Introduction
Osteosarcoma most often affects the long bones, such
as those in the arms and legs, and has an annual incidence
worldwide of ~one to three cases per million (1,2). It
has a bimodal distribution with peaks in the second decade of life
and late adulthood, and arises primarily in pre-teens and teens,
and is the third most common cancer in adolescence, and the most
common primary bone tumor in children (3,4). It
has been reported that osteosarcoma has a higher incidence in males
(5.4 per million males/year) than in females (4.0 per million
females/year) (5). The typical
clinical presentation of osteosarcoma includes pathologic
fractures, swelling and onset of pain in the affected bone, where
the pain is sufficiently intense to wake the patient from sleep.
~15 to 20% of patients will have clinically detectable metastases.
The most common site of metastasis occurs in the lung and bone is
the second most common metastasis site (6).
Cancer deaths caused by bone and joint malignancy
account for 8.9% of all childhood and adolescent cancer deaths, and
the death rates of osteosarcoma have been declining by ~1.3% per
year. The overall 5-year survival rate for osteosarcoma is 68%, and
the older patients have the poorest survival; however, it is not
correlated with gender (5).
Depending on the cancer stage, osteosarcoma treatment includes
surgery and chemotherapy (7).
Complete surgical excision and chemotherapy regimens are associated
with survival outcome significantly. In addition, tumor staging,
tumor size, anatomic location and presence of metastases, as well
as local recurrence are important to ensure an optimum prognosis
(5).
Because osteosarcoma cells secrete various growth
factors that induce angiogenesis to supply nutrition for their own
growth, agents that inhibit angiogenesis are good candidates for
antitumor drugs (8–10). Chondromodulin-I (ChM-I), a 25-kDa
glycoprotein purified from bovine epiphyseal cartilage on the basis
of growth-promoting activity for chondrocytes, was recently
identified as an angiogenesis inhibitor (11,12).
Human ChM-I cDNA revealed that the mature protein consists of 120
amino acids and is coded as the C-terminal part of a larger
transmembrane precursor (13,14).
It is reported that ChM-I is a homologous inhibitor and is
predominantly expressed in the avascular region of cartilage
(15).
It is well known that ChM-I exerts
anti-vascularization action; therefore, it is considered to have
antitumor properties (16). Tsai
et al (17) indicated that
ChM-I exhibits vascular targeting activity that is associated with
the induction of DR5-mediated endothelial cell apoptosis through
p53 upregulation, which suggests its potential as an antivascular
and antitumor therapeutic agent. Hsu et al (18) reported that CHM-I is a potent
inducer of apoptosis, which serves a role in the anticancer
activity of CHM-I. However, it has remained unclear whether ChM-I
exerts any direct antitumor effect on osteosarcoma cells.
Therefore, the present study aimed to evaluate the direct effects
of ChM-I on osteosarcoma cells. The current study indicated that
ChM-I directly suppresses the proliferation of osteosarcoma cells
in vitro, and also inhibits osteosarcoma cell growth in
vivo.
Materials and methods
Animals and patients
All animal experimental procedures in this study
were approved by the Ethics Committee of the Second Military
Medical University (Shanghai, China).
Human articular cartilage used in this study was
used to harvest the ChM-I gene, and was obtained from the femoral
condyles of patients undergoing knee joint replacement surgery at
Shanghai Changzheng Hospital, Second Military Medical University.
Written informed consent was obtained from all participants prior
to the study. A total of 6 patients were recruited to the present
study (male, 65 years). After surgically removing cartilage from
the knee joint, the chondrocytes were harvested by enzymatic
digestion.
Cell culture
The Saos-2 human osteosarcoma cell line was
purchased from the Shanghai Biological Cell Bank, Chinese Academy
of Sciences (Shanghai, China). Cells were cultured for 1 week
(after 2 passages) in Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS) in a humidified
atmosphere containing 5% CO2 at 37°C.
Small interfering RNA (siRNA)
siRNA transfection was used to knock down ChM-I mRNA
expression in Saos-2 cells. The siRNA sequences used were:
5′-UGGAUUUAUCCUACAGAUGCA-3′; 5′-CAUCUGUAGGAUAAAUCCAUA-3′. siRNA
transfection was performed according to the protocol of a previous
study (19).
ChM-I overexpression
A PcDNA3.1(+)-ChM-I plasmid was constructed to
overexpress ChM-I, and a ChM-I cDNA expression vector
(pcDNA3.1(+)-ChM-I) was constructed by Shanghai GeneChem Co., Ltd.
(Shanghai, China). In brief, the ChM-I gene was generated by
polymerase chain reaction (PCR) amplification from cartilage
tissue, and the plasmid pcDNA3.1(+)-ChM-I was generated according
to the cDNA sequence from GenBank (National Institutes of Health,
Bethesda, MD, USA). The plasmid pcDNA3.1(+) was extracted with a
Qiagen extraction kit (Qiagen China Co., Ltd., Shanghai, China)
according to the manufacturer's protocol. The PCR product was
subcloned into the BamHI (Clontech Laboratories, Inc.,
Mountain View, CA, USA) and HindIII (Clontech Laboratories,
Inc.) sites of pcDNA3.1 plasmid by T4 ligase (Clontech
Laboratories, Inc.). The recombinant plasmid was transfected into
293T cells using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.). Adenoviruses were plaque-purified and
propagated in 293T cells. The viral lysates were purified and
concentrated through two cycles of CsCl2 (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) step gradients, and the reconstructed
adenovirus was divided into several aliquots and stored at a titer
of 1.0×109 plaque-forming units/µl at 80°C according to
a previous study (20).
