Introduction
Peroxisome proliferator-activated receptor gamma
(PPAR γ) is a ligand-dependent nuclear receptor and plays a
critical role in fat metabolism and insulin sensitivity (1). PPAR γ is expressed in adipose
tissues, kidney, stomach, heart, liver, spleen, and brain (2). Recently, PPAR γ is best known for its
role in inflammatory and immunological responses (2–4).
Many inflammatory cells, such as lymphocytes, neutrophils, and
macrophages, express PPAR γ, and are the potential targets of PPAR
γ-mediated inhibitory functions (2). PPAR γ agonists have been demonstrated
as therapeutic agents in autoimmunity and allergic diseases
(2,3,5–7). In
monocyte/macrophage system, the relationship between PPAR γ
activation and negative regulation of cytokines production has been
demonstrated (4,8,9).
Other inflammatory cells, such as mast cells, may be potential
targets of PPAR γ. Although previous studies reported that mast
cells express PPAR γ (9–11), it remains obscure how PPAR γ
signaling affects the nature of mast cells. The function and
mechanism of PPAR γ in mast cells should be studied further.
Mast cells play an important role in immediate
hypersensitivity and allergic diseases via the cross-linking of
IgE/high affinity IgE receptors (FcRIs) by multivalent allergens
(12). The cross-linking results
in degranulation, prostaglandin and leukotriene synthesis, and
production of various cytokines and chemokines (13), which cause the early and late phase
of allergic responses. Mast cells derive from bone marrow stem
cells, subsequently flow into blood stream in the immature stage,
and differentiate at the peripheral tissues. The growth,
differentiation, maturation, and survival of mast cells in bone
marrow and in various tissues are largely mediated by signals of
kit receptor (CD117) and interleukin (IL)-3 receptor (14). After stimulation with stem cell
factor (SCF, kit ligand) and IL-3, the immature mast cells
differentiate into mature stage. PPAR γ expression has been
reported in bone marrow-derived mast cells (BMMCs), and PPAR γ
ligand administration can attenuate atopic and contact dermatitis
in mice (2). PPAR γ activation
dramatically suppresses neural stem cell differentiate into
maturate neurons (15). Thus, we
speculate that the development and maturation of bone marrow
progenitors might be regulated by PPAR γ pathway.
Given the evidence of negative effects of PPAR γ
agonist on allergic diseases, we have sought to examine how PPAR γ
agonist affects mast cell development from bone marrow progenitors.
Our findings indicate that PPAR γ agonist reduces the phenotypic
markers and viability of mast cells, inhibits degranulation, and
induces cell apoptosis. These data suggest that PPAR γ may be a
regulator of mast cells response and a novel therapeutical strategy
for treatment of mast cell-related diseases.
Materials and methods
Cell culture and reagents
Bone marrow cells were flushed and collected from
the femurs and tibias of C57BL/6J female mice (6–10 weeks old)
(Shanghai SLAC laboratory Animal Co., Ltd., Shanghai, China) and
cultured in complete RPMI (RPMI-1640; Corning Cellgro, Manassas,
VA, USA) containing 10% fetal bovine serum (FBS) (Gibco; Thermo
Fisher Scientific, Waltham, MA, USA), 2 mM L-glutamine, 100 U/ml
penicillin, 100 mg/ml streptomycin, 1 mM sodium pyruvate, and 1 mM
HEPES, supplemented with IL-3 (10 ng/ml) and SCF (10 ng/ml) (both
from Peprotech, Inc., Rocky Hill, NJ, USA), with or without
pioglitazone (PIO, 0–20 µM) (R&D Systems, Inc., Minneapolis,
MN, USA) or vehicle (DMSO). Cultures were incubated for the
indicated times. Every 4 days, the nonadherent population was
collected and half of the medium and cytokines were replaced. For
system evaluation, mice were treated with or without PIO
hydrochloride (30 mg/kg body weight/d; Actos; Takeda
Pharmaceuticals, Ltd., Osaka, Japan) by oral gavage for one week,
and the bone marrow cells were harvested for further study.
Assessment of differentiation
BMMCs were harvested after 1–8 weeks and
differentiation was monitored on a weekly basis. To detect the
development of BMMCs, cells were washed and assessed for surface
expression of CD117 and FcεRI α. Briefly, cells were stained with a
combination of PE-conjugated anti-mouse CD117 and FITC-conjugated
anti-mouse FcεRI α (BioLegend, Inc., San Diego, CA, USA) for 30 min
at 4°C. Then, cells were washed and subjected to flow cytometry
analysis (BD Biosciences, San Jose, CA, USA). Negative controls
included isotype-matched conjugated, nonspecific antibodies
(BioLegend, Inc.). Data were analyzed using the FlowJo software
(version 7.5.1; FlowJo, LLC, Ashland, OR, USA).
Reverse transcripton-polymerase chain
reaction analysis
Total RNA was extracted from BMMCs using Trizol
reagent (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer's instructions. The first strand cDNA was
reverse-transcribed from 1 µg of total RNA by using oligo
(dT)12–18 primer (Takara Bio, Inc., Otsu, Japan). The
reaction mixture was amplified with the oligonucleotides specific
for mast cell protease (MCP)-6, PPAR γ, and β-actin using ABI 7500
Real-Time PCR System and SYBR-Green PCR master mix (Takara).
Specific primer sequences are as follows: for MCP-6: sense
5′-CCACTGGTCTGCAAAGTGAA-3′ and antisense
5′-CAGAGGACAAGGAAGGCAAG-3′; for PPAR γ: sense
5′-TGACACAGAGATCGCATTCTGG-3′ and antisense 5′- ACA GAC ACG ACA TTC
AAT TGC C-3′; for β-actin: sense 5′-CCCATCTACGAGGGCTAT-3′ and
antisense 5′-TGTCACGCACGATTTCC-3′.
