Introduction
Lung cancer is a serious public health problem and
remains the most common cause of cancer-related mortality (1). Non-small cell lung cancer (NSCLC)
accounts for approximately 80% of all lung cancers (2). Although many advances have achieved
in cancer biology, such as the development of specific therapies
for distinct subtypes of NSCLC, appropriate treatment options for
the majority of patients are unsatisfactory (3). To identify new therapeutic targets
and options, it is of great necessity and importance to have a
better understanding of the pathogenesis of NSCLC.
Accumulating evidence suggests that, in many types
of malignancies, inflammatory signals could be regulated by
cytokines and chemokines, and thus exerting vital effects on tumor
progression including cancer cell proliferation, survival,
metastasis and angiogenesis (4).
The nine members of the interferon regulatory factor (IRF) family
are transcription factors that play diverse roles in innate and
adaptive immune responses, cell growth regulation, cell apoptosis,
and hematopoietic development (5–7).
Particularly, IRF4 is a diagnostic and prognostic
marker for various hematological malignancies (8). IRF4 is lymphocyte specific and is
overexpressed in Epstein-Barr virus transformed cells, multiple
myeloma, and human T cell leukemia virus 1 (HTLV1)-infected cell
lines and associated adult T cell lymphoma/leukemia (9–12).
There are several reports suggest a possible role of IRF4 in the
pathogenesis of chronic myeloid leukemia (13,14).
Besides, IRF4 upregulation has been proved to induce the growth of
lymphomas or multiple myeloma (10,15).
Yang et al (16) reported
that IRF 4 binding protein is a novel p53 target gene and
suppresses cisplatin-induced apoptosis of breast cancer cells. In a
previous study by Chen et al (17), IRF4 was shown to predict poorer
survival of NSCLC patients. Based on these investigations, it
raises the possibility that IRF4 may be involved in NSCLC tumor
progression, however, the underlying mechanism was not clear yet.
In the present study, we investigated whether IRF4 could exert
effects on human NSCLC and to explore the underlying mechanism in
the in vitro experiment. IRF4 expression was upregulated in
NSCLC tissues as compared to the corresponding adjacent non-tumor
tissue. In vitro experiments showed that IRF4 knockdown by
shRNA significantly reduced cell proliferation and colony number of
NSCLC cells; whereas IRF4 overexpression showed the absolutely
opposite results. Further investigations were performed to explore
the underlying mechanism in human NSCLC cells. The present study
suggests that IRF4 may be a new potential target for NSCLC
treatment.
Materials and methods
Human samples
Paired NSCLC (34 females and 20 males; with an
average age of 54; 22 patients with adenocarcinoma and 32 patients
with squamous cell carcinoma) and non-tumor adjacent lung tissues
(more than 5 cm from the edge of tumor) were obtained, with
informed consent, from 54 patients who underwent primary surgical
resection of NSCLC at the Second Affiliated Hospital of Soochow
University (SuZhou, China). Among them, 22 patients had negative
lymph nodes metastasis and 32 patients had positive lymph nodes
metastasis. 19 patients were at the I–II TNM stages and 35 were at
the III–IV TNM stages. None of the patients had received
preoperative radiotherapy or chemotherapy and chronic obstructive
pulmonary disease was excluded in the present study. Tissues were
obtained and frozen immediately with liquid nitrogen and stored in
a freezer at −70°C. The present study was approved by the Ethics
Committee of the Second Affiliated Hospital of Soochow University.
Written informed consent was obtained from all the
participants.
Quantitative real-time PCR
Total RNA was extracted from paired NSCLC and
non-tumor adjacent lung tissues with Trizol reagent (Invitrogen,
Carlsbad, CA, USA) according to the manufacturer's instructions.
First-strand cDNA was synthesized from total RNA using Super Script
II first-strand synthesis system (Invitrogen). Real-time PCR was
performed using an Applied Biosystems 7300 Sequence Detection
system (Applied Biosystems, Foster City, CA, USA). The primers for
IRF4: forward, 5′-CTACACCATGACAACGCCTTACC-3′ and reverse,
5′-GGCTGATCCGGGACGTAGT-3′. Notch1: forward,
5′-CTTAGATGTGCTGAGCGCGTCAATGTGTC-3′ and reverse,
5′-GCGCGATCCTTGATAACCTGCGGAT-3′. Notch 2: forward,
5′-CATAGAATGATTAGCAGAGAG-3′ and reverse,
5′-CAACATCAGAGCTAGCAAGAG-3′. GAPGH, forward
5′-GGTGGAGGTCGGGAGTCAACGGA-3′, reverse
5′-GAGGGATCTCGCTCCTGGAGGA-3′. The comparative threshold cycle (Ct)
method was used to analyze the results.
Western blot analysis
For western blot analysis, cell lysates were
prepared from cell lines with RIPA lysis buffer kit (Santa Cruz
Biotechnology, Santa Cruz, CA), and the protein concentrations were
quantified using a Bio-Rad protein assay (Bio-Rad, Hercules, CA).