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR was used to analyze ChM-I mRNA expression
levels in the ChM-I overexpression and siRNA groups, according to a
previous study (20). RNA
extraction buffer (catalog no. Z3051; Promega Corporation, Madison,
WI, USA), the miScript II RT kit (catalog no. 218160; Qiagen China
Co., Ltd.) and the Quntititect™ SYBR-Green PCR Reagent kit (Qiagen
China Co., Ltd.) were used. The primer sequences for ChM-I were as
follows: Forward, 3′-ATCAGCAGGAAGGGGAAAGC-5′ and reverse,
3′-TGGTCCCATCAGCATCAACC-5′, product size 308 bp. Primer sequences
for β-actin were: Forward, 3′-3′-TCCATCATGAAGTGTGACGT-5′ and
reverse, 3′-CTCAGGAGGAGCAATGATCT-5′, product size 161 bp. PCR was
performed on the corresponding cDNA synthesized from each sample in
triplicate with the GeneAmp 5700 system (Applied Biosystems; Thermo
Fisher Scientific. Inc.), according to the manufacturer's protocol.
Thermocycling conditions for PCR were as follows: An initial
predenaturation step at 95°C for 15 min, followed by 35 cycles of
denaturation at 94°C for 30 sec, annealing at 50°C for 30 sec,
extension at 72°C for 1 min and a final extension step at 72°C for
10 min. mRNA expression levels were normalized to 18S rRNA, and
data was analyzed using the 2−ΔΔCq method (21).
Western blotting
Western blot analysis was performed to analyze the
expression of ChM-I expression in the overexpression and siRNA
groups. Total protein from cultured cells were lysed using lysis
buffer supplemented with phenylmethylsulfonyl fluoride (1 mM) on
ice as described previously (19).
The protein concentration in each sample was determined using a
bicinchoninic acid assay kit (Thermo Fisher Scientific, Inc.). A
total of 20 µg protein was separated by 10% SDS-PAGE and then
transferred to a polyvinylidene difluoride membrane (EMD Millipore,
Billerica, MA, USA). Membranes were blocked with 5% non-fat milk
powder at room temperature for 1 h and incubated for 1 h with
primary antibodies at room temperature, including a
chondromodulin-I antibody (catalog no. NBP1-59348; Novus
Biologicals, LLC, Littleton, LO, USA) and a GAPDH antibody (catalog
no. NBP300-221H, Novus Biologicals, LLC) at 1:1,000 dilution. After
three 10-min washes in TBS with Tween-20, membranes were incubated
with secondary antibodies labeled with horseradish peroxidase
(catalog no. NBP600-894; Novus Biologicals, LLC; 1:1,000) for 1 h
at room temperature. At last, the signals were detected using an
Enhanced Chemiluminescence kit (Pierce; Thermo Fisher Scientific,
Inc.) and the membranes were scanned and analyzed to determine the
protein expression. Image J software version 1.46 (National
Institutes of Health, Bethesda, MD, USA) was used for densitometric
analysis. The protein expression was normalized to an endogenous
reference GAPDH and relative to the control.
DNA synthesis assay
A bromodeoxyuridine (BrdU) incorporation assay was
used to assess the proliferation of Saos-2 cells. Cells were
harvested with trypsin/EDTA and suspended in DMEM containing 10%
FBS. The cells were seeded at 2×104 cells/ml into a
96-well plate (100 µl/well) and cultured for 48 h at 37°C.
Following medium replacement with medium containing either 15 or 30
µg/ml recombinant human (rh)ChM-I, 20 µl 1X BrdU was added prior to
incubation of the cells for 12, 24, 36 or 48 h at room temperature.
Cells were labeled with BrdU during the last 3 h of this
incubation. The medium was then replaced with one containing either
15 or 30 µg/ml rhChM-I, BrdU was added, and the cells were cultured
for 12, 24, 36 or 48 h. BrdU incorporation by the cells was
measured at each time-point using a cell proliferation ELISA BrdU
colorimetric kit (catalog no. ab126556; Abcam, Cambridge, UK)
according to the manufacturer's protocol, in triplicate.
Quantification of BrdU was performed by measuring the absorbance at
450 nm, referenced at 655 nm using a DU 800 spectrophotometer
(Beckman Coulter, Inc., Brea, CA, USA).
In vitro cell proliferation assay
A Cell Counting Kit-8 (CCK-8; Dojindo Laboratories,
Kumamoto, Japan) was used to determine cell proliferation as
described previously (http://www.dojindo.com). For each group, a 200-µl
suspension containing 104 cells was added per well into
a 96-well plate and incubated overnight at 37°C. Following this,
culture medium was removed and fresh medium (100 µl/well) and 10 µl
CCK-8 solution were added, and cells were incubated for 4 h at
37°C. On days 1, 3, 5 and 7, the absorbance of the supernatant was
measured spectrophotometrically at 450 nm (Beckman Coulter, Inc.).
Cell counts were determined using a calibration curve and a Nikon
E400 microscope (Nikon Corporation, Tokyo, Japan), and the
experiments were performed in triplicate.
Cell invasion and migration
assays
Transwell (8-µm pore size, Corning Life Sciences,
Corning, NY, USA) was use to assessed cell invasion and migration
capacity. According to the manufacturer's protocol,
2×104 cells suspended in serum-free medium were seeded
into Transwell inserts either coated (for invasion assay) or
uncoated (for migration assay) with growth factor-reduced Matrigel
(BD Biosciences, Franklin Lakes, NJ, USA) (19). Bottom wells were incubated with
complete medium for 24 h, and the invaded cells were fixed with 4%
paraformaldehyde and stained with a crystal violet solution at room
temperature for 20 min, and the number of penetrating cells was
counted using a Nikon E400 microscope (Nikon Corporation).