Cell viability and characterization of
BMMCs
BMMCs were starved by replacing complete medium with
RPMI without serum and IL-3/SCF for 6 h. Cells were seeded into
96-well plates at a density of 25×104/ml and stimulated with or
without PIO in complete medium for 48 h. The relative cell
viability was determined using Alamar-Blue Cell Viability assay
according to the manufacturer's instructions (BioSource, Camarillo,
CA, USA). Briefly, at the end of incubation, each well was supplied
with 10 µl of AlamarBlue reagent. Plates were incubated at 37°C for
2 h, and absorbance was measured at 490 nm with Omega microplate
reader (Ingen Technologies, Inc., Alexandria, VA, USA).
BMMCs were harvested after 4 weeks and washed with
PBS. Cells were centrifuged onto glass slides. Cells were fixed in
Carnoy's solution. To investigate the maturation of mast cells,
granules were stained with toluidine blue (Sigma-Aldrich, St.
Louis, MO, USA) and photomicrographs were taken at a magnification
of ×400 using Leica Microsystems Inc. (Bannockburn, IL, USA).
Degranulation assay
The degranulation of BMMCs was detected by measuring
the activity of a granular enzyme, β-hexosaminidase. After 4 weeks,
BMMCs were sensitized with 100 ng/ml anti-DNP IgE overnight. The
sensitized cells were stimulated with 100 ng/ml of DNP-BSA in
Tyrode's buffer at 37°C for 30 min, and the supernatant and pellet
were collected. Release of β-hexosaminidase was evaluated using
p-nitrophenyl-N-acetyl-β-D-glucosamine (Sigma-Aldrich). The
percentage of β-hexosaminidase release was expressed as 100 ×
supernatant activity/(supernatant activity + cell lysate activity)
as described previously (16).
Apoptosis assay
Mast cells apoptosis was assessed by Annexin V
assay. Briefly, BMMCs were washed once in 1 ml of 1xAnnexin V
binding buffer. The supernatants were removed by centrifugation and
cells were suspended in 1xAnnexin V binding buffer. A 5 µl of
FITC-conjugated Annexin V (BD Biosciences) and 5 µl of prodium
iodide were added to the cells. Cells were mixed gently and
incubated for 15 min at room temperature in the dark, and diluted
with 400 µl of 1xAnnexin V binding buffer. Data were analyzed by
flow cytometry within 1 h.
Statistical analysis
Results are expressed as mean ± standard deviation
(SD). Statistical analysis was performed by Student's t-test or
ANOVA using SPSS Version 19 (IBM, Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of IL-3 and SCF on development
of mast cells cultured from bone marrow progenitors
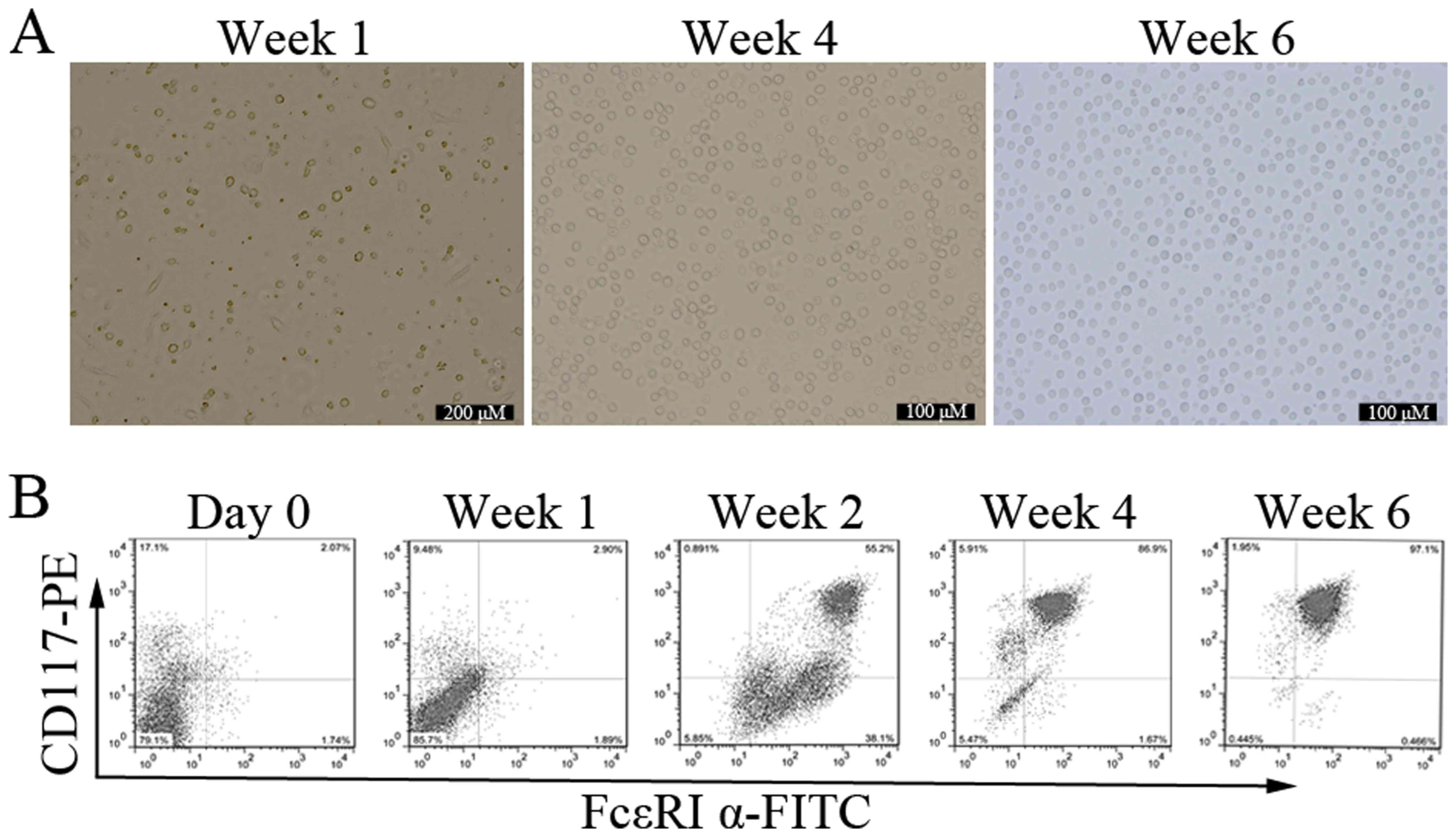
To observe the mature process of mast cells in
vitro, bone marrow cells were cultured with different
concentrations of IL-3 and SCF for up to 8 weeks. This system is
widely used to study mast cells development, and yields cells
closely resembling the in vivo counterparts (17). Since the driving effects of IL-3
and SCF on development of mast cells became more pronounced when
the concentration was 10 ng/ml, we conducted the subsequent
experiments with 10 ng/ml for IL-3 and SCF. After 2 days of
culture, adherent cells began to appear, and the cells were
relatively single, short shuttle-like or round. Suspension cells
varied widely in shape and size. With the extending of incubation
time, the number of adherent cells was less and less, and the
suspension cells number gradually increased. The form of suspension
cells was uniform, round-shaped, and bright by the end of week 6
(Fig. 1A).
