Introduction
Cytochrome P450 (CYP) are a family of
hemoproteins involved in multiple enzymatic reactions, such as drug
metabolism. Previous studies have indicated that gene polymorphisms
may be a major cause of the variability in drug metabolism that
exists among individuals. The CYP3A subfamilies are the most
abundant CYPs in the human liver and intestine. CYP3A
catalyzes the metabolism of drugs, and contributes to variations in
the bioavailability and clearance of CYPA3 substrates
(1–4). CYP3A4 is the most abundant
hepatic and intestinal CYP3A isoform. Researchers previously
thought that CYP3A4 was the primary contributor to
metabolism within this family, however, this view was recently
challenged by data that indicated that the relative contribution of
CYP3A5 may represent >50% of total CYP3A in
certain individuals and may exert a greater metabolic action than
previously considered in individuals that possess at least one
polymorphic CYP3A5 allele (3,5,6). The
wild type allele of CYP3A5 is defined as
CYP3A5*1. Mutations include the following:
CYP3A5*2; CYP3A5*3A; CYP3A5*3C;
CYP3A5*4; CYP3A5*5; CYP3A5*6; and
CYP3A5*7. The penetrance of polymorphic CYP3A5
genotypes varies among different ethnic groups and polymorphic
CYP3A5 genotypes have varying pharmacokinetics (PK) and
pharmacodynamics (PD) among CYP3A substrates, which include
the calcium channel blocker, amlodipine (7,8). The
effects of amlodipine vary depending on which CYP3A5 allele
an individual has. For example, among Korean and Chinese patients
that were treated with amlodipine, homozygous
CYP3A5*3 carriers (2,9)
exhibited a greater oral clearance of amlodipine and reduced mean
area under the plasma concentration-time curve compared with
CYP3A5*1 carriers (10–12).
Additionally, Zhang et al (2) demonstrated that
CYP3A5*3/*3 patients exhibited lower plasma
levels of amlodipine compared with CYP3A5*1 patients,
which indicated that patients with polymorphisms may have faster
amlodipine metabolism and increased CYP3A enzyme activity
(2). This heterogeneity in enzyme
action indicates the requirement for a reliable method to assess
the impact of polymorphisms in amlodipine metabolism. To
investigate the impact of CYP3A5 genetic polymorphisms on
amlodipine metabolism, the present study enrolled 57 Chinese
hypertensive patients with four different polymorphisms:
CYP3A5*1/*1 (n=7);
CYP3A5*1/*3 (n=28); CYP3A5*4
(n=2); and CYP3A5*6 (n=2). Changes in the blood
pressure of patients following administration of amlodipine were
recorded. In addition, a well-investigated and cost-effective
Saccharomyces cerevisiae expression system was constructed
to validate the results obtained in the in vivo
assessment.
Materials and methods
Patients and study design
The study protocol and consent for prospectively
enrolled patients were approved by the Ethics Committee of the
Third Xiangya Hospital (Changsha, China). Informed consent was
obtained from all individual participants included in the study.
The study included 57 hypertensive patients that were assigned to
receive amlodipine from a single center in China between May 2012
and February 2013. Patients were included in the study if they were
between the ages of 18 and 70, and diagnosed with a systolic blood
pressure ≥140 mmHg or a diastolic blood pressure ≥90 mmHg. Patients
were excluded if they exhibited any of the following complications:
Cerebrovascular accident history, heart failure, serious organic
heart disease, large aneurysm and aortic dissection, second degree
atrioventricular block, sick sinus syndrome, arrhythmia of clinical
significance, ventricular tachycardia, atrial fibrillation or
flutter, untreated thyroid disease, gastrointestinal lesions or
prior gastrointestinal surgeries that may affect drug absorption,
history of malignancy or any liver, kidney, heart and lung function
congruent with a life-threatening illness, liver dysfunction,
allergy to amlodipine, pregnancy or lactation, mental illness,
active use of other antihypertensive drugs during the trial period
or participating in other clinical trials within 3 months of
enrolling in the current study. The suitability of all subjects was
determined using medical history, routine physical examinations,
clinical laboratory testing, and measuring the heart rate and blood
pressure. Following a 1-week washout period, baseline blood
pressure values were recorded. All subjects received a single oral
dose of 5 mg/day amlodipine (one 5 mg tablet; Pfizer Inc., New
York, NY, USA) with water (200 ml) in the morning for 4 weeks. A
series of 5 ml venous blood samples were collected into heparinized
tubes from the antecubital vein of patients. Aliquots of plasma (2
ml) isolated from the blood were stored at −80°C until analysis.
The blood samples were collected at 0, 24, 48, 72 and 120 h, and
the plasma concentrations of amlodipine were measured through 4
weeks following the blood sample collection. The blood pressure of
patients was measured with the calibrated Electronic Blood Pressure
Monitor (HEM-7012; Omron Corporation, Kyoto, Japan) at 0 h on days
0 and 28 of drug treatment. DNA was extracted from patient
peripheral blood samples prior to the treatment. Genotypes for the
CYP3A5 polymorphisms were determined by polymerase chain reaction
(PCR)-restriction fragment length polymorphism as previously
described (13). Blood drug
concentrations of each CYP3A5 genotype group was measured with
liquid chromatography-mass spectrometry (LC-MS) (14,15)
and high performance liquid chromatography (HPLC) (16). The metabolic rate of amlodipine was
calculated in each group.
