
transferred onto Immobilon-P Membranes (EMD Millipore, Billerica,
MA, USA). The membranes were blocked with 5% non-fat milk powder in
buffer [10 mM Tris-HCl (pH 7.6), 100 mM NaCl and 0.1% Tween-20] for
2 h at room temperature and were subsequently incubated with the
appropriate primary antibodies overnight at 4°C. Anti-GRP-78 rabbit
anti-human polyclonal antibody (cat no. sc-13968; dilution, 1:200)
was obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA), mouse anti-human anti-C/EBP homologous protein (CHOP)
monoclonal antibody (cat no. ab11419; dilution, 1:1,000) from Abcam
(Hong Kong, China), rabbit anti-human anti-γ-H2AX
monoclonal antibody (cat no. 9718; dilution, 1:1,000) from Cell
Signaling Technology (Beverly, MA, USA), and mouse anti-human
anti-β-actin monoclonal antibody (cat no. 60008-1-Ig; dilution,
1:1,000) from Proteintech Group, Inc. (Chicago, IL, USA). Following
incubation, the membranes were incubated with horseradish
peroxidase-conjugated secondary antibody (Thermo Fisher Scientific,
Inc.) at a dilution of 1:2,000 for 1 h at room temperature.
Immunodetection was performed using enhanced chemiluminescence
reagents and images were captured using a Syngene Bio Imaging
system (Synoptics, Cambridge, UK). The protein levels were
normalized against those of β-actin, and the ratios of the
normalized protein are presented as the mean ± standard deviation
from three independent experiments. Protein levels were quantified
by densitometry using Quantity One software version 4.4.02 (Bio-Rad
Laboratories, Inc.).
Immunofluorescent staining and confocal
laser microscopy
The cells were seeded onto coverslips in 24-well
plates at a density of 5×104 cells/well 24 h prior to
treatment. Following exposure to 40 µg/ml myricetin for 0,
6, 12 and 24 h, the cells were fixed with 4% paraformaldehyde for
30 min at room temperature, stained with the nuclear stain Hoechst
33342 (2 µg/ml; Sigma-Aldrich) for 2 min at room
temperature, washed with PBS and examined using a confocal laser
microscope (FV1000; Olympus, Tokyo, Japan) to reveal chromatin
condensation. The expression levels of GRP-78, active Caspase 3 and
γ-H2AX were examined using an indirect
immunofluorescence method. Briefly, after the cells were cultured,
treated and fixed, as previously described, they were subsequently
permeabilized with 0.1% Triton X-100 for 5 min, blocked with bovine
serum albumin for 30 min and incubated with primary antibodies
against GRP-78, active caspase 3 and γ-H2AX (dilution,
1:100) overnight at 4°C. Following incubation, the cells were
incubated with fluorescein isothiocyanate/Texas Red-conjugated
secondary antibodies (dilution, 1:400; Santa Cruz Biotechnology,
Inc.) for 1 h at room temperature, stained with Hoechst 33342 (2
µg/ml) for 2 min at room temperature, washed with PBS three
times, and examined using the Olympus FV1000 confocal laser
microscope.
Statistical analysis
The data are representative of the results from
three independent experiments. Statistical analysis was performed
using one-way analysis of variance. The Tukey post-hoc test was
used to determine the significance of all pairwise comparisons of
interest. P<0.05 was considered to indicate a statistically
significant difference.
Results
Myricetin inhibits the viability of SKOV3
cells
Our previous results provided us with the
appropriate dose range for myricetin treatment (unpublished data).
SKOV3 cells were treated with the indicated doses of myricetin for
24 h. Myricetin inhibited the viability of SKOV3 cells in a
dose-dependent manner (Fig. 1A).
In addition, changes in cell morphology were also apparent
following myricetin treatment. Myricetin-treated cells appeared
more rounded and shrunken compared with the control group (Fig. 1B). The present study, therefore,
hypothesized that myricetin may induce apoptosis in SKOV3 cells,
and this was examined in detail using confocal microscopy.
