Introduction
Worldwide, liver cirrhosis is considered an
important global health burden, with more than one million deaths
in 2010 (1). Liver cirrhosis and
fibrosis are chronic inflammatory diseases in which excessive wound
healing is associated to multiple etiologies such as alcohol abuse,
metabolic disorders, immune disease and viral infections (2,3).
They are characterized by over deposition of extracellular matrix
(ECM), with reduced expression of matrix metalloproteinases (MMPs)
and increased expression of tissue inhibitor of metalloproteinases
(TIMPs), and the consequent loss of liver functions (4,5).
Fibrotic tissue increases the regenerative nodules and the
distortion of the vascular function is responsible for most of the
complications in this liver disease (6). The activation of hepatic stellate
cells (HSCs) to the miofibroblasts phenotype plays a remarkable
role during the pathogenesis of fibrosis and the establishment of
liver cirrhosis (7). Growing
evidence is demonstrating that inhibition of transforming growth
factor (TGF)-β, an activator of HSCs, shows promising effects in
the treatment of liver fibrosis (7–9).
TGF-β is considered as a crucial profibrogenic cytokine involved in
the fibroblasts recruitment, differentiation of miofibroblasts,
epithelial-mesenchymal transition (EMT) and deposition of ECM
(10,11).
Some drugs and pathways are being used as target for
the blockage of fibrotic process, like angiogenesis inhibitors,
antihypertensive drugs as angiotensin inhibitors, downregulation of
TGF-β, and cell therapy strategies (12). Recently doxazosin, an α-blocker of
adrenergic receptors, has been proposed as a potential antifibrotic
drug in an experimental model of cirrhosis induced with carbon
tetrachloride (CCl4). After 6 weeks of treatment with doxazosin, a
significant reduction in deposits of collagen type I was observed
in cirrhotic livers, which was associated with a decrease in TGF-β
synthesis (13). Nevertheless,
doxazosin antifibrotic effect at shorter periods of treatment has
not been studied on cirrhotic animals. Doxazosin is usually used to
treat patients with hypertension by avoiding the norepinephrine
binding to α1-adrenergic receptors (14). Moreover, doxazosin relieves benign
prostatic hypertrophy symptoms through the relaxing of the smooth
muscles surrounding the prostate, so that it can reduce the urine
retention and diminish bladder outlet obstruction (15).
In addition to reducing the complications in
cirrhotic patients, new treatments must be focused on ceasing the
progression of cellular damage, by promoting the regeneration of
parenchyma and the inhibition of fibrotic process. Gene therapy
coupled with standard drugs is a promising and viable alternative
to alleviate liver chronic diseases in order to recover
hepatocellular functions, improve portal pressure and arrest the
progression of the liver fibrosis (7,16).
Bone morphogenetic proteins (BMPs) are members of related proteins
that belong to the superfamily of TGF-β with 47 identified members
(17). BMPs have been described as
excellent inducers of bone and cartilage, due to their capacity to
induce cellular differentiation through cell signaling after
receptor binding. BMPs also play pleiotropic roles in various
biological processes in digestive tube, heart, brain, gonads, liver
and skin (18–21). After binding to their receptors, a
downstream Smad signaling is triggered with the consequent
activation of target genes, mainly related to cellular
proliferation and differentiation (22,23).
It has been reported that BMP-7 causes the inhibition of the
intracellular signaling initiated by TGF-β, through the blockage of
Smad 3/4, and diminishes the effects of TGF-β (24–26).
In absence of BMP-7, TGF-β binding to its receptor initiates the
phosphorylation of Smad-2 and Smad-3 proteins, which join Smad-4 to
form a complex which is translocated to the nucleus and activates
the collagen type I synthesis and EMT activation. Otherwise, BMP-7
downstream signaling via activation of Smad-1, Smad-5 and Smad-8
can antagonize the effect of Smad-2/-3 through the
heterodimerization with Smad-4, thereby Smad-3 is accumulated
outside the nucleus (27,28). Several studies show that
recombinant BMP-7 protein administered to animals reduces the
fibrosis in kidney, lung and liver, by downregulating fibrotic
markers such as collagen, α-smooth muscle actin (SMA) and TGF-β
expression (29–32).
In this study, we analyzed the effect of BMP-7 gene
therapy alone or in co-treatment with doxazosin in a model of liver
cirrhosis in hamster. Our results suggest that the strategy
combining BMP-7 gene therapy and doxazosin is useful for the
reversion of liver cirrhosis.