Construction of CYP3A5 gene
polymorphism expression vectors
The pYES2/CT vector was purchased from Invitrogen
(Thermo Fisher Scientific, Inc.). All primers used in this study
are presented in Table I (Shenzhen
Huada Gene Science and Technology Co., Ltd., Shenzhen, China)
(5). The total CYP3A5 RNA was
extracted from human liver cells (collected from a 33 year old male
patient who had undergone surgery and signed the informed consent)
following the Qiagen RNeasy Mini Kit (Qiagen, GmbH, Hilden,
Germany) protocol. The tissue sections were homogenized in RLT
buffer supplemented with 2 M dithiothreitol using a 1 ml syringe
and a needle. Homogenized cells were applied to an RNeasy spin
column, washed, and then eluted in 50 ml of water and stored at
−20°C until required. cDNA was obtained via RNA reverse
transcription using a reverse transcription kit (Qiagen, GmbH)
according to the manufacture's protocol. cDNA fragments were
amplified using PCR as presented in Tables II and III, according to the manufacturer's
protocol.
 | Table I.Primers used to construct the
CYP3A5 gene polymorphism expression vectors in this
study. |
Table I.
Primers used to construct the
CYP3A5 gene polymorphism expression vectors in this
study.
| Primer | Primer sequence
(5′-3′) |
|---|
| CYP3A5 | F:
aagcttATGGACCTCATCCCAAAT (Hind III) |
|
| R:
ctcgagTTCTCCACTTAGGGTTCC (Xho I) |
| CYP3A5*4 | F:
ATCCACGAGACCCCTTTGTGGAGAG |
|
| CAC |
|
| R:
AGGGGTCTCGTGGATTGTTGAGAGA |
|
| GTC |
| CYP3A5*6 | F:
CACTAAAAAGTTCCTAAAATTTGGTT |
|
| TCT |
|
| R:
GAACTTTTTAGTGCTCTCCACAAAGG |
|
| GGT |
| Table II.Polymerase chain reaction system. |
Table II.
Polymerase chain reaction system.
| Reagents | volume (µ1) |
|---|
| 10x LAmp
buffer | 2.5 |
| dNTP | 2 |
| 5x C solution | 5 |
| CYP3A5-CDS-F (10
uM) | 1 |
| CYP3A5-CDS-R (10
uM) | 1 |
| Template | 1 |
| Lamp DNA
polymerase | 0.25 |
|
ddH2O | Up to 25 |
| Table III.Polymerase chain reaction
condition. |
Table III.
Polymerase chain reaction
condition.
| Temperature
(°C) | Time |
|
|---|
| 94 | 4 min |
|
| 94 | 30 sec |
|
| 56 | 30 sec | 35 cycles |
| 72 | 1.30 min |
|
| 72 | 5 min |
|
The CYP3A5-CDS-F/CYP3A5-CDS-R primers are presented
in Table I, and GAPDH served as a
control. Gel electrophoresis products of PCR were analyzed using
agarose gel electrophoresis. Agarose gels of 2% (w/v) concentration
were made in 0.5 Tris/Borate/EDTA buffer. Loading dye was added to
the PCR products prior to being run at 120 V for 90 min alongside
Hyperladder II (Bioline UK, London England) for comparison.
Ethidium bromide at a concentration of 0.5 mg/ml (CLP, US) was used
to stain the DNA for visualization using a UV transilluminator.
RT-PCR products were alternatively analyzed using a standard
procedure on a Capillary Gel Electrophoresis CEQ 8000 Genetic
Analyser (Beckman Coulter, Inc., Brea, CA, USA).
The human liver cells were cultured in Dulbecco's
modified Eagle's medium (Invitrogen, Thermo Fisher Scientific,
Inc.) supplemented with 20% fetal bovine serum at a temperature of
37°C in an environment containing 5% CO2. PCR products
of the cDNA fragments (Table I)
were purified and ligated into the pMD18-T simple vector
(Invitrogen; Thermo Fisher Scientific, Inc.). The resulting
pMD18-T-CYP3A5 plasmid was transformed into E. coli JM109
(Auragene Bioscience Corporation Inc, Changsha, China). The E.
coli strains carrying the recombinant pMD18-T-CYP3A5 plasmid
were cultured on LB agar supplemented with 50 µg/ml ampicillin at
37°C overnight. A single colony was transferred into 5 ml of LB
medium with ampicillin at the same concentrations used in the solid
medium. The culture was incubated overnight at 37°C and 210 rpm.
Subsequently, plasmids were extracted. Plasmids were double-enzyme
digested with Hind III and Xho I (Thermo Fisher
Scientific. Inc.), and the target gene fragments were inserted into
the pYES2/CT plasmid that was digested with the same endonuclease
enzyme, and the resultant recombinant plasmids were transformed
into E. coli JM109 using the CYP3A5 expression vector and
cultured E. coli JM109 at a temperature of 37°C and a
revolution of 210 rpm/min in a medium containing 50 µg/ml
ampicillin (17). Positive clones
were screened for and the recombinant plasmid, CYP3A5-pYES2/CT, was
extracted. CYP3A5*4 and CYP3A5*6 mutational expression vectors were
constructed using site-specific mutagenesis of the wild type
CYP3A5- pYES2/CT gene (18). The
present study screened for positive clones and extracted the
recombinant plasmid of CYP3A5 gene polymorphism expression vectors.
The recombinant plasmid was transformed into Saccharomyces
cerevisiae INVSc1 (19). The
transformants were selected on Schaeffer's-glucose agar plates at
30°C for 2–4 days. The microsomal CYP3A5 yeast protein was
extracted, western blotting was performed as described previously
(20). The microsomal CYP3A5 yeast
proteins were separated on SDS polyacrylamide gels. Proteins were
probed with the anti his-V5 (Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA). The blots were scanned with an Odyssey imager
(LI-COR Biosciences, Lincoln, NE, USA).