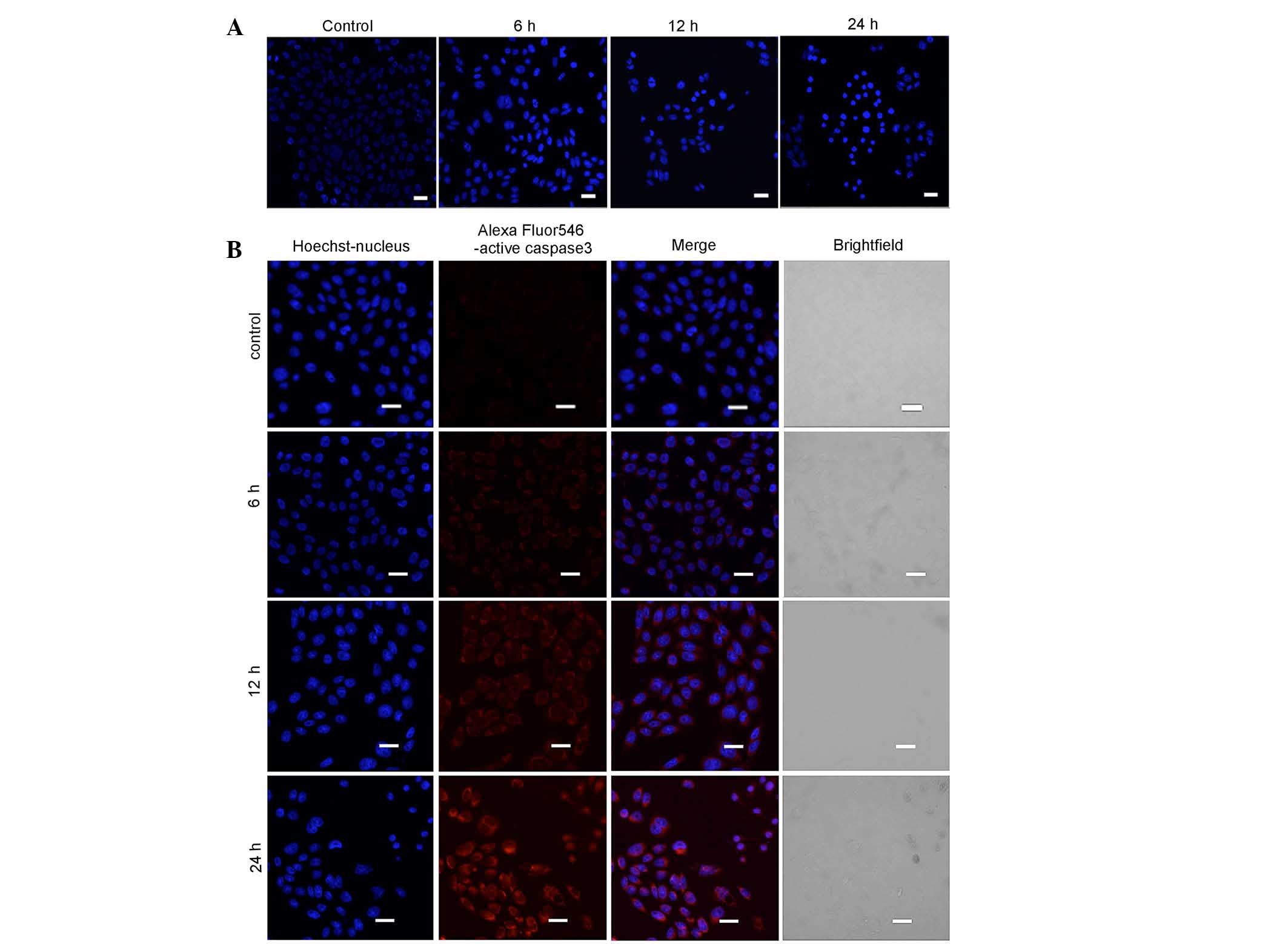
Myricetin triggers apoptosis in SKOV3
cells
Based on the above MTT results, SKOV3 cells were
treated with 40 µg/ml myricetin for 0, 6, 12 or 24 h,
stained with Hoechst 33342, and examined using confocal microscopy.
