Introduction
Alzheimer's disease (AD) is characterized clinically
by progressive memory loss and cognitive decline (1). In the United States, ~5 million
people have AD and with an aging population, ~16 million people are
predicted to be diagnosed with AD by middle of the century in the
US alone (2). Neuropathological
features of AD include extracellular aggregation of amyloid-β (Aβ)
and intraneuronal neurofibrillary tangles (aggregates of tau
composed primarily of hyperphosphorylated tau) (3). In addition, inflammation occurs in
the brains of patients with AD. In primary microglial and astrocyte
cell cultures obtained from postmortem AD patients and nondemented
elderly (ND) controls, and in neuroblastoma cell lines, the
plaque-associated cytokines interleukin (IL) 1β, IL6 and
hypoxia-inducible factor 1α (HIF1α) stimulated the production of
complement component (C) 1 s and C1r as a result of direct binding
and activation of C1 by Aβ (4).
Furthermore, patients with no history of dementia who exhibit
sufficient limbic Aβ deposits and neurofibrillary tangles at
autopsy to qualify for a diagnosis of AD, reveal levels of
inflammatory markers that are significantly greater than in typical
ND controls but markedly reduced compared with AD patients.
The mammalian target of rapamycin (mTOR) is a
phosphatidylinositol kinase-related serine-threonine kinase
(5) that modulates the activity of
ribosomal S6, nuclear factor κB (NFκB) and inhibitor of NFκB kinase
subunit β (IKKβ) via phosphorylation. As a consequence, mTOR
initiates translation and affects cell growth and proliferation.
Rapamycin (RAPA) is a macrolide antibiotic that was initially
developed as an antifungal agent, which has potent
immunosuppressive and antiproliferative properties (5), and is currently used to prevent
rejection in organ transplantation, particularly lung and kidney
(6). Lipopolysaccharide (LPS),
which is a component of the outer membrane of Gram-negative
bacteria, activates glial cells, inducing inflammation in the
periphery and in the central nervous system (CNS) (7). A local injection of LPS has been
demonstrated to induce inflammation in numerous tissue types
(8,9).
The present study examined the effects of RAPA on
the LPS-stimulated inflammatory response in SH-SY5Y human
neuroblastoma cells. In addition, the effects of RAPA treatment,
acting via mTOR signaling pathways on traumatic brain injury
recovery in a rat model of neuroinflammation, were
investigated.
Materials and methods
RAPA preparation
RAPA (Sigma-Aldrich, St. Louis, MO, USA) was
dissolved in deionized water (10 mg/ml) and stored at −20°C. For
in vivo experiments, this stock solutionwas diluted to 1
mg/ml immediately prior to intragastric administration. Rats in the
treatment group received RAPA (1 mg/kg) and control animals
received normal saline (NS), 30 min prior to surgery (10).
Cell culture and drug treatment
SH-SY5Y human neuroblastoma cells were obtained from
the Cell Bank of the Chinese Academy of Sciences (Shanghai, China).
The cells were cultured at 37°C in a humidified incubator
containing 5% CO2 in Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 15% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 2 mM L-glutamine. The cells were maintained
in the logarithmic phase of growth and subcultured every 3–4 days.
Prior to experimentation, the medium was removed and replaced with
0.01% trypsin-ethylenediaminetetraacetic acid (EDTA) (Gibco; Thermo
Fisher Scientific, Inc.), which had been warmed to 37°C. The cell
layer was completely covered and incubated at 37°C for ~3 min to
allow detachment of the cells. Subsequently, complete medium
(containing 0.02–0.03% trypsin) was added and the cell suspension
was transferred to a clean tube. The cells were centrifuged at 200
× g for 5 min at room temperature, the supernatant was
discarded and the cells were resuspended in fresh medium.
Experiments were performed 24 h later. The medium was removed and
replaced with medium supplemented with FBS containing LPS (20
µg/ml) or LPS (20 µg/ml) and RAPA (0.1, 1.0 or 10 nmol/ml). Cells
were incubated for 6, 12 or 24 h, and were subsequently digested
with trypsin for reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) and western blot analysis.
RT-qPCR
Total RNA samples were isolated from SH-SY5Y cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
total RNA products (1 µg) were immediately transcribed into cDNA
using a PrimeScript™RT Reagent kit with Genomic DNA Eraser (Takara
Biotechnology Co., Ltd.). qPCR was performed on a Chromo4
Four-Color Real-Time PCR Detection system (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) using the SYBR® Premix Ex
Taq™II (Tli RNaseH Plus) kit (Takara Biotechnology Co., Ltd.).
Cycling conditions were as follows: Initial denaturation at 95°C
for 30 sec, followed by 40 cycles of 95°C for 5 sec, 60°C for 20
sec and 72°C for 15 sec. Data was normalized using the
2−ΔΔCq method (11).
Primers were synthesized by Thermo Fisher Scientific, Inc.
(Table I).
 | Table I.Primer sequences. |
Table I.