An in vitro study of amlodipine
metabolism by CYP3A5 recombinant enzymes
A primary stock solution of amlodipine was prepared
in 50% methanol at a concentration of 100 µg/ml. Estazolam (Beijing
Yimin Pharmaceutical Co., Ltd., Beijing, China) was prepared from
the primary stock at a concentration of 0.4 µg/ml for use as an
internal standard. Working standard solutions of amlodipine were
prepared from the primary stock at concentrations of 100, 50, 25,
12.5, 5.25, 3.125, 1.5625 and 0.78125 ng/ml. All solutions were
stored at 4°C and warmed to room temperature before use. The
constructed CYP3A5 gene polymorphism systems were used to
metabolize amlodipine. The amount of drug metabolized was detected
using LC-MS and HPLC as previously described (15). Amlodipine solution (50 µl),
internal standard (50 µl), 500 µl blank serum (500 µl) and 1 M
sodium hydroxide (200 µl) were added to 1.5 ml methyl tert-butyl
ether. Following vortexing and centrifugation at a speed of 1,000 ×
g at 4°C for 10 min, the organic phase was dried at 45°C under
nitrogen, reconstituted in methanol (80%, 200 µl) and injected onto
a Phenomenex Luna column C18 (150×2 mm; Phenomenex, Inc., Torrance,
CA, USA). The mobile phase consisted of methanol (0.1% methanoic
acid) and 1 M ammonium acetate (0.1% methanoic acid;
methanol/ammonium acetate, 4/1) with a flow rate of 0.2 ml/min. A
standard curve was fitted using standard concentrations for the
X-axis and the area ratio of standard/internal standard for the
Y-axis. Values for the rate of metabolite formation from 0.2 mg/ml
of the recombinant enzyme were derived using the standard curve.
Km, Vmax and Vmax/Km
values were determined using GraphPad Prism 5.0 (GraphPad Software,
Inc., La Jolla, CA, USA) and the Michaelis-Menten equation
(6,21).
Statistical analysis
Values are presented as the mean ± standard
deviation. Statistical comparisons were performed using one-way
analysis of variance followed by the Newman-Keuls test (GraphPad
Prism 5.0; GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
Differences in amlodipine PD among
CYP3A5 gene polymorphisms
The present study assessed the genotype of 57
hypertensive patients from the Health Management Center of the
Third Xiangya Hospital. The number of patients with each
polymorphism were as follows: CYP3A5*1/*1 in 7 patients (12.3%);
CYP3A5*1/*3 in 28 patients (49.1%); CYP3A5*4 in 2 patients (3.5%);
and CYP3A5*6 in 2 patients (3.5%). The mean age of the patients (26
males and 13 females) was 45.61±2.33 years and the mean body mass
index was 24.15±1.46. Change in blood pressure prior to and
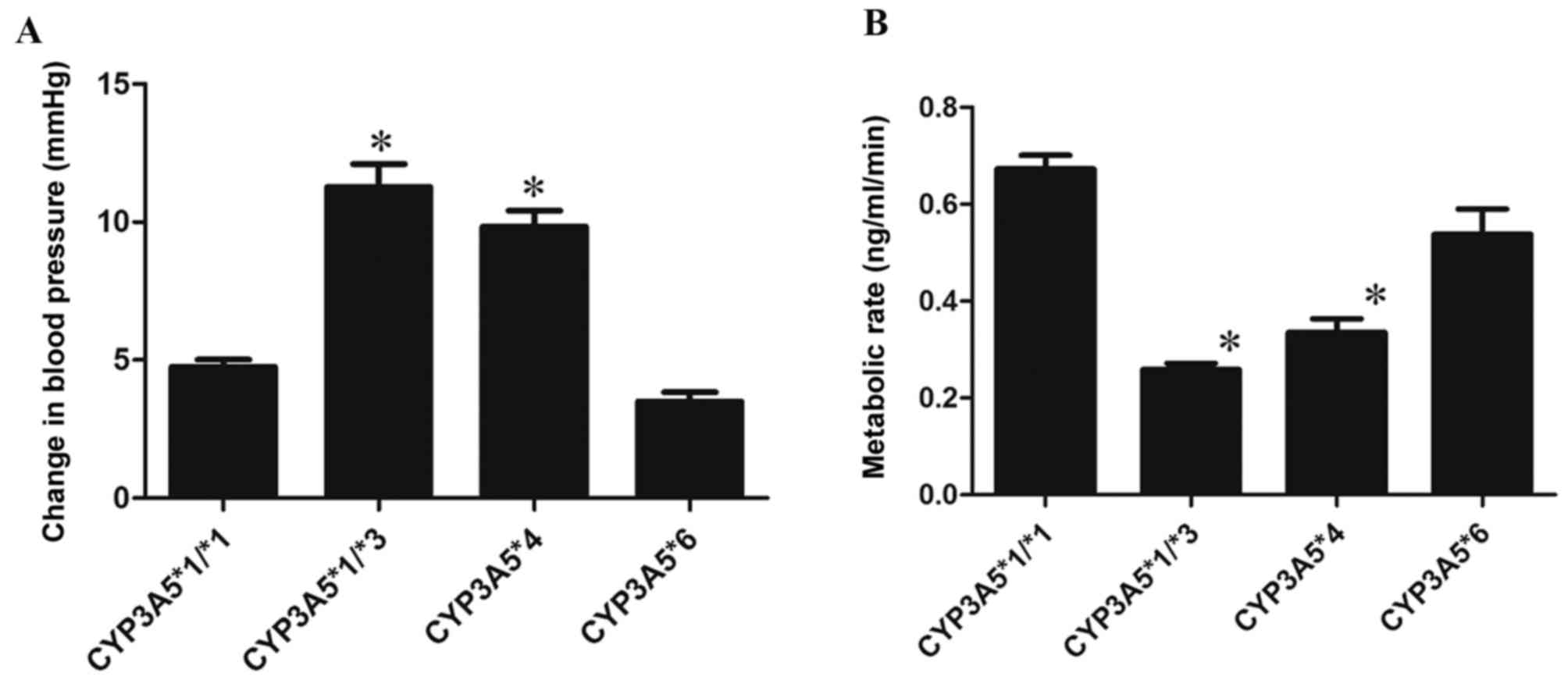
following treatment with amlodipine was also recorded (Table IV). Following administration of
amlodipine, blood pressure in all patients decreased. The magnitude
of the change in blood pressure varied among patients. Individuals
with the CYP3A5*1/*3 polymorphism exhibited the highest change in
blood pressure, followed by CYP3A5*4, CYP3A5*1/*1, and CYP3A5*6
(Fig. 1A).