Materials and methods
Animals
Six-to eight-weeks old male golden hamsters
(Mesocricetus auratus), from 100 to 150 g weight, were
maintained in the local Animal Core Facility with 12 h light/dark
cycles and fed with Nutricubos (Ralston Purina Company, St. Louis,
MO, USA) and water ad libitum. All animal experiments were
approved by the Animal Welfare and Research Ethics Committee of the
Autonomous University of Aguascalientes (Aguascalientes, Mexico),
and were conducted in accordance with institutional guidelines for
caring for experimental animals and the national regulatory norm
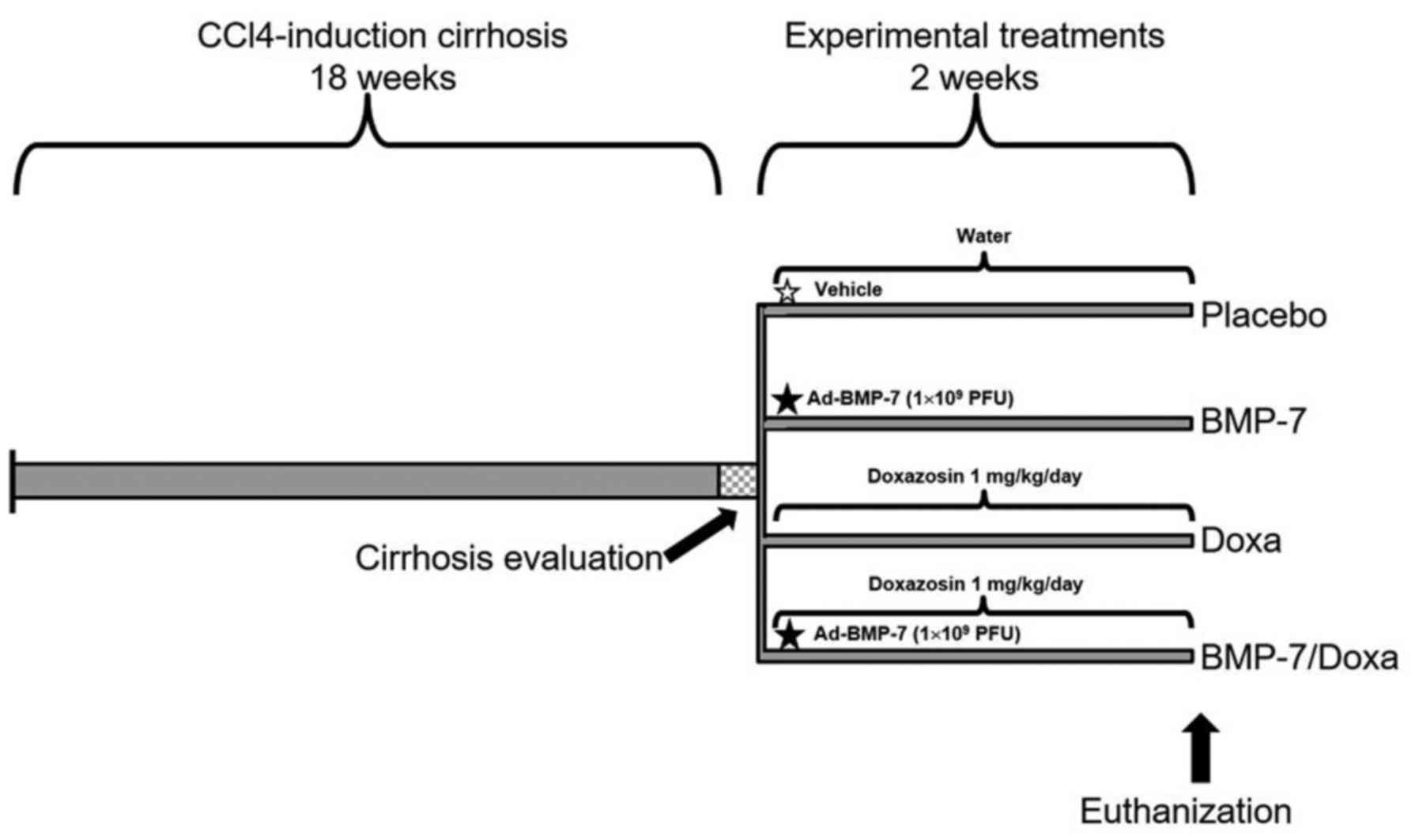
NOM-062-Z00-1999. Thirty animals were randomly distributed as
follows: i) intact hamsters (n=5) (Intact group) and ii)
experimental hamsters induced with cirrhosis by two intraperitoneal
injections of 50 mg/kg of CCl4 per week for 18 weeks (n=25). Intact
animals and five experimental hamsters (Cirrhotic group) were
sacrificed at week 18 to demonstrate the fibrosis/cirrhosis grade,
collecting blood and liver samples. Photographs in two positions,
liver anterior region and portal vein were documented with an
Olympus xD Master 2 (SP-55OUZ; Olympus Corporation, Tokyo,
Japan).
Adenoviral-BMP-7 transduction and
doxazosin therapies
Experimental hamsters were divided in four groups
with 5 animals per group: i) Placebo, treated orally with water and
via portal with a single injection of adenoviral storage buffer as
vehicle (10 mM Tris, Ph 8.0, 100 mM NaCl, and 50% glycerol); ii)
Doxa, orally treated with doxazosin 1 mg/kg/day; iii) BMP-7:
treated via portal with a single injection of 1×109 PFU
adenoviral vector Ad-BMP-7, as previously reported (33), and iv) BMP-7/Doxa, treated with the
same doses and times that BMP-7 and Doxa groups. All treatments
were administered for 2 weeks, once cirrhosis was established, and
later animals were euthanized for specimen collection of liver and
blood. Liver photographs were taken and tissue samples were fixed
in neutral formalin. Serum samples were frozen at −80°C until used.
The experimental protocol and treatment schedules for each group
are illustrated in Fig. 1.
Protein extraction and Western blot
analysis
To analyze the expression of BMP-7 and collagen type
I in the liver parenchyma, 100 mg of tissue was homogenized in 1 ml
of lysis buffer (Tris-HCl 50 mM pH 6.8, N-ethylmaleimide 5 mM,
iodoacetamide 3 mM, phenylmethanesulfonyl fluoride 1 mM and
tosyl-L-lysine chloromethyl ketone 3 mM) for total protein
extraction. The lysate was centrifuged at 40,000 × g for 1 h at
4°C. The supernatant was recovered and proteins quantified with the
Bradford method (34).
For western blot, 50 µg of each protein extract was
separated in a 12% SDS-PAGE gel, and proteins were transferred to a
PVDF membrane (Bio-Rad, Hercules, CA, USA). Blockage was achieved
for 1 h at room temperature with TBST (Tris-buffered saline/0.05%
Tween-20) and 5% skimmed milk. For immunodetection, membrane was
incubated for 1 h at room temperature with: 0.2 µg/ml goat
polyclonal anti BMP-7 (Santacruz Biotechnology, Dallas, Tx, USA), 1
µg/ml mouse monoclonal anti collagen type I (Sigma-Aldrich, St.
Louis, MO, USA) or 1 µg/ml rabbit polyclonal anti β-actin (Abcam,
Cambridge, UK). Then, blots were incubated with the alkaline
phosphatase (AP)-conjugated antibodies: 1 µg/ml rabbit anti-goat
(Merk-Millipore, Darmstadt, GE), 0.25 µg/ml goat anti-mouse
(Sigma-Aldrich), or 0.5 µg/ml goat anti-rabbit (Abcam),
respectively. After three washes with TBS, blotting was developed
with Sigma Fast BCIP®/NTB (Sigma-Aldrich). Analysis of
collagen type I, BMP-7 and β-actin expression was performed with
the ImageJ software (National Institutes of Health, Bethesda, MD,
USA). Mean of Intensity of collagen type I and BMP-7 bands were
normalized respect to that of β-actin, as internal control and
ratios BMP-7/collagen type I were calculated.