Primer sequences.
|
|
| Sequence
(5′-3′) |
|
|---|
|
|
|
|
|
|---|
| Target gene | GenBank accession
no. | Forward | Reverse | Product size
(bp) |
|---|
| GAPDH | NM_017008.4 |
CAGGGCTGCCTTCTCTTGTG |
GATGGTGATGGGTTTCCCGT | 172 |
| IL6 | NM_012589.2 |
AGAGACTTCCAGCCAGTTGC |
AGTCTCCTCTCCGGACTTGT | 85 |
| IL1β | NM_031512.2 |
CCTTGTCGAGAATGGGCAGT |
TTCTGTCGACAATGCTGCCT | 222 |
| HIF1α | NM_024359.1 |
AGCAATTCTCCAAGCCCTCC |
TTCATCAGTGGTGGCAGTTG | 111 |
Western blot analysis
Cells were trypsinized, washed twice with ice-cold
phosphate-buffered saline, lysed in radioimmunoprecipitation assay
buffer (20 mmol/l Tris-HCl, 150 mmol/l NaCl, 1 mmol/l EDTA, 1%
NP-40, 1 mmol/l Na3VO4, 1 mmol/l phenylmethylsulfonyl fluoride, 5
µg/ml leupetin, 5 µg/ml aprotinin and 5 µg/ml chymostatin; pH 7.4)
and were centrifuged at 3,000 × g for 5 min at room
temperature. Lysates were clarified and the protein concentrations
were determined according to the method of Lowry et al
(12), using bovine serum albumin
(Gibco; Thermo Fisher Scientific, Inc.) as the standard. Cellular
proteins (20 µg) were separated by 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and were transferred to
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). Membranes were blocked with 5% non-fat milk in Tris-buffered
saline (TBS) containing 0.05% Tween-20 (TBST) for 2 h at room
temperature. Subsequently, membranes were probed with the following
primary antibodies, diluted 1:1,000, overnight at 4°C: rabbit
anti-NF-κB (catalog no. 8242), rabbit anti-phosphorylated (p)-NF-κB
(catalog no. 3033), rabbit anti-IKKβ (catalog no. 8943), rabbit
anti-p-IKKβ (catalog no. 2694), rabbit anti-p-tau (catalog no.
15013), rabbit anti-p-S6 (catalog no. 4858) and rabbit anti-β-actin
(catalog no. 8457), purchased from Cell Signaling Technology, Inc.,
(Danvers, MA, USA). The membranes were then washed three times for
7 min with TBST and incubated with an anti-rabbit IgG conjugated to
horseradish peroxidase (1:5,000; catalog no. 7074; Cell Signaling
Technology, Inc.) for 1 h at room temperature. Protein bands were
visualized using SignalFire™ Elite Enhanced Chemiluminescence
reagent (Cell Signaling Technology, Inc.). The results were
normalized to β-actin using Image J software version 1.48 (National
Institutes of Health, Bethesda, MD, USA).
Animals and drug treatment
Ethical approval was provided by the Medical Ethics
Committee of Southern Medical University (Guangzhou, China). Female
Sprague Dawley (SD) rats (n=18; age, 6 weeks; weight, 170–180 g),
were purchased from Southern Medical University. Rats were
maintained at a constant humidity and temperature under a 12-h
light/dark cycle with ad libitum access to food and water.
The rats were randomly assigned to sham, LPS and LPS+RAPA groups
(n=6/group). In the LPS + RAPA group, RAPA (1 mg/kg) was
administered by gavage 30 min prior to surgery. Neuroinflammation
was induced as described by Espinosa-Oliva et al (13). Rats were anesthetized with an
intraperitoneal injection of 250 mg/kg ketamine (Zoetis, Inc.,
Florham Park, NJ, USA). Sterile techniques were used and the left
lateral ventricle was reached by stereotaxic surgery. With the
incisor bar placed 3.3 mm below the interaural line (horizontal
zero), the coordinates with respect to bregma for the guide cannula
were 1.0 mm anteroposterior, 1.5 mm lateral and 3.8 mm
dorsoventral. LPS (100 µg/kg) was then injected intracranially in
the LPS and LPS+RAPA groups. The sham group underwent surgery but
received no treatment.
Immunohistochemistry
Rats were anesthetized using 250 mg/kg ketamine and
2% xylazine (10 mg/kg, Sigma-Aldrich) for a duration of 6, 12 or 24
h following surgery, and were subsequently sacrificed by rapid
decapitation. The collected brain tissues were fixed in 4%
paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for 8 h at 0°C.
Fixed brains were paraffin embedded and cut into 2 µm sections
using a microtome (Leica Microsystems GmbH, Wetzlar, Germany).
Sections were briefly washed with TBS and incubated for 30 min in
3% H2O2 to quench endogenous peroxidase activity. The following
primary antibodies: Rabbit anti-HIF1α (1:100; catalog no. sc-10790;
Santa Cruz Biotechnology, Inc., Dallas; Texas; U.S.A) and rabbit
anti-p-S6 (1:100; catalog no. 4858; Cell Signaling Technology,
Inc.) were applied overnight at 4°C. Subsequently, sections were
washed three times with TBS and then incubated with
SignalStain® Boost Immunohistochemistry Detection
reagent (1:50; catalog no. 8114; Cell Signaling Technology, Inc.)
for 1 h at room temperature. Sections were then incubated with
3,3′-diaminobenzidine (ZSGB-BIO, Beijing, China). Finally, the
sections were dehydrated, mounted and examined under a light
microscope (Leica Microsystems GmbH). Positive cells were counted
using Image-Pro Plus software verison 7.0 (Media Cybernetics, Inc.,
Rockville, MD, USA).
Histological assays
Paraffin sections were stained with hematoxylin and
eosin (H&E) according to the manufacturer's protocol. The cells
which were swollen or were abnormally circular cells were defined
as positive cells, and these cells were counted using Image-Pro
Plus software verison 7.0.
Statistical analysis
Data are representative of a minimum of three
independent experiments, each of which were performed in
triplicate. Statistical analyses were conducted using GraphPad
Prism software, version 3.0 (GraphPad Software, Inc., La Jolla, CA,
USA). Data were analyzed using a one-way analysis of variance
followed by Tukey's post hoc test. Data are expressed as the
mean ± standard error. P<0.05 was considered to indicate a
statistically significant difference.