Scratch wound assay
Cells migration was measured by the scratch wound
assay (22). Cells were plated at
a density of 5×105 cells to 6-well-plates (Corning
Costar, Kaiserslautern, Germany) 24 h prior to the scratch. A cell
free area spanning approximately 500 µm in diameter was scratched
using a crystal 10 µl pipette tip after 24 h. The wells were washed
with medium to remove cellular debris. Cells were incubated with
DMEM without FBS. After 24 h, Gap width was measured using ImageJ
v1.46 (Wayne Rasband National Institutes of Health, Bethesda, MD,
USA).
Colony-formation assay
The colony-formation assay was performed to assess
the anchorage-dependent proliferation of cells. The log-phase cells
were harvested, and cultured in 6-well dishes as described
previously (23). The number of
colonies was stained with Coomassie Brilliant Blue (Sigma-Aldrich;
Merck KGaA) at room temperature for 15 min, and the visible
colonies were counted after 1, 2 and 3 weeks of incubation under a
phase contrast microscope. This assay was performed in triplicate
at least.
In vivo tumorigenicity assay
For in vivo assessment of tumor formation,
nude BALB/c athymic mice (male, aged 6 weeks, 20 g) purchased from
the Shanghai Laboratory Animal Centre (Shanghai, China) were
subcutaneously injected with 107 cultured human Saos-2
osteosarcoma cells suspended 500 µl sterile PBS in to evaluate the
effect of ChM-I on tumor growth in vivo. Once tumors reached
a volume of 200 mm3, mice (n=6/group) were injected with
5 mg/kg recombinant human (rh)ChM-I (experimental group) or 500 µl
PBS (control group). Mice received injections twice a week for 5
weeks. Tumor length and width were measured with calipers over a
period of 5 weeks. The tumor volume was calculated as (length ×
width2)/2. The mice were checked daily for up to 3 weeks
for tumor formation (24). Mice
were housed at a temperature of 25±12°C, humidity of 55±5% and 12 h
light dark cycle. Food and water were freely available.
Statistical analysis
The data are presented e.g., as the mean ± standard
deviation. One-way analysis of variance followed by Tukey's post
hoc test was performed to analyze the differences between groups
followed by Turkey post hoc test. P<0.05 was considered to
indicate a statistically significant difference. SPSS software
version 18.0 (SPSS, Inc., Chicago, IL, USA) was used for
analysis.
Results
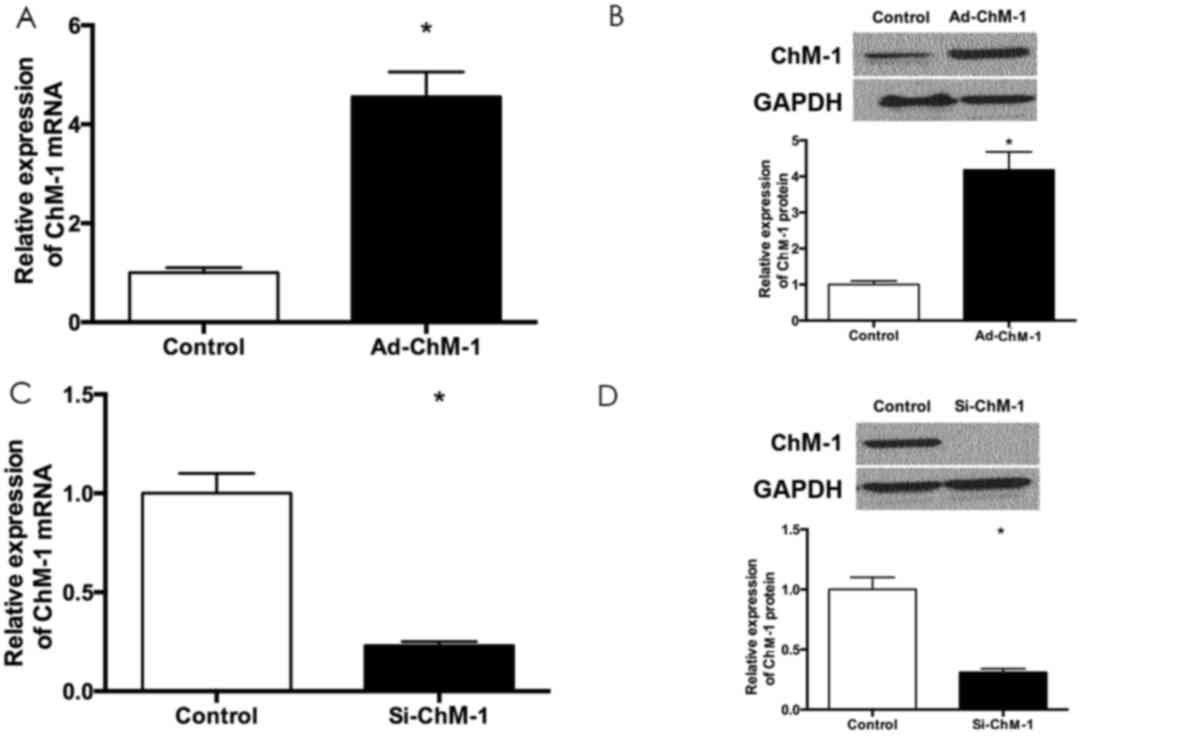
ChM-I expression is upregulated by
overexpression and downregulated by siRNA
As demonstrated by RT-qPCR and western blotting, the
expression of ChM-I was increased in the Ad-ChM-I group compared
with the control group (P<0.05), while the expression of ChM-I
was inhibited significantly by siRNA targeted to ChM-I (P<0.05;
Fig. 1).