Liver function tests
The serum markers of liver damage: alanine
aminotransferase (ALT), aspartate aminotransferase (AST) and
albumin were quantified in an Biosystems analyzer (bts-350;
Biosystems, Quezon City, Philippines).
Histochemical and Immunohistochemical
analysis
Tissues were paraffin-embedded and 5 µm sections
were obtained and stained with hematoxylin and eosin (H&E) and
Sirius red techniques to evaluate parenchymal architecture and
deposits of collagen type I or III as result of ECM rearrangement.
Percent of fibrotic area was determined with the measure of
fibrotic area in relation to the total area of the tissue.
Morphometric analysis was performed with Image Pro Plus Software
4.5.1 (Media Cybernetics, Bethesda, MD, USA). Immunohistochemistry
was performed to analyze BMP-7- and α-SMA-positive cells in liver
tissues. Briefly, liver tissue slides were incubated with 2 µg/ml
goat polyclonal anti-BMP-7 (Santa Cruz Biotechnology) or 10 µg/ml
rabbit polyclonal anti-α-SMA (Abcam) for 12 h at 4°C. As a
secondary antibody, 4 µg/ml rabbit anti-goat (Merk-Millipore)
conjugated with AP or EnVision+/Dual Link System-HRP anti
Rabbit/Mouse (Dako, Carpinteria, CA, USA), was respectively used.
The AP activity for BMP-7 detection was developed with the Sigma
Fast BCIP®/NTB (Sigma-Aldrich), and the HRP activity for
α-SMA detection with Sigma Fast Diaminobenzidine (Sigma-Aldrich).
Number of α-SMA-positive cells were counted in all slide and
reported as cells/mm2. Data was documented in a Zeiss
Axioscope 40/40FL microscope (Zeiss, Oberkochen, Germany) and
analyzed with the Image Pro Plus Software 4.5.1 (Media
Cybernetics).
Isolation of total RNA and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from 100 µg of liver of the
control and experimental animals with the SV Total RNA Isolation
System (Promega, Madison, WI, USA) following the manufacturer's
protocol. Total RNA was stored at −80°C until use. Reverse
transcription for cDNA synthesis was performed with 1 µg of total
RNA using the GoScript Reverse Transcription System (Promega) and
real time qPCR with the buffer GreenMaster with UNG-clear (Jena
Bioscience, Jena, Germany) in a StepOne Real-Time PCR Machine
(Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Oligonucleotides were designed to target on MMP-13, TIMP-2, and
actin as a reference control (Table
I). The RT-PCR program used for all targets was: 50°C for 2
min, 95°C for 45 sec, 40 cycles of 95°C for 45 sec and 60°C for 45
sec, and extension at 72°C for 12 sec. Relative quantification of
target mRNA expression was calculated by the ∆∆Ct method using
β-actin for normalization (35).
 | Table I.Oligonucleotides sequences. |
Table I.
Oligonucleotides sequences.
| Gene |
Oligonucleotides | Accession
number | Amplicon size |
|---|
| MMP-13 |
| XM_005077319.1 | 246 pb |
| F |
TGTCCTGGCCACTCTCTTCT |
|
|
| R |
GGGTCATCAAGTTTGCCAGT |
|
|
| TIMP-2 |
