Introduction
Abnormal vascular smooth muscle cell (VSMC) growth
contributes to the pathogenesis of vascular remodeling of large
arteries during hypertension. Autocrine and paracrine growth
factors may modulate VSMC growth, partly by promoting the secretion
of growth factors (1), including
platelet-derived growth factor (2,3).
Cyclophilin A (CyPA) has been identified to be a growth factor
secreted from VSMCs and has a direct mitogenic effect on the growth
of human aorta smooth muscle cells (4) and rat VSMCs (5). Additionally, CyPA mediates vascular
remodeling by promoting vascular smooth muscle cell proliferation
and inflammation (6). Previous
studies demonstrated that CyPA levels in rat serum gradually
increased with the development of hypertension (7), and were positively correlated with
systolic and diastolic blood pressure in untreated patients
(8). The extracellular matrix
metalloproteinase inducer basigin (CD147) was reported to be a
surface receptor for extracellular CyPA (9) and mediates CyPA-specific signaling
events. CyPA activates extracellular signal-regulated kinase (ERK)
1/2 via binding with CD147, leading to upregulation of cyclin D1
expression in cholangiocarcinoma cells (10). However, the kinetics of the
alterations in serum CyPA and its downstream signaling cascade
(CD147-ERK1/2-cyclin) during thoracic aorta remodeling have not
been completely elucidated. Therefore, the present study was
performed to further examine the kinetics of alterations in serum
CyPA and its downstream signaling cascade (CD147-ERK1/2-cyclin),
and to clarify the potential of CyPA to be a biomarker of the
extent of thoracic aorta remodeling.
Geranylgeranyl pyrophosphate (GGPP) and farnesyl
pyrophosphate (FPP) are essential for the geranylgeranylation and
farnesylation of Rho and Ras family small G proteins, respectively
(11). The downstream effectors of
Rho include the Rho kinase family members, consisting of
Rho-associated kinase 1 (ROCK1) and ROCK2, which are
serine-threonine kinases that are activated by Rho GTPases
(12), and recent studies have
demonstrated that ROCK2 regulates the secretion of proinflammatory
cytokines (13). It has been
demonstrated that 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA)
reductase inhibitors (statins) prevent the conversion of HMG-CoA to
mevalonate, and thereby inhibit the synthesis of the other products
of the mevalonate pathway, including GGPP and FPP. By inhibiting
the synthesis of GGPP, statins prevent the activation of Rho
GTPases and Rho-associated kinases (11). Simvastatin has been observed to
decrease reactive oxygen species (ROS) -mediated CyPA release in
cultured VSMCs, and the pan ROCK inhibitor Y27632 has exhibited
similar effects to simvastatin (14); however, it remains unclear whether
ROCK1 or ROCK2 regulates the secretion of CyPA. In addition, it is
uncertain whether simvastatin-inhibited CyPA secretion is mediated
by the Rho-Rho kinase pathway. These uncertainties provided a basis
for further investigation of whether ROCK1 or ROCK2 is involved in
CyPA secretion from VSMCs. Previous studies have demonstrated that
CyPA levels in rat serum gradually increase with the development of
hypertension (7) and contribute to
vascular remodeling (6).
Atorvastatin was reported to improve thoracic aortic remodeling in
spontaneously hypertensive rats (15). However, the novel underlying
mechanisms have not been completely elucidated. Available data
about the effect of simvastatin on thoracic aorta remodeling during
hypertension are limited. The above previous studies led to the
question of whether simvastatin may improve thoracic aorta
remodeling by regulating serum CyPA levels, and whether the Rho-Rho
kinase signaling pathway mediates simvastatin-inhibited CyPA
secretion. In addition, the present study aimed to examine whether
ROCK1 or ROCK2 regulates the secretion of CyPA.
The present study investigated whether simvastatin
was able to affect serum CyPA levels in hypertensive rats and
downstream signaling cascades in the remodeled thoracic aorta. The
mechanism underlying simvastatin-inhibited CyPA secretion was
additionally examined. The results of the present study
demonstrated that treatment with simvastatin diminished the
time-dependent upregulation of serum CyPA levels and
CD147-ERK1/2-cyclin signaling pathway in the remodeled thoracic
aorta. The secretion of CyPA is ROCK2-dependent. Additionally,
simvastatin-inhibited CyPA release was mediated by the Rho-Rho
kinase pathway in VSMCs.
Materials and methods
Reagents and cell culture
Simvastatin and LY83583 were purchased from
Calbiochem (Merck KGaA, Darmstadt, Germany). Geranylgeraniol (GGOL)
was purchased from Sigma-Aldrich (Merck KGaA).
Geranylgeranyltransferase inhibitor (GGTI) 298 and KD025 were
purchased from Cayman Chemical Company (Ann Arbor, MI, USA). The
above reagents were dissolved in dimethyl sulfoxide (DMSO) and
stored at −20°C. The final concentration of DMSO in the culture
medium was <0.1%. VSMCs were isolated from the thoracic aorta
media of male Sprague-Dawley rats (80–100 g) and maintained in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a
humidified atmosphere of 5% CO2/95% air. VSMCs were
confirmed by positive staining with α-smooth muscle actin antibody,
as described previously (16).