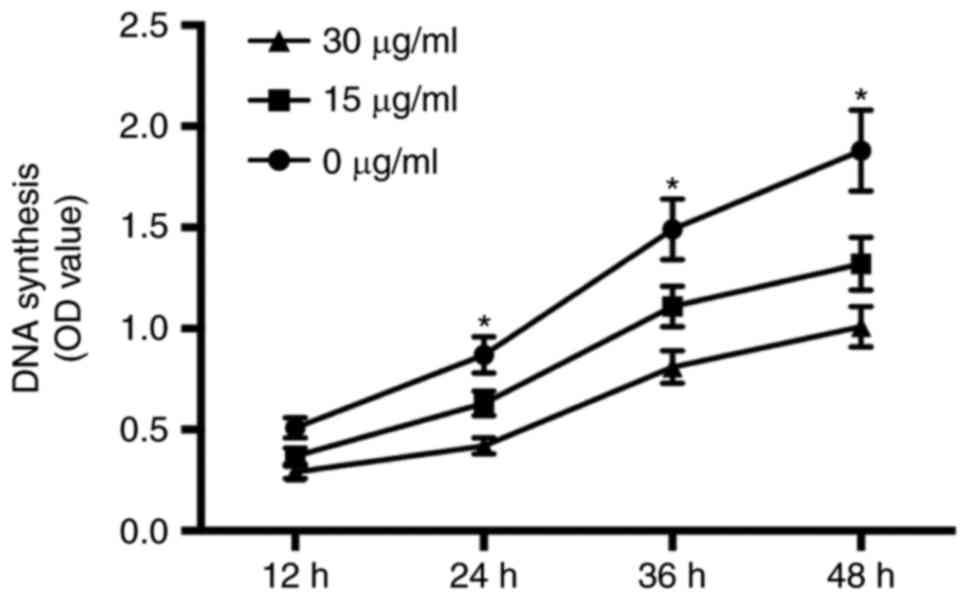
ChM-I inhibits DNA synthesis of
osteosarcoma cells
The effect of rhChM-I protein on DNA synthesis in
osteosarcoma cells was evaluated by a BrdU incorporation
assay. The results demonstrated that 15 and 30 µg/ml BrdU
suppressed DNA synthesis significantly at 24, 36 and 48 h
(P<0.05; Fig. 2).
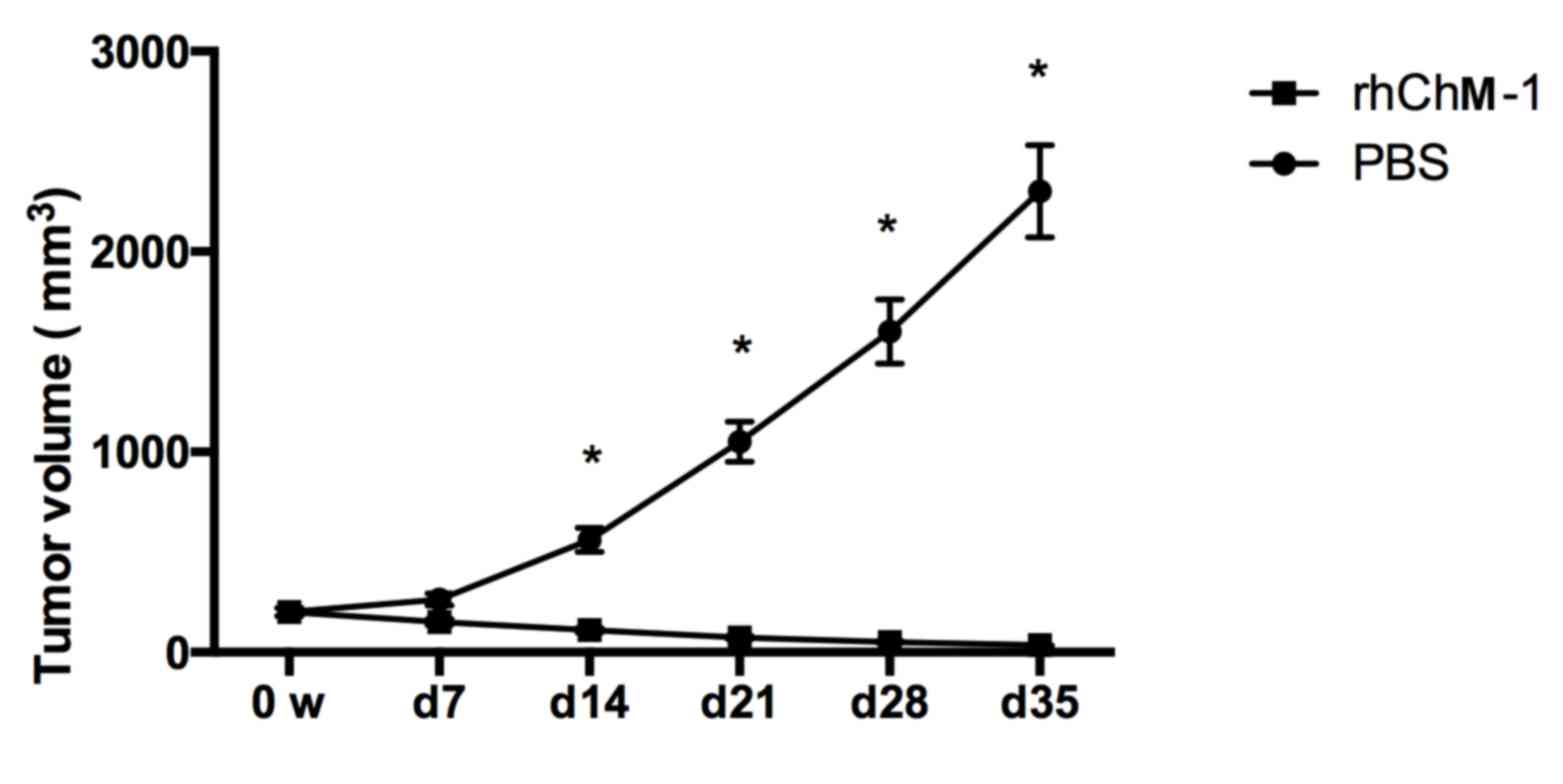
ChM-I suppresses tumor growth in
vivo
In order to evaluate the effect of ChM-I on tumor
growth in vivo, nude BALB/c athymic mice were injected with
human osteosarcoma cells. Upon reaching a volume of 200
mm3, the tumors were injected with rhChM-I protein
(experimental group) or PBS (control group). The tumors grew
exponentially in the PBS group, while the tumor in the experimental
group reduced significantly at 14 days after injection, and were
further reduced in a time-dependent manner (Fig. 3).
