Introduction
Ischemic stroke occurs due to the temporary or
permanent blockage of blood circulation in the brain in several
circumstances, including brain ischemia, cardiac arrest and
cardiovascular surgery (1–3). Transient cerebral ischemia, which is
a major cause of ischemic stroke, leads to selective neuronal
damage/death in vulnerable brain areas, including the cerebral
cortex, the striatum and the hippocampus (3,4). In
particular, the most vulnerable area to transient cerebral ischemia
is the CA1 region of the hippocampus, in which pyramidal neurons of
the stratum pyramidale of the CA1 region die several days following
ischemia-reperfusion injury (5,6).
One of mechanisms regarding neuronal damage/death
induced by cerebral ischemia is oxidative stress, which is
associated with the excessive production of reactive oxygen species
(ROS) (7,8). The accumulation of ROS in ischemic
conditions induces DNA damage, lipid peroxidation and changes in
cellular proteins (8,9). ROS is converted into nontoxic
compounds by enzymatic antioxidants including superoxide dismutases
(SODs), catalase (CAT) and glutathione peroxidase (GPX); known
antioxidant enzymes (8,10). Various antioxidants, including
antioxidant enzymes, have been recognized as beneficial in
therapies for neurologic diseases (11,12).
Many studies on neuroprotection by plant extracts
have been reported using animal models of cerebral ischemic insults
(13,14). Chrysanthemum indicum Linné
(Compositae; CIL) is a traditional herb used for medicines in East
Asia. It has been used for the treatment of immune-related
disorders, hypertension, infectious diseases and respiratory
illness (15,16). The major components of CIL are
bornyl acetate (10.00–21.33%), borneol (8.34–18.34%), camphor
(7.75–23.52%) and germacrene D (1.08–12.67%). Significant minor
components of CIL include α-terpineol (1.28–3.32%), terpinen-4-ol
(0.70–1.59%) and caryophyllene oxide (0.13–2.73%). However,
1,8-cineole is present at 30.41% in fresh flower oil and only
0.12–0.61% in the oil of air-dried and processed flowers (17,18).
CIL exhibits anti-bacterial, anti-viral, antioxidant,
anti-inflammatory and immunomodulatory functions (17,19).
To the best of the authors' knowledge, few studies regarding
neuroprotective effects of CIL and its antioxidant mechanism in
brain ischemic insults have been published; therefore, the present
study investigated the neuroprotective effect of CIL and whether
endogenous antioxidant enzymes, including SOD1, SOD2, CAT and GPX,
were associated with the neuroprotection in the hippocampus of the
gerbil, a good animal model of transient cerebral ischemia
(20,21).
Materials and methods
Preparation of extract from CIL
CIL was collected by Professor Jong Dai Kim in
Kangwon Province (South Korea), in October 2013 and maintained in a
deep freezer (−70°C). The CIL was extracted with 70% ethanol at
70°C for 4 h, and extraction was repeated three times. The extract
was filtered through Whatman filter paper (no. 2), concentrated
with a vacuum evaporator, and completely dried with a freeze-drier.
The extraction yield was 14.5%.
Groups of experimental animals
Male Mongolian gerbils (Meriones
unguiculatus; weight, 65–75 g; age, 6 months) were obtained
from the Experimental Animal Center, Kangwon National University,
Chuncheon, South Korea. The animals were housed in standard
conditions under adequate temperature (23°C) and humidity (60%)
control with a 12-h:12-h light:dark cycle, and were provided with
free access to food and water. All the experimental protocols were
approved (approval no. KW-130424-1) by the Institutional Animal
Care and Use Committee at Kangwon University and adhered to
guidelines that are in compliance with the current international
laws and policies (Guide for the Care and Use of Laboratory
Animals, The National Academies Press, 8th edition, 2011).
The experimental animals were divided into four
groups (n=7 at each time point in each groups): i)
Vehicle-sham-group, which was treated with vehicle and underwent no
ischemia; ii) CIL-sham-group, which was treated with CIL and
underwent no ischemia; iii) vehicle-ischemia-group, which was
treated with vehicle and underwent ischemia surgery; and iv)
CIL-ischemia-group, which was treated with CIL and underwent
ischemia surgery.
Administration with CIL
CIL extract was dissolved in saline and
administrated orally using a feeding needle once a day for seven
days prior to ischemia surgery, according to previously published
procedure (22). The preliminary
tests were carried out with 25, 50, 100 and 200 mg/kg CIL. There
were no neuroprotective effects in doses of 25, 50 and 100 mg/kg,
but protective effects were demonstrated in animals treated with
200 mg/kg. Thus, 200 mg/kg was selected. The last treatment was at
30 min prior to the surgery to maintain the level of CIL in animals
prior to and following surgery.
Induction of transient cerebral
ischemia
As previously described (23), the experimental animals were
anesthetized with a mixture of ~2.5% isoflurane (Baxtor Healthcare
Corp., Deerfield, IL, USA) in 33% O2 and 67%
NO2. Bilateral common carotid arteries were isolated and
occluded for 5 min using non-traumatic aneurysm clips. Rectal
temperature was controlled under normothermic (37±0.5°C) conditions
during the surgery with a rectal temperature probe (TR-100; Fine
Science Tools, Inc., Foster City, CA, USA).
Tissue preparation for histology
As previously described (24), gerbils (n=7 at each time point in
each group) were anesthetized with pentobarbital sodium at the
designated times and perfused transcardially with 0.1 M
phosphate-buffered saline (pH 7.4) followed by 4% paraformaldehyde
in 0.1 M phosphate-buffer (PB; pH 7.4). The brains were removed and
postfixed in the same fixative for 6 h, and the brain tissues were
sectioned on a cryostat (Leica Microsystems GmbH, Wetzlar, Germany)
into 30 µm coronal sections.