| AF260255.1 | 196 pb |
| F |
TCAAAGGCCCTGACAAAGAC |
|
|
| R |
AGGCTCTTCTTCTGGGTGGT |
|
|
| β-actin |
| NM_001281595.1 | 115 pb |
| F |
GCCCAGAGCAAGAGAGGTAT |
|
|
| R |
CACGCAGCTCGTTGTAGAAG |
|
|
Statistical analysis
GraphPad Prism V5 software (GraphPad Software, Inc.,
La Jolla, CA, USA) was employed for statistical analysis. Data are
expressed as the mean ± standard error of the mean (SEM) of five
animals. Significant differences between mean values were evaluated
by using the two tailed Student's t test. Statistical significance
was considered at P<0.05.
Results
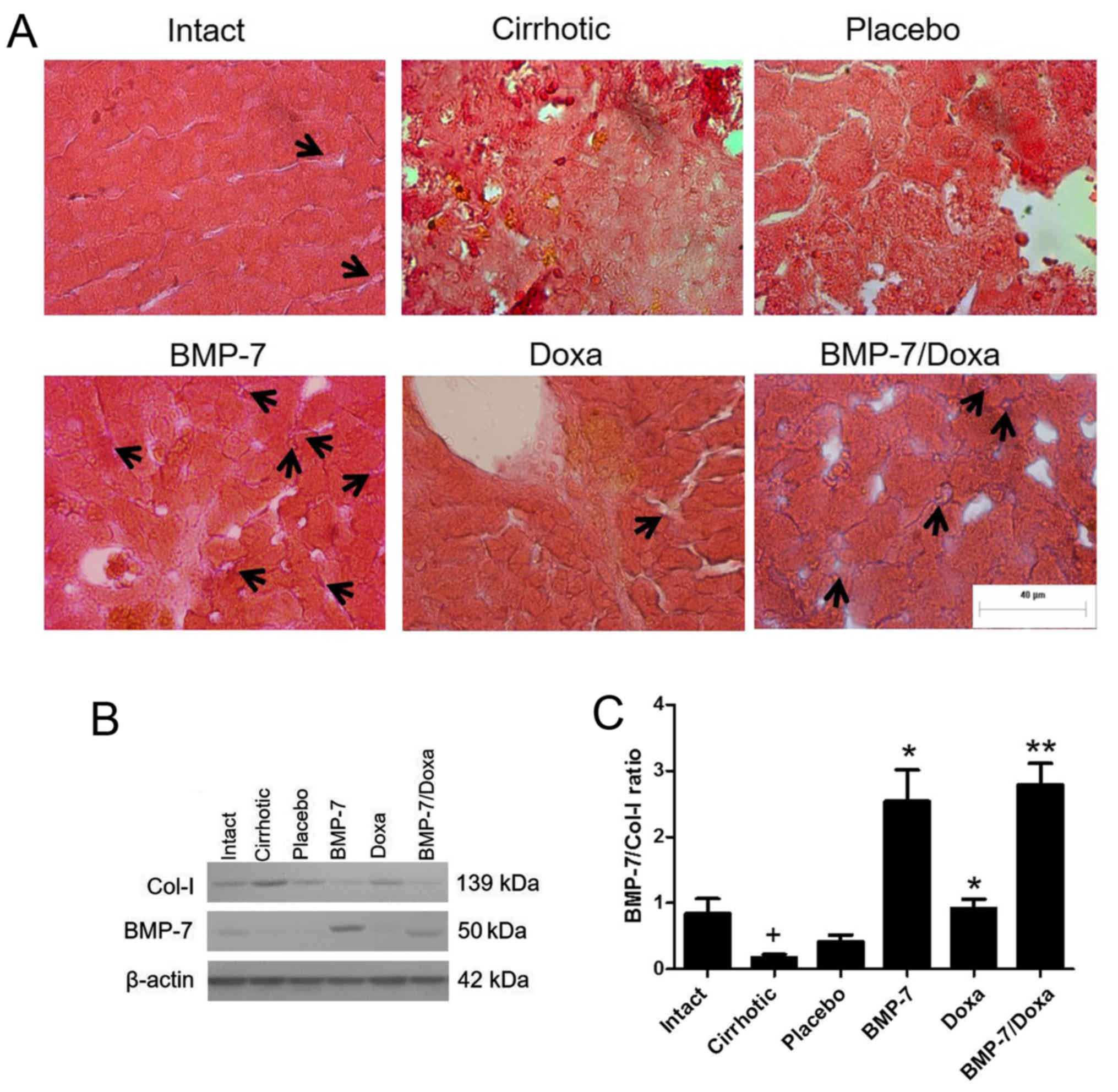
Expression of BMP-7 and its relationship with
collagen type I levels in hepatic parenchyma of transduced
CCl4-induced cirrhosis hamsters. The administration of
1×109 PFU of the vector via porta was efficiently
tolerated by animals. To confirm protein expression of BMP-7 in the
hepatic parenchyma of transduced animals, immunohistochemistry and
Western blot was developed in liver tissue (Fig. 2). Immunohistochemical examination
showed BMP-7 secreting hepatocytes in liver from BMP-7 and
BMP-7/Doxa animals (Fig. 2A, black
arrows). BMP-7 native production was also detected in Intact and
Doxa groups, and remains almost undetectable in Placebo and
Cirrhotic animals. Similar results were obtained by western blot
analysis, as seen in Fig. 2B.
Although basal expression of BMP-7 was present in Intact,
Cirrhotic, Placebo and Doxa groups, the expression of the protein
was much higher in the groups BMP-7 and BMP-7/Doxa after 2 weeks of
adenoviral delivery. Also, BMP-7 expression was correlated with
collagen type I production, since an increase in BMP-7 levels in
BMP-7 and BMP-7/Doxa groups was associated with a decrease in the
levels of collagen type I (Fig.
2B). The densitometric analysis revealed that cirrhotic animals
decreased BMP-7/Col-I ratio, but vector administration
significantly increased 6.1- and 6.7-fold BMP-7/Col-I ratio in
BMP-7 and BMP-7/Doxa groups, respect to Placebo animals (Fig. 2C). When doxazosin was administered
alone, although collagen type I expression was similar to Placebo
group, there was also an increase in BMP-7/Col-I ratio in these
animals that was 2-fold higher that in Placebo group.
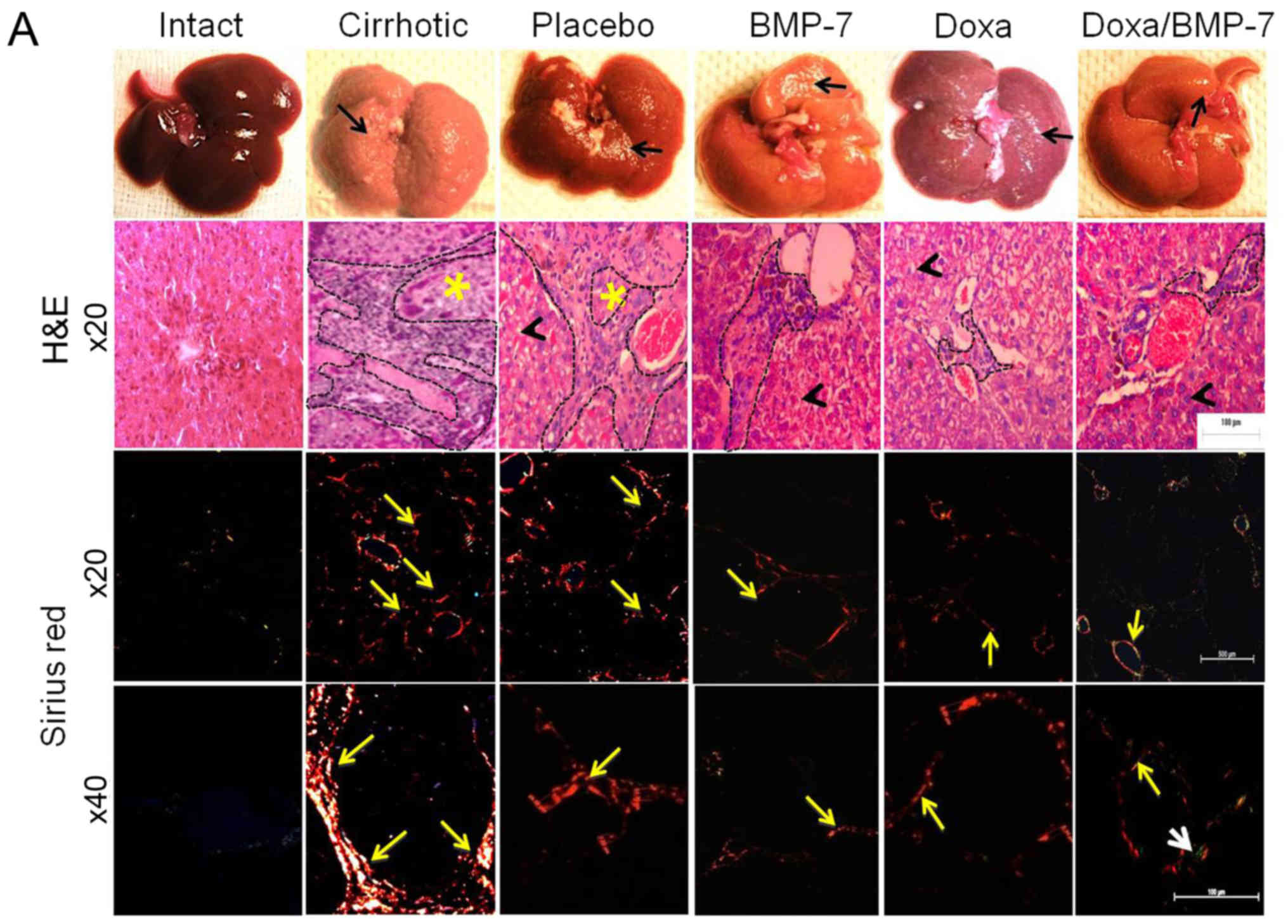
Regression of cirrhosis with
α-adrenergic receptor antagonist and therapy with BMP-7
The histological changes and accumulation of
collagen type I were evaluated by studying liver architecture and
collagen type I deposition by H&E and Sirius red staining under
light microscopy and polarized-light microscopy (Fig. 3A). The macroscopic observation of
Intact group livers evidenced normal aspect, shape and dark-browned
color. The H&E staining showed the liver parenchyma well
organized in acinus with a central vein. Moreover, Sirius red
staining did not reveal collagen type I accumulation at