ChM-I suppresses cells proliferation
in vitro
To assess the effect of ChM-I on the proliferation
of human Saos-2 osteosarcoma cells in vitro, cells
transfected with pcDNA3.1(+)-ChM-I and cells knocked down using
siRNA were used. The results demonstrated that ChM-I overexpression
suppressed cell proliferation (Fig.
4A; P<0.05), while ChM-I siRNA enhanced cell proliferation
(Fig. 4B; P<0.05) in
vitro.
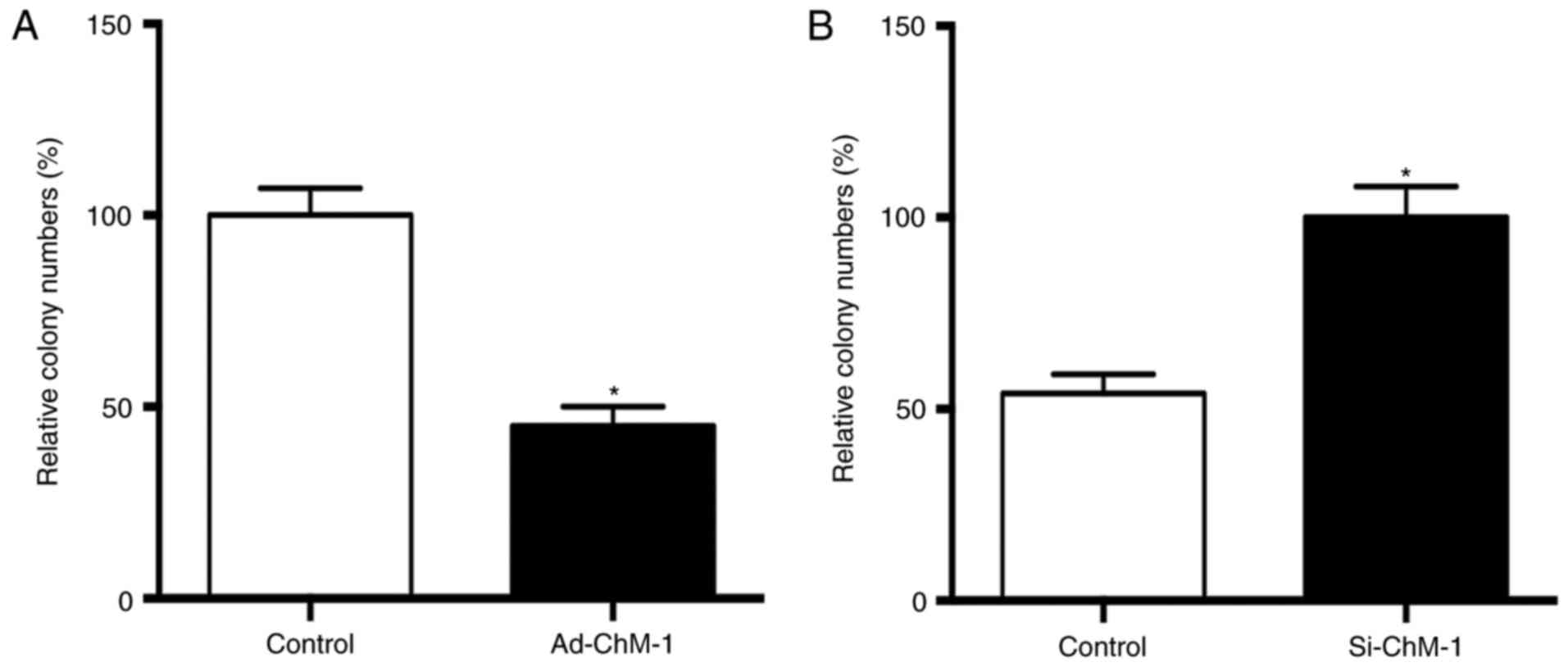
ChM-I suppresses the colony formation
ability of human osteosarcoma cells in vitro
In order to assess the effects of ChM-I on the
growth of osteosarcoma cells, an agarose-based colony formation
assay was performed according to a previous study (25). The results demonstrated that the
overexpression of ChM-I suppressed the colony formation ability of
human osteosarcoma cells (Fig. 5A;
P<0.05), while ChM-I knockdown enhanced the colony formation
ability of human osteosarcoma cells (Fig. 5B; P<0.05).