Whole-cell proteins (30 µg) were separated on 8% sodium dodecyl
sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to polyvinylidene difluoride membranes (Amersham Corp.,
Arlington Heights, IL). The membranes were incubated with primary
antibodies anti-IRF4 (1:500, cat. sc-6059; Santa Cruz
Biotechnology), anti-Notch1 (1:100, cat. Val1744, Cell Signaling,
Danvers, MA), anti-Notch2 (1:500, cat. D67C8; Cell Signaling),
anti-AKT (1:500, cat. 9272; Cell Signaling), anti-AKT
phosphorylation (1:500, cat. 9271; Cell Signaling) or anti-GAPDH
(1:1,000, cat. 2111; Cell Signaling) overnight at 4°C. The
horseradish peroxidase-conjugated secondary antibodies (1:1,000,
cat. A50-106P; Beijing Zhongshan Golden Bridge Biotechnology Co.,
Ltd., Beijing, China;) were subsequently used at room temperature
for 1 h. Signals were detected using enhanced chemiluminescence kit
(Wuhan Booute Biotechnology Co., Ltd, Wuhan, China; cat. no.
orb90504) and exposed to Kodak X-OMAT film (Eastman Kodak,
Rochester, NY, USA).
Cell lines and cell culture
Human lung adenocarcinoma cell line (A549) and lung
squamous cell carcinoma cell line (RERF-LC-AI, LC-AI) were obtained
from Shanghai Cell Bank (Chinese Academy of Sciences, China) and
maintained in −80°C freezer or cultured in RPMI (Roswell Park
Memorial Institute) 1640 medium containing 10% fetal bovine serum
(FBS) before experiments. Cells were maintained in 5%
CO2 incubator at 37°C and used for experiments in the
exponential phase of their growth. To detect the effect of Notch on
NSCLC cells, A549 and LC-AI were treated with 20 µM MK-0752, a
Notch pathway inhibitor (18), for
48 h.
Lentivirus packaging
For IRF4 knockdown, the lentivirus expressing short
hairpin RNA (shRNA) targeting the sequence of IRF4
(5′-GCCGTACAAAGTTCAGGATCC-3′) and the control sequence
(5′-TTCTCCGAACGTGTCACGT-3′) was purchased from Genechem Co. Ltd
(Shanghai, China). Lentivirus expressing scrambled shRNA was used
as a negative control (sh-Ctrl; Genechem Co. Ltd.). For IRF4
overexpression, the targeted IRF4 gene was amplified in human
sample by PCR with the following primers: forward:
5′-CACCATGACAACGCCTTACC-3′ and reverse 5′-CATTTTCACAAGCTGGGCCT-3′.
The PCR product containing PmeI and BstBI site introduced by primer
was digested, and then cloned into lentivirus vector to construct a
lentiviral vector carrying IRF4. The empty lentivirus vector was
used as a negative control (Ctrl). The shRNA-expressing,
overexpressing lentivirus and the corresponding controls were
transfected into 293 T cells, which are always used in producing
lentiviruses (19), together with
the lentivirus helper plasmids to generate respective lentivirus.
After transfection, the lentivirus supernatant was collected to
transfect A549 and LC-AI cells. Infectious lentivirus was harvested
48 h post-transfection, centrifuged to remove cell debris, and then
filtered through 0.45 µm cellulose acetate filters.
Cell proliferation assay
Cells were seeded into a 96-well plate
(1×104 cells/well) and cultured in 200 µl RPMI-1640. At
4 h before terminating culture, 20 µl MTT (5 mg/ml) was added into
each well. After cell culture, the supernatant was discarded and
100 µl dimethylsulfoxide (DMSO) was added into each well. After the
formazan granules were completely dissolved, a value of each well
was measured at 490 nm, and the corresponding cell proliferation
number was calculated.
Colony formation assay
NSCLC cells were suspended in 1.5 ml complete medium
supplemented with 0.45% low melting point agarose. The cells were
placed in 35 mm tissue culture plates containing 1.5 ml complete
medium and agarose (0.75%) on the bottom layer. The plates were
incubated at 37°C with 5% CO2 for 2 weeks. Cell colonies
were stained with 0.005% crystal violet and analyzed using a
microscope.
Statistical analysis
Data were presented as means ± SEM. The statistical
analysis was performed with SPSS 10.0 (SPSS, Inc., Chicago, IL,
USA). For two groups, Student's t test was performed to analysis
the data, and one-way ANOVA analysis was applied when three or more
groups exist. The associations between Notch1, Notch2 mRNA
expression levels and IRF4 mRNA level in NSCLC tissues were tested
with the linear regression analysis. P<0.05 was considered
statistically significant.