Results
RAPA attenuates LPS-induced increases
in IL1β, HIF1α and IL6 mRNA expression levels
SH-SY5Y cells were cultured with 20 µg/ml LPS and
0.1–10 nmol/ml RAPA for 6, 12 or 24 h. RT-qPCR revealed that IL1β
mRNA expression was significantly reduced following treatment with
RAPA compared with the LPS only group, at 6 (P=0.029; Fig. 1A), 12 (P=0.0011; Fig. 1B) and 24 h (P=0.003; Fig. 1C). In addition, HIF1α (P=0.0229 at
6 h, P=0.0005 at 12 h and P=0.0001 at 24 h vs. LPS alone; Fig. 1D-F) and IL6 (P=0.015 at 6 h,
P=0.005 at 12 h and P=0.0001 at 24 h vs. LPS alone; Fig. 1G-I) mRNA expression levels were
significantly reduced in the RAPA groups compared with the LPS only
group.
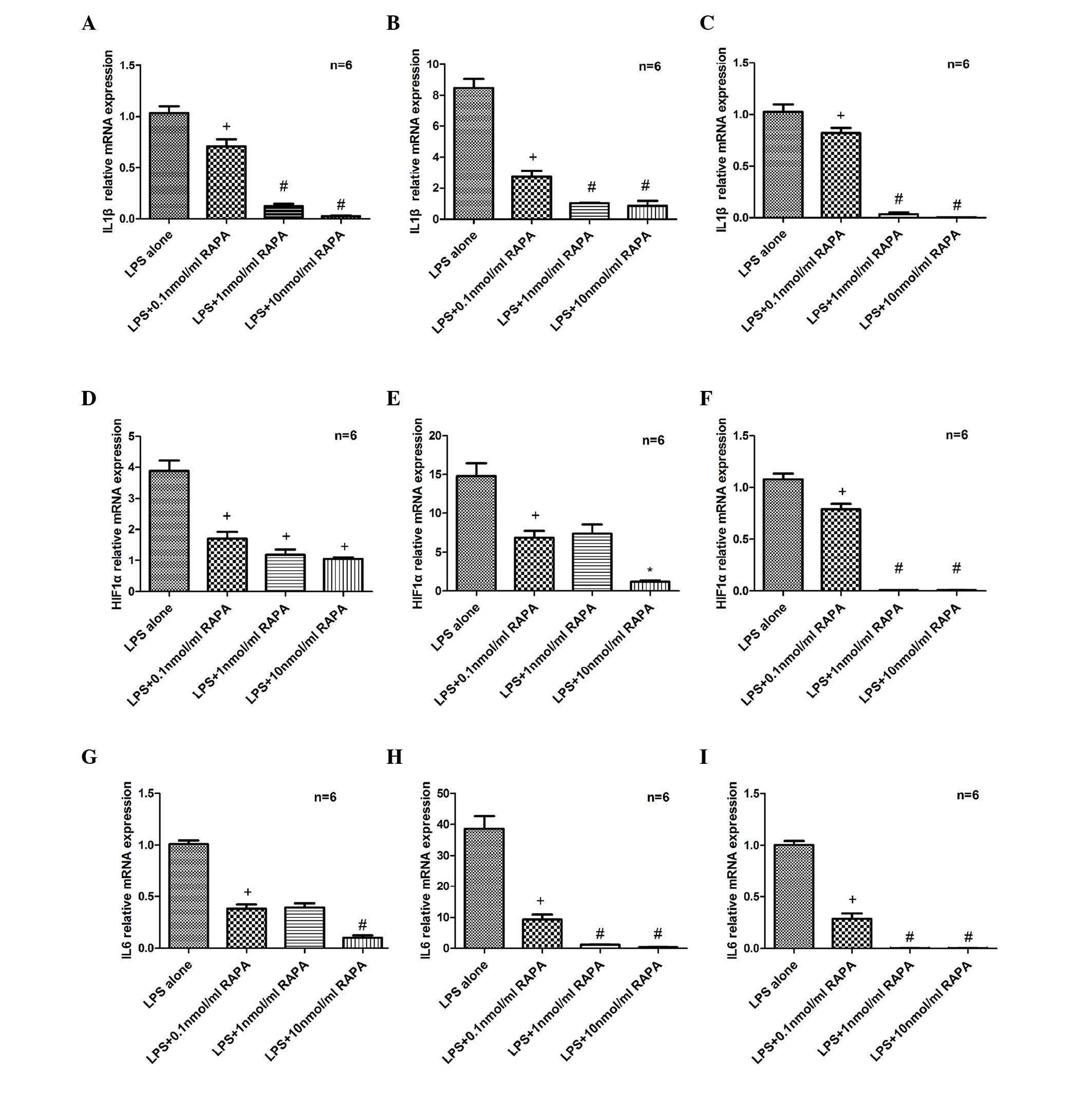 | Figure 1.Effects of RAPA on IL1β, HIF1α and
IL6 mRNA expression. Total RNA was prepared from SH-SY5Y cells
pretreated with LPS (20 µg/ml) for 30 min and exposed to RAPA (0–10
nmol/ml) for 6, 12 and 24 h. RT-qPCR analysis of IL1β following (A)
6, (B) 12 and (C) 24 h of RAPA treatment. RT-qPCR analysis of HIF1α
following (D) 6, (E) 12 and (F) 24 h of RAPA treatment RT-qPCR
analysis of IL6 following (G) 6, (H) 12 and (I) 24 h of RAPA
treatment. Data are presented as fold-change vs. the LPS alone
group. Each sample was measured in triplicate and the experiment
was repeated twice with similar results. Data are expressed as the
mean ± standard error from independent experiments.