The expression of two specific mast cell markers,
CD117 and FcεRI α, was investigated. Both positive cells are
defined mature mast cells (2). The
mean percentage of cells co-expressing CD117 and FcεRI α in freshly
isolated bone marrow progenitors was 2.13±0.56% (n=3). The
maturation of BMMCs was significantly enhanced in cells receiving
IL-3 and SCF with time. At the end of week 1, 2, 4, and 6, the
percentage of CD117/FcεRI α positive cells were 3.05±1.15%,
52.67±3.45%, 87.43±4.87%, and 98.16±3.97% respectively, n=3
(Fig. 1B). When BMMCs were
cultured for more than 8 weeks, the percentage of CD117/FcεRI α
positive cells decreased to about 60% (data not shown). These
results indicate that bone marrow progenitors can be stably induced
into mature mast cells in vitro in the presence of IL-3 and
SCF.
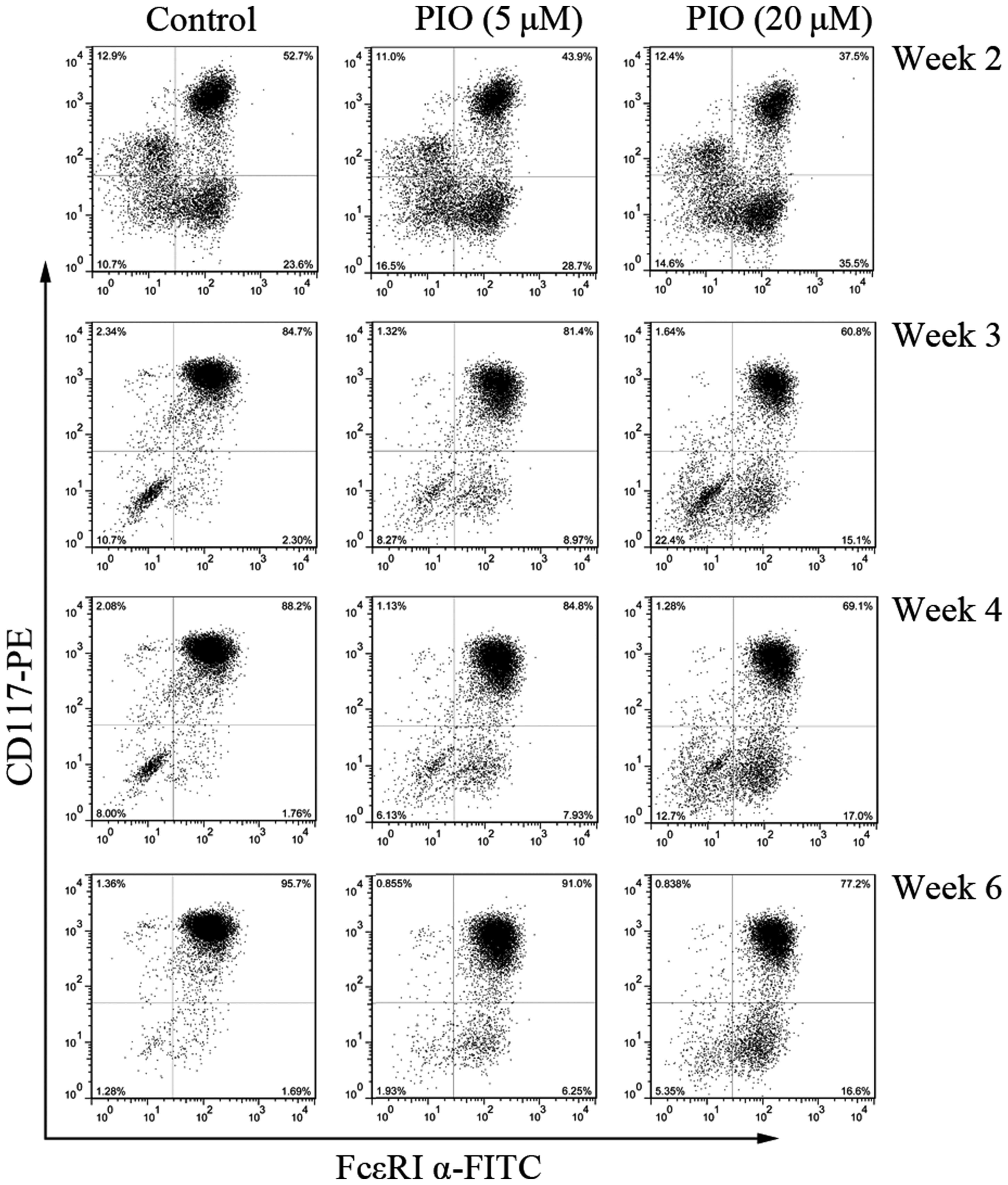
PPAR γ agonist inhibits cell-surface
antigen expression on BMMCs
PPAR γ plays an important role on mast cells
differentiation and is potentially useful for the therapy in
allergic disorders (2,3,7,9). To
evaluate the effects of PPARγ agonist on mast cells development, we
measured the expression of CD117/FcεRI α after culture in IL-3 and
SCF with different concentrations of PIO (0, 5, and 20 µM). PIO
treatment decreased cell surface expression of CD117 and FcεRI α in
a concentration-dependent manner, with significant reduction in
surface antigens expression at 20 µM (Fig. 2).