| Table IV.Antihypertensive effects of CYP3A5
gene polymorphisms. |
Table IV.
Antihypertensive effects of CYP3A5
gene polymorphisms.
|
|
| Systolic BP
(mmHg) | Diastolic BP
(mmHg) |
|---|
|
|
|
|
|
|---|
| Polymorphism | Number of
patients | Prior to amlodipine
administration | Following
amlodipine administration | Difference | Prior to amlodipine
administration | Following
amlodipine administration | Difference |
|---|
| CYP3A5*1/*1 | 7 | 145.21±19.4 | 137.54±17.0 | 7.67±3.2 | 92.94±12.4 | 88.18±11.3 | 4.76±2.5 |
| CYP3A5*1/*3 | 28 | 144.67±17.6 | 127.41±8.3 |
17.27±6.7a | 89.53±12.6 | 78.27±5.9 |
11.27±81.4a |
| CYP3A5*4 | 2 | 156.26±19.3 | 141.16±17.9 |
15.53±10.4a | 94.17±12.2 | 84.93±12.3 |
9.83±5.9a |
| CYP3A5*6 | 2 | 150.29±13.4 | 143.81±13.1 |
6.48±4.7b | 93.03±11.4 | 89.52±11.9 |
3.51±3.4b |
In vivo drug metabolism of
amlodipine
The differences in amlodipine PK among different
CYP3A5 gene polymorphisms were compared. As presented in
Table V, CYP3A5 recombinant
enzymes influenced the PK of amlodipine in hypertensive patients.
There were differences in the metabolic rate among the
polymorphisms; individuals with the CYP3A5*6 polymorphism
exhibited the highest metabolic ratio of amlodipine, followed by
CYP3A5*1/*1, CYP3A5*4 and CYP3A5*1/*3.
Compared with CYP3A5*1/*1, there were significant
differences in CYP3A5*1/*3 and CYP3A5*4 (P<0.05).
There was no statistically significant difference between
CYP3A5*6 and CYP3A5*1/*1 (P=0.5; Fig. 1B). These results may explain the
differences in the antihypertensive effect of amlodipine in
individuals with different CYP3A5 polymorphisms.
| Table V.Amlodipine metabolic ratio of CYP3A5
gene polymorphisms. |
Table V.
Amlodipine metabolic ratio of CYP3A5
gene polymorphisms.
| Polymorphism | Number of
patients | V max
(ng/ml/min) |
|---|
| CYP3A5*1/*1 | 7 | 0.6727±0.028 |
| CYP3A5*1/*3 | 28 |
0.2584±0.013a |
| CYP3A5*4 | 2 |
0.3347±0.029a |
| CYP3A5*6 | 2 | 0.7174±0.053 |
Construction and verification of the
Saccharomyces cerevisiae expression system for CYP3A5 gene
polymorphisms
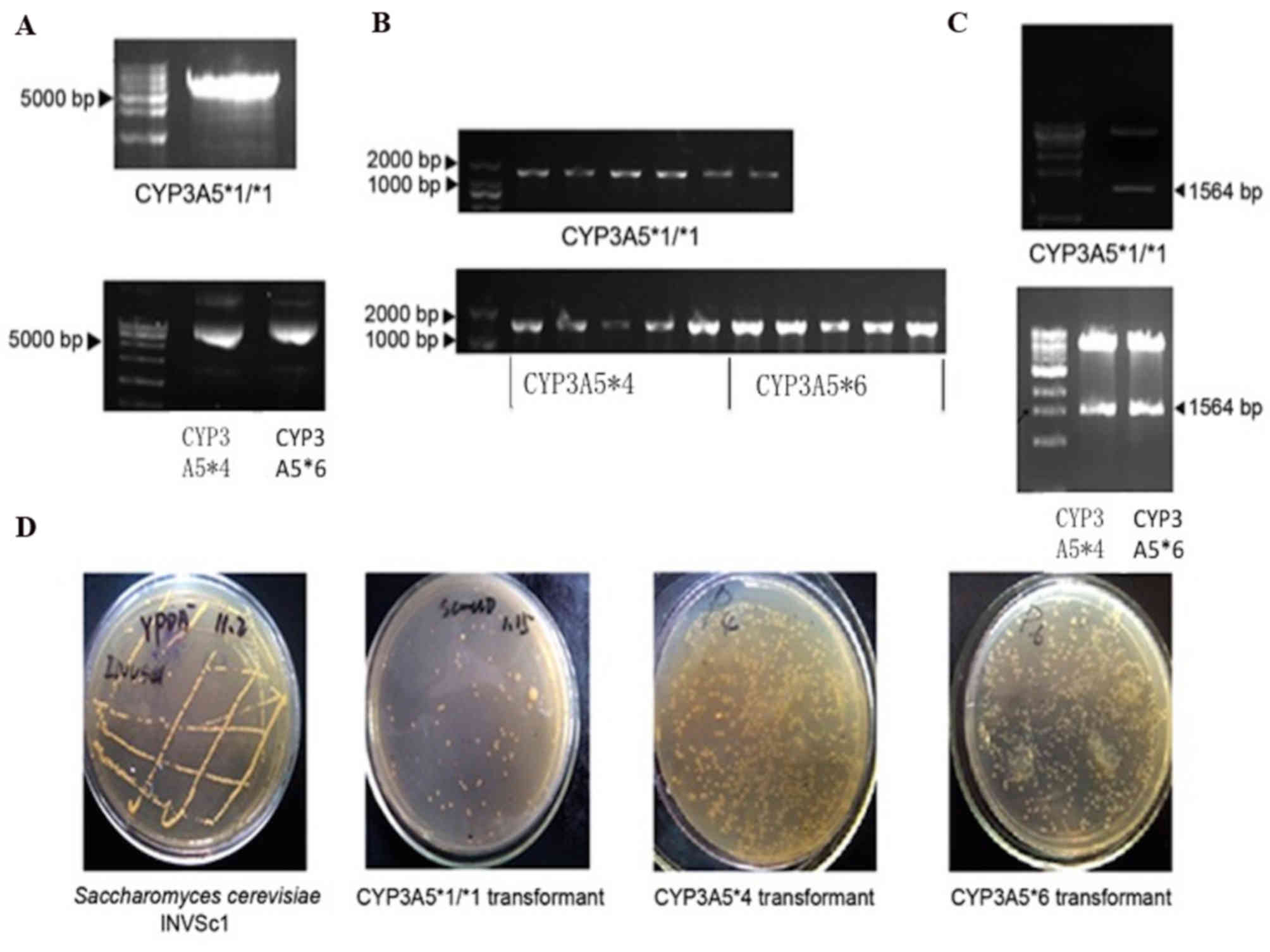
CYP3A5 gene polymorphism expression vectors were
successfully constructed for CYP3A5*1/*1, CYP3A5*4 and CYP3A5*6,
however, this was not achieved for CYP3A*1/*3. The site-specific
mutagenesis failed in the CYP3A5*1/*3 mutation as the CYP3A5*1/*3