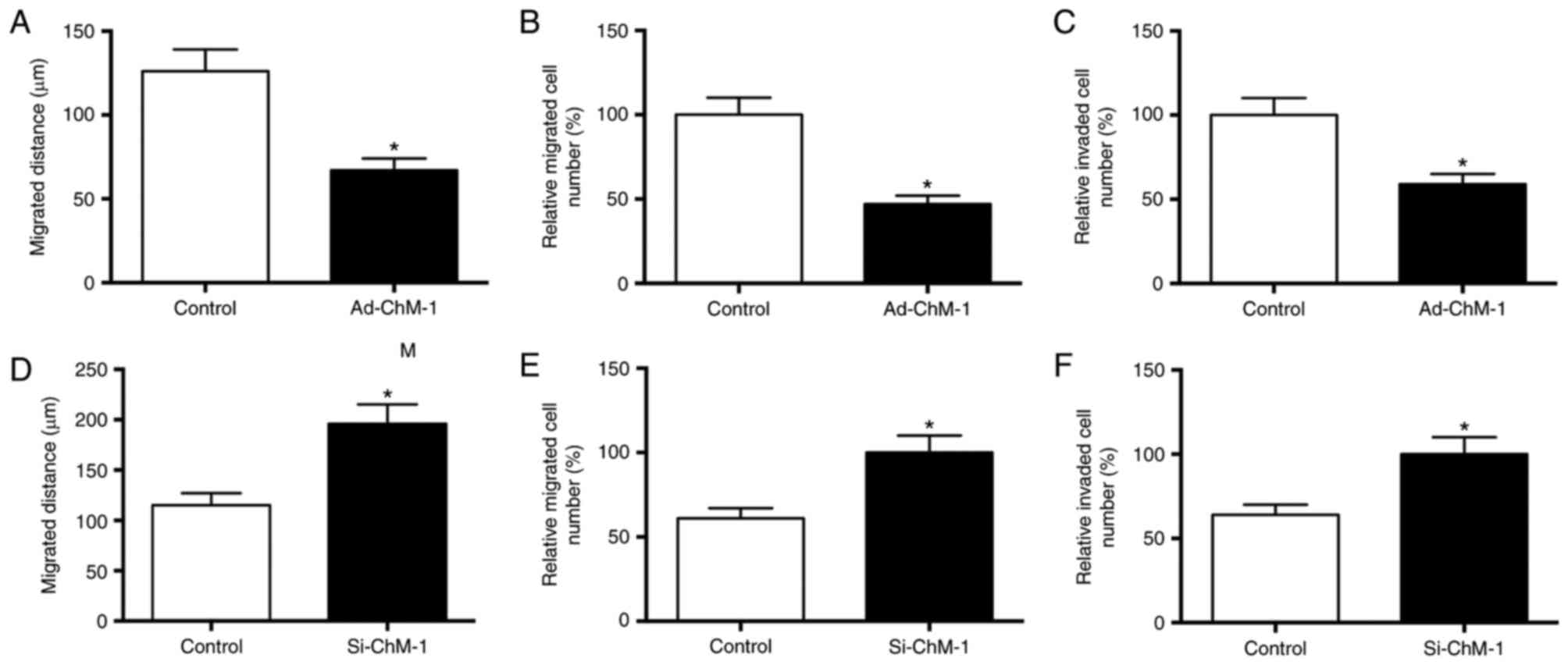
ChM-I suppresses osteosarcoma cell
migration and invasion
As presented in Fig.
6, compared with control cells, cells in the ChM-I
overexpression group demonstrated decreased cell migration in the
scratch wound assay, while cells in the ChM-I knockdown group
demonstrated increased migration ability. The results were further
confirmed by a three-dimensional cell migration assay and an
invasion assay, which demonstrated that ChM-I overexpression
resulted in a significant reduction in the migration and invasion
ability, while ChM-I knockdown resulted in a significant increase
in migration and invasion ability (Fig. 6).
Discussion
Osteosarcoma, the most common bone tumor occurs in
children and adolescents, is an aggressive and highly malignant
cancer (26,27). It occurs slightly more commonly in
male than female patients, and has a predilection for the
metaphyseal regions adjacent to physes with the greatest growth in
the distal femur, proximal tibia and proximal humerus (28,29).
In ~15–20% of patients, osteosarcoma has spread by the time it is
diagnosed (30). Therefore, early
diagnosis and early treatment serves a very important role in the
prognosis of patients. Certain methods have been used to cure this
disease, such as surgery, and chemo- and radiotherapy, which
depends on the stage of tumor development (31–33).
Angiogenesis is considered to serve a crucial role in tumor growth,
tumor propagation and metastasis formation; tumors cannot grow
beyond 1–2 mm in diameter without angiogenesis (34). Therefore, anti-angiogenic drugs and
treatments have been developed to inhibit the formation of new
blood vessels and block the blood supply of tumors in order to
inhibit or cure tumors (35,36).
It has been reported that ChM-I, a 25-kDa
glycoprotein purified from cartilage on the basis of
growth-promoting activity for chondrocytes, was recently identified
as an angiogenesis inhibitor (13,14).
ChM-I promotes growth and colony formation of cultured
chondrocytes, but inhibits angiogenesis in vitro and in
vivo. A previous study demonstrated that ChM-I inhibited
vascular invasion into cartilage that was ectopically induced by
demineralized bone matrix in nude mice, leading to the suppression
of replacement of cartilage by bone in vivo (13). Klinger et al (37) revealed that adeno-associated virus
(AAV) vectors carrying ChM-I complementary DNA (AAV-ChM-I)
efficiently produced ChM-I, which had strong antiangiogenic
effects, as verified by the inhibition of tube formation of
endothelial cells. Additionally, previous studies have demonstrated
that ChM-I exerts an antitumor effect, which may be based on the
effect of angiogenesis inhibition (16,38).