The nuclei of myricetin-treated cells appeared more condensed, when
compared with the untreated cells (Fig. 2A).
Caspase 3 is considered to be the primary executor
of apoptosis, and cleaved Caspase 3 (active Caspase 3) is used as a
biomarker for apoptosis. As shown in Fig. 2B, increasing concentrations of
myricetin generated higher levels of red fluorescence. This
indicated that myricetin induced the activation of Caspase 3 in a
time-dependent manner (Fig. 2b).
Together, these results indicated that myricetin triggered
apoptosis in SKOV3 cells.
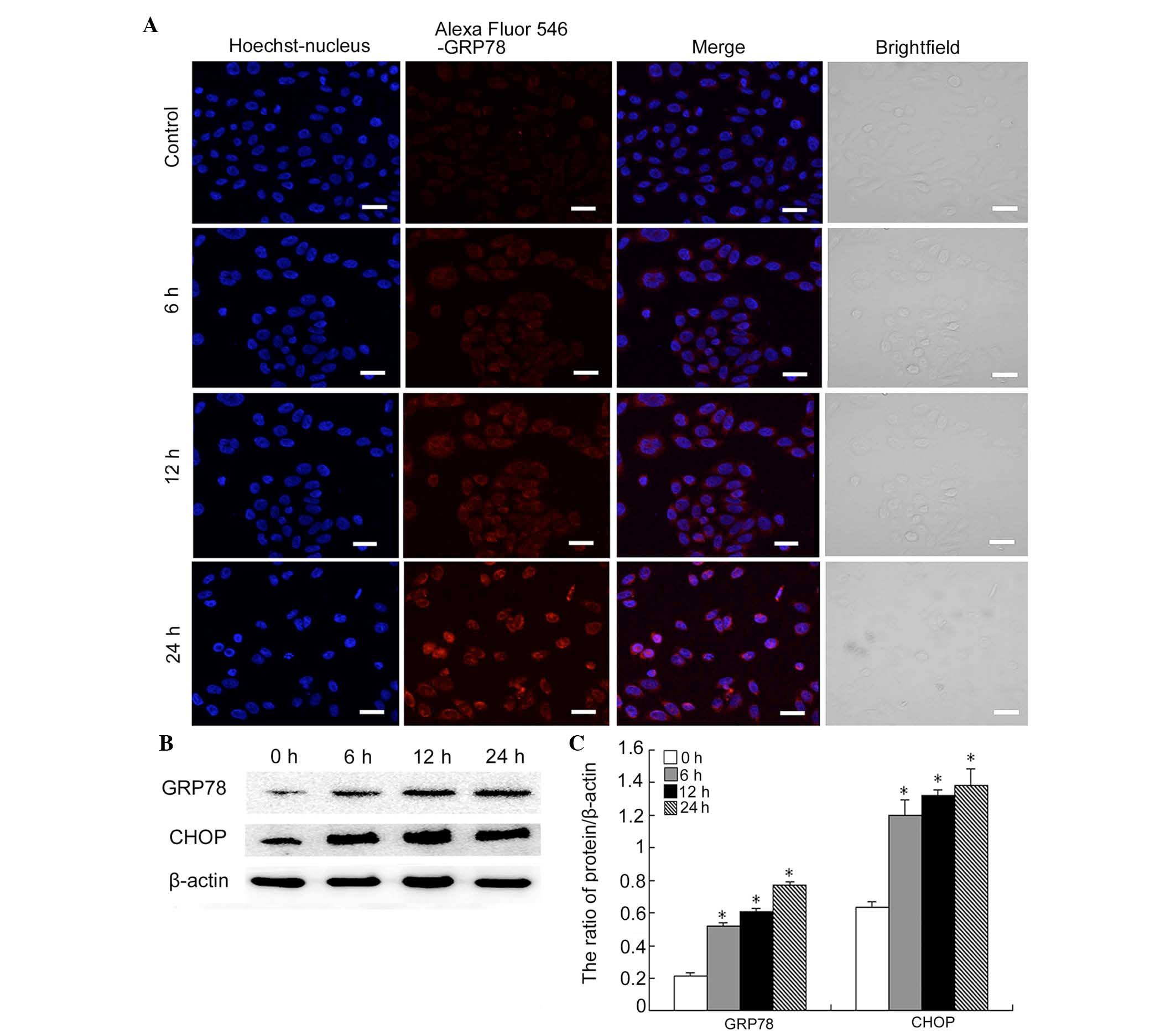
Myricetin induces ER stress-associated
apoptosis in SKOV3 cells
GRP-78 is an ER chaperone molecule, which increases
following ER stress (19). In
order to determine whether myricetin induced ER stress, confocal
microscopy was used to detect the expression of GRP-78 in
myricetin-treated cells. Myricetin was found to increase the mean
fluorescence intensity of GRP-78 in SKOV3 cells, which became
notable following 24 h of treatment (Fig. 3A).