mutation occurs in the CYP3A5 gene intron. The introduction of the
nucleotides required to form each CYP3A5 polymorphism was confirmed
with a PCR electrophoretogram (Fig.
2A). Expression vectors constructed by the PCR method were used
as a template for PCR amplification. PCR amplification products
were subjected to agarose gel electrophoresis. In addition, agarose
gel electrophoresis was performed to analyze the constructed
expression vector. The results demonstrated that the expression
vector was successfully constructed (Fig. 2B). Double digestion of the
expression vectors was performed to further confirm that the
expression vectors were correct. The digestion products were
assessed using agarose gel electrophoresis. The results
demonstrated that the expression vectors were successfully
constructed (Fig. 2C).
Additionally, Saccharomyces cerevisiae formed into
monoclonal cells, indicating that the CYP3A5 gene polymorphism
expression vectors were successfully transformed into the INVSC1
yeast cells (Fig. 2D).
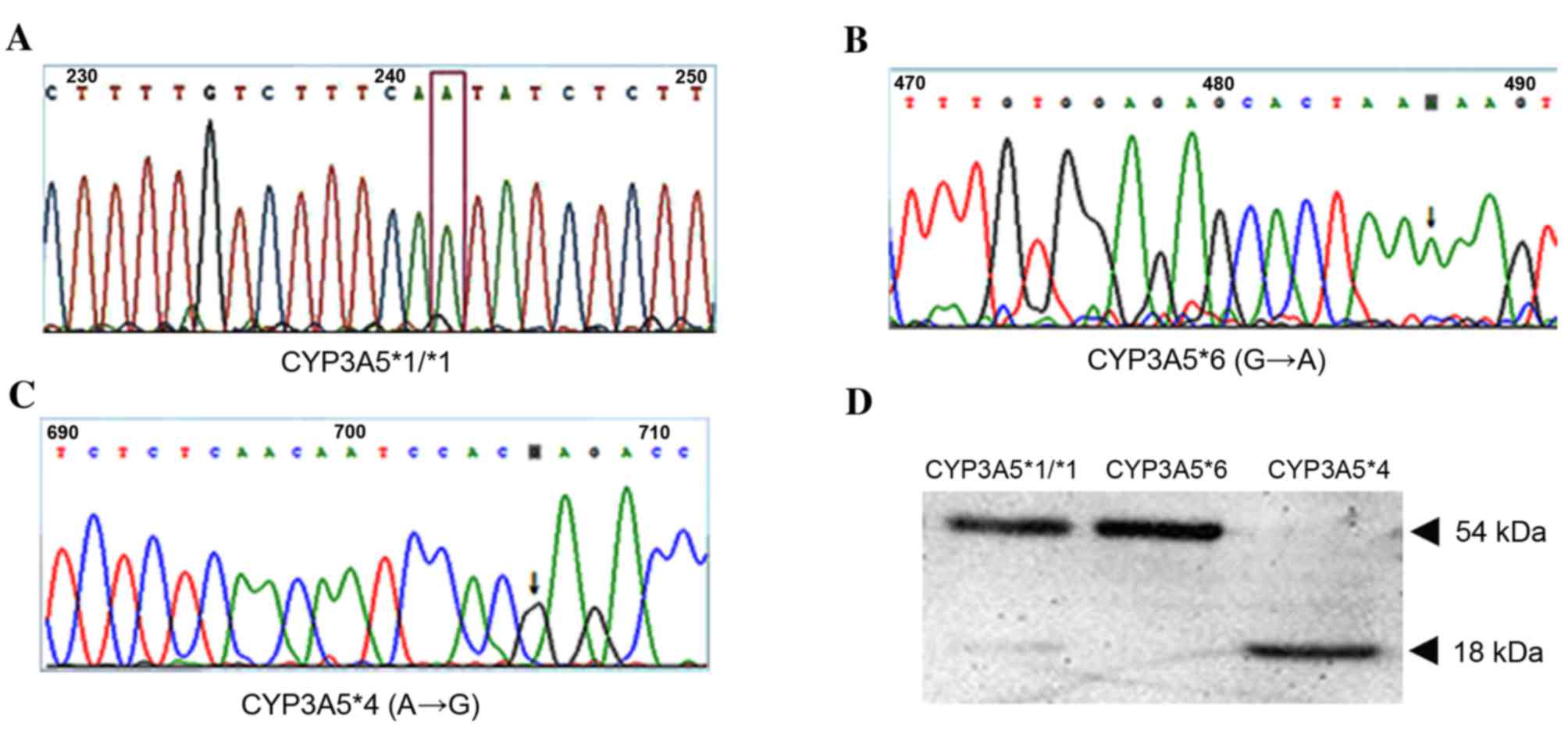
Bidirectional sequencing comparisons were performed using positive
bacterial clones of the double digestion vectors (Fig. 3). CYP3A5*4, the constructed gene
fragment exhibited a base group mutation from A to G at bp 599,
without the presence of other mutations, indicating that the
fragment was CYP3A5*4. CYP3A5*6, the constructed gene fragment
exhibited base group mutation from G to A at bp 624, without the
presence of other mutations, indicating that the fragment was
CYP3A5*6. The results demonstrated that the present study
successfully constructed recombinant yeast expression systems for
three single nucleotide mutants of human cytochrome CYP3A5,
including CYP3A5*1/*1, CYP3A5*4 and CYP3A5*6. No additional
mutations, apart from the desired point mutations created in the
expression vectors, were identified. The gene sequencing results
demonstrated that there were mutations in the required base locus
while other bases were normal (Fig.
3A-C). These results confirmed that the CYP3A5 gene
polymorphism vectors were successfully constructed.
PK prediction model for CYP3A5 gene
polymorphisms
To assess the effects of CYP3A5 gene polymorphisms
on drug metabolism, the present study constructed a PK prediction
model by expressing CYP3A5 gene polymorphism vectors in
Saccharomyces cerevisiae. After the CYP3A5 gene polymorphism