Animal model and primary culture of
VSMCs
The present study was approved by the Animal Care
and Use Committee of Guangzhou Medical University (Guangzhou,
China). All animal experimental procedures were performed in line
with the Guide for the Care and Use of Laboratory Animals issued by
the Ministry of Science and Technology of China. A total of 96 male
adult rats were used for these experiments. Two-kidney, two clip
renovascular hypertensive rats were induced as described by our
previous studies (7,16). U-shaped silver clips with an
internal diameter of 0.3 mm were placed around the right and left
renal arteries of 8 to 10-week old healthy normotensive male
Sprague-Dawley rats (80–100 g), after anaesthetized by 10% chloral
hydrate (300 mg/kg; intraperitoneal injection). The criterion for
hypertension was a systolic blood pressure of ≥150 mmHg. Following
3 days of adaptation, rats were randomly allocated to sham,
hypertension (Htn) or simvastatin groups (Sim; hypertensive rats
treated with simvastatin). Hypertension was induced using U-shaped
silver clips, as described above. The Sim group received
simvastatin by daily gavage (10 mg/kg/day) from the second day
post-clipping. The sham-operated rats were treated in the same
manner without the placement of silver clips, and served as the
control. All groups were investigated for 1, 4, 8 and 12 weeks
following surgery. The rats were injected with penicillin G (25,000
U intramuscular) prior to surgery to mitigate the risk of
infection. Systolic blood pressure was detected preoperatively and
every 3 days post-surgery under a conscious state using the
indirect tail-cuff method (Powerlab 4/30; AD Instruments, Sydney,
New South Wales, Australia), subsequent to warming of the rats at
35°C for 5 min under mild restraint. The average of three pressure
readings was recorded for each measurement. Animals were supplied
by the Experimental Animal Center of Sun Yat-Sen University
(Guangzhou, China). All rats were maintained in pathogen-free
facilities with a 12-h light/dark cycle at 23–25°C, with a relative
humidity of 55–60% and ad libitum access to food and
water.
To investigate whether CyPA secretion from VSMCs was
influenced by treatment with simvastatin in rats, VSMCs of the
thoracic aorta from sham, Htn and Sim rats at the 12-week point
were isolated and cultured in DMEM containing 10% FBS in an
incubator with 95% O2 and 5% CO2 at 37°C, as
previously described (17). CyPA
in conditioned medium was detected by western blotting as
previously reported (14).
Histological analysis of thoracic
aorta specimens, and hematoxylin and eosin (H&E) staining
Following anesthetization at 1, 4, 8 and 12 weeks
with 10% chloral hydrate, the thorax was opened, perfusion
commenced at a pressure of 55±5 mmHg, and the right atrium incised
to allow drainage of blood and perfusate. The same pressure was
used in all animals so that all aortas would be fixed under uniform
tension. Perfusion with 0.1 mol/l phosphate buffer containing
heparin (100 U/kg) for 5 min was followed by 4°C fixative solution
containing 4% freshly depolymerized paraformaldehyde in 0.1 mol/l
phosphate buffer for 15 min. At a defined distance from the aortic
arch (1 cm), a segment of the thoracic aorta was dissected free of
perivascular tissue, cleaned and immersed in 4% fresh
paraformaldehyde for an additional 3–4 h at 4°C. Following
fixation, each thoracic aorta was cut transversely into 5–6 rings
of 2–3-mm thickness, which were subsequently embedded in paraffin.
Sections (4-µm thick) of paraffin-embedded thoracic aortas that
included the entire circumference of each ring were sectioned from
five different blocks in each animal. Sections were stained with
H&E for ~2 min at room temperature for general morphology and
quantitative morphometric analysis. Thoracic aortas without
perfusion were rapidly frozen at −80°C for subsequent western blot
analysis.
Image acquisition, methods of
measurement and calculation of vascular remodeling parameters
All stained histological sections of thoracic aortas
were visualized under light microscopy (Eclipse-80i; Nikon
Corporation, Tokyo, Japan), and digital images of thoracic aortas
were captures using the imaging software NIS-Elements F v4.00.00
(Nikon Corporation) at magnification, ×40 and ×400, marked with a
micrometer scale. Integrated Performance Primitives software
(version 6.0; Intel Corporation, Santa Clara, CA, USA) was used to
measure the perimeter of the lumen (C) and media (without
adventitia) of sections from each thoracic aorta at magnification,
×40. The diameter of lumen (the internal diameter, Di) and external
diameter (De) were calculated using the formula: D=C/π, according
to a previously-published method with minor modifications (18,19).
The media thickness was calculated by subtraction of Di/2 from
De/2. The cross-sectional area occupied by the lumen was calculated
from its mean diameter (πD2/4). Medial area was
calculated by subtraction of the inner outlined area from the outer
outlined area according to the formula: Media
CSA=π(De2-Di2)/4. Remodeling was assessed by
media thickness and media area in paraffin-embedded tissue sections
of thoracic aortas with H&E staining.
ELISA for measurement of rat serum
CyPA
Rat serum CyPA was assayed as our previous
experiment (7), using a rat
cyclophilin A ELISA kit (cat. no. CSB-E14928r; Cusabio, Wuhan,
China). A total of 3–5 ml blood was collected with a serum vacuumed
blood collection tube, clotted overnight at 4°C, and centrifuged
for 15 min at 1,000 × g at 4°C. The separated serum was stored in
fresh tubes in aliquots at −80°C. Fresh samples were used for the
ELISA, according to the manufacturer's protocol. The optical
density of each well was read at 450 nm using a Multiskan GO
1.00.38 microplate reader (Thermo Fisher Scientific, Inc.). A
standard curve was produced and the CyPA concentration of each
sample was calculated.