The percentage of cells co-expressing CD117 and
FcεRI α was consistently reduced by greater than 28% by week 2 and
3 with PIO at 20 µM. However, CD117 expression was more sensitive
to the inhibitory effect of PIO than that of FcεRI α. Similar
results were obtained at the end of week 4 and 6. These results
indicate that PPAR γ agonist can decrease the expression of
specific mast cell antigens and inhibit mast cells maturation.
PPAR γ agonist suppresses mast cells
viability and induces cell apoptosis
The inhibitory effects of PIO on BMMCs could be due
to proapoptotic effect. To demonstrate the effects of PPAR γ
agonist on BMMCs, cell viability was determined by Alamar-Blue
assay. After 4 weeks of culture, the percentage of mature BMMCs was
increased to more than 85% (Fig.
1B). Before cell viability assay, cells were starved for 6 h in
medium without FBS and cytokines. Then BMMCs were cultured in
complete medium with IL-3 and SCF for a further 7 days, with or
without PIO (5–20 µM). In the presence of PIO, the total number of
mast cells was decreased and the cell viability was inhibited in a
dose-dependent manner (Fig. 3A).
The viability of BMMCs cultured with PIO (20 µM) was decreased to
50.4±7.71% (n=3). We hypothesized that PIO could induce BMMCs
programmed death. Therefore, the effect of PIO on cell apoptosis
was evaluated 48 h after incubation in the presence or absence of
PIO (20 µM). The percentage of Annexin V+PI+
cells was 19.2±1.51% in PIO treated cultures, vs. 14.9±0.87% in
control (P<0.05) (n=3) (Fig.
3B).
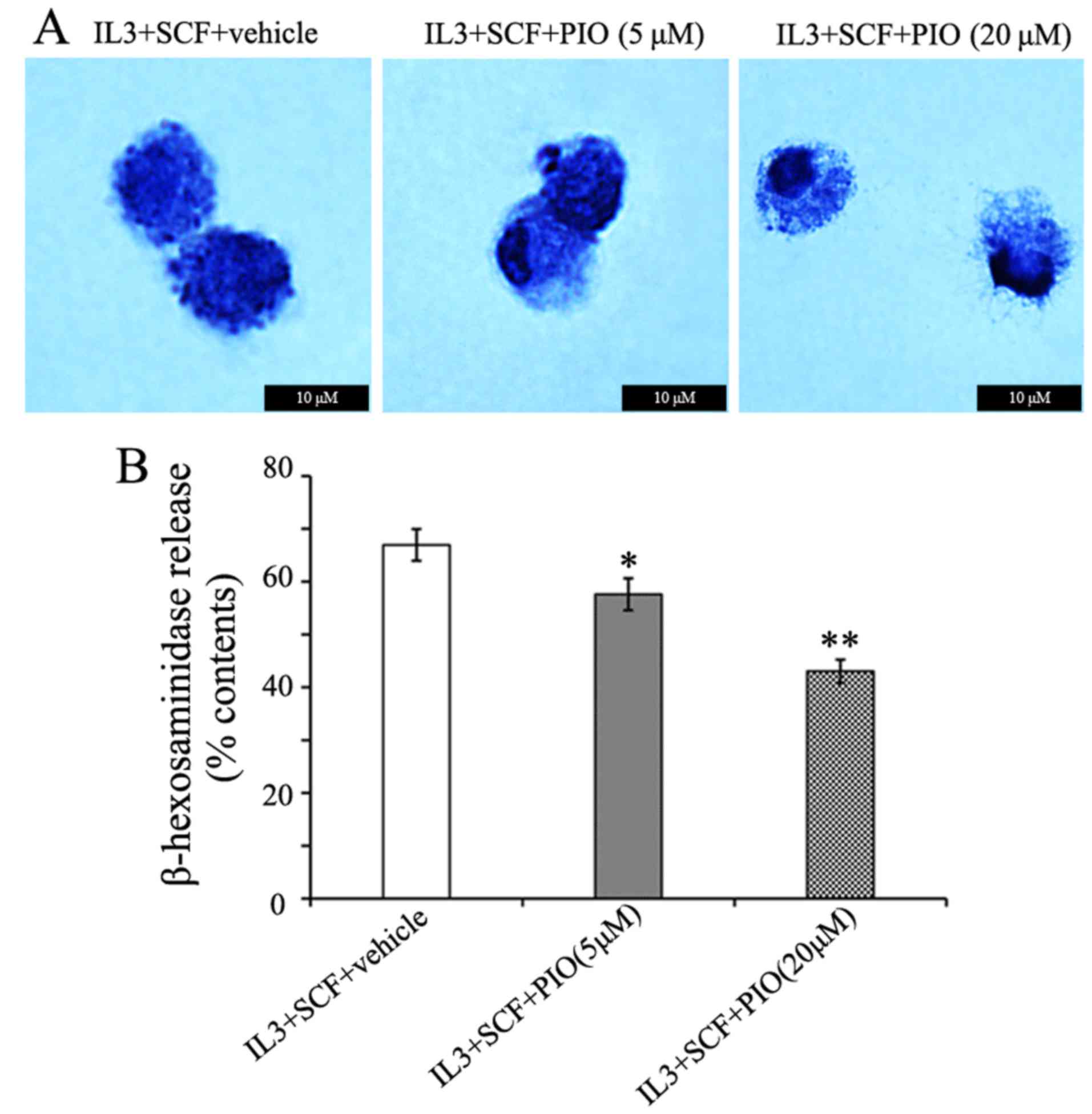
PPAR γ agonist inhibits mast cells
granule formation and the release of β-hexosaminidase
The cross-linking of IgE/FcεRI complexes by
allergens has been shown to coincide with mast cells granulation
(13,18), and enhances the characteristic mast
cells morphology. In the present study, PIO played a weak role in
FcεRI α expression, therefore we determined the effects of PIO on
mast cells morphology. BMMCs cultured in IL-3 and SCF with PIO
(5–20 µM) for 4 weeks were centrifuged onto glass slides and
stained with toluidine blue. The cells had a homogeneous appearance
at either concentration of PIO. However, the treatment of PIO
altered the appearance of cytoplasmic granules from these cells.