Immunohistochemistry
Immunohistochemistry was performed according to the
previously published procedure (24). In short, the sections were
incubated with diluted mouse anti-neuronal nuclei (NeuN; 1:800;
cat. no. MAB377), sheep anti-copper, zinc-superoxide dismutase
(SOD1; 1:1,000; cat. no. 574597) and sheep anti-mangan-superoxide
dismutase (SOD2; 1:1,000; cat. no. 574596; all from EMD Millipore,
Billerica, MA, USA), rabbit anti-catalase (CAT; 1:500; cat. no.
ab52477) and sheep anti-glutathione peroxidase (GPX; 1:1,000; cat.
no. ab21966; both from Abcam, Cambridge, MA, UK). Thereafter the
tissues were exposed to biotinylated horse anti-mouse (1:250; cat.
no. BA-2000), rabbit anti-sheep (1:250; cat. no. BA-6000) and goat
anti-rabbit immunoglobulin (Ig)G (1:250; cat. no. BA-1000) and
streptavidin peroxidase complex (1:200, all from Vector
Laboratories, Burlingame, CA, USA) and were visualized with
3,3′-diaminobenzidine (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany).
Fluoro-Jade B histofluorescence
staining
Fluoro-Jade B (F-J B) histofluorescence staining was
conducted according to a published procedure (25) in order to examine neuronal death.
In brief, the sections were immersed in a solution containing 1%
sodium hydroxide, transferred to a solution of 0.06% potassium
permanganate and transferred to a 0.0004% F-J B (Histo-Chem Inc.,
Jefferson, AR, USA) staining solution. The stained sections were
observed using an epifluorescent microscope (Zeiss AG, Oberkochen,
Germany) with a blue (450–490 nm) excitation source and a barrier
filter.
Data analysis
Data were analyzed according to published procedure
(26). Briefly, to quantitatively
analyze immunoreactivities of antioxidant enzymes, the
immunoreactivity of SOD1, SOD2, CAT and GPX-immunoreactive
structures was evaluated on the basis of optical density (OD),
which was obtained following the transformation of the mean gray
level using the formula: OD = log (256/mean gray level). A portion
of the OD of an image file was calibrated in Adobe Photoshop 8.0
(Adobe Systems, Inc., San Jose, CA, USA) and then analyzed as a
percentage, with the sham-operated-group designated as 100%, in
ImageJ version 1.59 (National Institutes of Health, Bethesda, MD,
USA). For cell counting, NeuN- and F-J B-positive cells were imaged
from the stratum pyramidale through an AxioM1 light microscope
(Zeiss AG) equipped with a digital camera (Axiocam; Zeiss AG)
connected to a PC monitor. The mean number of NeuN- and F-J
B-positive cells was counted in a 200×200 µm square applied
approximately at the center of the CA1 region. Cell counts were
obtained by averaging the total cell numbers from each animal per
group and analyzing them as a percentage, with the
vehicle-sham-group designated as 100%.
Statistical analysis
The data was presented as mean ± standard error of
mean of the means among the groups and were statistically analyzed
by analysis of variance with a post hoc Bonferroni's multiple
comparisons test, in order to present differences among
experimental groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Antioxidant immunoreactivities
SOD1 immunoreactivity
SOD1 immunoreactivity was easily detected in
pyramidal cells of the hippocampal CA1 region of the
vehicle-sham-group (Fig. 1A). In
the vehicle-ischemia-group, SOD1 immunoreactivity was significantly
decreased in the CA1 pyramidal cells 2 days following
ischemia-reperfusion and SOD1 immunoreactivity was hardly detected
in the CA1 pyramidal cells 5 days following ischemia-reperfusion
(Fig. 1C, E and G).
 | Figure 1.SOD1 immunohistochemistry in the
hippocampal CA1 region of the (A) vehicle-sham-, (B) CIL-sham-, (C
and E) vehicle-ischemia- and (D and F) CIL-ischemia-groups
following ischemia-reperfusion. SOD1 immunoreactivity is easily
observed in the SP in the vehicle-sham-group. In the
vehicle-ischemia-group, SOD1 immunoreactivity is hardly observed in
the SP (asterisk) 5 days following ischemia-reperfusion. In the
CIL-sham-group, SOD1 immunoreactivity is significantly increased
compared with the vehicle-sham-group, and the immunoreactivity is
well detected until 5 days following ischemia-reperfusion. Scale
bar, 100 µm. (G) ROD as % values of SOD1 immunoreactivity in the SP
of each group (*P<0.05 vs. vehicle-sham-group;
#P<0.05 vs. corresponding vehicle-ischemia-group;
†P<0.05 vs. respective pre-time point group). The
bars indicate the means ± standard error of mean. SP, stratum
pyramidale; SOD, superoxide dismutase; CIL, Chrysanthemum
indicum Linné extract; ROD, relative optical density; SO,
stratum oriens; SR, stratum radiatum. |
In the CIL-sham-group, SOD1 immunoreactivity in the
CA1 pyramidal cells was significantly higher compared with the
vehicle-sham-group (Fig. 1B and
G). In the CIL-ischemia-group, SOD1 immunoreactivity in the CA1
pyramidal cells was reduced following ischemia-reperfusion;
however, the SOD1 immunoreactivity was significantly higher
compared with the corresponding vehicle-ischemia-group (Fig. 1D, F and G). In particular, 5 days
following ischemia-reperfusion in the CIL-ischemia-group, numerous
SOD1-immunoreactive CA1 pyramidal cells were observed (Fig. 1F).
SOD2 immunoreactivity
SOD2 immunoreactivity was also clearly identified in
the CA1 pyramidal cells in the vehicle-sham-group (Fig. 2A). SOD1 immunoreactivity in the CA1
pyramidal cells was significantly decreased 2 days and barely
detected 5 days following ischemia-reperfusion in the
vehicle-ischemia-group (Fig. 2C, E and
G).