However, to the best of our knowledge, no previous studies have
demonstrated the effect of ChM-I on osteosarcoma. Therefore, the
present study aimed to determine the possible direct antitumor
effects of ChM-I in osteosarcoma.
The results demonstrated that ChM-I not only has the
ability to suppress angiogenesis, but also to suppress tumor cell
proliferation in vitro and inhibit tumor growth in
vivo. Zhang et al (19)
also reported that ChM-I functions as a tumor suppressor in gastric
adenocarcinoma, which indicated that ChM-I is a potential tumor
suppressor in gastric cancer, suggesting that it may be useful as a
biomarker for the treatment and prognosis of gastric cancer.
In the present study, the results of the BrdU
incorporation assay demonstrated that ChM-I inhibits DNA synthesis
of osteosarcoma cells. ChM-I was demonstrated to exhibit antitumor
effects based on direct inhibition of tumor-cell growth. The
results of the CCK-8 assay indicated that overexpression of ChM-I
inhibited the proliferation of human osteosarcoma cells
significantly, while ChM-I knockdown enhanced the proliferation of
human osteosarcoma cells. The direct antitumor effects of ChM-I on
human osteosarcoma cells were further proven by a colony formation
assay. The agarose-based clonogenic assay demonstrated that ChM-I
inhibited the proliferation of osteosarcoma cells significantly. In
a study by Shao et al (20), they transfected ChM-I into human
breast cancer cells, and observed that it inhibited the growth of
human breast cancer cells. In the present study, the in vivo
xenograft study demonstrated that ChM-I inhibited osteosarcoma
growth in nude mice. The anti-cancer efficacy of ChM-I may be due
to its marked impact on the tumor microenvironment with additional
direct suppression of tumor cell proliferation, which has been
demonstrated to be exerted via activation of transforming growth
factor β in tumor cells overexpressing this protein (39).
In conclusion, the present study demonstrated that
ChM-I exerts antitumor effects by directly suppressing the
proliferation and growth of osteosarcoma cells. The direct
antitumor effects and anti-angiogenic effects make ChM-I a
promising potential drug in treating osteosarcoma.
Acknowledgements
The present study was supported by the Rizhao
Committee of Science and Technology of Shandong, China (grant no.
201539).
References
|
1
|
Kansara M, Teng MW, Smyth MJ and Thomas
DM: Translational biology of osteosarcoma. Nat Rev Cancer.
14:722–735. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Pisani P and Ferlay J: Global
cancer statistics. CA Cancer J Clin. 49:33–64. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bielack S, Kempf-Bielack B, Von Kalle T,
Schwarz R, Wirth T, Kager L and Whelan J: Controversies in
childhood osteosarcoma. Minerva Pediatr. 65:125–48. 2013.PubMed/NCBI
|
|
4
|
Moore DD and Luu HH: Osteosarcoma. Cancer
Treat Res. 162:65–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fan Z, Patel S, Lewis VO, Guadagnolo BA
and Lin PP: Should High-grade extraosseous osteosarcoma Be treated
with multimodality therapy like other soft tissue sarcomas? Clin
Orthop Relat Res. 473:3604–3611. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jones KB, Ferguson PC, Lam B, Biau DJ,
Hopyan S, Deheshi B, Griffin AM, White LM and Wunder JS: Effects of
neoadjuvant chemotherapy on image-directed planning of surgical
resection for distal femoral osteosarcoma. J Bone Joint Surg Am.
94:1399–1405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peng N, Gao S, Guo X, Wang G, Cheng C, Li
M and Liu K: Silencing of VEGF inhibits human osteosarcoma
angiogenesis and promotes cell apoptosis via VEGF/PI3K/AKT
signaling pathway. Am J Transl Res. 8:1005–1015. 2016.PubMed/NCBI
|
|
10
|
Dossi R, Frapolli R, Di Giandomenico S,
Paracchini L, Bozzi F, Brich S, Castiglioni V, Borsotti P, Belotti
D, Uboldi S, et al: Antiangiogenic activity of trabectedin in
myxoid liposarcoma: Involvement of host TIMP-1 and TIMP-2 and tumor
thrombospondin-1. Int J Cancer. 136:721–729. 2015.PubMed/NCBI
|
|
11
|
Miura S, Kondo J, Takimoto A, Sano-Takai
H, Guo L, Shukunami C, Tanaka H and Hiraki Y: The N-terminal
cleavage of chondromodulin-I in growth-plate cartilage at the
hypertrophic and calcified zones during bone development. PLoS One.
9:e942392014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aoyama T, Okamoto T, Fukiage K, Otsuka S,
Furu M, Ito K, Jin Y, Ueda M, Nagayama S, Nakayama T, et al:
Histone modifiers, YY1 and p300, regulate the expression of
cartilage-specific gene, chondromodulin-I, in mesenchymal stem
cells. J Biol Chem. 285:29842–29850. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hiraki Y and Shukunami C: Chondromodulin-I
as a novel cartilage-specific growth-modulating factor. Pediatr
Nephrol. 14:602–605. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hiraki Y, Inoue H, Iyama K, Kamizono A,
Ochiai M, Shukunami C, Iijima S, Suzuki F and Kondo J:
Identification of chondromodulin I as a novel endothelial cell
growth inhibitor. Purification and its localization in the
avascular zone of epiphyseal cartilage. J Biol Chem.