Increased and sustained ER stress can cause an
apoptotic response. To determine whether the response to myricetin
treatment caused ER stress-associated apoptosis, the levels of CHOP
were investigated (20). Western
blot analysis indicated that the protein levels of CHOP markedly
increased at 6, 12 and 24 h following myricetin treatment (Fig. 3B and C). Consistent with the data
from the confocal experiments, GRP-78 levels also increased in a
time-dependent manner (Fig. 2B and
C). These results indicated that myricetin triggers ER
stress-associated apoptosis.
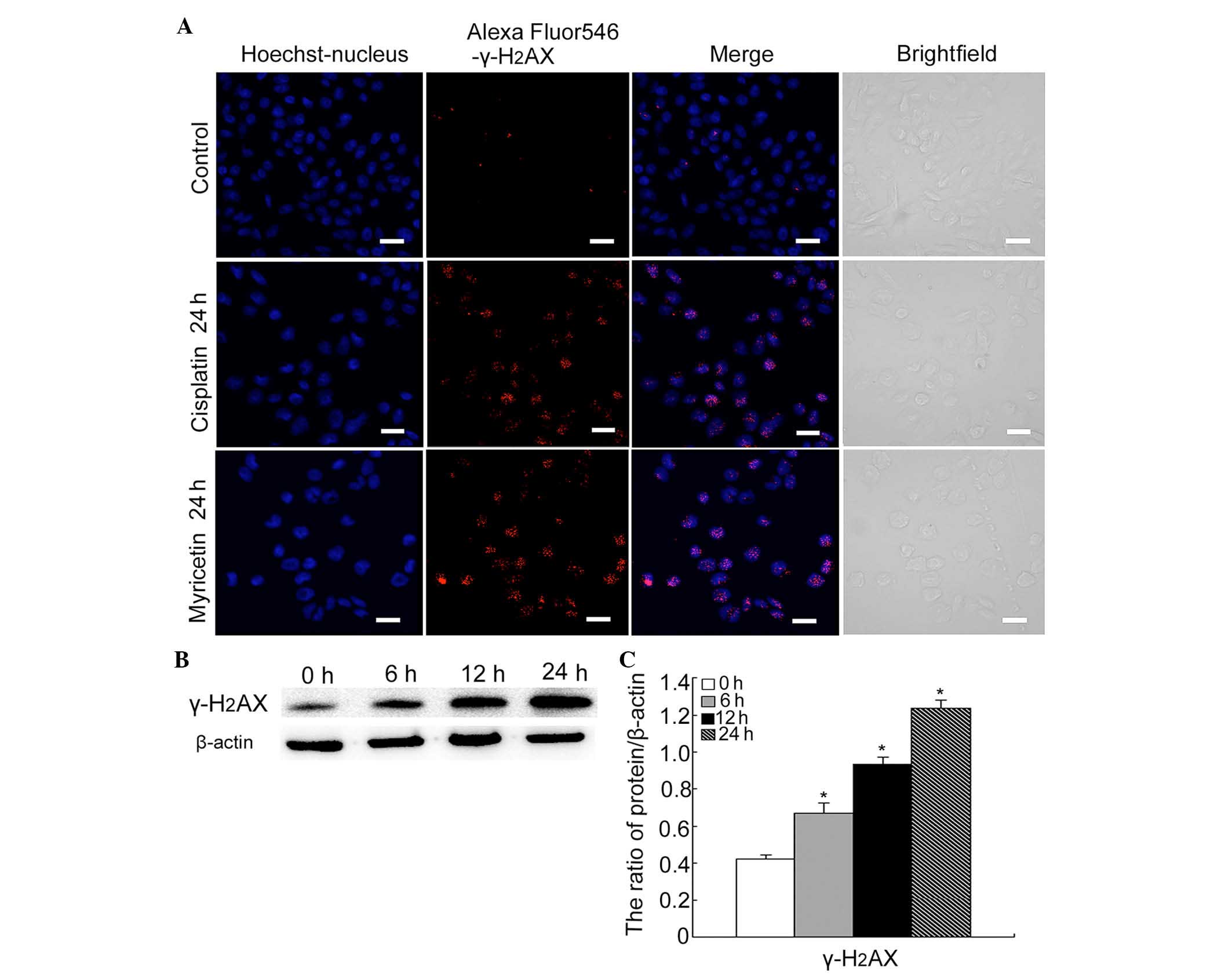
Myricetin induces DNA DSBs in SKOV3
cells
Cisplatin is one of the classic chemotherapeutic
drugs known to induce a marked apoptotic response by inhibiting DNA
replication and damage (21,22).
It was therefore hypothesized that myricetin can also induce DNA
DSBs. It has been reported that following a DSB, histone
H2AX is rapidly phosphorylated (becoming
γ-H2AX) near the site of the DSB, and is involved in the
recruitment of other factors that contribute to lesion repair. The
levels of γ-H2AX have been shown to be downregulated
when the DNA is repaired. If the lesion remains unrepaired (or the
repair process is delayed), then the levels of γ-H2AX
remain high; therefore, the levels of γ-H2AX comprise an
ideal indicator of the degree of unrepaired DSBs, which can then
contribute to cell death (23,24).
Firstly, using confocal microscopy, the expression
of γ-H2AX was detected in cells treated with either
cisplatin (6 µg/ml) or myricetin (40 µg/ml).