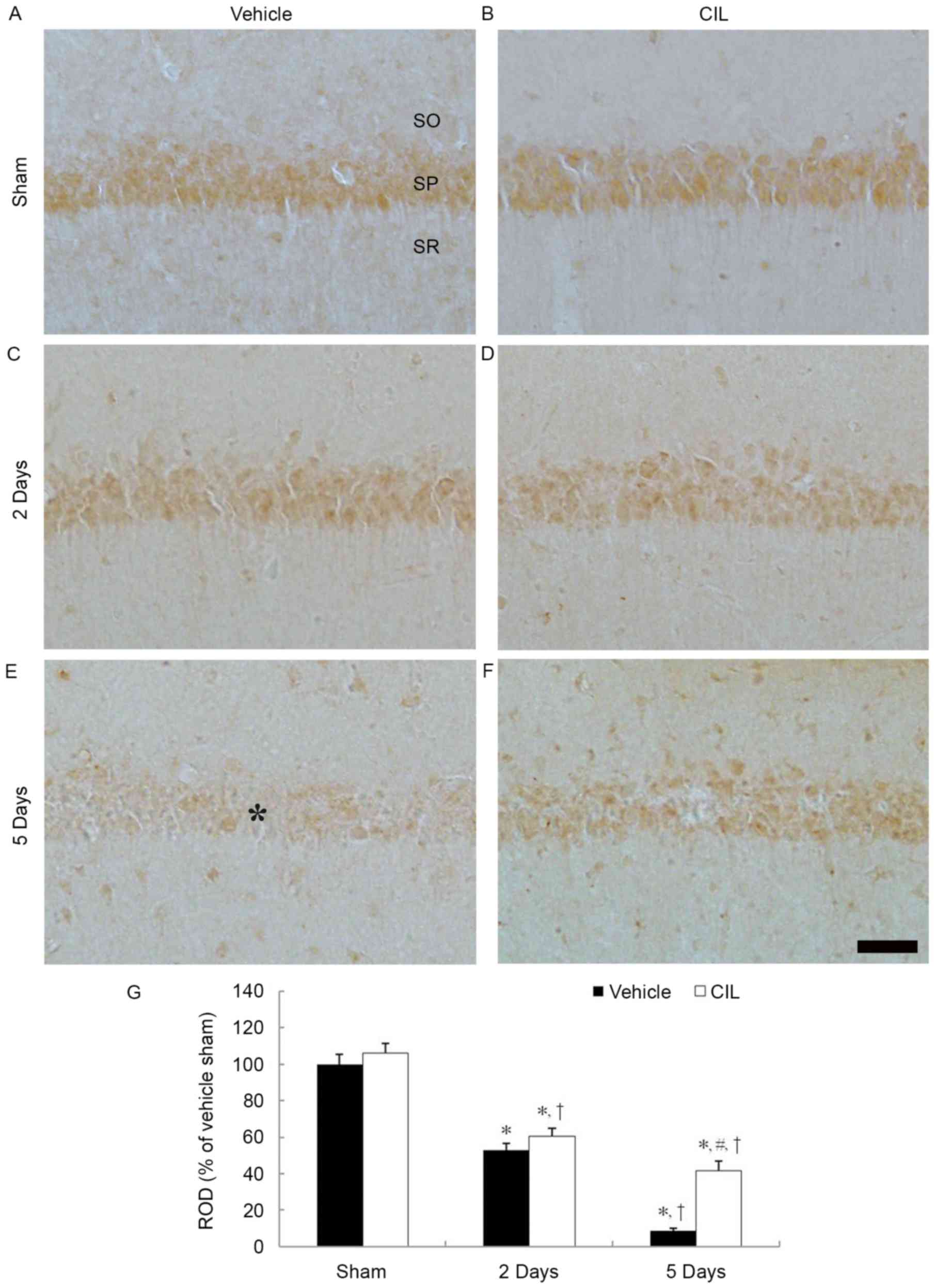 | Figure 2.SOD2 immunohistochemistry in the
hippocampal CA1 region of the (A) vehicle-sham-, (B) CIL-sham-, (C
and E) vehicle-ischemia- and (D and F) CIL-ischemia-groups
following ischemia-reperfusion. SOD2 immunoreactivity is identified
in the SP in the vehicle-sham-group, and SOD2 immunoreactivity in
the SP (asterisk) is barely observable 5 days following
ischemia-reperfusion. In the CIL-sham-group, SOD2 immunoreactivity
is similar to that of the vehicle-sham-group, and, in the
CIL-ischemia-group, SOD2 immunoreactivity in the SP is higher than
that corresponding vehicle-sham-group. Scale bar, 100 µm. (G) ROD
as % values of SOD2 immunoreactivity in the SP of each group
(*P<0.05 vs. vehicle-sham-group; #P<0.05 vs.
corresponding vehicle-ischemia-group; †P<0.05 vs.
respective pre-time point group). The bars indicate the means ±
standard error of mean. SP, stratum pyramidale; SOD, superoxide
dismutase; CIL, Chrysanthemum indicum Linné extract; ROD,
relative optical density; SO, stratum oriens; SR, stratum
radiatum. |
In the CIL-sham-group, SOD2 immunoreactivity in the
CA1 pyramidal cells was slightly increased compared with the
vehicle-sham-group; however, it was not statistically significant
(Fig. 2B and G). In the
CIL-ischemia-group, the changing pattern of SOD2 immunoreactivity
in the CA1 pyramidal cells was similar to that of the SOD1
immunoreactivity (Fig. 2D, F and
G).
CAT immunoreactivity
In the vehicle-sham-group, CAT immunoreactivity was
clearly observed in the pyramidal cells of the stratum pyramidale
layer of the CA1 region (Fig. 3A).
In the vehicle-ischemia-group, CAT immunoreactivity was
significantly decreased in the CA1 pyramidal cells at 2 days
post-ischemia (Fig. 3C and G). At
5 days post-ischemia, CAT immunoreactivity in the CA1 pyramidal
cells was barely observable; however, CAT immunoreactivity was
newly expressed in non-pyramidal cells in the strata oriens and
radiatum of the CA1 region (Fig. 3E
and G).