+P<0.05 vs. LPS alone; #P<0.05 vs. 0.1
nmol/ml RAPA; and *P<0.05 vs. 1 nmol/ml RAPA. RAPA, rapamycin;
IL, interleukin; HIF1α, hypoxia-inducible factor 1α; LPS,
lipopolysaccharide; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction. |
RAPA attenuate LPS-induced increases
in p-S6, p-tau, p-NFκB and p-IKKB
SH-SY5Y cells were treated with 20 µg/ml LPS and
0.1–10.0 nmol/ml RAPA for 6, 12 or 24 h. Western blot analysis
(Fig. 2A and B) revealed that
expression levels of p-NFκB decreased gradually with increasing
concentrations of RAPA at 6 (P=0.0025; Fig. 2C), 12 (P=0.0011; Fig. 2D) and 24 h (P=0.0014; Fig. 2E). A similar pattern was observed
for p-IKKβ (P=0.0125 at 6 h, P=0.0115 at 12 h and P=0.001 at 24 h
vs. LPS alone; Fig. 2F-H), p-tau
(P=0.019 at 6 h, P=0.025 at 12 h and P=0.003 at 24 h vs. LPS alone;
Fig. 2I-K) and p-S6 (P=0.005 at 6
h, P=0.001 at 12 h and P=0.0005 at 24 h vs. LPS alone; Fig. 2L-N). At 6 h, suppression of p-NFκB
(Fig. 2C), p-IKKβ (Fig. 2F) and p-S6 (Fig. 2L) began and appeared to be
dose-dependent. In addition, the inhibitory effect on p-NFκB lasted
for 12 h (Fig. 2D), and even 24 h
(Fig. 2E). Similarly, expression
of p-IKKβ was decreased at 12 (Fig.
2G) and 24 h (Fig. 2H).
Therefore, critical components of the NFκB signaling pathway,
p-NFκB and p-IKKβ, were suppressed by RAPA and the
neuroinflammation protein, p-tau was decreased. In addition, the
downstream protein of the mTOR signaling pathway, p-S6 was
downregulated by RAPA.
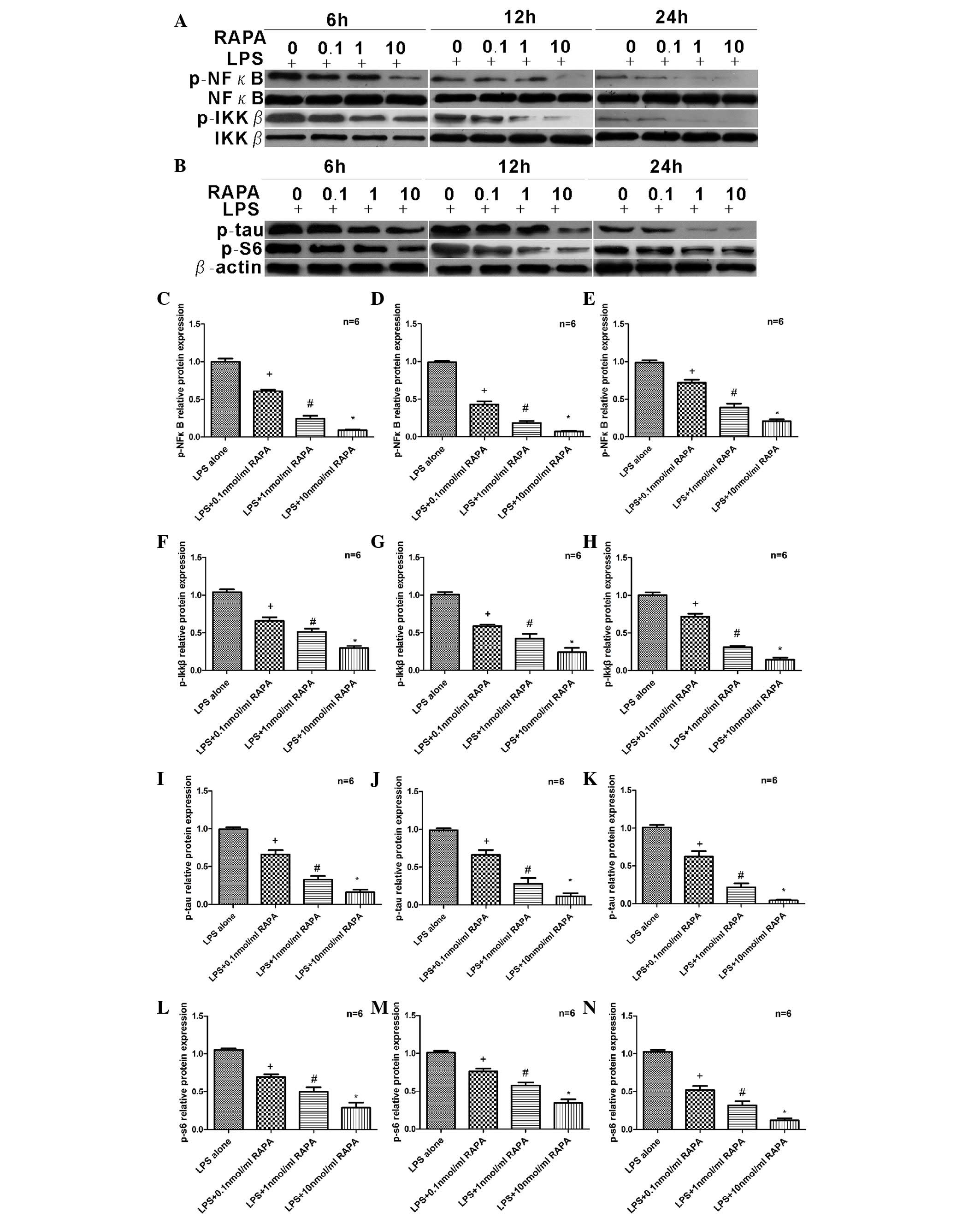 | Figure 2.Effects of RAPA on protein expression
levels of p-NFκB, p-IKKβ, p-tau and p-S6. (A and B) Proteins
extracted from SHSY5Y cells pretreated with 20 µg/ml LPS for 30 min
and exposed to 0–10 nmol/ml RAPA for 6, 12 and 24 h were subjected
to western blot analysis. Protein expression levels of p-NFκB were
analyzed following (C) 6, (D) 12 and (E) 24 h of RAPA treatment.