Cisplatin-treated cells were used as the positive control. The
results revealed that both cisplatin and myricetin increased the
formation of the γ-H2AX foci (Fig. 4A). Secondly, western blotting was
used to detect the protein levels of γ-H2AX in
myricetin-treated cells. It was revealed that myricetin increased
the levels of γ-H2AX at 6, 12 and 24 h treatment
(Fig. 4B and C).
Discussion
Myricetin is abundant in nature and can be found in
walnuts, vegetables and fruits (25). Myricetin has numerous biological
functions, including antioxidant and anticarcinogenic functions. In
addition, myricetin has been reported to inhibit RNA/DNA
replication and repair (26–28).
A number of its biological functions suggest its potential use in
clinical medicine. In HCT-15 colon cancer cells, myricetin induced
cytotoxicity, which was mediated by a mitochondrial pathway
(25). A different study indicated
that myricetin can be used as auxiliary therapy, increasing the
sensitivity of esophageal carcinoma cells to 5-fluorouracil in
vitro and in vivo (29). In the present study, myricetin
inhibited viability, induced nuclear fragmentation, upregulated the
level of active Caspase 3, and subsequently induced apoptosis in
SKOV3 cells. Apoptosis can be governed by extrinsic or intrinsic
pathways; however, this study focused on the effect of myricetin on
the intrinsic pathways, and in particular the ER stress-associated
pathway.
ER is an organelle responsible for a number of
functions, including the maintenance of intracellular calcium
balance and protein synthesis, modification and processing.