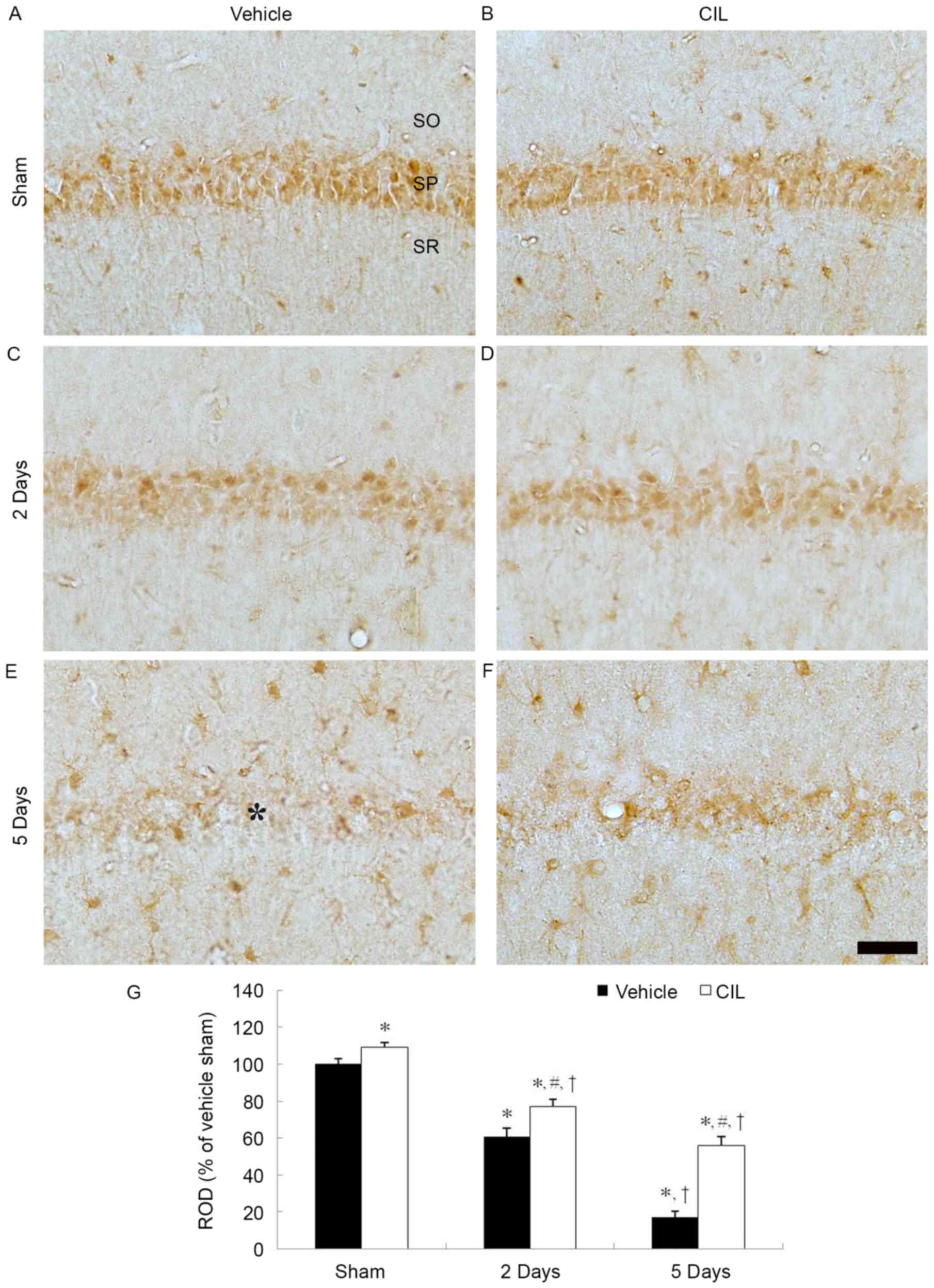 | Figure 3.CAT immunohistochemistry in the
hippocampal CA1 region of the (A) vehicle-sham, (B) CIL-sham-, (C
and E) vehicle-ischemia- and (D and F) CIL-ischemia-groups
following ischemia-reperfusion. CAT immunoreactivity is markedly
decreased in the SP (asterisk) of the vehicle-ischemia-group at 5
days following ischemia-reperfusion; however, in the CIL-sham- and
CIL-ischemia-groups, CAT immunoreactivity in the SP is
significantly higher compared with the vehicle-sham- and
vehicle-ischemia-groups. Scale bar, 100 µm. (G) ROD as % values of
CAT immunoreactivity in the SP of each group (*P<0.05 vs.
vehicle-sham-group; #P<0.05 vs. corresponding
vehicle-ischemia-group; †P<0.05 vs. respective
pre-time point group). The bars indicate the means ± standard error
of mean. SP, stratum pyramidale; CAT, catalase; CIL,
Chrysanthemum indicum Linné extract; ROD, relative optical
density; SO, stratum oriens; SR, stratum radiatum. |
In the CIL-sham-group, CAT immunoreactivity in the
CA pyramidal cells was significantly increased compared with the
vehicle-sham-group (Fig. 3B and
G). In the CIL-ischemia-group, although the CAT
immunoreactivity in the CA1 pyramidal cells was decreased following
ischemia-reperfusion, the CAT immunoreactivity was significantly
higher compared with the corresponding vehicle-ischemia-group
(Fig. 3D, F and G).
GPX immunoreactivity
GPX immunoreactivity in the vehicle-sham-group was
easily detected in the CA1 pyramidal cells (Fig. 4A). GPX immunoreactivity in the CA1
pyramidal cells was markedly decreased at 2 days post-ischemia and
barely identified at 5 days post-ischemia (Fig. 4C, E and G).
 | Figure 4.GPX immunohistochemistry in the
hippocampal CA1 region of the (A) vehicle-sham-, (B) CIL-sham-, (C
and E) vehicle-ischemia- and (D and F) CIL-ischemia-groups
following ischemia-reperfusion. GPX immunoreactivity is detected
well in the SP of the vehicle-sham-group; the immunoreactivity is
significantly increased in the CIL-sham-group. In the
vehicle-ischemia-group, GPX immunoreactivity is hardly observed in
the SP at 5 days post-ischemia; however, in the CIL-ischemia-group,
GPX immunoreactivity is significantly higher than that in the
vehicle-ischemia-group. Scale bar, 100 µm. (G) ROD as % values of
GPX immunoreactivity in the SP of each group (*P<0.05 vs.
vehicle-sham-group; #P<0.05 vs. corresponding
vehicle-ischemia-group; †P<0.05 vs. respective
pre-time point group). The bars indicate the means ± standard error
of mean. GPX, glutathione peroxidase; CIL, Chrysanthemum
indicum Linné extract; ROD, relative optical density; SO,
stratum oriens; SP, stratum pyramidale; SR, stratum radiatum. |
GPX immunoreactivity in the CA1 pyramidal cells of
the CIL-sham-group was significantly higher compared with the
vehicle-sham-group (Fig. 4B and
G). In the CIL-ischemia-group, GPX immunoreactivity in the CA1
pyramidal cells was decreased following ischemia-reperfusion;
however, the GPX immunoreactivity was significantly higher compared
with the corresponding vehicle-ischemia-group (Fig. 4D, F and G).