Results
IRF4 is upregulated in human NSCLC
tissues
To investigate the role of IRF4 in NSCLC
development, we first analyzed the expression of IRF4 in NSCLC
tissues (n=54, 22 cases with adenocarcinoma and 32 cases with
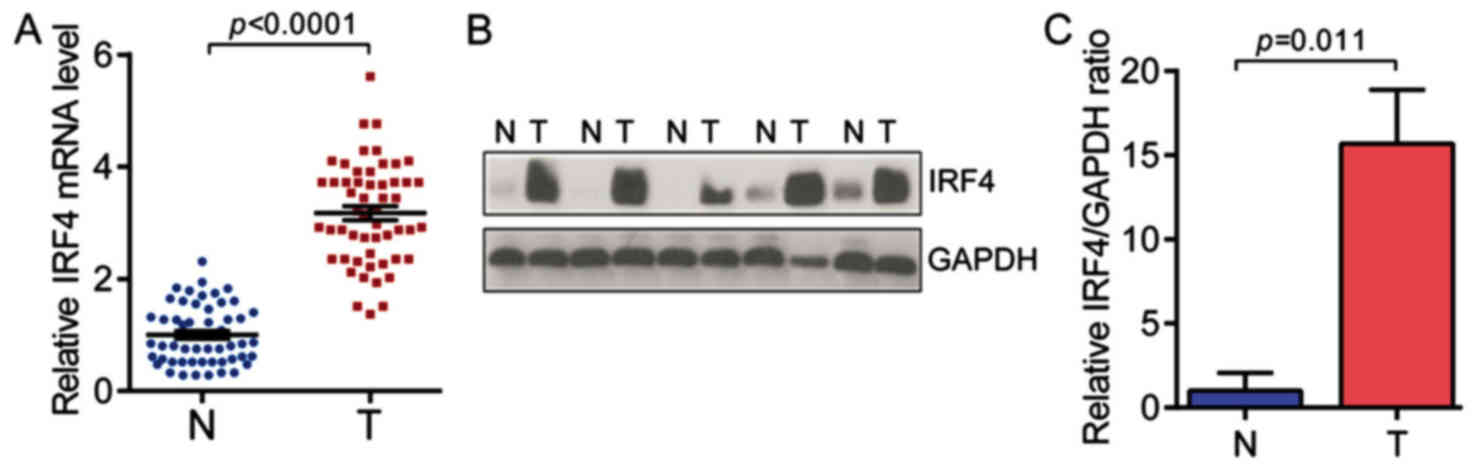
squamous cell carcinoma). As shown in Fig. 1A, the expression levels of IRF4
mRNA were significantly highe rin NSCLC tissues as compared with
non-tumor adjacent normal tissues (n=54 for each group). Moreover,
the expression levels of IRF4 protein were significantly higher in
NSCLC tissues (Fig. 1B and C).
These findings indicate the potential role of IRF4 in NSCLC
development and progression.
IRF4 level is positively correlated
with the expression of Notch in NSCLC tissues
Then, we want to further explore the underlying
mechanism of the tumorigenic role of IRF4 in NSCLC. Notch, firstly
identified in human T-cell neoplasia, is a potent regulator for
cellular differentiation, development, proliferation and survival
(20–22). Besides, previous study indicated
that IRF4 acts as a critical regulator of Notch signaling during
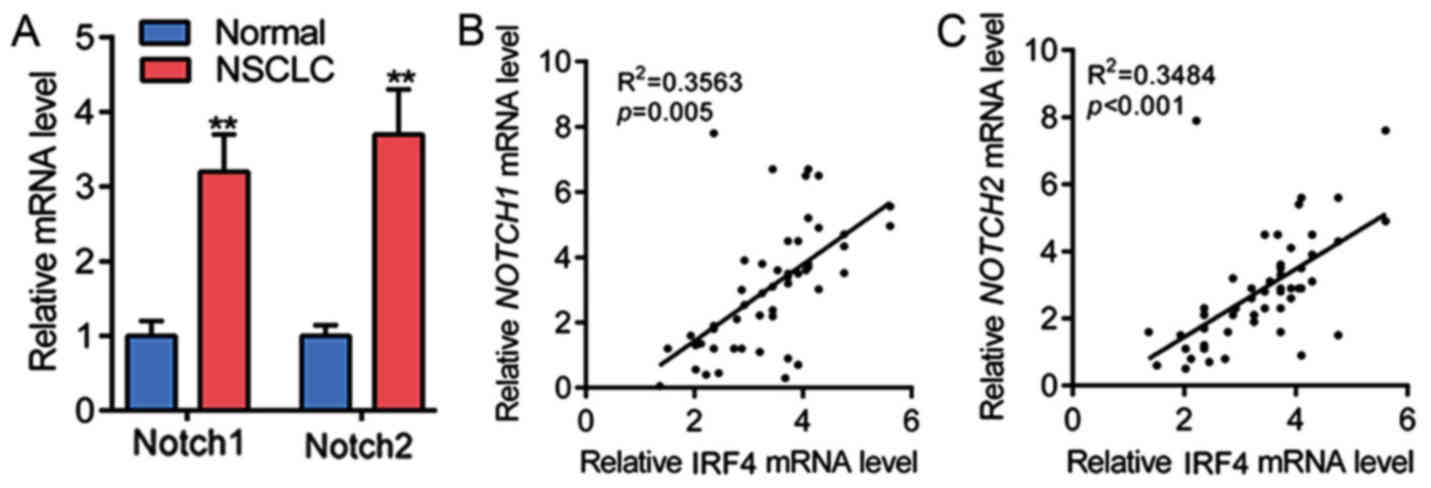
chronic lymphocytic leukemia (CLL) development (23). Therefore, the mRNA levels of Notch1
and Notch2 in NSCLC tissues were determined, as shown in Fig. 2A, Notch1 and Notch2 mRNA expression
were significantly upregulated in NSCLC tissues as compared with
non-tumor adjacent normal tissues. Subsequently, linear regression
analysis showed that IRF4 mRNA expression was positively correlated
with the mRNA level of Notch1 in 54 cases of NSCLC tissues
(Fig. 2B). The linear regression
analysis also showed that IRF4 mRNA expression was positively
correlated with the mRNA level of Notch2 in NSCLC tissues in 54
cases of NSCLC tissues (Fig. 2C).
These results showed that IRF4 level was positively correlated with
the expression of Notch in NSCLC tissues.