Protein expression levels of p-IKKβ were analyzed following (F) 6,
(G) 12 and (H) 24 h of RAPA treatment. Protein expression levels of
p-tau were analyzed following (I) 6, (J) 12 and (K) 24 h of RAPA
treatment. Protein expression levels of p-S6 were analyzed
following (L) 6, (M) 12 and (N) 24 h of RAPA treatment. Protein
expression levels of all proteins examined decreased following RAPA
treatment, in a dose-dependent manner, at all time points. Data are
presented as fold-change vs. the LPS alone group. Each sample was
measured in triplicate and the experiment was repeated twice with
similar results. Data are expressed as the mean ± standard error
from independent experiments. +P<0.05 vs. LPS alone;
#P<0.05 vs. 0.1 nmol/ml RAPA; and *P<0.05 vs. 1
nmol/ml RAPA. RAPA, rapamycin; p-, phosphorylated; NFκB, nuclear
factor κB; IKKβ, inhibitor of NFκB kinase subunit β; LPS,
lipopolysaccharide. |
RAPA increases the anti-inflammatory
properties of neurons
Female SD rats were randomly divided into sham, LPS
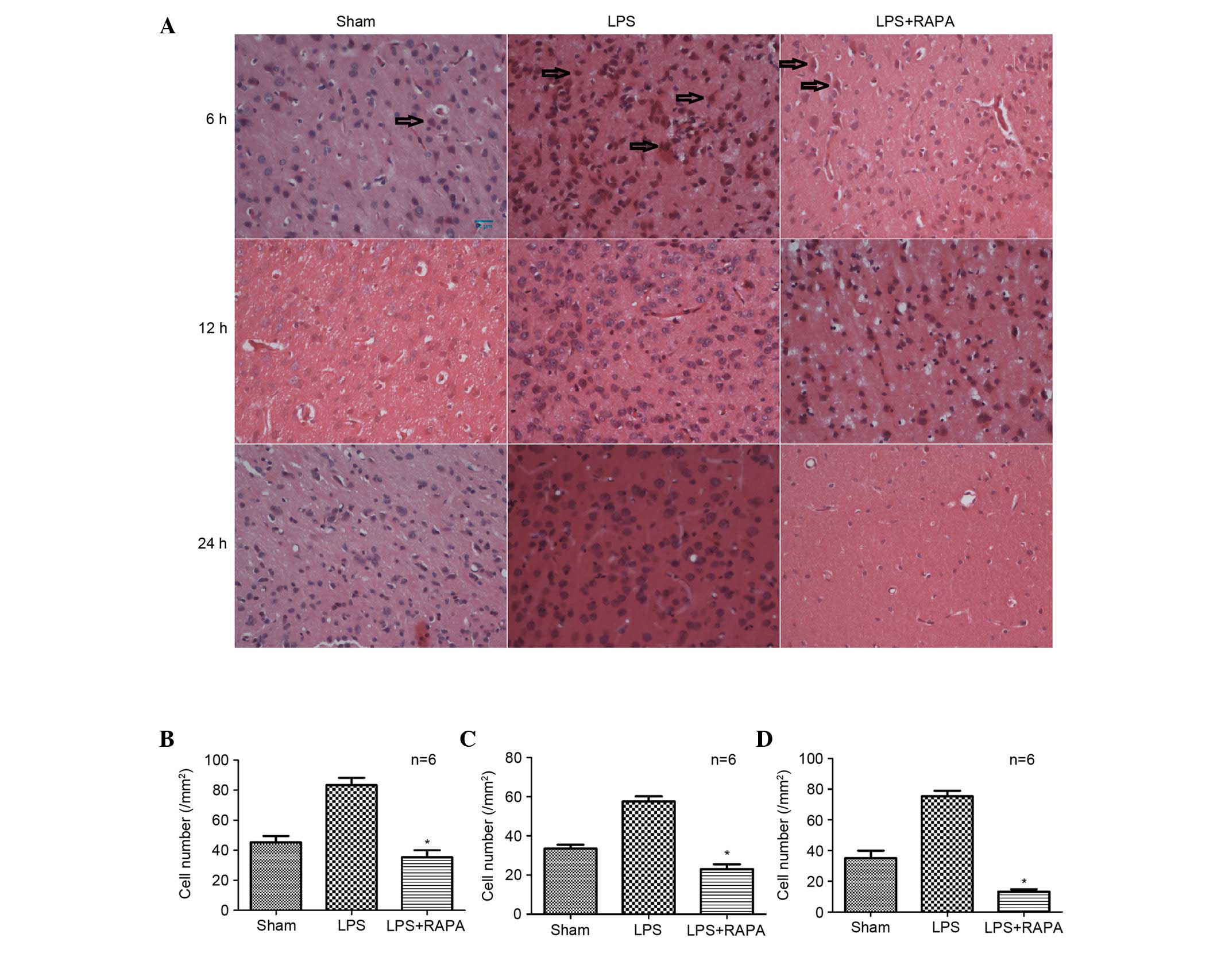
and LPS + RAPA groups. Cerebral cortex sections stained with
H&E revealed that treatment with LPS caused cells to become
swollen and abnormally circular, in a time-dependent manner
(P=0.029 at 6 h, P=0.018 at 12 h and P=0.003 at 24 h vs. sham;
Fig. 3A-D); this damage was
abrogated by RAPA (P=0.009 at 6 h, P=0.001 at 12 h and P=0.0002 at
24 h vs. LPS; Fig. 3A-D). The
swollen and abnormally circular cells within sections were counted
to quantify the LPS-induced inflammation. The inflammatory response
in the neurons increased in the LPS group; however, this increase
was reduced over time in the LPS + RAPA group [6 (Fig. 3B), 12 (Fig. 3C) and 24 h (Fig. 3D)]. Detection of HIF1α (Fig. 4A) and p-S6 (Fig. 4B) by immunohistochemistry
determined that expression levels of HIF1α in rat cerebral cortex
increased following LPS injection (P=0.0021 at 6 h, P=0.0011 at 12
h and P=0.00022 at 24 h vs. sham; Fig.