While cells cultured with vehicle and PIO at 5 µM showed prominent
granule formation, PIO at 20 µM greatly inhibited granule formation
and resulted in empty vacuoles and swelling of granules (Fig. 4A). The functional effects of PIO on
histamine synthesis and release were determined by measuring
β-hexosaminidase activity. We found that the level of
β-hexosaminidase release were reduced 25% by PIO administration at
20 µM (Fig. 4B). These results
indicate that PIO inhibits granule formation and function of mast
cells.
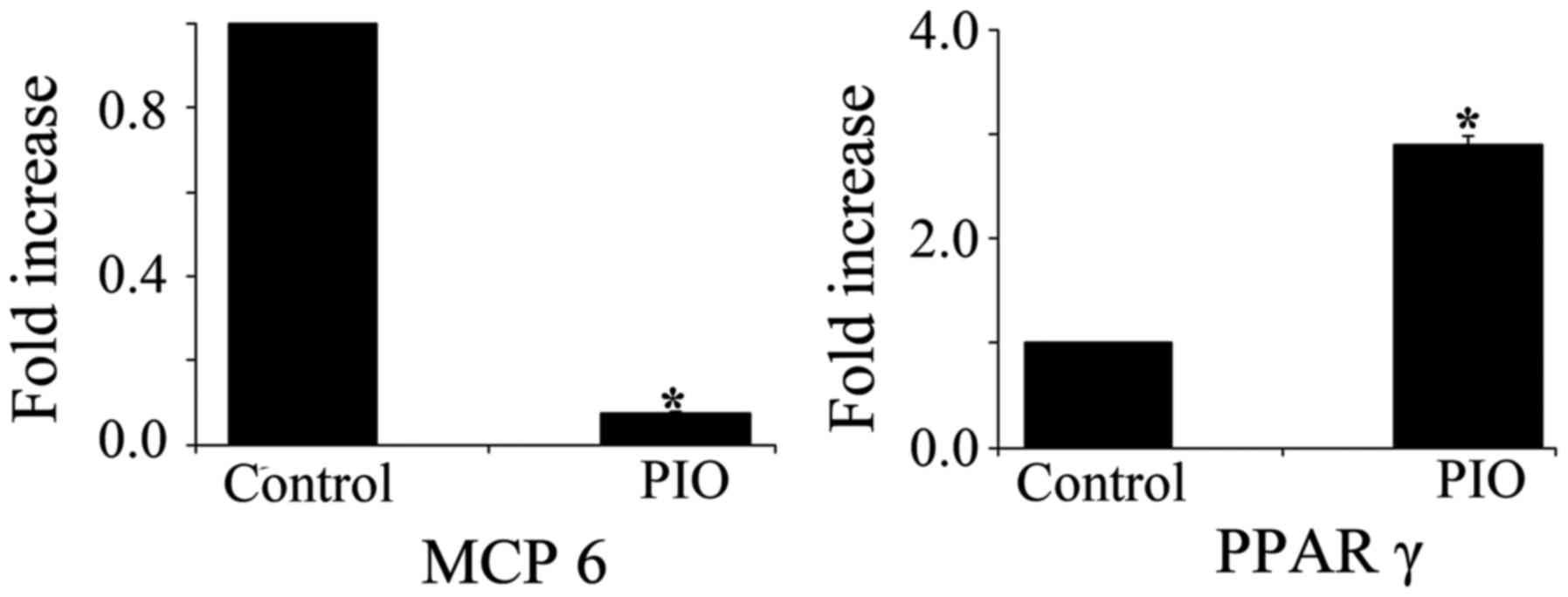
PPAR γ agonist alters the expression
of MCP-6 and PPAR γ mRNAs
It has been reported that MCP-6, a mature mast
cells-specific tryptase, is a potent inflammatory mediator and is
needed for the development of airway hyperresponsiveness (19,20).
To determine whether PPAR γ activation affects mast cells
maturation, we measured the expression levels of MCP-6 and
PPAR γ mRNA in BMMCs treated with PIO. MCP-6 mRNA
expression level was significantly decreased in BMMCs treated with
PIO for 4 weeks when compared with that of control cells. In
contrast, PIO treatment induced a 3-fold increase in PPAR γ
mRNA expression in BMMCs (Fig.
5).