IRF4 promotes cell growth of human
NSCLC cells
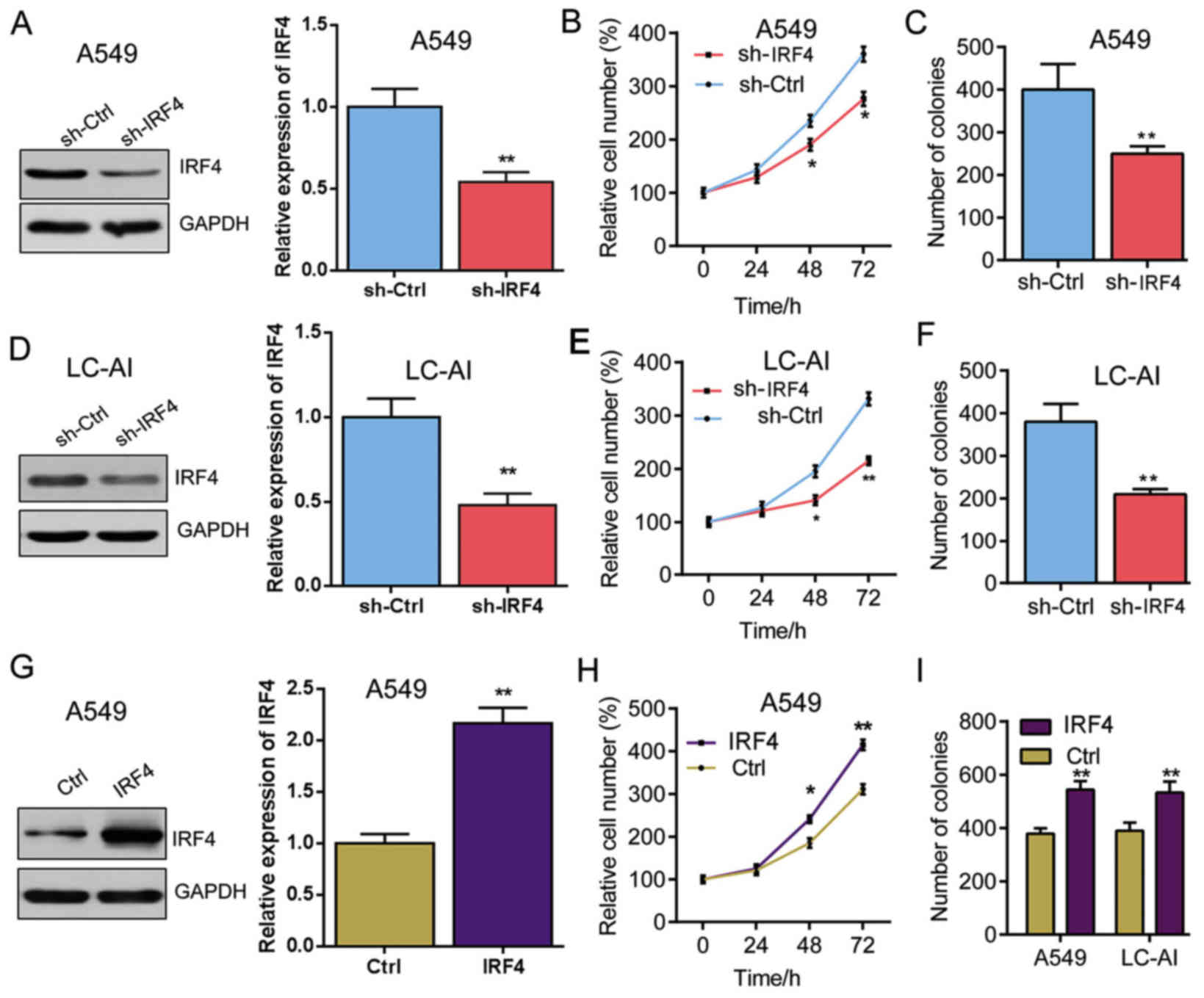
To understand the tumorigenic role of IRF4, we
knock-downed the expression of IRF4 with lentivirus expressing
sh-IRF4 in A549 and LC-AI cells, respectively. The IRF4 expression
was significantly decreased by IRF4 knockdown in A549, which is
determined by western blot analysis (Fig. 3A). As shown in Fig. 3B, cell proliferation was
significantly suppressed in A549 that with IRF4 knockdown in a
time-dependent manner. And IRF4 knockdown also markedly reduced the
colony number in A549 cells (Fig.
3C). Similarly, IRF4 expression was also significantly
inhibited in the LC-AI cells (Fig.
3D) and the cell proliferation rate and colony number of LC-AI
cells were markedly reduced by IRF4 knockdown (Fig. 3E and F).
Additionally, IRF4-overexpressing lentivirus (IRF4)
was also generated and infectedA549 cells, which was confirmed by
western blotting analysis (Fig.
3G). In contrast to the results of IRF4 knockdown, IRF4
overexpression significantly promoted the proliferation and
increased colony number of A549 cells (Fig. 3H and I). Besides, similar results
were found in LC-AI cells (Fig.
3I). These results indicate that IRF4 promotes NSCLC cell
proliferation and increases colony number.