expression vectors were transformed into yeast, the transformants
were selected on Schaeffer's-glucose agar plates. Subsequently,
western blotting was performed to detect protein expression. The
results (Fig. 3D) demonstrated
that CYP3A5*1/*1 and CYP3A5*6 proteins were detected at 54 kDa;
whereas CYP3A5*4 was detected at 18 kDa. These results demonstrate
that the CYP3A5 gene polymorphism sequences were successfully
transformed into the Saccharomyces cerevisiae expression
vectors.
In vitro drug metabolism of amlodipine
by CYP3A5 recombinant enzymes
The standard curve was fitted using standard
concentrations for the X-axis and the area ratio of
standard/internal standard for the Y-axis (Fig. 4A and B). Values for the rate of
metabolite formation using 0.2 mg/ml of the recombinant enzymes
were derived using the standard curve. The Vmax of
amlodipine metabolized by CYP3A5*1/*1, CYP3A5*4 and CYP3A5*6 was
0.61±0.02, 0.42±0.02 and 0.66±0.04 pmol/min/pmol P450,
respectively. The Km of CYP3A5*1/*1, CYP3A5*4 and
CYP3A5*6 was 0.32±0.12 µmol/l, 0.31±0.05 µmol/l and 0.32±0.08
µmol/l, respectively. The drug clearance
(Vmax/Km) for CYP3A5*1/*1, CYP3A5*4 and
CYP3A5*6 was 1.91±0.05, 1.35±0.15 and 2.06±0.05 µl/min/pmol P450,
respectively (Fig. 4C). The
results demonstrated that CYP3A5 recombinant enzymes influenced the
PK of amlodipine. There was a significant difference in the drug
clearance between CYP3A5*4 and CYP3A5*1/*1 (P<0.05). The
difference between CYP3A5*6 and CYP3A5*1/*1 was not statistically
significant (P>0.05).
Discussion
CYP3A5 has an important role in the
metabolism of various drugs. The major polymorphisms of
CYP3A5 are CYP3A5*1, CYP3A5*2,
CYP3A5*3, CYP3A5*4,
CYP3A5*6 and CYP3A5*7 (22,23).
Previous studies have reported the successful construction of an
in vitro Saccharomyces cerevisiae expression systems for
CYP3A4, CYP2D6 and CYP2C9 (22,24).
To the best of our knowledge, the present study is the first to
construct the in vitro CYP3A5 gene polymorphism
Saccharomyces cerevisiae expression systems for
CYP3A5*1/*1, CYP3A5*4 and
CYP3A5*6, and to compare the in vitro
metabolic rate with in vivo rates.
Amlodipine is a CYP3A5 substrate that is
widely used for the treatment of hypertension. Numerous studies
have demonstrated that the CYP3A5 gene polymorphisms affect
the PK and PD of amlodipine (24–27).