Neuroprotective effects
NeuN-positive neurons
NeuN-positive neurons were identified in the stratum
pyramidale of the hippocampus proper (CA1-3 regions) of the
vehicle-sham-group (Fig. 5A and
B). The distribution of NeuN-positive neurons in the
CIL-sham-group was similar to the vehicle-sham-group and the number
of NeuN-positive neurons remained unchanged (Fig. 5D, E and M).
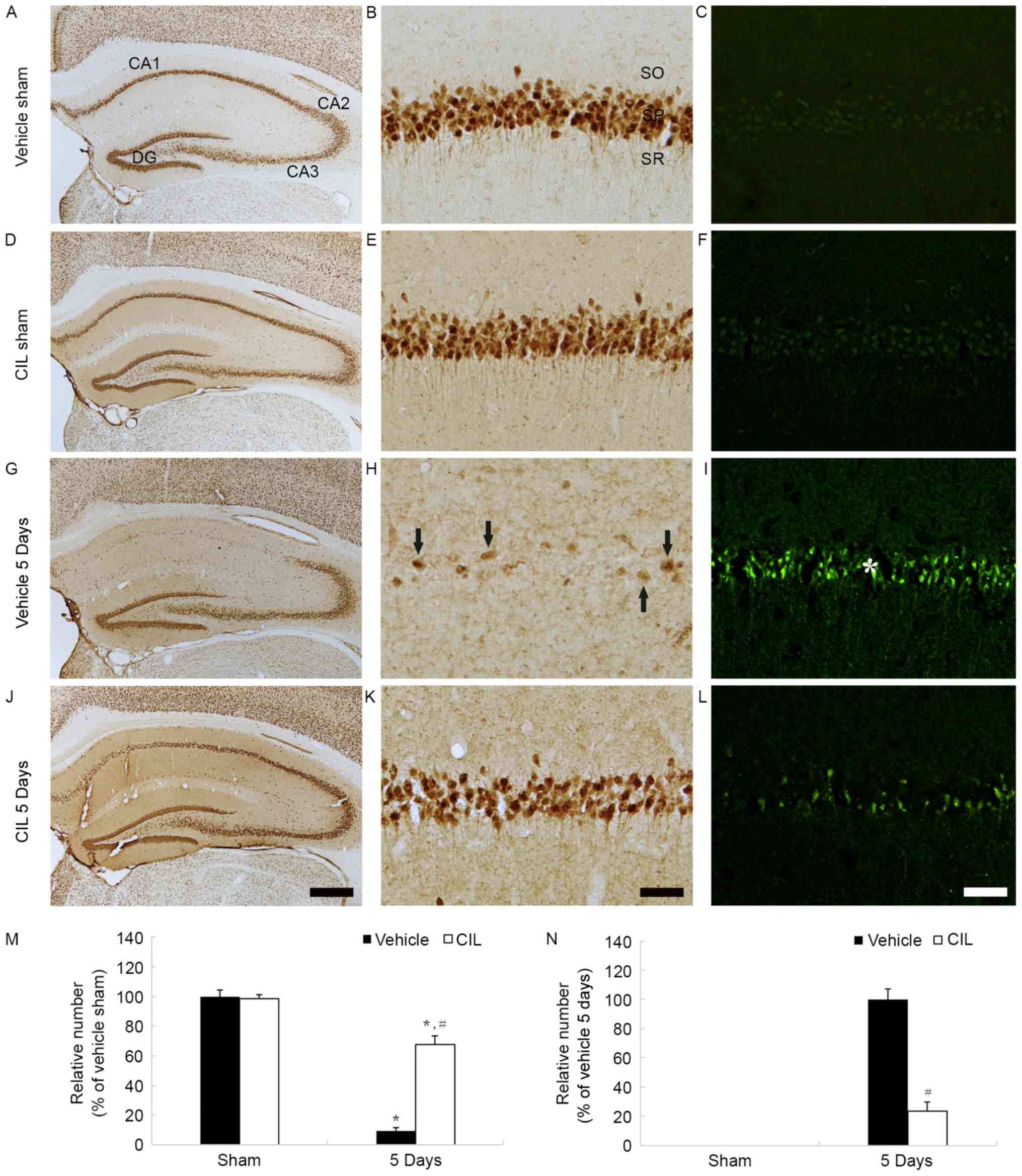 | Figure 5.NeuN-(left and middle columns) and
F-J B-(right column) positive cells of (A-C) vehicle-sham-, (D-F)
CIL-sham-, (G-I) vehicle-ischemia-(J-L) and CIL-ischemia-groups 5
days following ischemia-reperfusion. In the vehicle-ischemia-group,
a few NeuN-(arrows) and numerous F-J B-(asterisk) are detected in
the SP of the CA1 region. In the CIL-ischemia-group, numerous NeuN-
and few F-J B-positive cells are observed in the SP at 5 days
post-ischemia. Scale bar, (A, D, G and J) 50 µm and (B, C, E, F, H,
I, K and L) 100 µm. (M and N) Relative analysis as % of the number
of NeuN- and F-J B-positive cells in the CA1 region (*P<0.05 vs.
respective vehicle-sham-group; #P<0.05 vs.
corresponding vehicle-ischemia-group). The bars indicate the means
± standard error of mean. SP, stratum pyramidale; NeuN, neuronal
nuclei; CIL, Chrysanthemum indicum Linné extract; F-J B,
Fluoro-Jade B; SO, stratum oriens; SR, stratum radiatum. |
In the vehicle-ischemia-group, a small number of
NeuN-positive neurons were observed in the stratum pyramidale of
the CA1 region, and none in the CA2-3 region, 5 days following
ischemia-reperfusion (Fig. 5G and
H); the relative number of NeuN-positive neurons was ~9%
compared with the vehicle-sham-group (Fig. 5M). However, in the
CIL-ischemia-group, numerous NeuN-positive neurons were detected in
the stratum pyramidale of the CA1 region 5 days following
ischemia-reperfusion (Fig. 5J and
K); the protected neurons were ~68% of the vehicle-sham-group
(Fig. 5M); this finding was
identical to our previously study (22).
F-J B-positive cells
In the vehicle-sham- and CIL-sham-groups, F-J
B-positive cells were not observed in the stratum pyramidale of the
CA1 region (Fig. 5C, F and N). In
the vehicle-ischemia-group, however, numerous F-J B-positive cells
were detected in the stratum pyramidale of the CA1 region 5 days
following ischemia-reperfusion (Fig.