IRF4 activates notch-Akt signaling in
NSCLC cells
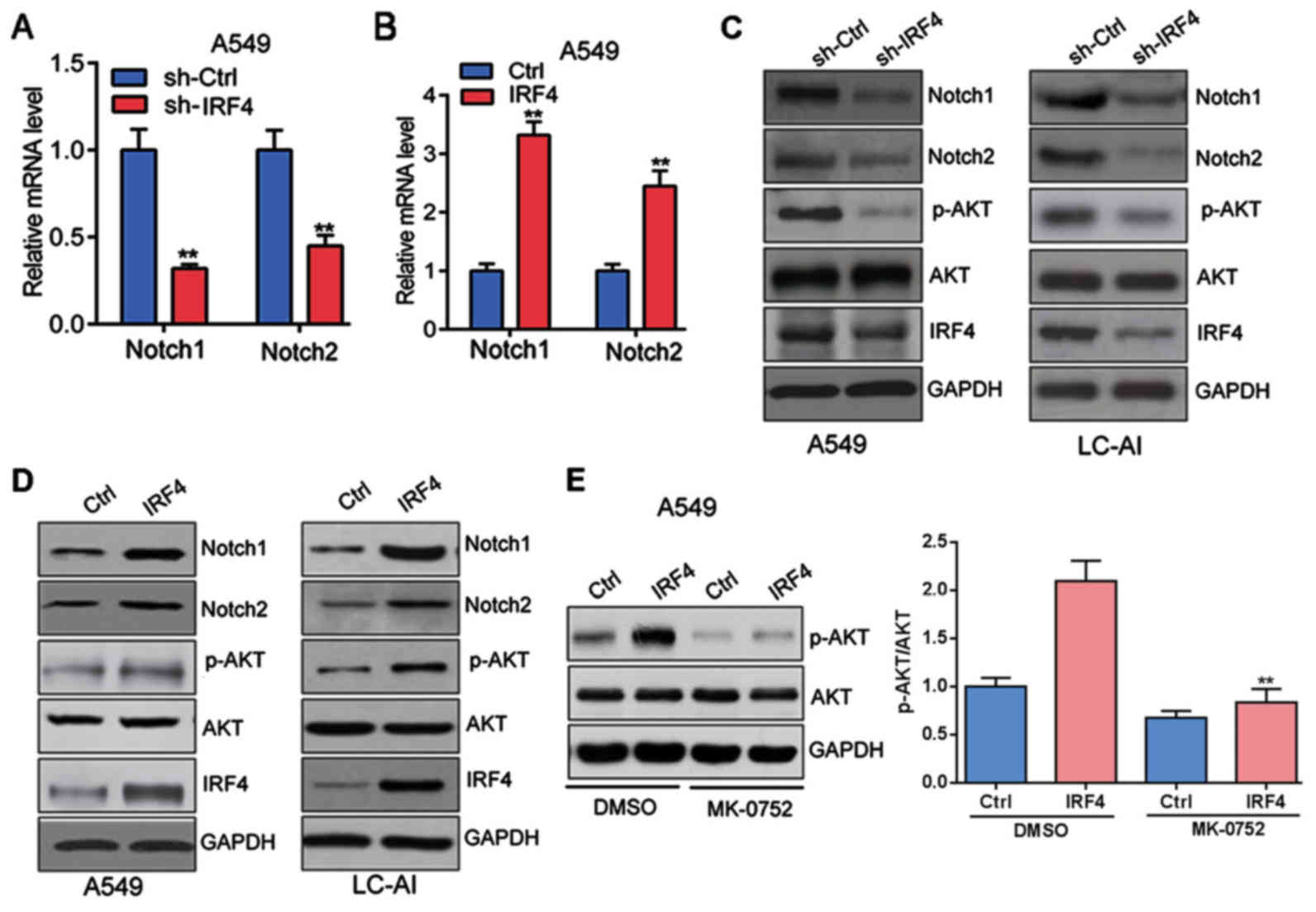
Furthermore, as shown in Fig. 4A and B, IRF4 knockdown
significantly decreased both the mRNA expression of Notch1 and
Notch2 in A549 cells; whereas IRF4 overexpression markedly
increased the mRNA expression of Notch1 and Notch2 in A549
cells.
There is growing evidence that Notch regulates the
AKT pathway in several normal and cancer cell types (24), we then investigated whether
Notch/AKT pathway was activated by IRF4. Indeed, IRF4 knockdown
significantly suppressed the protein expression of Notch1, Notch2
and phosphorylation of AKT (p-AKT) in A549 and LC-AI cells
(Fig. 4C); whereas IRF4
overexpression markedly facilitated the protein expressionof
Notch1, Notch2 and p-AKT in A549 and LC-AI cells (Fig. 4D). Furthermore, A549 cells with
IRF4 overexpression (IRF4) were treated with Notch pathway
inhibitor MK-0752 (20 µM) or DMSO for 48 h. Western blotting was
performed and the results showed that MK-0752 could significantly
reverse the effect of IRF4 overexpressionon activation of Akt
signaling in A549 cells (Fig. 4E).
Totally, those investigations indicated that IRF4 activates
Notch-Akt signaling in NSCLC cells.