The in vitro drug metabolism results of amlodipine by
CYP3A5 recombinase in the present study demonstrated that
there were significant differences between CYP3A5*4
and CYP3A5*1/*1
(CYP3A5*1/*1> CYP3A5*6 >
CYP3A5*4), while the in vivo PK results for
amlodipine demonstrated that CYP3A5*6 exhibited the
highest metabolic ratio of amlodipine, followed by
CYP3A5*1/*1 and CYP3A5*4. There
was a significant difference between the in vitro and in
vivo PK results obtained for CYP3A5*1/*1
and CYP3A5*4 (P<0.05) however no significant
difference between CYP3A5*1/*1 and
CYP3A5*6.
The reaction ratio of in vivo CYP3A5*6
was much larger compared with CYP3A5*1/*1 and
CYP3A5*4 polymorphisms, which is in contrast to the
in vitro drug metabolism results. This may be due to the
fact that there were 2 patients (3.51%) with the
CYP3A5*6 allele, which is higher than the normal
CYP3A5*6 allele frequency. The CYP3A5*1
allele frequency is 25% in Chinese populations, and the
CYP3A5*3 allele frequency is >25%. However, the
CYP3A5*6 allele is not as common in Asian
populations, including Chinese and Japanese populations (5,28–30).
Further studies that focus on the mechanisms and frequency of the
CYP3A5*6 allele are required. Large clinical trials
should be conducted to clarify the difference between the PK
effects of these three gene polymorphisms. The antihypertensive
effects of amlodipine among different patients were determined.
CYP3A5*1/*3 exhibited the largest
antihypertensive effect, followed by CYP3A5*4,
CYP3A5*1/*1 and CYP3A5*6. The in
vivo PK data demonstrated that CYP3A5*1/*1
had the largest reaction ratio. Thus, the antihypertensive results
are consistent with the PK results. The current study had certain
limitations that must be stated. The current study failed to
construct the in vitro CYP3A5*1*3 polymorphism
Saccharomyces cerevisiae expression vector as the
CYP3A5*1/*3 mutation occurs in the
CYP3A5 gene intron and it was difficult to mutate the
intron. In future studies, we hope to identify another method to
solve this problem. Additionally, only 2 patients were included in
CYP3A5*4 and CYP3A5*6 groups, and the
small patient population and the retrospective nature of the study
do not allow any conclusions to be drawn about the effectiveness of
this prediction method. Studies consisting of larger groups of
patients with long-term follow-up are required to confirm these
results.
In conclusion, the present study indicates that the
CYP3A5 gene polymorphism Saccharomyces cerevisiae
expression system is a potential tool for the prediction of drug
metabolism, particularly for novel drugs prior to their application
in clinical trials. This expression system may also aid in the
development of individualized clinical medications.
Acknowledgements
This study was supported by grants from the
National Science and Technology Major Projects for ‘Major New Drugs
Innovation and Development’ (grant no. 2012ZX09303014001), the
National Natural Science Foundation of China (grant no. 81273594)
and Fundamental Research Funds for the Central Universities of
Central South University (grant no. 2016zzts153).
References
|
1
|
Emoto C and Iwasaki K: Approach to predict
the contribution of cytochrome P450 enzymes to drug metabolism in
the early drug-discovery stage: The effect of the expression of
cytochrome b(5) with recombinant P450 enzymes. Xenobiotica.
37:986–999. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang YP, Zuo XC, Huang ZJ, Cai JJ, Wen J,
Duan DD and Yuan H: CYP3A5 polymorphism, amlodipine and
hypertension. J Hum Hypertens. 28:145–149. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin YS, Dowling AL, Quigley SD, Farin FM,
Zhang J, Lamba J, Schuetz EG and Thummel KE: Co-regulation of
CYP3A4 and CYP3A5 and contribution to hepatic and intestinal
midazolam metabolism. Mol Pharmacol. 62:162–172. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shin J, Pauly DF, Pacanowski MA, Langaee
T, Frye RF and Johnson JA: Effect of cytochrome P450 3A5 genotype
on atorvastatin pharmacokinetics and its interaction with
clarithromycin. Pharmacotherapy. 31:942–950. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kuehl P, Zhang J, Lin Y, Lamba J, Assem M,
Schuetz J, Watkins PB, Daly A, Wrighton SA, Hall SD, et al:
Sequence diversity in CYP3A promoters and characterization of the
genetic basis of polymorphic CYP3A5 expression. Nat Genet.
27:383–391. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Williams JA, Ring BJ, Cantrell VE, Jones
DR, Eckstein J, Ruterbories K, Hamman MA, Hall SD and Wrighton SA:
Comparative metabolic capabilities of CYP3A4, CYP3A5, and CYP3A7.
Drug Metab Dispos. 30:883–891. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Walker DK, Humphrey MJ and Smith DA:
Importance of metabolic stability and hepatic distribution to the
pharmacokinetic profile of amlodipine. Xenobiotica. 24:243–250.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sugimoto K, Uno T and Tateishi T: Effects
of the CYP3A5 genotype on omeprazole sulfoxidation in CYP2C19 PMs.
Eur J Clin Pharmacol. 64:583–587. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim KA, Park PW, Lee OJ, Choi SH, Min BH,
Shin KH, Chun BG, Shin JG and Park JY: Effect of CYP3A5*3 genotype
on the pharmacokinetics and pharmacodynamics of amlodipine in
healthy Korean subjects. Clin Pharmacol Ther. 80:646–656. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Elmachad M, Elkabbaj D, Elkerch F, Laarabi
FZ, Barkat A, Oualim Z and Sefiani A: Frequencies of CYP3A5*1/*3
variants in a Moroccan population and effect on tacrolimus daily
dose requirements in renal transplant patients. Genet Test Mol
Biomarkers. 16:644–647. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Provenzani A, Notarbartolo M, Labbozzetta
M, Poma P, Vizzini G, Salis P, Caccamo C, Bertani T, Palazzo U,
Polidori P, et al: Influence of CYP3A5 and ABCB1 gene polymorphisms
and other factors on tacrolimus dosing in Caucasian liver and
kidney transplant patients. Int J Mol Med. 28:1093–1102.