PPAR γ agonist maintains the
inhibitory effects on bone marrow cells in vitro
To determine the role of PPAR γ agonist on BMMCs
development after systemic administration, bone marrow cells from
mice with or without oral gavage with PIO for 7 days were cultured
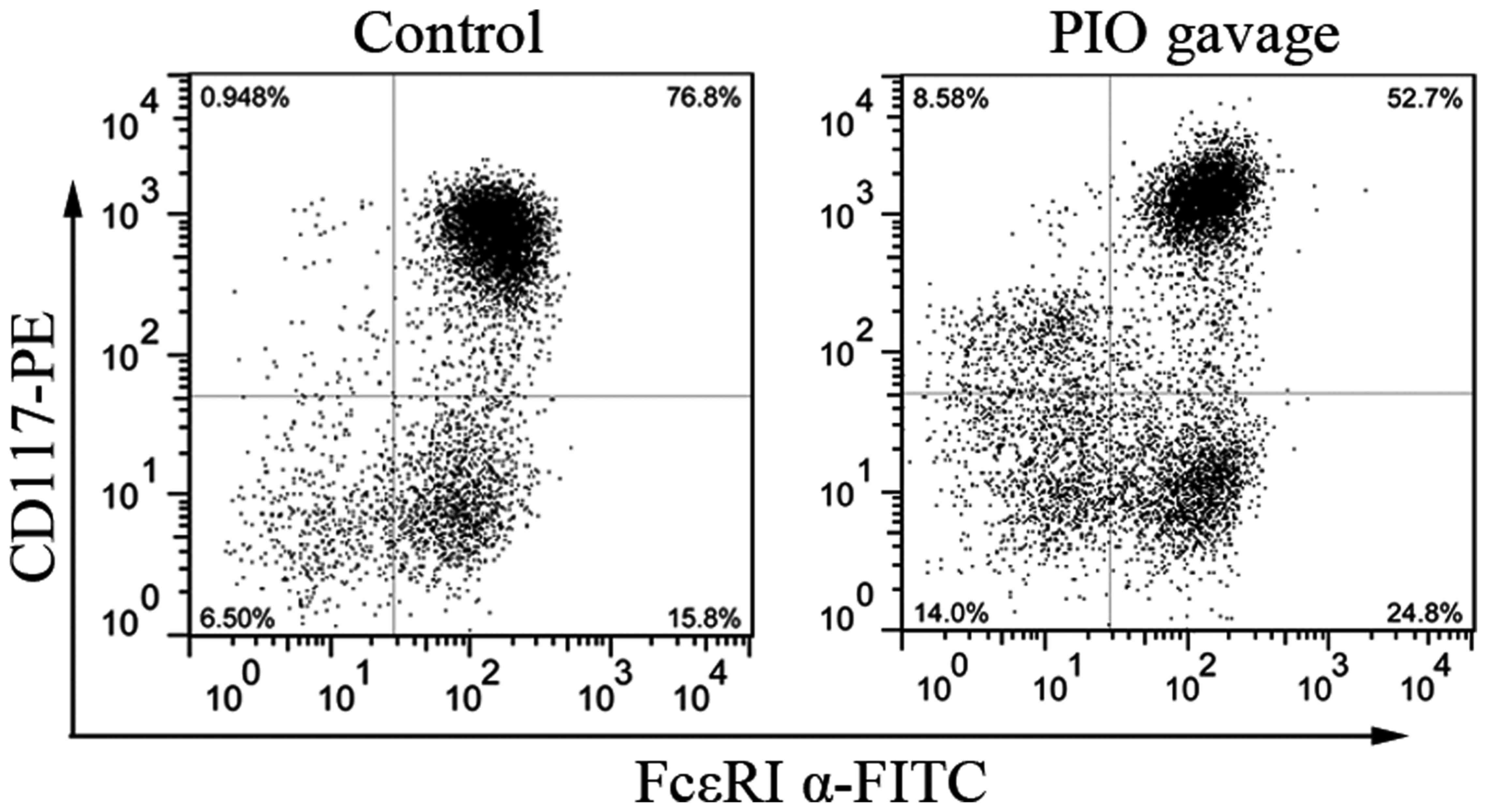
for 3 weeks with IL-3 and SCF. As seen in Fig. 6, the percentage of CD117/FcεRI α
double-positive cells from PIO gavage mice is lower than that from
control mice (56±4.2% vs. 78±3.5%, P<0.05, n=3). These data
indicate that mast cells development from PIO-treated mice is
suppressed partially by PPAR γ activation even in the culture
condition without PIO.
Discussion
We previously reported that PPAR γ agonist
attenuated allergic inflammation in a mouse model of allergic
rhinitis (21). PIO is the
effective agent in suppressing allergic inflammation. However,
there are limited studies on the action mechanism of PPAR γ in mast
cells, which are the total valve of allergic diseases. In the
present study, we investigated the effects of PPAR γ agonist on
mouse mast cells development. Our results show the inhibitory
effects of PPAR γ on mast cells maturation and function. More
importantly, PPAR γ agonist maintains the inhibitory effects on
BMMCs in vitro.
SCF and IL-3 signaling pathways regulate mast cells
development and growth. Mast cells derive from multipotential
hematopoietic stem cells and their differentiation and maturation
occur when they reach the target organ (22,23).
Mast cell progenitors leave the bone marrow, invade connective or
mucosal tissue, and then proliferate and differentiate into
morphologically identifiable mast cells (24). The phenotype of mast cells is
determined by each site of final differentiation (24,25).
SCF binds its receptor, kit, which is distributed on the surface of
mast cells. SCF and kit signaling is essential for the development
of murine mast cells (13). Mast
cells can be developed at high efficiency by culturing BMMCs in
IL-3-containing media. IL-3 stimulates the proliferation of BMMCs
and enhances SCF-dependent mast cells development at low cell
densities (26). In the present
study, we optimized the culture conditions with IL-3 (10 ng/ml) and
SCF (10 ng/ml). We observed a gradual increase in mature mast cells
number during the first 4 weeks of culture, especially by the end
of week 2 when the percentage of CD117/FcεRI α double-positive
cells was dramatically increased to 55%. After 4 weeks, bone marrow
progenitors became uniform in appearance. By the end of week 6, the
percentage of CD117/FcεRI α double-positive cells reached to
>97%. The bone marrow progenitors were effectively induced into
mature mast cells under IL-3 and SCF stimulation in vitro.
We used this culture model for the present study.
PPAR γ is one of the master regulators on mast cells
maturation and potentially useful for therapy in various disorders
involving mast cells activation (2). However, the roles of PPAR γ in mast
cells are too variable to conclude a simple regulatory mechanism of
inflammation (9). Spencer et
al (11), demonstrated that
PIO reduced adipose tissue inflammation through reduction of mast
cells number. As shown in Fig. 2,
PIO greatly inhibited BMMCs maturation in a concentration-dependent
manner at the indicated time points. While under certain
concentration of PIO, the percentage of mature mast cells was
gradually increased. After 8 weeks of culture, there was no
difference in the differentiated degree between the treated and
untreated BMMCs with PIO (data not shown). The results suggest that
PIO inhibits early-stage mast cells maturation.
The number of mast cells in inflamed tissue can be
regulated by proliferation, migration, and survival (and apoptosis)
(13). The inhibitory effects of
PPAR γ on surface antigens expression and granule formation could
be duo to apoptosis. In the present study, we have demonstrated
that PPAR γ agonist is able to inhibit BMMCs cells viability in a
dose-dependent manner. However, Maeyama et al (9) reported the opposite result that
rosiglitazone, another PPAR γ agonist, increased cell viability of
BMMCs from wide type mice. One of the possible explanations is that
they obtained BMMCs with an additional 4-week culture with IL-3
alone before adding SCF. In addition, PIO acting via PPAR γ played
a proapoptotic role in murine mast cells progenitors. The result is
consistent with our previous finding in a mouse model of allergic
rhinitis (7). Our observation
could suggest that PPAR γ can modulate the expression of
proapoptotic molecules in mast cells.