Western blot analysis of the
expression of CyPA, CD147, phosphorylated-ERK1/2 and cyclins
Western blot analysis was performed as described
previously (20). Total proteins
were extracted from the media of rat thoracic aortas. Equal amounts
of total protein (20–30 µg) were loaded on an 8–12% SDS-PAGE gel
and transferred to nitrocellulose membranes (EMD Millipore,
Billerica, MA, USA). Membranes were blocked at room temperature for
1 h in 5% non-fat milk in TBST buffer [10 mM Tris (pH 7.5), 150 mM
NaCl, 0.1% Tween-20], incubated overnight at 4°C with the following
primary antibodies: Phospho (p)-ERK1/2 (cat. no. 4370; 1:1,000),
ERK1/2 (cat. no. 9102; 1:1,000), cyclin D1 (cat. no. 2978; 1:500),
CyPA (cat. no. 2175; 1:1,000), β-actin (cat. no. 8457; 1:2,000; all
from Cell Signaling Technology, Inc., Danvers, MA, USA), CD147 (cat
no. SAB4502896; 1:500; Sigma-Algrich; Merck KGaA), cyclin A (cat.
no. sc-239; 1:500) and cyclin E (cat. no. sc-247; 1:500; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). They were subsequently
incubated for 1 h at room temperature with the appropriate
anti-rabbit (cat. no. 7074) and anti-mouse (cat. no. 7076)
secondary horseradish peroxidase-conjugated antibodies (1:2,000;
Cell Signaling Technology, Inc.). Blots were incubated with
chemiluminescence substrates (cat. no. 34079; Pierce; Thermo Fisher
Scientific, Inc.) and were visualized using a gel image system. The
integrated optical density of target bands was determined using
Gel-Pro analyzer software v6.3 (Media Cybernetics, Inc., Rockville,
MD, USA). β-actin was used as an internal control to guarantee
equal loading and transfer of protein.
Assay of medium CyPA
concentration
CyPA in the medium was assayed as previously
reported (14). Following
pretreatment with simvastatin (5 µM), GGOL (10 µM), GGTI 298 (10
µM) and KD025 (10 µM) for 30 min at 37°C, VSMCs were exposed to 1
µmol/l LY83583 for 2 h at 37°C in a CO2 incubator.
Conditioned medium (CM) from VSMCs in each group was collected and
centrifuged at 4°C for 10 min at 800 × g to remove cell debris. The
medium was subsequently concentrated 100-fold using a Centricon
Plus-20 filter (EMD Millipore). CyPA in CM and total cell lysate
were detected using western blot analysis, as aforementioned.
Statistical analysis
Quantitative results were expressed as the mean ±
standard deviation of at least 3 independent experiments.
Statistical significance of differences between means was assessed
by one-way analysis of variance using SPSS 19.0 software (IBM
Corp., Armonk, NY, USA), and post hoc multiple comparisons were
analyzed using Bonferroni testing. P<0.05 was considered to
indicate a statistically significant difference.
Results
Simvastatin alleviates
hypertension-induced remodeling of the rat thoracic aorta
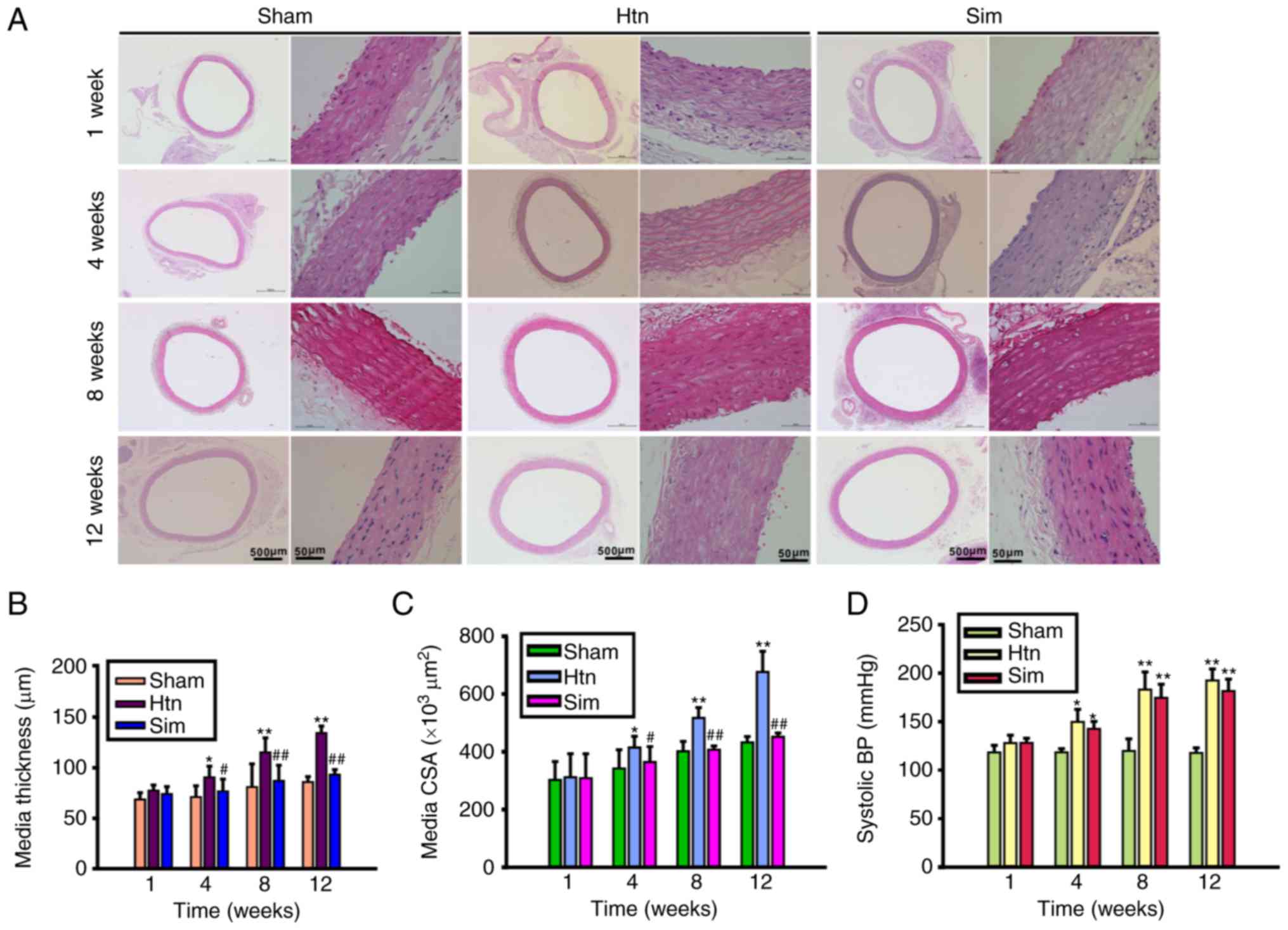
The present study evaluated the effect of