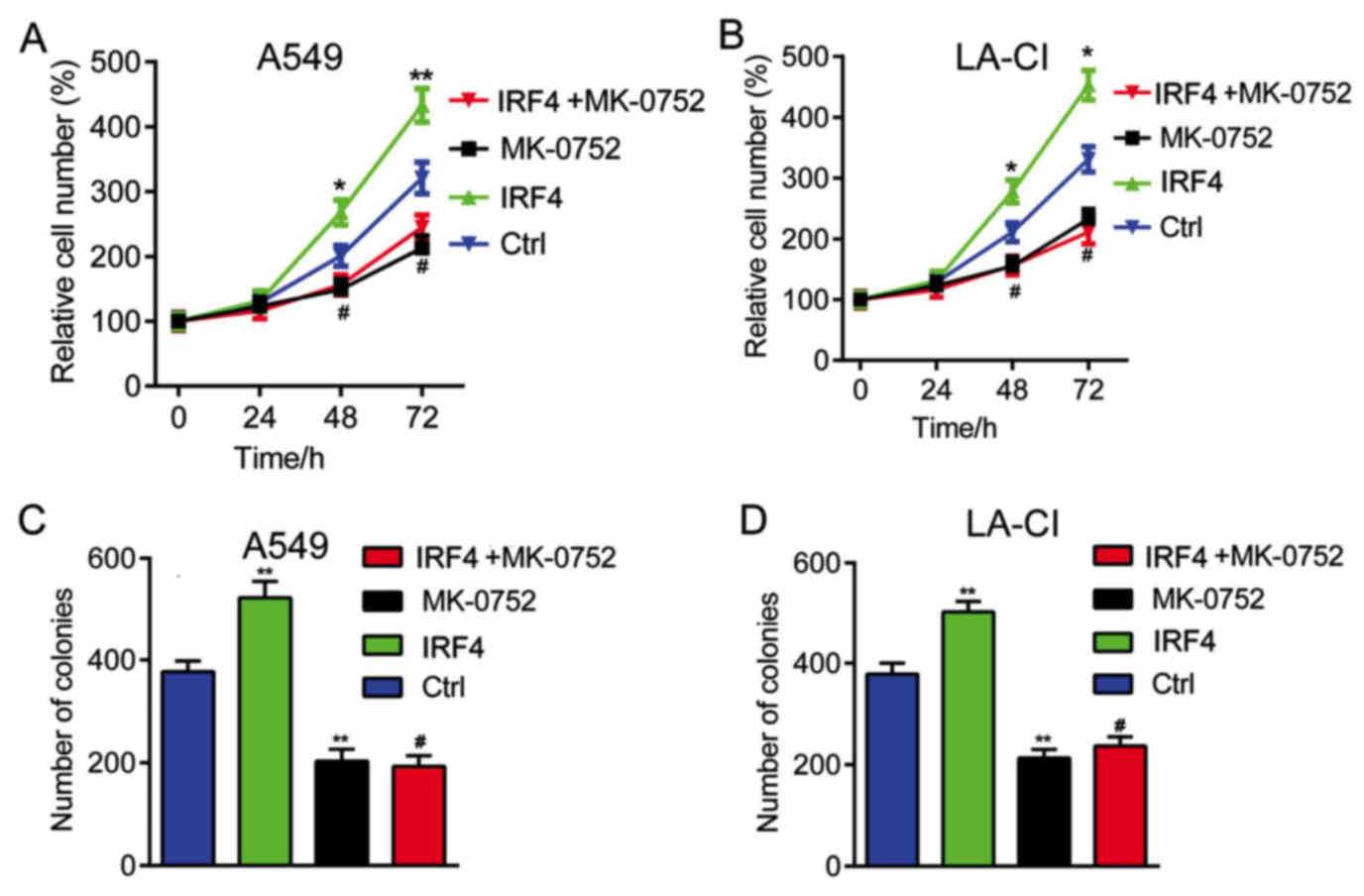
Notch inhibition reverses the effects
of IRF4 overexpression on cell growth
Finally, we explored whether Notch-Akt pathway was
involved in the effects of IRF4 on human NSCLC cell growth. As
shown in Fig. 5, IRF4
overexpression (IRF4) could significantly increased cell
proliferation and colony number. However, when treated with Notch
pathway inhibitor MK-0752 (IRF4+MK-0752), the enhancement of cell
proliferation and colony number induced by IRF4 overexpression were
significantlyreversed. It seems that Notch inhibition significantly
reverses the effects of IRF4 overexpression on human NSCLC cell
growth.
Discussion
In the present study, the function of IRF4 in human
NSCLC was investigated. IRF4 mRNA and protein were found highly
expressed in NSCLC tissues as compared to the corresponding
adjacent non-tumor tissues, indicating its potential role in NSCLC
development and progression. The cell proliferation rate and colony
number of A549 and LC-AI were significantly declined when IRF4
expression was knocked down. However, the cell proliferation rate
and colony number of A549 and LC-AI cells were markedly increased
when IRF4 expression was overexpressed. These investigations
validated the important role of IRF4 in NSCLC cell growth. This is
the first evidence demonstrating the oncogenic role of IRF4 in
NSCLC. IRF4 is overexpressed in many hematological malignancies,
serves as a diagnostic and prognostic marker of hematological
malignancies and is a promising therapeutic target for treatment of
hematological malignancies (25).
In a previous study by Chen et al (17), IRF4 was shown to predict poorer
survival of NSCLC patients. To some extent, te present study is
consistent with our study. By contrast, Wu et al (26) demonstrated that IRF4 was a
protective prognostic factor in NSCLC patients. These contradictory
data may suggest that IRF4 may reflect tumor-infiltrating
lymphocyte activity in tissue section.
Besides, in the present study, Notch1 and Notch2
expression were significantly upregulated in NSCLC tissues as
compared to corresponding adjacent non-tumor tissues, also
indicating their potential role in NSCLC development and
progression. Notch1 and Notch2 expression were found positively
correlated with IRF4 level in NSCLC tissues. The expression of
Notch1, Notch2 and p-AKT were found regulated by IRF4 in NSCLC
cells. In the cells with IRF4 overexpressed that treated with
MK-0752, it showed the significant decreased cell proliferation
rate and colony number as compared with cells treated with DMSO.