Numerous cellular pathological and physiological processes can
cause ER stress (30,31), which, in turn, can trigger the
unfolded protein response (UPR) to recover ER homeostasis. UPR
signaling pathways include the inositol-requiring enzyme (IRE)1,
activating transcription factor (ATF)6 and protein kinase RNA-like
endoplasmic reticulum kinase (PERK) pathways. GRP-78, an ER stress
molecular chaperone, is upregulated following ER stress, and has a
positive regulatory role in the prevention of apoptosis (32); however, severe and persistent ER
stress has been reported to induce apoptosis even when GRP-78
protein is overexpressed (33).
CHOP is a downstream element of the ER stress pathway and a
convergence point of the IRE1, ATF6 and PERK pathways (34). CHOP is known to have a regulatory
role as an apoptotic switch. It has been shown that a high
expression of CHOP contributes to ER stress-mediated apoptosis. An
increasing number of studies have identified ER as a target of
apoptosis-inducing drugs in cancer cells (35–37).
The present study demonstrated that myricetin
upregulated the level of GRP-78 and CHOP in a time-dependent
manner. It further revealed that myricetin induces ER
stress-associated apoptosis in SKOV3 cells. Under normal
conditions, DSBs occur and are quickly repaired by DNA repair
mechanisms. Failure of these mechanisms leads to unrepaired DSBs,
ultimately resulting in apoptosis (38,39).
The expression of γ-H2AX comprises a marker for
unrepaired DSBs. The present results showed that myricetin induces
DNA DSBs in SKOV3 cells, suggesting that myricetin may target DNA
damage repair.
In conclusion, myricetin treatment was revealed to
inhibit viability and induce apoptosis through ER stress and DNA
DSBs in SKOV3 human ovarian cancer cells.
Acknowledgments
The present study was supported by the National
Nature and Science foundation of China (nos. NSFC81372793 and
81272876), the Department of Education of Jilin Province Project
(no. 2013361) and the Scientific Research Foundation of Jilin
Province for University Students. The authors would like to thank
Director Benjamin Shaw from Liwen Bianji (Edanz Group China) for
the language editing of this manuscript.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tumanian SV and Iartseva DV: Effect of
hepatic functional activity of the liver and endogenous
intoxication in patients with ovarian cancer. Khirurgiia (Mosk).
45–47. 2014.In Russian.
|
|
3
|
Shilpa V, Bhagat R, Premalata CS, Pallavi
VR, Ramesh G and Krishnamoorthy L: Relationship between promoter
methylation & tissue expression of MGMT gene in ovarian cancer.
Indian J Med Res. 140:616–623. 2014.
|
|
4
|
Shibata Y: Initial safety and efficacy of
cisplatin and gemcitabine combination chemotherapy for unresectable
biliary tract cancer. Gan To Kagaku Ryoho. 41:2599–2602. 2014.
|
|
5
|
Yang J, Shi Y, He X, Dong M, Zhang C, Liu
P, Zhou S, Qin Y, Gui L, Yang S and Sun Y: A pilot study of the
safety and efficacy of dexamethasone, ifosfamide, methotrexate and
gemcitabine chemotherapy for natural killer/T-cell lymphoma. Leuk
Lymphoma. 56:2218–2221. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu X, Ye F, Wu J, How B, Li W and Zhang
DY: Signaling proteins and pathways affected by flavonoids in
leukemia cells. Nutr Cancer. 67:238–249. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schwarz D, Kisselev P, Schunck WH and
Roots I: Inhibition of 17β-estradiol activation by CYP1A1:
Genotype- and regioselective inhibition by St. John's Wort and
several natural polyphenols. Biochim Biophys Acta. 1814:168–174.
2011. View Article : Google Scholar
|
|
8
|
Kozyra M and Skalicka-Woźniak K:
Quantitative analysis of flavonoids and phenolic acids from
inflorescences and aerial parts of selected Cirsium species using
ASE method. Acta Pol Pharm. 71:877–881. 2014.PubMed/NCBI
|
|
9
|
Tsui KC, Chiang TH, Wang JS, Lin LJ, Chao
WC, Chen BH and Lu JF: Flavonoids from gynostemma pentaphyllum
exhibit differential induction of cell cycle arrest in H460 and
A549 cancer cells. Molecules. 19:17663–17681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu C, Wang W, Tian B, Liu X, Qu X, Zhai Z,
Li H, Liu F, Fan Q, Tang T, et al: Myricetin prevents titanium
particle-induced osteolysis in vivo and inhibits RANKL-induced
osteoclastogenesis in vitro. Biochem Pharmacol. 93:59–71. 2015.