4C-E); however, this increase was abrogated over time by RAPA
treatment, at 6 (P=0.001 vs. LPS; Fig.
4C), 12 (P=0.005 vs. LPS; Fig.
4D) and 24 h (P=0.003 vs. LPS; Fig. 4E). A similar pattern was observed
for p-S6 expression levels, with increased levels following LPS
injection (P=0.021 at 6 h, P=0.011 at 12 h and P=0.013 at 24 h vs.
sham), abrogated by RAPA treatment (P=0.0029 at 6 h, P=0.0021 at 12
h and P=0.0022 at 24 h vs. LPS; Fig.
4F-H).
Discussion
The present study demonstrated that, following an
LPS-induced neuroinflammatory response, RAPA significantly altered
the expression of key proteins and cytokines, and modulated the
neuronal inflammatory response. It has previously been demonstrated
that RAPA inhibits the mTOR complex 1 (mTORC1) signaling pathway
and prevents phosphorylation of proteins involved in transcription,
translation and cell cycle control (14).
Although inflammation has long been implicated in AD
pathogenesis, its importance has recently become more widely
recognized. Increasing evidence has suggested that inflammation is
critical for the initiation and progression of AD; it has therefore
been identified as a potential therapeutic target. AD has been
associated with polymorphisms in genes involved in inflammation
(15). IL1 is an immunoregulatory
cytokine that has been shown to be overexpressed in affected
regions of the cerebral cortex in patients with AD, as determined
by quantitative assays of tissue IL1 concentrations and analysis of
IL1-immunoreactive microglia (16,17).
IL6 is a pleiotropic cytokine that mediates inflammation affecting
the growth and differentiation of cells in the CNS (18,19).
Although IL6 is widely accepted to be involved in various CNS
disorders, there is evidence to suggest that, in certain
circumstances, IL6 may have anti-inflammatory effects as well as
other beneficial properties. For example, IL6 has been demonstrated
to be critical in the regulation of neuronal survival and function
(20–24). In the present study, RT-qPCR
analyses revealed that IL1β and IL6 mRNA expression levels were
significantly suppressed by RAPA treatment. These
anti-neuroinflammatory effects may provide neuroprotection.
HIF1α, which is the functional subunit of HIF1, is
typically activated under conditions of hypoxia (25,26).
HIF-1 has been demonstrated to be an essential transcription factor
that mediates oxygen homeostasis and exhibits an adaptive response
to the pathological conditions that result from hypoxia (27), upregulating genes that protect
against cerebrovascular diseases and neurodegenerative disorders
(28). A previous study suggested
that HIF1α may be a novel therapeutic target for the treatment of
cerebral ischemia (29). When
inflammation occurs under conditions of hypoxia, an increase in the
mRNA expression levels of HIF1α and the activation of mTOR have
been observed (30,31). Using RT-qPCR and
immunohistochemistry, the present study demonstrated that in the
presence of RAPA, the HIF1α levels were markedly decreased in
vivo and in vitro from 6 to 24 h. This may downregulate
the expression of genes responsible for protection against cerebral
inflammatory injury, and thus provide neuroprotection. Certain
previous studies have demonstrated that myeloid-specific deletion
of HIF-1α impaired inflammatory responses, which was associated
with a metabolic defect characterized by lower glycolytic rates and
energy generation in the absence of HIF-1α (32,33).
mTOR integrates signaling pathways that respond to
growth factors, energy metabolism, nutrients and in addition,
stress (34). Postmortem evidence
from the brains of patients with AD indicated that expression
levels of p-S6 are increased compared with brains from age-matched
controls, thus suggesting that mTORC1 activity is elevated in AD
brains (35–41) and that events downstream of mTORC1
are involved in AD (42). The
increase in mTOR-dependent signaling has also been positively and
significantly correlated with total tau and p-tau expression
levels, and the decades-long continuous synthesis of tau in
degenerating neurons resulted from upregulated mTOR-dependent
signaling (40). Evidence has
indicated a direct association between the neuropathological
effects of Aβ and activation of the NFκB signaling cascade
(43). Aβ induces the accumulation
of inhibitor of κB, a downstream transcriptional target of the NFκB
signaling pathway. The results of the present study revealed that
RAPA inhibited the expression of the mTORC1 downstream target,
p-S6, and the NFκB downstream targets, p-NFκB and p-IKKβ, in
addition to suppressing p-tau protein expression levels. This is
indicative of anti-neurodegenerative effects mediated by RAPA via
mTORC1 and the downstream NFκB signaling pathway (Fig. 5).
Activated microglia produce cytotoxic molecules,
including nitric oxide, oxygen radicals, arachidonic acid
derivatives and in addition, cytokines (1,15,44–46).
Numerous mechanisms may underlie the contribution of post-traumatic
inflammation to neuronal damage (10); the anti-inflammatory effects of
early RAPA treatment may reduce microglial activation and enhance
recovery from head trauma. RAPA is non-cytotoxic in vivo and
may be used as an anti-angiogenic agent to inhibit tumor growth
(6). The lack of toxicity of RAPA
adds weight to the importance of the neuroprotective effects of
RAPA revealed in the present study.