Mechanically, our study suggests that the oncogenic role of IRF4 in
NSCLC was, at least partially, through regulating Notch expression
and activating Notch-Akt signaling pathway. Previous study
indicated that low level of IRF4 is a common feature of CLL, and
the deregulated IRF4-Notch axis may represent a major pathway in
the molecular pathogenesis of CLL (23). It suggests the positive relation
between IRF4 and Notch signaling pathway, to some extent, this is
consistent with our study. Besides, Previous study also identified
Nedd4 as a key IRF4 target gene involved in impeding the responses
of CLL cells and their precursors to Notch signaling (23), whether Nedd4 acts as a linkage
between IRF4 and Notch signaling in our study remain further
exploration.
It is well-known that Notch receptors (Notch1-4) are
critical for tumorigenesis (27).
Upon binding to its ligands, Notch receptors are proteolytically
processed bymetal loproteases and the g-secretase complex. This
process releases the intracellular domain of Notch, which
thentranslocates into the nucleus to modulate expression of its
target genes (28). Our study is
consistent with a serious of previous studies that indicating the
tumorgenic role of Notch1 (29–31).
Particularly, a previous work has shown that Notch1 signaling
promotes tumorigenicity and survival in NSCLC cells in vitro
(32). The present study also
showed that Notch2 mRNA was upregulated in NSCLC tissues,
consistently, the transcription factors Notch2 was upregulated in
invasive cancer cells in all 11 minimally invasive adenocarcinomas
(33). In embryonal brain tumors,
Notch2 also functions as an oncogene (34). However, a study reported that
Notch2 mediates differentiation and has tumor suppressor functions
during lung carcinogenesis (3).
Notch2 also functions as a tumor suppressor in bronchial epithelial
cells modulating tumor initiation and progression (35).
There is growing evidence that Notch regulates the
AKT pathway in several normal and cancer cell types (24). The present study showed IRF4
upregulated the expression of Notch-1, Notch-2 and p-Akt, it seems
that Notch-1 and Notch-2 have consistently regulative effects on
p-Akt levels. Differently, aprevious study indicated that Notch-1
led to Akt phosphorylation and promoted cell survival, whereas
Notch-2 signaling led to Akt dephosporylation and suppressed cell
survival of malignant mesothelioma cells (35).
In summary, our results indicate that IRF4 acts as a
tumor promoter in human NSCLC, at least partially, through
activating Notch-Akt signaling pathway.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yuan X, Wu H, Han N, Xu H, Chu Q, Yu S,
Chen Y and Wu K: Notch signaling and EMT in non-small cell lung
cancer: Biological significance and therapeutic application. J
Hematol Oncol. 7:872014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baumgart A, Mazur P, Anton M, Rudelius M,
Schwamborn K, Feuchtinger A, Behnke K, Walch A, Braren R, Peschel
C, et al: Opposing role of Notch1 and Notch2 in a Kras(G12D)-driven
murine non-small cell lung cancer model. Oncogene. 34:578–588.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coffelt SB, Hughes R and Lewis CE:
Tumor-associated macrophages: Effectors of angiogenesis and tumor
progression. Biochimica et Biophysica Acta (BBA)-Reviews on Cancer.
1796:11–18. 2009. View Article : Google Scholar
|
|
5
|
Honda K and Taniguchi T: IRFs: Master
regulators of signalling by Toll-like receptors and cytosolic
pattern-recognition receptors. Nat Rev Immunol. 6:644–658. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mamane Y, Heylbroeck C, Génin P, Algarté
M, Servant MJ, LePage C, DeLuca C, Kwon H, Lin R and Hiscott J:
Interferon regulatory factors: The next generation. Gene. 237:1–14.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taniguchi T, Ogasawara K, Takaoka A and
Tanaka N: IRF family of transcription factors as regulators of host
defense. Annual Rev immunol. 19:623–655. 2001. View Article : Google Scholar
|
|
8
|
Wang L and Ning S: Interferon regulatory
factor 4 is activated through c-Src-mediated tyrosine
phosphorylation in virus-transformed cells. J Virol. 87:9672–9679.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Spender LC, Lucchesi W, Bodelon G,
Bilancio A, Karstegl CE, Asano T, Dittrich-Breiholz O, Kracht M,
Vanhaesebroeck B and Farrell PJ: Cell target genes of Epstein-Barr
virus transcription factor EBNA-2: Induction of the p55alpha
regulatory subunit of PI3-kinase and its role in survival of EREB2.
5 cells. J Gen Virol. 87:2859–2867. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iida S, Rao PH, Butler M, Corradini P,
Boccadoro M, Klein B, Chaganti RS and Dalla-Favera R: Deregulation
of MUM1/IRF4 by chromosomal translocation in multiple myeloma. Nat
Genet. 17:226–230. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mamane Y, Grandvaux N, Hernandez E, Sharma
S, Innocente SA, Lee JM, Azimi N, Lin R and Hiscott J: Repression
of IRF-4 target genes in human T cell leukemia virus-1 infection.
Oncogene. 21:6751–6765. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sharma S, Mamane Y, Grandvaux N, Bartlett
J, Petropoulos L, Lin R and Hiscott J: Activation and regulation of
interferon regulatory factor 4 in HTLV type 1-infected T
lymphocytes. AIDS Res Hum Retroviruses. 16:1613–1622. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ortmann CA, Burchert A, Hölzle K, Nitsche
A, Wittig B, Neubauer A and Schmidt M: Down-regulation of
interferon regulatory factor 4 gene expression in leukemic cells
due to hypermethylation of CpG motifs in the promoter region.