simvastatin on hypertension-induced remodeling of the rat thoracic
aorta. Representative images of transverse sections of
H&E-stained thoracic aorta and morphometric analysis of
thoracic aortic remodeling are presented in Fig. 1A-C. The thoracic aorta media
thickness (Fig. 1B), and the media
cross sectional area (Fig. 1C) of
hypertensive rats were significantly increased compared with those
of sham controls at 4, 8 and 12 weeks post-clipping. The results of
the present study indicated that the rat thoracic aortas from the
Htn group had undergone remodeling. However, treatment with
simvastatin ameliorated hypertension-induced thoracic aorta
remodeling at 4, 8 and 12 weeks, in a time-dependent manner.
Treatment with simvastatin slightly, although not significantly,
decreased the elevated systolic blood pressure at 4, 8 and 12 weeks
post-surgery (Fig. 1D). The
results of the present study suggested that treatment with
simvastatin attenuated thoracic aorta remodeling without a
significant lowering of blood pressure, indicating that the
beneficial effect of simvastatin on thoracic aorta remodeling
involves other mechanisms likely not associated with blood
pressure.
Treatment with simvastatin
significantly decreases serum CyPA and CyPA secretion from primary
VSMCs of thoracic aortas
It has been reported that pretreatment with
simvastatin inhibited CyPA secretion from cultured VSMCs induced by
ROS (14). The present study
further examined whether the rat serum CyPA concentration may be
influenced by treatment with simvastatin. The ELISA results
illustrated that serum CyPA levels were significantly elevated at
4, 8 and 12 weeks post-surgery, concomitant with the progress of
thoracic aorta remodeling, compared with the sham group (Fig. 2). The results of the present study
indicated that serum CyPA levels may be an indicator of the degree
of thoracic aorta remodeling. However, serum CyPA was decreased by
treatment with simvastatin for 4, 8 and 12 weeks, suggesting that
simvastatin may inhibit CyPA secretion in hypertensive rats. In
order to confirm further whether simvastatin may affect CyPA
secretion from VSMCs, VSMCs from the thoracic aortas of sham, Htn
and simvastatin-treated rats at the 12-week point were isolated and
cultured. The level of CyPA in the CM was determined by western
blotting. Notably, the LY83583-induced CyPA secretion from VSMCs of
Htn (12 weeks) rats was increased compared with that of sham (12
weeks) rats, suggesting that CyPA secretion from VSMCs increased
with the progress of thoracic aorta remodeling. Additionally,
LY83583-induced CyPA secretion from VSMCs of simvastatin-treated
rats (12 weeks) was decreased compared with that of Htn rats (12
weeks). These results further demonstrated that treatment with
simvastatin inhibited the secretion of CyPA from VSMCs during
remodeling of the thoracic aorta.