magnification, ×20 and ×40. In the Cirrhotic group, livers had
granulated surface (black arrow); the H&E images showed a wide
area of inflammatory infiltrate and possible fibrosis (black dashed
line) that delimited the regenerative nodules (yellow asterisk).
Sirius red staining evidenced several circles of collagen type I
surrounding regenerative nodules (×20, yellow arrow), in which
collagen type I accumulated in the periphery (×40, yellow arrow).
The livers in the Placebo group were similar to the Cirrhotic
group, with granulated surface (black arrow). We observed an
inflammatory infiltrate (black dashed outline) in the tissue
architecture, and hepatocytes appeared edematous (black arrowhead).
After observation at magnification, ×20 and ×40 with Sirius red
staining, liver slides had lesser aggregation of collagen type I
fibers (yellow arrows) when compared to the Cirrhotic group. Livers
from BMP-7 group had reduced granulation on their surface (black
arrow), liver parenchyma showed less inflammatory infiltrate (black
dashed line) surrounding hepatocytes with notable reduction of
edema (black arrowhead), and reduced fibers of collagen type I
(yellow arrows). Doxazosin-treated cirrhotic hamsters had little
granulation on the liver surface (black arrow); like in the BMP-7
group, scarce inflammatory infiltrate (black dashed line) and some
edematous hepatocytes were observed (black arrowhead), and collagen
type I fibers (yellow arrows) were markedly reduced when compared
to Placebo group. In the group that received the double treatment
(BMP-7/Doxa), the surfaces of the livers were barely granulated
(black arrow), liver parenchyma showed few inflammatory infiltrate
(black dashed line) surrounding morphologically healthy hepatocytes
(black arrowhead). Outstandingly, the observation of the Sirius red
staining showed scarce deposits of collagen type I (yellow arrows)
and merging fibers of collagen type III (white arrow).
The measurement of the fibrotic area clearly
demonstrated a significant 7.8-fold increment of collagen type I
deposits in liver of cirrhotic animals. Placebo group showed a
spontaneous 33.6% decrease of these collagen deposits. However, the
treatment with adenoviral BMP-7-transduction induced a reduction of
51.6% of collagen type I deposits, and Doxa group had 47.1% reduced
collagen type I accumulation, when compared to Placebo group. The
percent of the fibrotic area was reduced 37.4% in cirrhotic hamster
treated with doxazosin and BMP-7 in comparison to the Placebo group
(Fig. 3B). BMP-7 did not improve
the antifibrogenic effect of doxazosin, as there were not
differences between Doxa and BMP-7/Doxa groups. These data
demonstrated that both adenoviral-BMP-7 gene and doxazosin
treatment are potential candidates for decreasing liver fibrosis,
although combination of both therapies do not improve their
individual antifibrotic effect once cirrhosis is stablished.