View Article : Google Scholar
|
|
11
|
Weng CJ and Yen GC: Flavonoids, a
ubiquitous dietary phenolic subclass, exert extensive in vitro
anti-invasive and in vivo anti-metastatic activities. Cancer
Metastasis Rev. 31:323–351. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun F, Zheng XY, Ye J, Wu TT, Wang JL and
Chen W: Potential anticancer activity of myricetin in human T24
bladder cancer cells both in vitro and in vivo. Nutr Cancer.
64:599–606. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Choi HN, Kang MJ, Lee SJ and Kim JI:
Ameliorative effect of myricetin on insulin resistance in mice fed
a high-fat, high-sucrose diet. Nutr Res Pract. 8:544–549. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brüning A, Kimmich T, Brem GJ, Buchholtz
ML, Mylonas I, Kost B, Weizsäcker K and Gingelmaier A: Analysis of
endoplasmic reticulum stress in placentas of HIV-infected women
treated with protease inhibitors. Reprod Toxicol. 50:122–128. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Marí M, Morales A, Colell A, García-Ruiz C
and Fernández-Checa JC: Mitochondrial cholesterol accumulation in
alcoholic liver disease: Role of ASMase and endoplasmic reticulum
stress. Redox Bio. 3:100–108. 2014. View Article : Google Scholar
|
|
16
|
Bazyka DA, Muzalevska KD, Maznichenko OL
and Belyaev OA: Expression of γ-H2AX histone in lymphocytes of the
Chornobyl 'Shelter' object staff exposed to ionizing radiation in
occupational limits. Probl Radiac Med Radiobiol. 19:186–191.
2014.In English, Ukrainian. PubMed/NCBI
|
|
17
|
Valdiglesias V, Giunta S, Fenech M, Neri M
and Bonassi S: γ-H2AX as a marker of DNA double strand breaks and
genomic instability in human population studies. Mutat Res.
753:24–40. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Reynolds M, Armknecht S, Johnston T and
Zhitkovich A: Undetectable role of oxidative DNA damage in cell
cycle, cytotoxic and clastogenic effects of Cr(VI) in human lung
cells with restored ascorbate levels. Mutagenesis. 27:437–443.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fan L, Li A, Li W, Cai P, Yang B, Zhang M,
Gu Y, Shu Y, Sun Y, Shen Y, et al: Novel role of Sarco/endoplasmic
reticulum calcium ATPase 2 in development of colorectal cancer and
its regulation by F36, a curcumin analog. Biomed Pharmacother.
68:1141–1148. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu Y, Yu H, Qin H, Kang J, Yu C, Zhong J,
Su J, Li H and Sun L: Inhibition of autophagy enhances cisplatin
cytotoxicity through endoplasmic reticulum stress in human cervical
cancer cells. Cancer Lett. 314:232–243. 2012. View Article : Google Scholar
|
|
21
|
Maheshwari RA, Sailor GU, Patel L and
Balaraman R: Amelioration of cisplatin-induced nephrotoxicity by
statins. Indian J Pharmacol. 45:354–358. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lingeman RG, Hickey RJ and Malkas LH:
Expression of a novel peptide derived from PCNA damages DNA and
reverses cisplatin resistance. Cancer Chemother Pharmacol.
74:981–993. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bazyka DA, Muzalevska KD, Maznichenko OL
and Belyaev OA: Expression of γ-H2AX histone in lymphocytes of the
Chornobyl 'Shelter' object staff exposed to ionizing radiation in
occupational limits. Probl Radiac Med Radiobiol. 19:186–191.