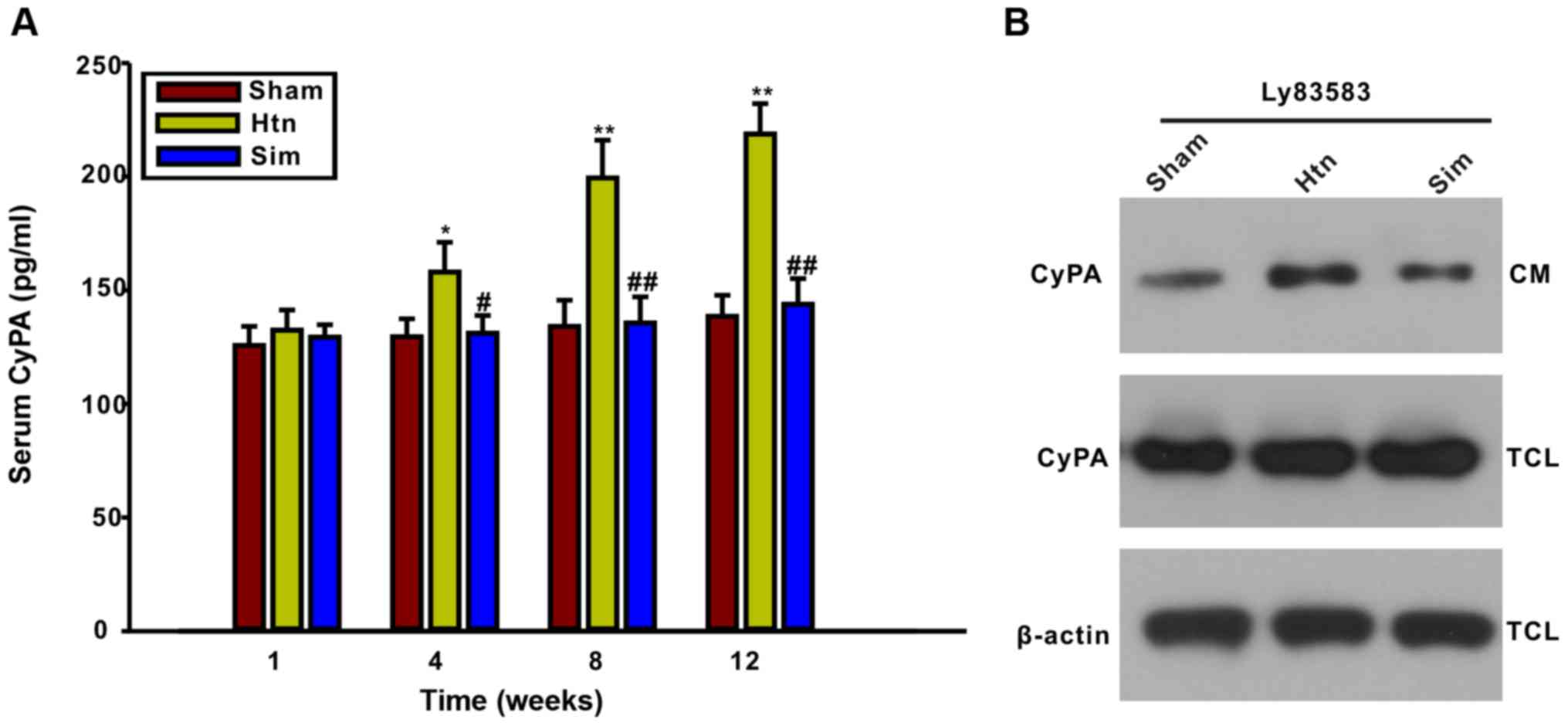 | Figure 2.Rat serum CyPA and CyPA secretion in
primary VSMCs from thoracic aortas is markedly decreased by
treatment with simvastatin. (A) Serum CyPA levels gradually
increased during the development of thoracic aorta remodeling, as
assayed using a rat CyPA ELISA kit. Treatment with simvastatin for
4, 8 and 12 weeks reduced the serum CyPA concentration. (B)
LY83583-induced CyPA secretion from primary VSMCs of
simvastatin-treated rats was decreased compared with that from
VSMCs of Htn rats at 12 weeks. Primary VSMCs were isolated from
sham, Htn and simvastatin-treated rats at 12 weeks and cultured
in vitro. Primary VSMCs were exposed to 1 µmol/l LY83583 for
2 h. The CyPA level in the CM and TCL were detected by western
blotting. Data are expressed as mean ± standard deviation from 3
independent experiments. *P<0.05, **P<0.01 vs. sham;
#P<0.05, ##P<0.01 vs. Htn. Htn,
hypertension; Sim, simvastatin; CM, conditioned medium; TCL, total
cell lysate; CyPA, cyclophilin A. |
Treatment with simvastatin reduces
upregulation of the CD147-ERK1/2-cyclin pathway with the extent of
thoracic aorta remodeling
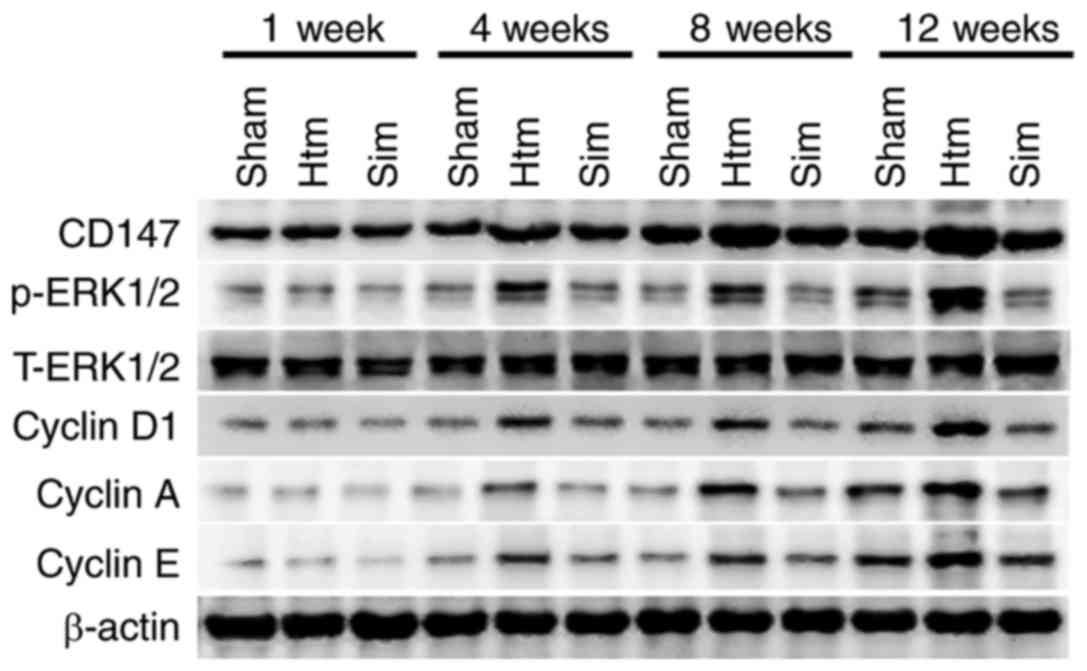
In order to determine the association between the
CD147-ERK1/2-cyclin pathway and the progression of remodeling
during hypertension, protein levels of CD147,
phosphorylated-ERK1/2, cyclin D1, cyclin A and cyclin E in rat
thoracic aortas were detected using western blotting. As presented
in Fig. 3, CD147,
phosphorylated-ERK1/2, cyclin D1, cyclin A and cyclin E were
markedly increased in thoracic aortas with the progression of
remodeling. The results of the present study indicated that the
CD147-ERK1/2-cyclin pathway was activated and upregulated with the
process of remodeling of the rat thoracic aorta. Treatment with
simvastatin for 4, 8 and 12 weeks significantly diminished these
alterations. The results of the present study implied that
simvastatin may improve remodeling of the rat thoracic aorta by
diminishing the upregulation and activation of the
CD147-ERK1/2-cyclin pathway.