5I and N). However, in the CIL-ischemia-group, only a few F-J
B-positive cells were detected, and the relative number of F-J
B-positive cells was ~24% that of the vehicle-ischemia-group
(Fig. 5L and N); this finding was
identical to a previous study of the authors (22).
Discussion
Just five min of transient cerebral ischemia results
in the damage/death of pyramidal neurons in the hippocampus and
neuronal death occurs selectively in the hippocampal CA1 region
(27,28). It has been reported (6,29)
that pyramidal cells of the stratum pyramidale in the hippocampal
CA1 region die several days following transient cerebral ischemia.
Mongolian gerbils have been generally used as a good experimental
animal model of transient cerebral ischemia as the posterior
communicating arteries in Willis' circle, which connect the
vertebrabasilar and carotid arterial system, are lacking in gerbils
and transient cerebral ischemia can easily be made by the ligation
of bilateral common carotid arteries (24,30,31).
The present study observed the death of pyramidal
cells in the hippocampal CA1 region by NeuN immunohistochemistry
and F-J B histofluorescence; a noticeable loss of CA1 pyramidal
cells was identified in the stratum pyramidale of the CA1 region 5
days following ischemia-reperfusion. This result corresponds to
findings of previous studies (23,32,33).
In addition, it was identified that the oral pre-treatment of 200
mg/kg CIL to the gerbils protected CA1 pyramidal cells (~67% of the
sham-operated gerbils) from 5 min of transient cerebral ischemia;
this finding was identical to a previous study of the authors
(22).
Although mechanisms regarding neuronal death by
transient cerebral ischemic insult are complex, it has been
demonstrated that, among the mechanisms, endogenous antioxidant
enzymes are associated with neuroprotection via the efficient
scavenging of ROS (2,9,34,35).
Excessive ROS production is a cause of neuronal damage/death
following ischemia-reperfusion injury and has been implicated in
the development of numerous neurologic disorders and brain
dysfunctions (10,36–38).
Accumulated ROS cause the injurious modification of cellular
elements including DNA, proteins and lipids; eventually, the
accumulated ROS can impair cellular function and result in neuronal
damage/death (39,40). In the present study, SOD1, SOD2,
CAT and GPX immunoreactivities of the vehicle-ischemia-group were
significantly decreased and barely identified in the CA1 pyramidal
cells 5 days following ischemia-reperfusion. This result is
coincident with the finding of a previous study using gerbils
(40).
Kim et al (41) investigated the protective effect of
Chrysanthemum indicum ethanol extract against
cisplatin-induced nephrotoxicity in vitro. Their findings
may be associated with the antioxidative effects of
Chrysanthemum indicum ethanol extract since the
Chrysanthemum indicum ethanol extract pre-treated group
demonstrated a recovery of serum renal function index with
ameliorated oxidative stress; the effect has not been investigated
in any ischemic stroke model and may be a subject for future
studies. In the present study, CIL pre-treatment significantly
enhanced the immunoreactivities of SOD1, CAT and GPX, although not
SOD2, in the CA1 pyramidal cells of the vehicle-sham-group, and
SOD1, SOD2, CAT and GPX immunoreactivities in the CA1 pyramidal
cells were significantly higher compared with the
vehicle-ischemia-group. These results suggested that the
administration of CIL increases antioxidant enzymes and it exhibits
neuroprotection following transient cerebral ischemia.
ROS are scavenged by SODs, GPX and CAT, and
functions of the antioxidant enzymes have been studied by a number
of researchers. It has been reported that SOD1 overexpression
demonstrated a neuroprotective effect in the hippocampal CA1 region
against cerebral ischemic insults in rodents (42,43).
Kondo et al (44) reported
that SOD1 knockout mice had demonstrated the increase of cell death
and edema of the brain following focal cerebral ischemia, and
Murakami et al (45)
demonstrated that, in SOD2 knockout mice, exacerbated infarct
volume was identified in the brain following permanent focal
cerebral ischemia, and suggested that SOD2 was an important enzyme
in protecting brain from ischemic injury. It has also been reported
that the administration of PEP-1-CAT fusion protein demonstrated
significant neuroprotection in the hippocampal CA1 region following
transient cerebral ischemia (46).
Furthermore, it was recently reported that GPX, which is another
antioxidant enzyme contributing to H2O2
scavenging, exhibited a stronger neuroprotective antioxidant
against oxidative stress than SOD (8).
It has been demonstrated that CIL is associated with
the inhibition of inflammatory responses (15,16,19,22,47).
Cheng et al (15) reported
that a butanol soluble fraction of CIL possessed anti-inflammatory,
immunomodulatory and mononuclear phagocytic activities by the
enhancement of serum IgG and IgM levels in response to sheep red
blood cells in cyclophosphamide-induced mice, and Cheon et
al (19) demonstrated that CIL
suppressed the production of inflammatory mediators and
proinflammatory cytokines via the downregulation of nuclear factor
κB and mitogen-activated protein kinases in RAW264.7 macrophages
(15,19). Previously, Kim et al
(48) reported that CIL protected
against 1-methyl-4-phenylpridinium ions and
lipopolysaccharide-induced cytotoxicity in a cellular model of
Parkinson's disease. In addition, Yoo et al (22) recently reported that CIL
pre-treatment increased anti-inflammatory cytokines in the
hippocampus and that the increased anti-inflammatory cytokines were
associated with neuroprotection in the gerbil hippocampus induced
by transient cerebral ischemia.
In brief, the present study identified that CIL
pre-treatment enhanced SOD1, CAT and GPX, although not SOD2, in
pyramidal cells in the gerbil hippocampal CA1 region and protected
the cells from transient cerebral ischemia. These results indicated
that CIL-mediated neuroprotective effect may be associated with
increases of antioxidant enzymes in the CA1 pyramidal cells and
suggested that CIL may be used for the prevention of ischemic
damage in the brain.
Acknowledgements
The present study was supported by the Bio-Synergy
Research Project (grant no. NRF-2015M3A9C4076322) of the Ministry
of Science, ICT and Future Planning through the National Research
Foundation, and by the Bio and Medical Technology Development
Program of the NRF funded by the Korean Government, Minister of
Science, ICT and Future Planning (grant no.
NRF-2015M3A9B6066835).