2014.In English, Ukrainian. PubMed/NCBI
|
|
24
|
Thompson CM, Seiter J, Chappell MA,
Tappero RV, Proctor DM, Suh M, Wolf JC, Haws LC, Vitale R, Mittal
L, et al: Synchrotron-based imaging of chromium and γ-H2AX
immunostaining in the duodenum following repeated exposure to
Cr(VI) in drinking water. Toxicol Sci. 143:16–25. 2015. View Article : Google Scholar :
|
|
25
|
Kim ME, Ha TK, Yoon JH and Lee JS:
Myricetin induces cell death of human colon cancer cells via
BAX/BCL2-dependent pathway. Anticancer Res. 34:701–706.
2014.PubMed/NCBI
|
|
26
|
Zhang XH, Chen SY, Tang L, Shen YZ, Luo L,
Xu CW, Liu Q and Li D: Myricetin induces apoptosis in Hepg2 cells
through Akt/P70s6k/bad signaling and mitochondrial apoptotic
pathway. Anticancer Agents Med Chem. 13:1575–1581. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nirmala P and Ramanathan M: Effect of
myricetin on 1,2 dimethylhydrazine induced rat colon
carcinogenesis. J Exp Ther Oncol. 9:101–108. 2011.PubMed/NCBI
|
|
28
|
Zhang S, Wang L, Liu H, Zhao G and Ming L:
Enhancement of recombinant myricetin on the radiosensitivity of
lung cancer A549 and H1299 cells. Diagn Pathol. 9:682014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang L, Feng J, Chen X, Guo W, Du Y, Wang
Y, Zang W, Zhang S and Zhao G: Myricetin enhance chemosensitivity
of 5-fluorouracil on esophageal carcinoma in vitro and in vivo.
Cancer Cell Int. 14:712014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang D, Gao L, Wang T, Qiao Z, Liang Y and
Zhang P: Hypoxia triggers endothelial endoplasmic reticulum stress
and apoptosis via induction of VLDL receptor. FEBS Lett.
588:4448–4456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yamamoto S, Yamashita A, Arakaki N, Nemoto
H and Yamazaki T: Prevention of aberrant protein aggregation by
anchoring the molecular chaperone αB-crystallin to the endoplasmic
reticulum. Biochem Biophys Res Commun. 455:241–245. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Komatsu M and Ichimura Y: Physiological
significance of selective degradation of p62 by autophagy. FEBS
Lett. 584:1374–1378. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zheng YZ, Cao ZG, Hu X and Shao ZM: The
endoplasmic reticulum stress markers GRP-78 and CHOP predict
disease-free survival and responsiveness to chemotherapy in breast
cancer. Breast Cancer Res Treat. 145:349–358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu H, Su J, Xu Y, Kang J, Li H, Zhang L,
Yi H, Xiang X, Liu F and Sun L: p62/SQSTM1 involved in cisplatin
resistance in human ovarian cancer cells by clearing ubiquitinated
proteins. Eur J Cancer. 47:1585–1594. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang D, Gao L, Wang T, Qiao Z, Liang Y and
Zhang P: Hypoxia triggers endothelial endoplasmic reticulum stress
and apoptosis via induction of VLDL receptor. FEBS Lett.
588:4448–4456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ahmad M, Hahn IF and Chatterjee S: GRP-78
up-regulation leads to hypersensitization to cisplatin in A549 lung
cancer cells. Anticancer Res. 34:3493–3500. 2014.PubMed/NCBI
|
|
37
|
Chen Z, Ballar P, Fu Y, Luo J, Du S and
Fang S: The E3 ubiquitin ligase gp78 protects against ER stress in
zebrafish liver. J Genet Genomics. 41:357–368. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Macfie A, Hagan E and Zhitkovich A:
Mechanism of DNA-protein cross-linking by chromium. Chem Res
Toxicol. 23:341–347. 2010. View Article : Google Scholar
|
|
39
|
Panier S and Boulton SJ: Double-strand
break repair: 53BP1 comes into focus. Nat Rev Mol Cell Biol.
15:7–18. 2014. View
Article : Google Scholar
|