272:32419–32426. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yukata K, Matsui Y, Shukunami C, Takimoto
A, Hirohashi N, Ohtani O, Kimura T, Hiraki Y and Yasui N:
Differential expression of Tenomodulin and Chondromodulin-1 at the
insertion site of the tendon reflects a phenotypic transition of
the resident cells. Tissue Cell. 42:116–120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Oshima Y, Sato K, Tashiro F, Miyazaki J,
Nishida K, Hiraki Y, Tano Y and Shukunami C: Anti-angiogenic action
of the C-terminal domain of tenomodulin that shares homology with
chondromodulin-I. J Cell Sci. 117:2731–2744. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsai AC, Pan SL, Sun HL, Wang CY, Peng CY,
Wang SW, Chang YL, Kuo SC, Lee KH and Teng CM: CHM-1, a new
vascular targeting agent, induces apoptosis of human umbilical vein
endothelial cells via p53-mediated death receptor 5 up-regulation.
J Biol Chem. 285:5497–5506. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hsu SC, Yang JS, Kuo CL, Lo C, Lin JP,
Hsia TC, Lin JJ, Lai KC, Kuo HM, Huang LJ, et al: Novel quinolone
CHM-1 induces apoptosis and inhibits metastasis in a human
osterogenic sarcoma cell line. J Orthop Res. 27:1637–1644. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang P, Wang Y, Xu P, Song S, Zhu X, Shi
Z, Gao S and Feng X: Chondromodulin-1 functions as a tumor
suppressor in gastric adenocarcinoma. Int J Oncol. 47:941–950.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shao J, Gan L and Wang J: Transfection of
chondromodulin I into human breast cancer cells and its effect on
the inhibition of cancer cell growth. Mol Med Rep. 13:4303–4308.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roloff F, Ziege S, Baumgärtner W, Wewetzer
K and Bicker G: Schwann cell-free adult canine olfactory
ensheathing cell preparations from olfactory bulb and mucosa
display differential migratory and neurite growth-promoting
properties in vitro. BMC Neurosci. 14:1412013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Franken NA, Rodermond HM, Stap J, Haveman
J and van Bree C: Clonogenic assay of cells in vitro. Nat Protoc.
1:2315–2319. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li P, Chen X, Su L, Li C, Zhi Q, Yu B,
Sheng H, Wang J, Feng R, Cai Q, et al: Epigenetic silencing of
miR-338-3p contributes to tumorigenicity in gastric cancer by
targeting SSX2IP. PLoS One. 8:e667822013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shingu T, Holmes L, Henry V, Wang Q, Latha
K, Gururaj AE, Gibson LA, Doucette T, Lang FF, Rao G, et al:
Suppression of RAF/MEK or PI3K synergizes cytotoxicity of receptor
tyrosine kinase inhibitors in glioma tumor-initiating cells. J
Transl Med. 14:462016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang Z, Li X, Yang Y, He Z, Qu X and Zhang
Y: Long noncoding RNAs in the progression, metastasis, and
prognosis of osteosarcoma. Cell Death Dis. 7:e23892016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
PDQ Pediatric Treatment Editorial Board, .
Osteosarcoma and Malignant Fibrous Histiocytoma of Bone Treatment
(PDQ®)PDQ Cancer Information Summaries [Internet].
Health Professional Version. National Cancer Institute (US);
Bethesda: 2002
|
|
28
|
Abarrategi A, Tornin J, Martinez-Cruzado
L, Hamilton A, Martinez-Campos E, Rodrigo JP, González MV, Baldini
N, Garcia-Castro J and Rodriguez R: Osteosarcoma: Cells-of-Origin,
cancer stem cells, and targeted therapies. Stem Cells Int.
2016:36317642016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li CJ, Liu XZ, Zhang L, Chen LB, Shi X, Wu
SJ and Zhao JN: Advances in Bone-targeted drug delivery systems for
neoadjuvant chemotherapy for osteosarcoma. Orthop Surg. 8:105–110.
2016. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Friebele JC, Peck J, Pan X, Abdel-Rasoul M
and Mayerson JL: Osteosarcoma: A meta-analysis and review of the
literature. Am J Orthop (Belle Mead NJ). 44:547–553.
2015.PubMed/NCBI
|
|
31
|
Li S, Sun W, Wang H, Zuo D, Hua Y and Cai
Z: Research progress on the multidrug resistance mechanisms of
osteosarcoma chemotherapy and reversal. Tumour Biol. 36:1329–1338.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dass CR and Choong PF: Gene therapy for
osteosarcoma: Steps towards clinical studies. J Pharm Pharmacol.
60:405–413. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Clark JC, Dass CR and Choong PF: A review
of clinical and molecular prognostic factors in osteosarcoma. J
Cancer Res Clin Oncol. 134:281–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kerbel RS: Tumor angiogenesis: Past,
present and the near future. Carcinogenesis. 21:505–515. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Maione F and Giraudo E: Tumor
angiogenesis: Methods to analyze tumor vasculature and vessel
normalization in mouse models of cancer. Methods Mol Biol.
1267:349–365. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kubota Y: Tumor angiogenesis and
anti-angiogenic therapy. Keio J Med. 61:47–56. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Klinger P, Surmann-Schmitt C, Brem M,
Swoboda B, Distler JH, Carl HD, von der Mark K, Hennig FF and Gelse
K: Chondromodulin 1 stabilizes the chondrocyte phenotype and
inhibits endochondral ossification of porcine cartilage repair
tissue. Arthritis Rheum. 63:2721–2731. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Miura S, Kondo J, Kawakami T, Shukunami C,
Aimoto S, Tanaka H and Hiraki Y: Synthetic disulfide-bridged cyclic
peptides mimic the anti-angiogenic actions of chondromodulin-I.
Cancer Sci. 103:1311–1318. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: A convenient and inexpensive method for analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|