Each mast cell has approximately 75 granules
containing a host of mediators with diverse biological roles that
can be performed or formed de novo (27). Mature mast cells express mouse
MCP-4, 5, 6, and 7, and contain a histamine content that is at
least ten-times higher than that of immature ones (2). Histamine is involved in various
physiological responses and is released from the cytoplasmic
granules upon IgE-dependent antigen stimulation (28). Histamine promotes granule
maturation in mast cells and acts as a proinflammatory mediator
(29). Degranulation of mast cells
was decreased by PIO treatment in a dose-dependent manner,
concomitant with defective granule formation. Thus, these findings
highlight two possibilities: one is PIO directly inhibited granule
formation of mast cells, and the other is that the decreased
release of histamine affected granule maturation. MCP-6, one of
mast cell-derived granule components, is secreted from mature mast
cells (30). MCP-6 can activate
the protease-activated receptors 1 and 2 to modulate the activities
of target cells (31). It has been
reported that MCP-6 is a proinflammatory mediator in various
conditions (32–34). The development of airway high
reactivity in a model of allergic asthma is dependent on MCP-6
(19). In our study, the
expression of MCP-6 mRNA was decreased after treatment with
PIO, which is consistent with the inhibited formation of the
granule by PIO. Recent studies have shown that PPAR γ ligands
suppressed antigen-induced cytokine production by MBBCs at both
protein and mRNA levels (9,10).
As shown in the present study, PPAR γ mRNA expression of
BMMCs was increased by PIO treatment. These results suggested that
PPAR γ agonist might amplify the inhibitory effects of PPAR γ on
BMMCs by increasing PPAR γ expression.
PPAR γ agonist plays an inhibitory effect on mast
cells maturation in vitro, which raises a possibility that
PPAR γ is involved in the physiological property of mast cell
progenitors. We demonstrated our hypothesis by culturing BMMCs
prepared from mice with PIO gavage. Even under the same culture
condition in vitro, the percentage of CD117/FcεRI α
double-positive cells was significantly decreased in BMMCs from PIO
gavage mice than that from control mice. Systemic treatment of PIO
suppressed surface antigens expression of mature mast cells in
vitro. Thus, PPAR γ activation can maintain the inhibitory
effects on bone marrow cells for a while in absence of PIO.
In summary, the present study has demonstrated that
PPAR γ affects mouse mast cell progenitors by inhibiting granule
formation and suppressing degranulation in response to
FcεRI-antigen stimulation and that PPAR γ agonist attenuates BMMCs
in vitro differentiation after systemic treatment and
notably increases apoptosis. Consistent with our previous in
vivo studies (7,21), PPAR γ agonists might have clinical
potentials for allergic disorders, such as allergic rhinitis,
asthma, and atopic dermatitis.
Acknowledgements
We would like to thank Ying Lu and Bing Li
(Translational medicine research center, Shanghai East Hospital,
Tongji University School of Medicine) for flow cytometry technical
support and cell biology experiments.
The present study was supported by National Natural
Science Foundation of China (W.W., 81300809) and Key Disciplines
Group Construction Project of Pudong Health Bureau of Shanghai
(PWZxq2014-09).
Glossary
Abbreviations
Abbreviations:
|
BMMCs
|
bone marrow-derived mast cells
|
|
IL-3
|
interleukin-3
|
|
PPAR
|
peroxisome proliferator-activated
receptor
|
|
PIO
|
pioglitazone
|
|
SCF
|
stem cell factor
|
References
|
1
|
Kersten S, Desvergne B and Wahli W: Roles
of PPARs in health and disease. Nature. 405:421–424. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tachibana M, Wada K, Katayama K, Kamisaki
Y, Maeyama K, Kadowaki T, Blumberg RS and Nakajima A: Activation of
peroxisome proliferator-activated receptor gamma suppresses mast
cell maturation involved in allergic diseases. Allergy.
63:1136–1147. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao Y, Huang Y, He J, Li C, Deng W, Ran X
and Wang D: Rosiglitazone, a peroxisome proliferator-activated
receptor-γ agonist, attenuates airway inflammation by inhibiting
the proliferation of effector T cells in a murine model of
neutrophilic asthma. Immunol Lett. 157:9–15. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marcone S, Haughton K, Simpson PJ, Belton
O and Fitzgerald DJ: Milk-derived bioactive peptides inhibit human
endothelial-monocyte interactions via PPAR-γ dependent regulation
of NF-κB. J Inflamm (Lond). 12:12015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Housley WJ, Adams CO, Vang AG, Brocke S,
Nichols FC, LaCombe M, Rajan TV and Clark RB: Peroxisome
proliferator-activated receptor gamma is required for
CD4+ T cell-mediated lymphopenia-associated
autoimmunity. J Immunol. 187:4161–4169. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim SH, Hong JH and Lee YC: Ursolic acid,
a potential PPARγ agonist, suppresses ovalbumin-induced airway
inflammation and Penh by down-regulating IL-5, IL-13 and IL-17 in a
mouse model of allergic asthma. Eur J Pharmacol. 701:131–143. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang W, Zhu Z, Zhu B and Ma Z: Peroxisome
proliferator-activated receptor-gamma agonist induces regulatory T
cells in a murine model of allergic rhinitis. Otolaryngol Head Neck
Surg. 144:506–513. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meng Y, Chen C, Tian C, Du J and Li HH:
Angiotensin II-induced Egr-1 expression is suppressed by peroxisome
proliferator-activated receptor-γ ligand 15d-PGJ2 in macrophages.