References
|
1
|
Roger VL, Go AS, Lloyd-Jones DM, Adams RJ,
Berry JD, Brown TM, Carnethon MR, Dai S, de Simone G, Ford ES, et
al: Heart disease and stroke statistics-2011 update: A report from
the American heart association. Circulation. 123:e18–e209. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liang G, Shi B, Luo W and Yang J: The
protective effect of caffeic acid on global cerebral
ischemia-reperfusion injury in rats. Behav Brain Funct. 11:182015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
White BC, Grossman LI and Krause GS: Brain
injury by global ischemia and reperfusion: A theoretical
perspective on membrane damage and repair. Neurology. 43:1656–1665.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Butler TL, Kassed CA, Sanberg PR, Willing
AE and Pennypacker KR: Neurodegeneration in the rat hippocampus and
striatum after middle cerebral artery occlusion. Brain Res.
929:252–260. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Crain BJ, Westerkam WD, Harrison AH and
Nadler JV: Selective neuronal death after transient forebrain
ischemia in the Mongolian gerbil: A silver impregnation study.
Neuroscience. 27:387–402. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kirino T: Delayed neuronal death.
Neuropathology. 20:(Suppl). 95–97. 2000. View Article : Google Scholar
|
|
7
|
Lewén A, Matz P and Chan PH: Free radical
pathways in CNS injury. J Neurotrauma. 17:871–890. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee JC and Won MH: Neuroprotection of
antioxidant enzymes against transient global cerebral ischemia in
gerbils. Anat Cell Biol. 47:149–156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park JH, Park OK, Yan B, Ahn JH, Kim IH,
Lee JC, Kwon SH, Yoo KY, Lee CH, Hwang IK, et al: Neuroprotection
via maintenance or increase of antioxidants and neurotrophic
factors in ischemic gerbil hippocampus treated with tanshinone I.
Chin Med J (Engl). 127:3396–3405. 2014.PubMed/NCBI
|
|
10
|
Chan PH: Reactive oxygen radicals in
signaling and damage in the ischemic brain. J Cereb Blood Flow
Metab. 21:2–14. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Delanty N and Dichter MA: Antioxidant
therapy in neurologic disease. Arch Neurol. 57:1265–1270. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pastore A, Petrillo S, Piermarini E and
Piemonte F: Systemic redox biomarkers in neurodegenerative
diseases. Curr Drug Metab. 16:46–70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Duan X, Wang W, Liu X, Yan H, Dai R and
Lin Q: Neuroprotective effect of ethyl acetate extract from
gastrodia elata against transient focal cerebral ischemia in rats
induced by middle cerebral artery occlusion. J Tradit Chin Med.
35:671–678. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Surapaneni S, Prakash T, Ansari M,
Manjunath P, Kotresha D and Goli D: Study on cerebroprotective
actions of Clerodendron glandulosumleaves extract against
long term bilateral common carotid artery occlusion in rats. Biomed
Pharmacother. 80:87–94. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cheng W, Li J, You T and Hu C:
Anti-inflammatory and immunomodulatory activities of the extracts
from the inflorescence of Chrysanthemum indicum Linné. J
Ethnopharmacol. 101:334–337. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee DY, Choi G, Yoon T, Cheon MS, Choo BK
and Kim HK: Anti-inflammatory activity of Chrysanthemum
indicum extract in acute and chronic cutaneous inflammation. J
Ethnopharmacol. 123:149–154. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shunying Z, Yang Y, Huaidong Y, Yue Y and
Guolin Z: Chemical composition and antimicrobial activity of the
essential oils of Chrysanthemum indicum. J Ethnopharmacol.
96:151–158. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang C, Qin MJ, Shu P, Hong JL, Lu L and
He DX: Chemical variations of the essential oils in flower heads of
Chrysanthemum indicum L. From China. Chem Biodivers.
7:2951–2962. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheon MS, Yoon T, Lee DY, Choi G, Moon BC,
Lee AY, Choo BK and Kim HK: Chrysanthemum indicum Linné
extract inhibits the inflammatory response by suppressing NF-kappaB
and MAPKs activation in lipopolysaccharide-induced RAW 264.7
macrophages. J Ethnopharmacol. 122:473–477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dekanski D, Selaković V, Piperski V,
Radulović Z, Korenić A and Radenović L: Protective effect of olive
leaf extract on hippocampal injury induced by transient global
cerebral ischemia and reperfusion in Mongolian gerbils.
Phytomedicine. 18:1137–1143. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rocher MN, Carré D, Spinnewyn B, Schulz J,
Delaflotte S, Pignol B, Chabrier PE and Auguet M: Long-term
treatment with standardized Ginkgo biloba extract (EGb 761)
attenuates cognitive deficits and hippocampal neuron loss in a
gerbil model of vascular dementia. Fitoterapia. 82:1075–1080. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yoo KY, Kim IH, Cho JH, Ahn JH, Park JH,
Lee JC, Tae HJ, Kim DW, Kim JD, Hong S, et al: Neuroprotection of
Chrysanthemum indicum Linne against cerebral
ischemia/reperfusion injury by anti-inflammatory effect in gerbils.
Neural Regen Res. 11:270–277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim IH, Yan BC, Park JH, Yeun GH, Yim Y,
Ahn JH, Lee JC, Hwang IK, Cho JH, Kim YM, et al: Neuroprotection of
a novel synthetic caffeic acid-syringic acid hybrid compound
against experimentally induced transient cerebral ischemic damage.
Planta Med. 79:313–321. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim DW, Lee JC, Cho JH, Park JH, Ahn JH,
Chen BH, Shin BN, Tae HJ, Seo JY, Cho JH, et al: Neuroprotection of
ischemic preconditioning is mediated by anti-inflammatory, not
pro-inflammatory, cytokines in the gerbil hippocampus induced by a
subsequent lethal transient cerebral ischemia. Neurochem Res.
40:1984–1995. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Candelario-Jalil E, Alvarez D, Merino N
and León OS: Delayed treatment with nimesulide reduces measures of
oxidative stress following global ischemic brain injury in gerbils.
Neurosci Res. 47:245–253. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee JC, Kim IH, Cho GS, Park JH, Ahn JH,
Yan BC, Kwon HM, Kim YM, Cheon SH, Cho JH, et al: Ischemic
preconditioning-induced neuroprotection against transient cerebral
ischemic damage via attenuating ubiquitin aggregation. J Neurol
Sci. 336:74–82. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Petito CK, Olarte JP, Roberts B, Nowak TS
Jr and Pulsinelli WA: Selective glial vulnerability following
transient global ischemia in rat brain. J Neuropathol Exp Neurol.