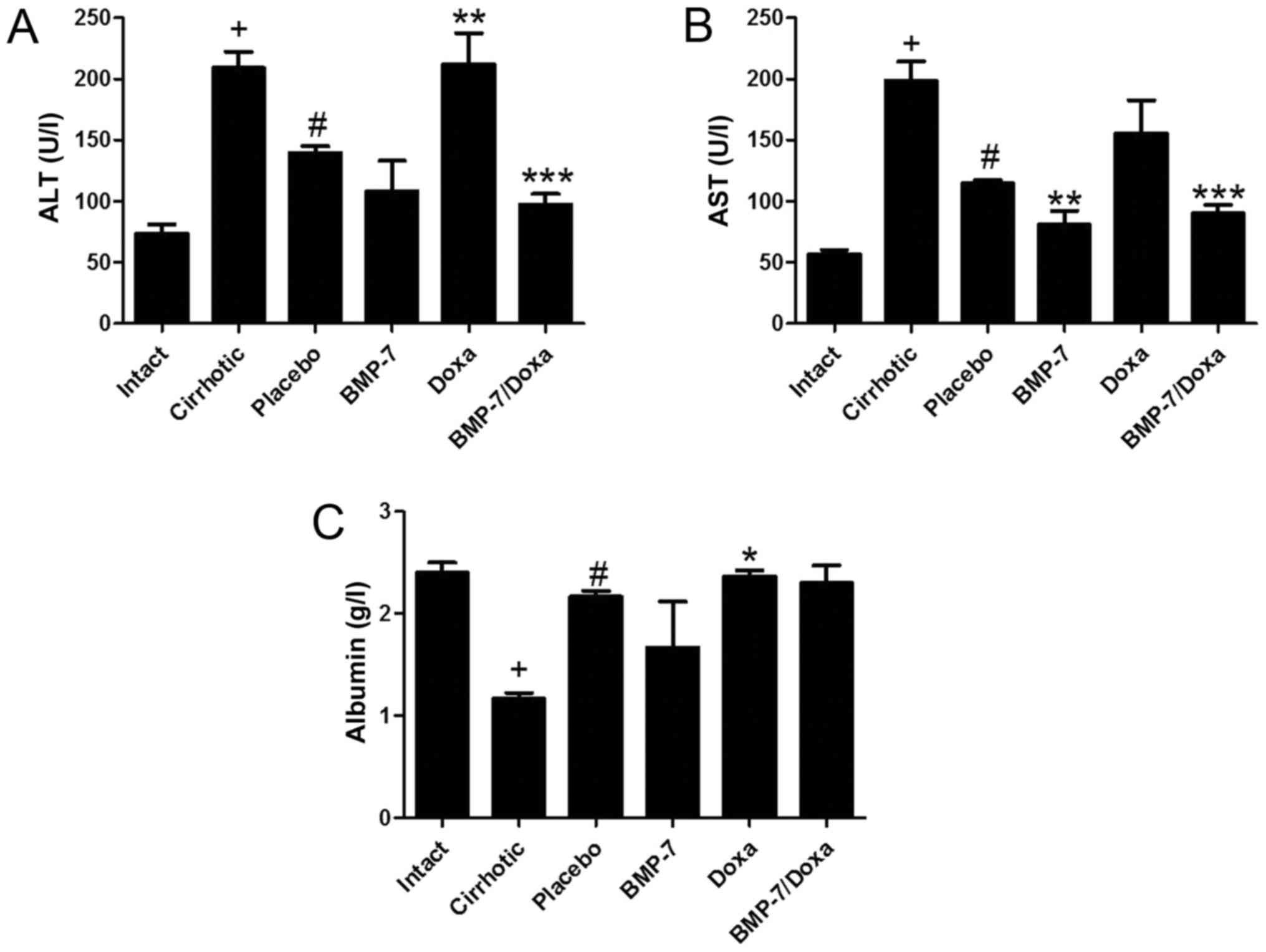
Recovery of liver function in BMP-7
transduced and/or Doxa treated animals
The plasma levels of liver enzymes ALT and AST, and
albumin were measured to determine the liver function in
experimental groups (Fig. 4).
Liver function tests supported the histological findings in 50
mg/kg CCl4-induced hamsters treated with BMP-7 and BMP-7/Doxa. As
expected, the cirrhotic group had significantly increased levels of
ALT and AST, and diminished concentration of albumin when compared
to Intact group. Animals from Placebo group showed a spontaneous
recovery of these biochemical markers of hepatic damage, as they
decrease in 33.2 and 42.3% the levels of ALT and AST and increase
in 86.2% albumin concentration in blood. In the BMP-7 group,
adenoviral-BMP-7 transduction significantly lowered the levels of
AST (29.6%) as compared to the Placebo group, although ALT and
albumin remained without significant changes. However, after two
weeks of doxazosin treatment (Doxa group), ALT levels and albumin
were 34.1 and 6% higher than in the Placebo group, respectively,
which presumably indicates hepatotoxic activity of doxazosin.
Moreover, adenoviral-BMP-7 transduction and doxazosin treatment
induced significant diminution of ALT and AST levels, while
concentration of albumin was unchanged, as compared to Placebo
group. These results suggest that BMP-7 over-expression combined
with doxazosin may reduce the biochemical abnormalities after
hepatic damage induced with CCl4, even when two weeks of doxazosin
treatment appear to induce toxic effects in the cirrhotic
liver.
Analyses of hepatic fibrosis by
immunohistochemistry for HSCs
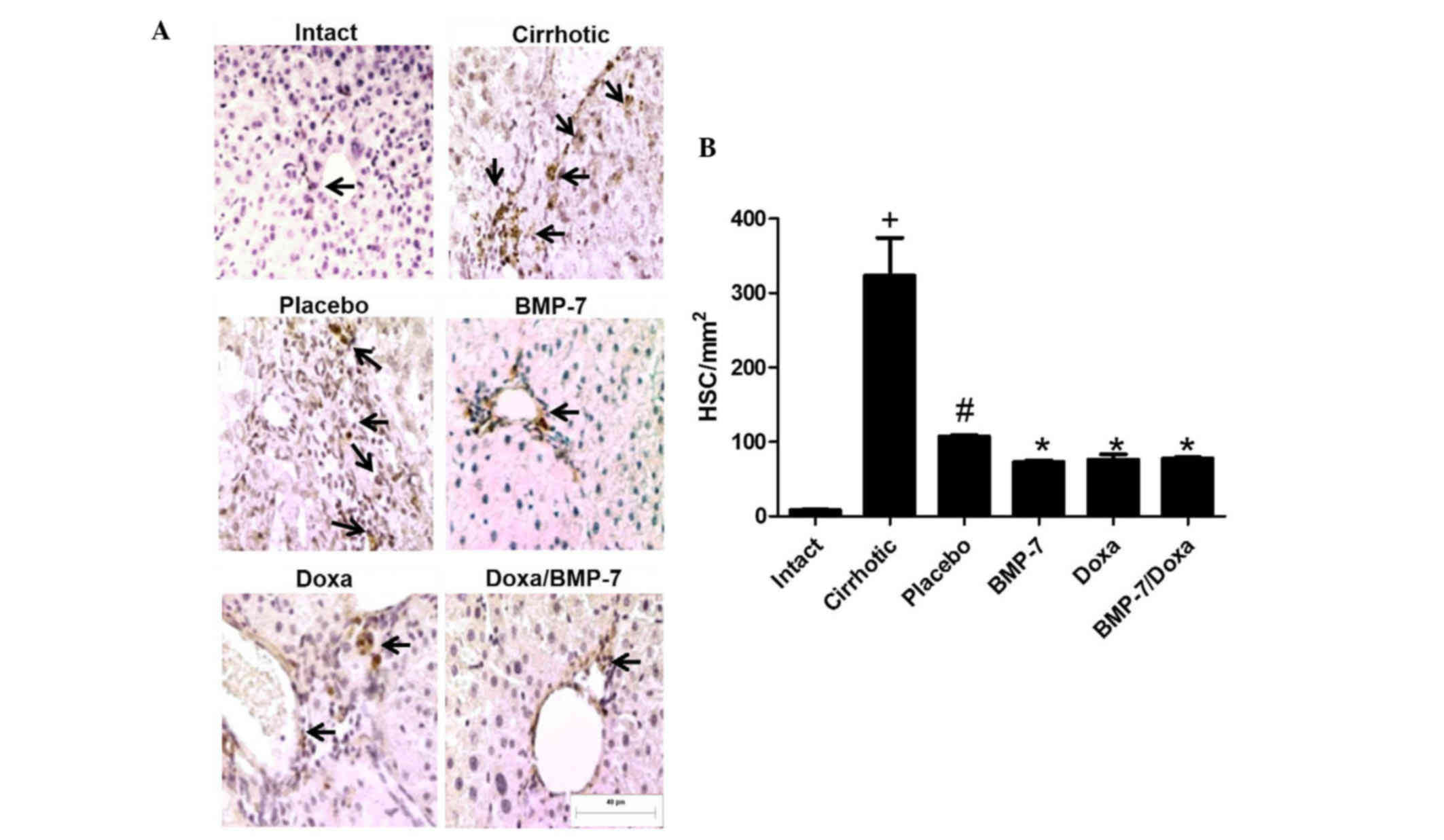
To correlate changes in number of activated HSCs to
the fibrosis reduction, we analyzed the number of α-SMA-positive
cells in hepatic tissue from 50 mg/kg CCl4-induced hamsters treated
with adenoviral-BMP-7 transduction and/or doxazosin (Fig. 5A). As expected, the amount of
activated HSCs in liver of cirrhotic animals was 40.4-fold higher
than in Intact hamsters. When cirrhotic animals were placebo