Cell Physiol Biochem. 35:689–698. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maeyama K, Emi M and Tachibana M: Nuclear
receptors as targets for drug development: Peroxisome
proliferator-activated receptor gamma in mast cells: Its roles in
proliferation and differentiation. J Pharmacol Sci. 97:190–194.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sugiyama H, Nonaka T, Kishimoto T,
Komoriya K, Tsuji K and Nakahata T: Peroxisome
proliferator-activated receptors are expressed in mouse bone
marrow-derived mast cells. FEBS Lett. 467:259–262. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Spencer M, Yang L, Adu A, Finlin BS, Zhu
B, Shipp LR, Rasouli N, Peterson CA and Kern PA: Pioglitazone
treatment reduces adipose tissue inflammation through reduction of
mast cell and macrophage number and by improving vascularity. PLoS
One. 9:e1021902014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sly LM, Kalesnikoff J, Lam V, Wong D, Song
C, Omeis S, Chan K, Lee CW, Siraganian RP, Rivera J and Krystal G:
IgE-induced mast cell survival requires the prolonged generation of
reactive oxygen species. J Immunol. 181:3850–3860. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Okayama Y and Kawakami T: Development,
migration, and survival of mast cells. Immunol Res. 34:97–115.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma P, Mali RS, Munugalavadla V, Krishnan
S, Ramdas B, Sims E, Martin H, Ghosh J, Li S, Chan RJ, et al: The
PI3K pathway drives the maturation of mast cells via microphthalmia
transcription factor. Blood. 118:3459–3469. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wada K, Nakajima A, Katayama K, Kudo C,
Shibuya A, Kubota N, Terauchi Y, Tachibana M, Miyoshi H, Kamisaki
Y, et al: Peroxisome proliferator-activated receptor gamma-mediated
regulation of neural stem cell proliferation and differentiation. J
Biol Chem. 281:12673–12681. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suzuki K and Verma IM: Phosphorylation of
SNAP-23 by IkappaB kinase 2 regulates mast cell degranulation.
Cell. 134:485–495. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kashyap M, Bailey DP, Gomez G, Rivera J,
Huff TF and Ryan JJ: TGF-beta1 inhibits late-stage mast cell
maturation. Exp Hematol. 33:1281–1291. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lantz CS and Huff TF: Murine
KIT+ lineage- bone marrow progenitors express Fc
gamma-RII but do not express Fc epsilon-RI until mast cell granule
formation. J Immunol. 154:355–362. 1995.PubMed/NCBI
|
|
19
|
Cui Y, Dahlin JS, Feinstein R, Bankova LG,
Xing W, Shin K, Gurish MF and Hallgren J: Mouse mast cell
protease-6 and MHC are involved in the development of experimental
asthma. J Immunol. 193:4783–4789. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vangansewinkel T, Geurts N, Quanten K,
Nelissen S, Lemmens S, Geboes L, Dooley D, Vidal PM, Pejler G and
Hendrix S: Mast cells promote scar remodeling and functional
recovery after spinal cord injury via mouse mast cell protease 6.
FASEB J. 30:2040–2057. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang W, Zhu Z, Zhu B and Ma Z:
Pioglitazone attenuates allergic inflammation and induces
production of regulatory T lymphocytes. Am J Rhinol Allergy.
24:454–458. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gurish MF and Austen KF: Developmental
origin and functional specialization of mast cell subsets.
Immunity. 37:25–33. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rao KN and Brown MA: Mast cells:
Multifaceted immune cells with diverse roles in health and disease.
Ann N Y Acad Sci. 1143:83–104. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen CC, Grimbaldeston MA, Tsai M,
Weissman IL and Galli SJ: Identification of mast cell progenitors
in adult mice. Proc Natl Acad Sci USA. 102:11408–11413. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kitamura Y, Oboki K and Ito A: Molecular
mechanisms of mast cell development. Immunol Allergy Clin North Am.
26:387–405; v. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Saito H: Culture of human mast cells from
hemopoietic progenitors. Methods Mol Biol. 315:113–122.
2006.PubMed/NCBI
|
|
27
|
Succar J, Douaiher J, Lancerotto L, Li Q,
Yamaguchi R, Younan G, Pejler G and Orgill DP: The role of mouse
mast cell proteases in the proliferative phase of wound healing in
microdeformational wound therapy. Plast Reconstr Surg. 134:459–467.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Metcalfe DD, Baram D and Mekori YA: Mast
cells. Physiol Rev. 77:1033–1079. 1997.PubMed/NCBI
|
|
29
|
Nakazawa S, Sakanaka M, Furuta K,
Natsuhara M, Takano H, Tsuchiya S, Okuno Y, Ohtsu H, Nishibori M,
Thurmond RL, et al: Histamine synthesis is required for granule
maturation in murine mast cells. Eur J Immunol. 44:204–214. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hirai S, Ohyane C, Kim YI, Lin S, Goto T,
Takahashi N, Kim CS, Kang J, Yu R and Kawada T: Involvement of mast
cells in adipose tissue fibrosis. Am J Physiol Endocrinol Metab.
306:E247–E255. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu ZQ, Song JP, Liu X, Jiang J, Chen X,
Yang L, Hu T, Zheng PY, Liu ZG and Yang PC: Mast cell-derived
serine proteinase regulates T helper 2 polarization. Sci Rep.
4:46492014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
McNeil HP, Shin K, Campbell IK, Wicks IP,
Adachi R, Lee DM and Stevens RL: The mouse mast cell-restricted
tetramer-forming tryptases mouse mast cell protease 6 and mouse
mast cell protease 7 are critical mediators in inflammatory
arthritis. Arthritis Rheum. 58:2338–2346. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shin K, Nigrovic PA, Crish J, Boilard E,
McNeil HP, Larabee KS, Adachi R, Gurish MF, Gobezie R, Stevens RL
and Lee DM: Mast cells contribute to autoimmune inflammatory
arthritis via their tryptase/heparin complexes. J Immunol.
182:647–656. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hamilton MJ, Sinnamon MJ, Lyng GD,
Glickman JN, Wang X, Xing W, Krilis SA, Blumberg RS, Adachi R, Lee
DM and Stevens RL: Essential role for mast cell tryptase in acute
experimental colitis. Proc Natl Acad Sci USA. 108:290–295. 2011.
View Article : Google Scholar : PubMed/NCBI
|