2011.PubMed/NCBI
|
|
12
|
Amundsen R, Åsberg A, Ohm IK and
Christensen H: Cyclosporine A- and tacrolimus-mediated inhibition
of CYP3A4 and CYP3A5 in vitro. Drug Metab Dispos. 40:655–661. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guo C, Pei QI, Tan H, Huang Z, Yuan H and
Yang G: Effects of genetic factors on the pharmacokinetics and
pharmacodynamics of amlodipine in primary hypertensive patients.
Biomed Rep. 3:195–200. 2015.PubMed/NCBI
|
|
14
|
Choi M, Eren-Dogu ZF, Colangelo C,
Cottrell J, Hoopmann MR, Kapp EA, Kim S, Lam H, Neubert TA,
Palmblad M, et al: ABRF proteome informatics research group (iPRG)
2015 study: Detection of differentially abundant proteins in
label-free quantitative LC-MS/MS experiments. J Proteome Res Jan.
3:2017.(Epub ahead of print).
|
|
15
|
Beresford AP, Macrae PV, Alker D and
Kobylecki RJ: Biotransformation of amlodipine. Identification and
synthesis of metabolites found in rat, dog and human
urine/confirmation of structures by gas chromatography-mass
spectrometry and liquid chromatography-mass spectrometry.
Arzneimittelforschung. 39:201–209. 1989.PubMed/NCBI
|
|
16
|
Chennuru LN, Choppari T, Nandula RP, Zhang
T and Franco P: Direct separation of pregabalin enantiomers using a
zwitterionic chiral selector by high performance liquid
chromatography coupled to mass spectrometry and ultraviolet
detection. Molecules. 21:E15782016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Colson CA, Cornelis PE, Digneffe CS and
Walon C: Genetically engineered microorganisms for massive
production of amylolytic enzymes and process for preparing same. US
Patend 4469791 A. Filed February 13, 1981; issued September 4.
1984.
|
|
18
|
Narita V, Widyanto RM, Pambudi S and
Sudiro TM: Cloning of dengue virus type 3 (Indonesian Strain
D3-1703) non structural-1 gene into pYES2/CT vector. Makara Sains.
15:173–178. 2011.
|
|
19
|
Costa IM, Schultz L, de Araujo Bianchi
Pedra B, Leite MS, Farsky SH, De Oliveira MA, Pessoa A and Monteiro
G: Recombinant L-asparaginase 1 from Saccharomyces cerevisiae: An
allosteric enzyme with antineoplastic activity. Sci Rep.
6:362392016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huang Y, Liu XL, Wen J, Huang LH, Lu Y,
Miao RJ, Liu X, Li Y, Xing XW and Yuan H: Downregulation of the β1
adrenergic receptor in the myocardium results in insensitivity to
metoprolol and reduces blood pressure in spontaneously hypertensive
rats. Mol Med Rep. 15:703–711. 2017.PubMed/NCBI
|
|
21
|
Guengerich FP, Brian WR, Iwasaki M, Sari
MA, Bäärnhielm C and Berntsson P: Oxidation of dihydropyridine
calcium channel blockers and analogues by human liver cytochrome
P-450 IIIA4. J Med Chem. 34:1838–1844. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ha HR, Chen J, Freiburghaus AU and Follath
F: Metabolism of theophylline by cDNA-expressed human cytochromes
P-450. Br J Clin Pharmacol. 39:321–326. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ho H, Pinto A, Hall SD, Flockhart DA, Li
L, Skaar TC, Cadman P, O'Connor DT, Wagner U, Fineberg NS and
Weinberger MH: Association between the CYP3A5 genotype and blood
pressure. Hypertension. 45:294–298. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Granvil CP, Yu AM, Elizondo G, Akiyama TE,
Cheung C, Feigenbaum L, Krausz KW and Gonzalez FJ: Expression of
the human CYP3A4 gene in the small intestine of transgenic mice: In
vitro metabolism and pharmacokinetics of midazolam. Drug Metab
Dispos. 31:548–558. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang L, Miyaki K, Wang W and Muramatsu M:
CYP3A5 polymorphism and sensitivity of blood pressure to dietary
salt in Japanese men. J Hum Hypertens. 24:345–350. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Stopher DA, Beresford AP, Macrae PV and
Humphrey MJ: The metabolism and pharmacokinetics of amlodipine in
humans and animals. J Cardiovasc Pharmacol. 12 Suppl 7:S55–S59.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Faulkner JK, McGibney D, Chasseaud LF,
Perry JL and Taylor IW: The pharmacokinetics of amlodipine in
healthy volunteers after single intravenous and oral doses and
after 14 repeated oral doses given once daily. Br J Clin Pharmacol.
22:21–25. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Balram C, Zhou Q, Cheung YB and Lee EJ:
CYP3A5*3 and *6 single nucleotide polymorphisms in three distinct
Asian populations. Eur J Clin Pharmacol. 59:123–126. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fukuen S, Fukuda T, Maune H, Ikenaga Y,
Yamamoto I, Inaba T and Azuma J: Novel detection assay by PCR-RFLP
and frequency of the CYP3A5 SNPs, CYP3A5*3 and *6, in a Japanese
population. Pharmacogenetics. 12:331–334. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Thompson EE, Kuttab-Boulos H, Witonsky D,
Yang L, Roe BA and Di Rienzo A: CYP3A variation and the evolution
of salt-sensitivity variants. Am J Hum Genet. 75:1059–1069. 2004.
View Article : Google Scholar : PubMed/NCBI
|