Nucleic Acids Res. 33:6895–6905. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schmidt M, Hochhaus A, König-Merediz SA,
Brendel C, Proba J, Hoppe GJ, Wittig B, Ehninger G, Hehlmann R and
Neubauer A: Expression of interferon regulatory factor 4 in chronic
myeloid leukemia: Correlation with response to interferon alfa
therapy. J Clin Oncol. 18:3331–3338. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tsuboi K, Iida S, Inagaki H, Kato M,
Hayami Y, Hanamura I, Miura K, Harada S, Kikuchi M, Komatsu H, et
al: MUM 1/IRF4 expression as a frequent event in mature lymphoid
malignancies. Leukemia. 14:449–456. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang M, Yuan F, Li P, Chen Z, Chen A, Li S
and Hu C: Interferon regulatory factor 4 binding protein is a novel
p53 target gene and suppresses cisplatin-induced apoptosis of
breast cancer cells. Mol Cancer. 11:542012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen HY, Yu SL, Chen CH, Chang GC, Chen
CY, Yuan A, Cheng CL, Wang CH, Terng HJ, Kao SF, et al: A five-gene
signature and clinical outcome in non-small-cell lung cancer. N
Engl J Med. 356:11–20. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Blackman SC, Podtelezhnikov A, Railkar RA,
Loboda A, Tanis K, Klappenbach JA, Watters J, Iannone R, Herman G
and Bergtrom DA: Abstract 26: Notch pathway inhibition with MK-0752
leads to dose- and time-dependent transcriptional alterations in
proliferation, PI3K, and Wnt pathway genes in plucked human hair
follicles. Cancer Res. 70:17–21. 2010. View Article : Google Scholar
|
|
19
|
Zhao J and Lever AM: Lentivirus-mediated
gene expression. Methods Mol Biol. 366:343–355. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Harper JA, Yuan JS, Tan JB, Visan I and
Guidos CJ: Notch signaling in development and disease. Clin Genet.
64:461–472. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kadesch T: Notch signaling: The demise of
elegant simplicity. Curr Opin Genet Dev. 14:506–512. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ellisen LW, Bird J, West DC, Soreng AL,
Reynolds TC, Smith SD and Sklar J: TAN-1, the human homolog of the
Drosophila notch gene, is broken by chromosomal translocations in T
lymphoblastic neoplasms. Cell. 66:649–661. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shukla V, Shukla A, Joshi SS and Lu R:
Interferon regulatory factor 4 attenuates Notch signaling to
suppress the development of chronic lymphocytic leukemia.
Oncotarget. 7:41081–41094. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu H, Bhaijee F, Ishaq N, Pepper DJ,
Backus K, Brown AS, Zhou X and Miele L: Correlation of Notch1, pAKT
and nuclear NF-κB expression in triple negative breast cancer. Am J
Cancer Res. 3:230–239. 2013.PubMed/NCBI
|
|
25
|
Gualco G, Weiss LM and Bacchi CE:
MUM1/IRF4: A Review. Appl Immunohistochem Mol Morphol. 18:301–310.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu YY, Hwang YT, Perng WC, Chian CF, Ho
CL, Lee SC, Chang H, Terng HJ and Chao TY: CPEB4 and IRF4
expression in peripheral mononuclear cells are potential prognostic
factors for advanced lung cancer. J Formos Med Assoc. 116:114–122.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bray SJ: Notch signalling: A simple
pathway becomes complex. Nat Rev Mol Cell Biol. 7:678–689. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Weijzen S, Rizzo P, Braid M, Vaishnav R,
Jonkheer SM, Zlobin A, Osborne BA, Gottipati S, Aster JC, Hahn WC,
et al: Activation of Notch-1 signaling maintains the neoplastic
phenotype in human Ras-transformed cells. Nat Med. 8:979–986. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Allen TD, Rodriguez EM, Jones KD and
Bishop JM: Activated Notch1 induces lung adenomas in mice and
cooperates with Myc in the generation of lung adenocarcinoma.
Cancer Res. 71:6010–6018. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Osanyingbemi-Obidi J, Dobromilskaya I,
Illei PB, Hann CL and Rudin CM: Notch signaling contributes to lung
cancer clonogenic capacity in vitro but may be circumvented in
tumorigenesis in vivo. Mol Cancer Res. 9:1746–1754. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Baumgart A, Seidl S, Vlachou P, Michel L,
Mitova N, Schatz N, Specht K, Koch I, Schuster T, Grundler R, et
al: ADAM17 regulates epidermal growth factor receptor expression
through the activation of Notch1 in non-small cell lung cancer.
Cancer Res. 70:5368–5378. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mimae T, Okada M, Hagiyama M, Miyata Y,
Tsutani Y, Inoue T, Murakami Y and Ito A: Upregulation of notch2
and six1 is associated with progression of early-stage lung
adenocarcinoma and a more aggressive phenotype at advanced stages.
Clin Cancer Res. 18:945–955. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fan X, Mikolaenko I, Elhassan I, Ni X,
Wang Y, Ball D, Brat DJ, Perry A and Eberhart CG: Notch1 and notch2
have opposite effects on embryonal brain tumor growth. Cancer Res.
64:7787–7793. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Graziani I, Eliasz S, De Marco MA, Chen Y,
Pass HI, De May RM, Strack PR, Miele L and Bocchetta M: Opposite
effects of Notch-1 and Notch-2 on mesothelioma cell survival under
hypoxia are exerted through the Akt pathway. Cancer Res.
68:9678–9685. 2008. View Article : Google Scholar : PubMed/NCBI
|