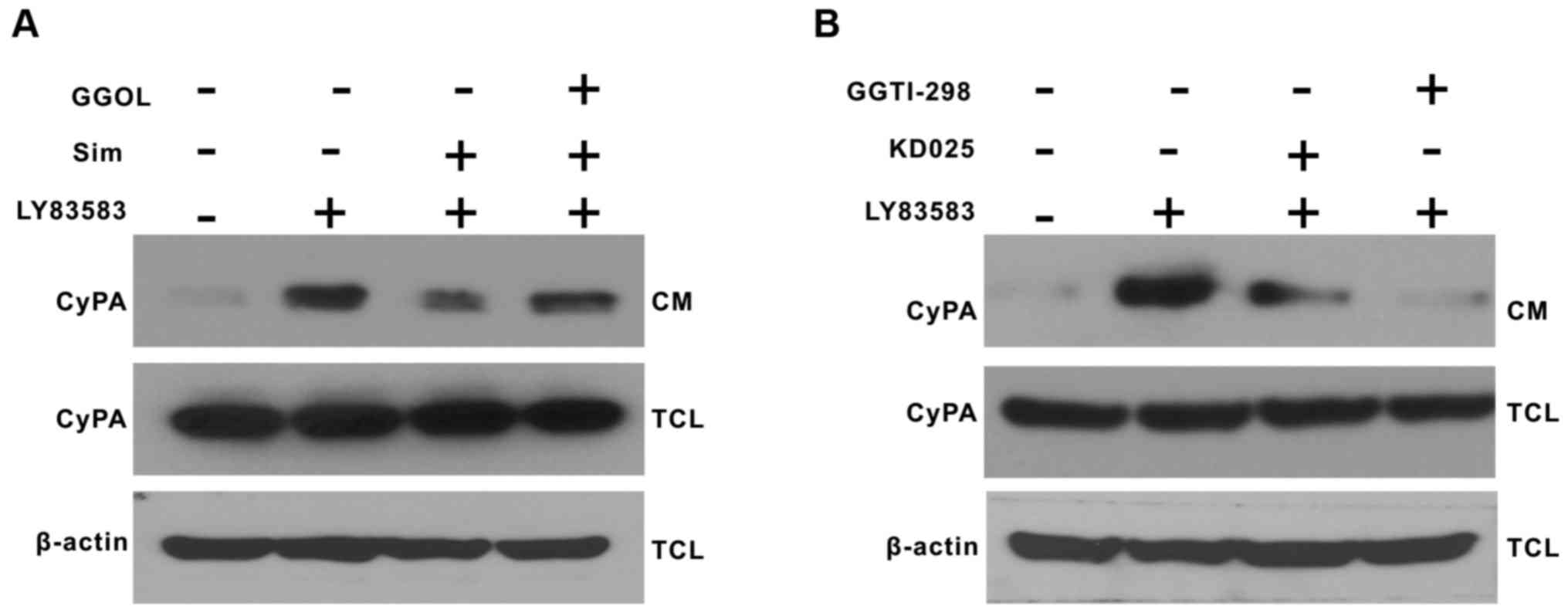
Rho-ROCK2 pathway mediates the
inhibitory action of simvastatin on CyPA secretion induced by
LY83583 in VSMCs
Previous studies have suggested that the pleiotropic
effects of statins may be mediated through inhibition of the
Rho/Rho kinase pathway (16,21).
In order to further demonstrate whether Rho-ROCK2 mediates CyPA
secretion from VSMCs, and whether simvastatin inhibits CyPA
secretion by inhibiting the Rho-ROCK2 kinase pathway. VSMCs were
pretreated with GGOL (to stimulate protein geranylgeranylation),
KD025 (a selective ROCK2 inhibitor) and GGTI-298 (a geranylgeranyl
transferase-I inhibitor) for 30 min prior to the addition of 1
µmol/l LY83583 for 2 h. Pretreatment with GGOL partially reversed
the inhibitory effect of simvastatin on CyPA secretion induced by
LY83583 (Fig. 4A). KD025 and
GGTI-298 inhibited CyPA secretion, suggesting that Rho and ROCK2
may mediate CyPA secretion (Fig.