57:231–238. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sugawara T, Fujimura M, Morita-Fujimura Y,
Kawase M and Chan PH: Mitochondrial release of cytochrome c
corresponds to the selective vulnerability of hippocampal CA1
neurons in rats after transient global cerebral ischemia. J
Neurosci. 19:RC391999.PubMed/NCBI
|
|
29
|
Zhang QG, Wang R, Han D, Dong Y and Brann
DW: Role of Rac1 GTPase in JNK signaling and delayed neuronal cell
death following global cerebral ischemia. Brain Res. 1265:138–147.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Levine S and Sohn D: Cerebral ischemia in
infant and adult gerbils. Relation to incomplete circle of Willis.
Arch Pathol. 87:315–317. 1969.PubMed/NCBI
|
|
31
|
Martínez NS, Machado JM, Pérez-Saad H,
Coro-Antich RM, Berlanga-Acosta JA, Salgueiro SR, Illera GG, Alba
JS and del Barco DG: Global brain ischemia in Mongolian gerbils:
Assessing the level of anastomosis in the cerebral circle of
Willis. Acta Neurobiol Exp (Wars). 72:377–384. 2012.PubMed/NCBI
|
|
32
|
Kim IH, Yoo KY, Park JH, Yan BC, Ahn JH,
Lee JC, Kwon HM, Kim JD, Kim YM, You SG, et al: Comparison of
neuroprotective effects of extract and fractions from Agarum
clathratum against experimentally induced transient cerebral
ischemic damage. Pharm Biol. 52:335–343. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Park JH, Shin BN, Chen BH, Kim IH, Ahn JH,
Cho JH, Tae HJ, Lee JC, Lee CH, Kim YM, et al: Neuroprotection and
reduced gliosis by atomoxetine pretreatment in a gerbil model of
transient cerebral ischemia. J Neurol Sci. 359:373–380. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Numagami Y, Sato S and Ohnishi ST:
Attenuation of rat ischemic brain damage by aged garlic extracts: A
possible protecting mechanism as antioxidants. Neurochem Int.
29:135–143. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tu Q, Wang R, Ding B, Zhong W and Cao H:
Protective and antioxidant effect of Danshen polysaccharides on
cerebral ischemia/reperfusion injury in rats. Int J Biol Macromol.
60:268–271. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chan PH: Oxygen radicals in focal cerebral
ischemia. Brain Pathol. 4:59–65. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rodrigo R, Fernández-Gajardo R, Gutiérrez
R, Matamala JM, Carrasco R, Miranda-Merchak A and Feuerhake W:
Oxidative stress and pathophysiology of ischemic stroke: Novel
therapeutic opportunities. CNS Neurol Disord Drug Targets.
12:698–714. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mantha AK, Sarkar B and Tell G: A short
review on the implications of base excision repair pathway for
neurons: Relevance to neurodegenerative diseases. Mitochondrion.
16:38–49. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sugawara T and Chan PH: Reactive oxygen
radicals and pathogenesis of neuronal death after cerebral
ischemia. Antioxid Redox Signal. 5:597–607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Park JH, Cho JH, Kim IH, Ahn JH, Lee JC,
Chen BH, Shin BN, Tae HJ, Yoo KY, Hong S, et al: Oenanthe
Javanica extract protects against experimentally induced
ischemic neuronal damage via its antioxidant effects. Chin Med J
(Engl). 128:2932–2937. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim TW, Kim YJ, Park SR, Seo CS, Ha H,
Shin HK and Jung JY: Chrysanthemum indicum attenuates
cisplatin-induced nephrotoxicity both in vivo and in vitro. Nat
Prod Commun. 10:397–402. 2015.PubMed/NCBI
|
|
42
|
Murakami K, Kondo T, Epstein CJ and Chan
PH: Overexpression of CuZn-superoxide dismutase reduces hippocampal
injury after global ischemia in transgenic mice. Stroke.
28:1797–1804. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chan PH, Kawase M, Murakami K, Chen SF, Li
Y, Calagui B, Reola L, Carlson E and Epstein CJ: Overexpression of
SOD1 in transgenic rats protects vulnerable neurons against
ischemic damage after global cerebral ischemia and reperfusion. J
Neurosci. 18:8292–8299. 1998.PubMed/NCBI
|
|
44
|
Kondo T, Reaume AG, Huang TT, Carlson E,
Murakami K, Chen SF, Hoffman EK, Scott RW, Epstein CJ and Chan PH:
Reduction of CuZn-superoxide dismutase activity exacerbates
neuronal cell injury and edema formation after transient focal
cerebral ischemia. J Neurosci. 17:4180–4189. 1997.PubMed/NCBI
|
|
45
|
Murakami K, Kondo T, Kawase M, Li Y, Sato
S, Chen SF and Chan PH: Mitochondrial susceptibility to oxidative
stress exacerbates cerebral infarction that follows permanent focal
cerebral ischemia in mutant mice with manganese superoxide
dismutase deficiency. J Neurosci. 18:205–213. 1998.PubMed/NCBI
|
|
46
|
Kim DW, Kim DS, Kim MJ, Kwon SW, Ahn EH,
Jeong HJ, Sohn EJ, Dutta S, Lim SS, Cho SW, et al: Imipramine
enhances neuroprotective effect of PEP-1-Catalase against ischemic
neuronal damage. BMB Rep. 44:647–652. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kim JE, Jun S, Song M, Kim JH and Song YJ:
The extract of Chrysanthemum indicum Linne inhibits EBV
LMP1-induced NF-κB activation and the viability of EBV-transformed
lymphoblastoid cell lines. Food Chem Toxicol. 50:1524–1528. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kim IS, Ko HM, Koppula S, Kim BW and Choi
DK: Protective effect of Chrysanthemum indicum Linne against
1-methyl-4-phenylpridinium ion and lipopolysaccharide-induced
cytotoxicity in cellular model of Parkinson's disease. Food Chem
Toxicol. 49:963–973. 2011. View Article : Google Scholar : PubMed/NCBI
|