In conclusion, the results of the present study
demonstrated that RAPA suppresses the neuroinflammation induced by
LPS, in vitro and in vivo. RAPA may therefore be a
potential therapeutic agent for the treatment of early AD, or for
the treatment of neuroinflammation in non-AD patients to prevent
the onset of AD. However, further studies are required to confirm
the suitability of RAPA as a treatment strategy.
Acknowledgements
The authors would like to thank Mr. Sheng-fa Li
(Huzihou First Hospital, Huizhou, China) and Mr. Wen-ji Zhao (Inner
Mongolia People's Hospital, Hohhot, China) for providing reagents,
instructive suggestions and valuable comments.
References
|
1
|
Giulian D: Microglia and the immune
pathology of Alzheimer disease. Am J Hum Genet. 65:13–18. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Alzheimer's Association: 2014 Alzheimer's
disease facts and figures. Alzheimers Dement. 10:e47–e92. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Golde TE, Petrucelli L and Lewis J:
Targeting Abeta and tau in Alzheimer's disease, an early interim
report. Exp Neurol. 223:252–266. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Veerhuis R, Janssen I, De Groot CJ, Van
Muiswinkel FL, Hack CE and Eikelenboom P: Cytokines associated with
amyloid plaques in Alzheimer's disease brain stimulate human glial
and neuronal cell cultures to secrete early complement proteins,
but not C1-inhibitor. Exp Neurol. 160:289–299. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baker H, Sidorowicz A, Sehgal SN and
Vézina C: Rapamycin (AY-22,989), a new antifungal antibiotic. III.
In vitro and in vivo evaluation. J Antibiot (Tokyo). 31:539–545.
1978. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Majumder PK, Febbo PG, Bikoff R, Berger R,
Xue Q, McMahon LM, Manola J, Brugarolas J, McDonnell TJ, Golub TR,
et al: mTOR inhibition reverses Akt-dependent prostate
intraepithelial neoplasia through regulation of apoptotic and
HIF-1-dependent pathways. Nat Med. 10:594–601. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bonow RH, Aïd S, Zhang Y, Becker KG and
Bosetti F: The brain expression of genes involved in inflammatory
response, the ribosome, and learning and memory is altered by
centrally injected lipopolysaccharide in mice. Pharmacogenomics J.
9:116–126. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hauss-Wegrzyniak B, Lukovic L, Bigaud M
and Stoeckel ME: Brain inflammatory response induced by
intracerebroventricular infusion of lipopolysaccharide: An
immunohistochemical study. Brain Res. 794:211–224. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Burguillos MA, Hajji N, Englund E, Persson
A, Cenci AM, Machado A, Cano J, Joseph B and Venero JL:
Apoptosis-inducing factor mediates dopaminergic cell death in
response to LPS-induced inflammatory stimulus: Evidence in
Parkinson's disease patients. Neurobiol Dis. 41:177–188. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Erlich S, Alexandrovich A, Shohami E and
Pinkas-Kramarski R: Rapamycin is a neuroprotective treatment for
traumatic brain injury. Neurobiol Dis. 26:86–93. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lowry OH, Rosebrough NJ, Farr AL and
Randall RJ: Protein measurement with the Folin phenol reagent. J
Biol Chem. 193:265–275. 1951.PubMed/NCBI
|
|
13
|
Espinosa-Oliva AM, de Pablos RM and
Herrera AJ: Intracranial injection of LPS in rat as animal model of
neuroinflammation. Methods Mol Biol. 1041:295–305. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vignot S, Faivre S, Aguirre D and Raymond
E: mTOR-targeted therapy of cancer with rapamycin derivatives. Ann
Oncol. 16:525–537. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Akiyama H, Barger S, Barnum S, Bradt B,
Bauer J, Cole GM, Cooper NR, Eikelenboom P, Emmerling M, Fiebich
BL, et al: Inflammation and Alzheimer's disease. Neurobiol aging.
21:383–421. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Griffin WS, Sheng JG, Roberts GW and Mrak
RE: Interleukin-1 expression in different plaque types in
Alzheimer's disease: Significance in plaque evolution. J
Neuropathol Exp Neurol. 54:276–281. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Griffin WS, Sheng JG, Royston MC,
Gentleman SM, McKenzie JE, Graham DI, Roberts GW and Mrak RE:
Glial-neuronal interactions in Alzheimer's disease: The potential
role of a ‘cytokine cycle’ in disease progression. Brain Pathol.
8:65–72. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Heinrich PC, Horn F, Graeve L, Dittrich E,
Kerr I, Müller-Newen G, Grötzinger J and Wollmer A: Interleukin-6
and related cytokines: Effect on the acute phase reaction. Z
Ernahrungswiss (37 Suppl 1). 43–49. 1998.
|
|
19
|
Hirano T, Nakajima K and Hibi M: Signaling
mechanisms through gp130: A model of the cytokine system. Cytokine
Growth Factor Rev. 8:241–252. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Campbell IL: Transgenic mice and cytokine
actions in the brain: Bridging the gap between structural and
functional neuropathology. Brain Res Brain Res Rev. 26:327–336.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gadient RA and Otten UH: Interleukin-6
(IL-6)-a molecule with both beneficial and destructive potentials.
Prog Neurobiol. 52:379–390. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
März P, Cheng JG, Gadient RA, Patterson
PH, Stoyan T, Otten U and Rose-John S: Sympathetic neurons can
produce and respond to interleukin 6. Proc Natl Acad Sci USA.
95:3251–3256. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rodriguez M, Pavelko KD, McKinney CW and
Leibowitz JL: Recombinant human IL-6 suppresses demyelination in a
viral model of multiple sclerosis. J Immunol. 153:3811–3821.