4B). The results of the present study further elucidated that
the Rho-ROCK2 pathway may partially mediate the inhibitory effect
of simvastatin on CyPA secretion induced by LY83583.
Discussion
The primary findings of the present study were that
the administration of simvastatin significantly decreased the serum
levels of CyPA and ameliorated the remodeling of thoracic aortas in
hypertensive rats. Upregulation of the CD147-ERK-cyclin signaling
pathway in remodeled thoracic aortas was alleviated by treatment
with simvastatin. The Rho-ROCK2 pathway partly mediated
simvastatin-inhibited CyPA release from cultured VSMCs.
CyPA was previously demonstrated to promote vascular
remodeling by inducing VSMC proliferation (6), and LY83583-induced CyPA secretion was
dose-dependently inhibited by simvastatin in cultured VSMCs
(14). It was previously
demonstrated that serum CyPA levels were gradually increased with
the development of hypertension in rats (7), and that the levels were positively
correlated with systolic and diastolic blood pressure in untreated
patients (8). Therefore, the
present study further investigated whether serum CyPA levels may be
influenced by treatment with simvastatin in hypertensive rats. The
results of the present study demonstrated that CyPA levels were
upregulated with the process of thoracic aorta remodeling, and that
treatment with simvastatin significantly decreased the serum CyPA
levels and improved thoracic aorta remodeling during hypertension.
Through detection of CyPA secretion from primary VSMCs from the
sham, hypertensive and simvastatin group at 12 weeks, in response
to LY83583, it was observed that the CyPA secretion of primary
VSMCs from the simvastatin group was decreased compared with that
from the hypertensive group. This result illustrated that CyPA
secretion from VSMCs was enhanced by hypertension, while this
hypertension-induced CyPA secretion was inhibited by treatment with
simvastatin. Since CyPA contributes to vascular remodeling
(6), it is possible that
simvastatin may improve thoracic aorta remodeling during
hypertension via the inhibition of CyPA release from VSMCs and the
downregulation of serum CyPA levels.
CyPA has been suggested to mediate vascular
remodeling by stimulating ERK1/2, VSMC growth and inflammation
(5,6). Secreted CyPA was recently reported to
mediate the G1/S phase transition of cholangiocarcinoma cells via
the CD147-ERK1/2 pathway (10).
Therefore, the present study further investigated alterations to
downstream signaling molecules and the influence of simvastatin in
the thoracic aorta. Western blotting demonstrated that CD147,
phosphorylated-ERK1/2 and certain cyclins (cyclin D1, cyclin E and
cyclin A) were time-dependently upregulated with the development of
thoracic aorta remodeling. Treatment with simvastatin decreased the
upregulation of CD147, phosphorylated-ERK1/2 and cyclins in the
thoracic aorta, and alleviated thoracic aorta remodeling during
hypertension. VSMC proliferation is controlled by various cell
cycle proteins. In particular, cyclin D1, cyclin E and cyclin A
induce cell cycle progression from the G1 phase to the S phase
(22), and VSMC proliferation
following angioplasty in the rat carotid artery is associated with
a temporally- and spatially-coordinated expression of cyclin E,
cyclin A and proliferating cell nuclear antigen (23). These results indicated that
simvastatin may improve thoracic aorta remodeling during
hypertension in part by impeding the upregulation of CD147,
phosphorylated-ERK1/2 and cyclins in thoracic aortas.
Due to the downregulation of serum CyPA levels by
simvastatin, the present study examined the underlying mechanisms
through which simvastatin may decreased CyPA release from VSMCs.
GGOL, which promotes Rho geranylgeranylation and the subsequent
activation of Rho kinase, prevented the inhibitory effect of
simvastatin on CyPA secretion. GGTI-298 (a geranylgeranyl
transferase-I inhibitor) and KD025 (a selective ROCK2 inhibitor)
inhibited CyPA secretion in VSMCs. The results of the present study
demonstrated that activation of the Rho-ROCK2 pathway promoted CyPA
secretion from VSMCs, while inhibition of the Rho-ROCK2 pathway
diminished CyPA secretion, and that simvastatin decreased CyPA
secretion in VSMCs by inhibiting the Rho-ROCK2 pathway.
In conclusion, serum CyPA concentration and
CD147-ERK1/2-cyclins were time-dependently upregulated with the
development of thoracic aorta remodeling, whereas treatment with
simvastatin decreased the upregulation of the CD147-ERK1/2-cyclin
pathway in thoracic aortas and CyPA secretion from VSMCs. The
Rho-ROCK2 pathway mediated the inhibitory effect of simvastatin on
CyPA secretion from VSMCs.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81302771),
the Natural Science Foundation of Guangdong Province (grant no.
2014A030313087), the Science and Technology Program of Guangzhou
City (grant no. 201607010255), the Medical Scientific Research
Foundation of Guangdong Province, China (grant no. A2016382), the
National College Students' Innovation Entrepreneurship Training
Plan Program of China (grant no. 201410570003), the College
Students' Science and Technology Innovation Cultivation Special
Funds Program of Guangdong Province (grant no. pdjh2015b0438), the
Guangzhou Medical University College Students Science Technology
Innovation Project (grant no. 2013A0014), the Scientific Research
Foundation of Guangzhou Medical University and the Foundation for
Excellent Teachers by Guangzhou Medical University, China.