administered there was a spontaneous decrease of 66.9% in the
number of liver activated HSCs. In the BMP-7 and Doxa groups,
α-SMA-positive cells were 31.7 and 29% significantly lower than in
the Placebo. Combination of BMP-7-adenoviral transduction and Doxa
did not improve reduction of α-SMA-positive cells in liver
parenchyma once cirrhosis was stablished (Fig. 5B). Taken together, these data
indicated that Doxa and BMP-7 exert the antifibrotic effect, at
least in part, by reduction of HSCs; although the combination of
both treatments does not increase the reduction of this activated
cell population.
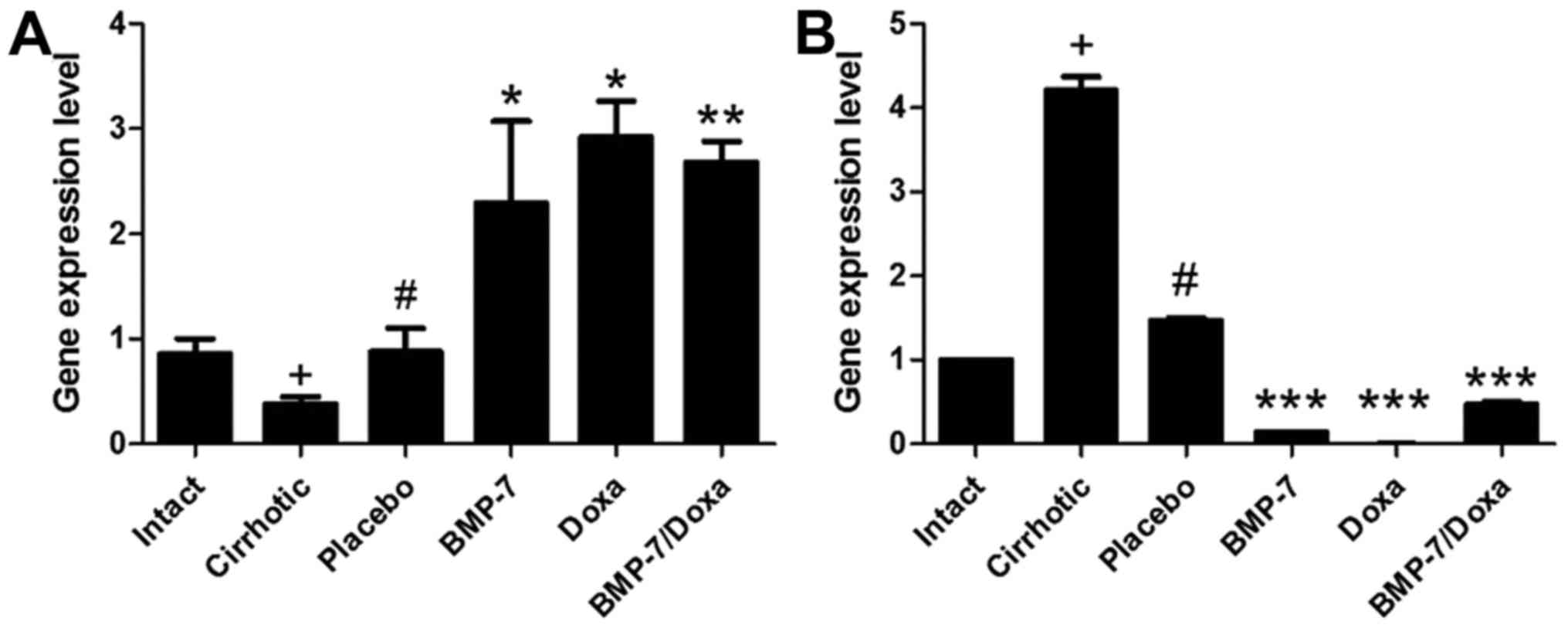
Changes in MMP-13 and TIMP-2
expression are associated to antifibrotic effect of experimental
treatments
We further examined MMP-13 (Fig. 6A) and its inhibitor TIMP-2
(Fig. 6B) mRNA expression, as both
molecules are involved in ECM remodeling. As it is known that
occurs in cirrhotic damage, MMP-13 expression was significant
decreased in liver of Cirrhotic animals while that of TIMP-2 was
increased. Placebo administration increased 2.3-fold MMP-13
expression and diminished in a 65.2% TIMP-2 mRNA levels. Consistent
with the histological data, the experimental treatments were
associated with upregulated MMP-13 mRNA expression, as well as
downregulated TIMP-2 mRNA. In the BMP-7 group, MMP-13 and TIMP-2
were inversely regulated with increment of 2.6-fold and reduction
of 11.2-fold, respectively, when compared to Placebo group. Also,
Doxa treatment promoted a 3.3-fold increased expression of MMP-13
and almost undetectable expression of TIMP-2. Nevertheless,
BMP-7/Doxa group had 3.0-fold increased expression of MMP-13 mRNA
and TIMP-2 mRNA was 3.1-fold lowered than Placebo group. These data
suggest that BMP-7-adenoviral transduction and/or doxazosin
treatments induce decrement in the collagen type I deposition by
increasing MMP-13 expression and reducing TIMP-2 expression. These
finding are consistent with collagen type I accumulation in the
cirrhotic animals. However, the combined BMP-7/Doxa therapy does
not exceed the benefits observed in the separates treatments.