1994.PubMed/NCBI
|
|
24
|
Yang P, Wen H, Ou S, Cui J and Fan D: IL-6
promotes regeneration and functional recovery after cortical spinal
tract injury by reactivating intrinsic growth program of neurons
and enhancing synapse formation. Exp Neurol. 236:19–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jeong YJ, Cho HJ, Magae J, Lee IK, Park KG
and Chang YC: Ascofuranone suppresses EGF-induced HIF-1α protein
synthesis by inhibition of the Akt/mTOR/p70S6K pathway in
MDA-MB-231 breast cancer cells. Toxicol Appl Pharmacol.
273:542–550. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shin JM, Jeong YJ, Cho HJ, Park KK, Chung
IK, Lee IK, Kwak JY, Chang HW, Kim CH, Moon SK, et al: Melittin
suppresses HIF-1α/VEGF expression through inhibition of ERK and
mTOR/p70S6K pathway in human cervical carcinoma cells. PloS one.
8:e693802013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
White NM, Masui O, Newsted D, Scorilas A,
Romaschin AD, Bjarnason GA, Siu KW and Yousef GM: Galectin-1 has
potential prognostic significance and is implicated in clear cell
renal cell carcinoma progression through the HIF/mTOR signaling
axis. Br J Cancer. 110:1250–1259. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Singh N, Sharma G and Mishra V: Hypoxia
inducible factor-1: Its potential role in cerebral ischemia. Cell
Mol Neurobiol. 32:491–507. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Umschweif G, Alexandrovich AG, Trembovler
V, Horowitz M and Shohami E: Hypoxia-inducible factor 1 is
essential for spontaneous recovery from traumatic brain injury and
is a key mediator of heat acclimation induced neuroprotection. J
Cereb Blood Flow Metab. 33:524–531. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Argyriou P, Papageorgiou SG, Panteleon V,
Psyrri A, Bakou V, Pappa V, Spathis A, Economopoulou P,
Papageorgiou E, Economopoulos T and Rontogianni D:
Hypoxia-inducible factors in mantle cell lymphoma: Implication for
an activated mTORC1->HIF-1α pathway. Ann Hematol. 90:315–322.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Weng Q, Zhang J, Cao J, Xia Q, Wang D, Hu
Y, Sheng R, Wu H, Zhu D, Zhu H, et al: Q39, a quinoxaline
1,4-Di-N-oxide derivative, inhibits hypoxia-inducible factor-1α
expression and the Akt/mTOR/4E-BP1 signaling pathway in human
hepatoma cells. Invest New Drugs. 29:1177–1187. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Palazon A, Goldrath AW, Nizet V and
Johnson RS: HIF transcription factors, inflammation, and immunity.
Immunity. 41:518–528. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cramer T, Yamanishi Y, Clausen BE, Förster
I, Pawlinski R, Mackman N, Haase VH, Jaenisch R, Corr M and Nizet
V: HIF-1alpha is essential for myeloid cell-mediated inflammation.
Cell. 112:645–657. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Laplante M and Sabatini DM: mTOR signaling
at a glance. J Cell Sci. 122:3589–3594. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
An WL, Cowburn RF, Li L, Braak H,
Alafuzoff I, Iqbal K, Iqbal IG, Winblad B and Pei JJ: Up-regulation
of phosphorylated/activated p70 S6 kinase and its relationship to
neurofibrillary pathology in Alzheimer's disease. Am J Pathol.
163:591–607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chang RC, Wong AK, Ng HK and Hugon J:
Phosphorylation of eukaryotic initiation factor-2alpha (eIF2alpha)
is associated with neuronal degeneration in Alzheimer's disease.
Neuroreport. 13:2429–2432. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Griffin RJ, Moloney A, Kelliher M,
Johnston JA, Ravid R, Dockery P, O'Connor R and O'Neill C:
Activation of Akt/PKB, increased phosphorylation of Akt substrates
and loss and altered distribution of Akt and PTEN are features of
Alzheimer's disease pathology. Journal of Neurochemistry.
93:105–117. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Onuki R, Bando Y, Suyama E, Katayama T,
Kawasaki H, Baba T, Tohyama M and Taira K: An RNA-dependent protein
kinase is involved in tunicamycin-induced apoptosis and Alzheimer's
disease. EMBO J. 23:959–968. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peel AL and Bredesen DE: Activation of the
cell stress kinase PKR in Alzheimer's disease and human amyloid
precursor protein transgenic mice. Neurobiol Dis. 14:52–62. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pei JJ, Björkdahl C, Zhang H, Zhou X and
Winblad B: p70 S6 kinase and tau in Alzheimer's disease. J
Alzheimers Dis. 14:385–392. 2008.PubMed/NCBI
|
|
41
|
Pei JJ and Hugon J: mTOR-dependent
signalling in Alzheimer's disease. J Cell Mol Med. 12:2525–2532.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lipton JO and Sahin M: The neurology of
mTOR. Neuron. 84:275–291. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tan L, Schedl P, Song HJ, Garza D and
Konsolaki M: The Toll->NFkappaB signaling pathway mediates the
neuropathological effects of the human Alzheimer's Abeta42
polypeptide in Drosophila. PloS one. 3:e39662008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Garwood CJ, Cooper JD, Hanger DP and Noble
W: Anti-inflammatory impact of minocycline in a mouse model of
tauopathy. Front Psychiatry. 1:1362010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Walker D and Lue LF: Anti-inflammatory and
immune therapy for Alzheimer's disease: Current status and future
directions. Curr Neuropharmacol. 5:232–243. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu B, Gao HM and Hong JS: Parkinson's
disease and exposure to infectious agents and pesticides and the
occurrence of brain injuries: Role of neuroinflammation. Environ
Health Perspect. 111:1065–1073. 2003. View Article : Google Scholar : PubMed/NCBI
|