References
|
1
|
Berk BC: Vascular smooth muscle growth:
Autocrine growth mechanisms. Physiol Rev. 81:999–1030.
2001.PubMed/NCBI
|
|
2
|
Dobrian A, Wade SS and Prewitt RL: PDGF-A
expression correlates with blood pressure and remodeling in 1K1C
hypertensive rat arteries. Am J Physiol. 276:H2159–H2167.
1999.PubMed/NCBI
|
|
3
|
Naftilan AJ, Pratt RE and Dzau VJ:
Induction of platelet-derived growth factor A-chain and c-myc gene
expressions by angiotensin II in cultured rat vascular smooth
muscle cells. J Clin Invest. 83:1419–1424. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang H, Li M, Chai H, Yan S, Lin P,
Lumsden AB, Yao Q and Chen C: Effects of cyclophilin A on cell
proliferation and gene expressions in human vascular smooth muscle
cells and endothelial cells. J Surg Res. 123:312–319. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jin ZG, Melaragno MG, Liao DF, Yan C,
Haendeler J, Suh YA, Lambeth JD and Berk BC: Cyclophilin A is a
secreted growth factor induced by oxidative stress. Circ Res.
87:789–796. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Satoh K, Matoba T, Suzuki J, O'Dell MR,
Nigro P, Cui Z, Mohan A, Pan S, Li L, Jin ZG, et al: Cyclophilin A
mediates vascular remodeling by promoting inflammation and vascular
smooth muscle cell proliferation. Circulation. 117:3088–3098. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tang FC, Wang HY, Ma MM, Guan TW, Pan L,
Yao DC, Chen YL, Chen WB, Tu YS and Fu XD: CyPA-CD147-ERK1/2-cyclin
D2 signaling pathway is upregulated during rat left ventricular
hypertrophy. Sheng Li Xue Bao. 67:393–400. 2015.PubMed/NCBI
|
|
8
|
Chang CS, Su SL, Chang CC, Lee KW, Kuo CL,
Huang CS, Tseng WM and Liu CS: Cyclophilin-A: A novel biomarker for
untreated male essential hypertension. Biomarkers. 18:716–720.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yurchenko V, Zybarth G, O'Connor M, Dai
WW, Franchin G, Hao T, Guo H, Hung HC, Toole B, Gallay P, et al:
Active site residues of cyclophilin A are crucial for its signaling
activity via CD147. J Biol Chem. 277:22959–22965. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Obchoei S, Sawanyawisuth K, Wongkham C,
Kasinrerk W, Yao Q, Chen C and Wongkham S: Secreted cyclophilin A
mediates G1/S phase transition of cholangiocarcinoma cells via
CD147/ERK1/2 pathway. Tumour Biol. 36:849–859. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Goldstein JL and Brown MS: Regulation of
the mevalonate pathway. Nature. 343:425–430. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Riento K and Ridley AJ: Rocks:
Multifunctional kinases in cell behaviour. Nat Rev Mol Cell Biol.
4:446–456. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zanin-Zhorov A, Weiss JM, Nyuydzefe MS,
Chen W, Scher JU, Mo R, Depoil D, Rao N, Liu B, Wei J, et al:
Selective oral ROCK2 inhibitor down-regulates IL-21 and IL-17
secretion in human T cells via STAT3-dependent mechanism. Proc Natl
Acad Sci USA. 111:pp. 16814–16819. 2014; View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Suzuki J, Jin ZG, Meoli DF, Matoba T and
Berk BC: Cyclophilin A is secreted by a vesicular pathway in
vascular smooth muscle cells. Circ Res. 98:811–817. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ge CJ, Hu SJ, Wu YS and Chen NY: Effects
of atorvastatin on vascular remodeling in spontaneously
hypertensive rats. J Zhejiang Univ Sci. 4:612–615. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma MM, Li SY, Wang M and Guan YY:
Simvastatin attenuated cerebrovascular cell proliferation in the
development of hypertension through Rho/Rho-kinase pathway. J
Cardiovasc Pharmacol. 59:576–582. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Freeman EJ, Chisolm GM, Ferrario CM and
Tallant EA: Angiotensin-(1–7) inhibits vascular smooth muscle cell
growth. Hypertension. 28:104–108. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Short D: Morphology of the intestinal
arterioles in chronic human hypertension. Br Heart J. 28:184–192.
1966. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wiener J, Loud AV, Giacomelli F and
Anversa P: Morphometric analysis of hypertension-induced
hypertrophy of rat thoracic aorta. Am J Pathol. 88:619–634.
1977.PubMed/NCBI
|
|
20
|
Pan L, Yao DC, Yu YZ, Li SJ, Chen BJ, Hu
GH, Xi C, Wang ZH, Wang HY, Li JH and Tu YS: Necrostatin-1 protects
against oleic acid-induced acute respiratory distress syndrome in
rats. Biochem Biophys Res Commun. 478:1602–1608. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Laufs U, Kilter H, Konkol C, Wassmann S,
Böhm M and Nickenig G: Impact of HMG CoA reductase inhibition on
small GTPases in the heart. Cardiovasc Res. 53:911–920. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sherr CJ: G1 phase progression: Cycling on
cue. Cell. 79:551–555. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wei GL, Krasinski K, Kearney M, Isner JM,
Walsh K and Andrés V: Temporally and spatially coordinated
expression of cell cycle regulatory factors after angioplasty. Circ
Res. 80:418–426. 1997.PubMed/NCBI
|