Discussion
Cirrhosis represents the major pathological outcome
of continuous insult to the liver and it also precludes the
development of hepatocellular carcinoma in some patients.
Currently, drugs targeting fibrosis in clinical practice are
attracting the interest of researchers, who have generated
innovative therapeutic strategies addressed to target fibrogenic
cytokines, such as TGF-β, with blocking peptides (36), or through the inhibition of HSCs
activation (37). Thus, in the
present study, we evaluated whether the drug therapy with the
α-adrenergic blocker doxazosin or gene therapy through adenoviral
transduction of BMP-7, could have antifibrotic effects as
individual or combined treatments in a hamster model of
CCl4-induced cirrhosis.
Spontaneous reversion of the liver cirrhosis after
cessation of the CCl4-induction has been well documented (38–40).
We observed spontaneous improvement in the fibrotic process and
liver function in animals from Placebo group. Chávez et al
demonstrated spontaneous resolution of liver fibrosis in vehicle
treated CCl4-induced rats. The placebo treatment also diminished
the levels of ALT in CCl4-induced cirrhosis even after 4 weeks of
discontinued induction (41).
Besides, other authors reported that the spontaneous reversal of
cirrhosis after 6 weeks of vehicle daily administration was
accompanied by diminution of α-SMA positive cells and
down-expression of TIMP-1, as well by up-expression of MMP-13
(42). In our study, drug therapy
with doxazosin and BMP-7 showed greater recovery than spontaneous
reversion, outstanding their promissory role in the treatment of
fibrotic liver.
BMP-7 has inhibitory downstream activity over TGF-β
signaling, as Smad-1, −5 and −8, avoiding the activation of
Smad-3/4 mediated by TGF-β (24–26,43).
Thus, when BMP-7 was administered, fibrosis deposition, some liver
functions, HSCs hyperplasia, and expression changes in MMP and TIMP
were favored to retrieve the functionality of the liver. Studies
have also demonstrated how oral administration of a recombinant
adeno-associated virus (AAV)-mediated BMP-7 transduction reduces
the hepatic fibrosis and suppresses the activation of HSCs
(44). Moreover, gene delivery of
BMP-7 via polyethylenimine-conjugated gold nanoparticles in corneal
tissue demonstrated diminution of α-SMA positive cells and
fibronectin, in model of corneal fibrosis after photorefractive
keratectomy in rabbits (45).
Besides, administration of recombinant human BMP-7 reduced tubular
damage and tubulointerstitial fibrosis in streptozotocin-induced
diabetic mice with renal disease (46). Recently, natural compounds have
shown to promote the activation of BMP-7/Smad signaling, which in
turn modified the serum, biochemical and histological markers of
hepatic damage, indicating a diminution of liver fibrosis in a
model of bile duct ligation-induced liver fibrosis (47).
Previous studies have reported a significant
reduction in deposits of collagen type I in cirrhotic hamsters
treated with Doxazosin, when it was administered during 6 weeks
after liver damage development (13). The effect was related with a
decrease in TGF-β synthesis. We showed that when Doxazosin is only
administered for 2 weeks, the beneficial effect on cirrhosis is
maintained, as demonstrated by the decrease in the fibrotic area
and collagen type I deposit in the liver of cirrhotic CCl4-induced
hamsters. The antifibrotic effect of doxazosin has also been
reported in renal fibrosis and myocardial interstitial fibrosis
(48–50). As HSCs, the main fibrinogenic cells
in liver, express α1-AR (51,52)
we argue that is the selective inhibition of this receptor in HSCs
which is causing the decrease in number of activated cells and
therefore in collagen synthesis. Some differences are reported
between 2 or 6 weeks of treatment with Doxazosin in relation to
liver enzymes. When it was administered thought 6 weeks ALT was
significantly decreased (13), but
no significant change in liver enzymes is observed after 2 weeks of
treatment. However, the increase in plasma albumin after 2 weeks of
treatment is indicative of liver function recovery.
As both experimental therapies are targeted to
blockade (BMP-7) or inhibit (doxazosin) TGF-β secretion, we
hypothesized that co-administration of both therapies could have an
improved effect on ECM deposition in the treated hamsters. In the
present study, adenoviral-BMP-7 transduction as well as doxazosin
therapy, demonstrated to reduce, significantly, the deposition of
ECM, particularly of collagen type I, and to diminish the
expression of α-SMA in activated HSCs. We found that combination of
both treatments do have neither synergic nor an additive effect on
liver cirrhosis. Because of both treatments are targeted to
blockade the same fibrogenic mechanisms (13,31),
probably the total capacity of inhibition has been achieved by
individual therapies and then the combination is not able to
improve the antifibrotic effect.
In addition, ALT levels were significantly reduced
when combined therapies were administered, but AST and albumin
productions were unchanged by comparison to BMP-7. It is not
surprising that doxazosin treatment maintains increased levels of
AST or ALT after two weeks of treatment, but apparent liver
toxicity to the liver is reduced when BMP-7 is also administered.
Previously, doxazosin was reported to cause liver toxicity at doses
of 1 mg/kg/day at six weeks in the same model (13), moreover in the present study this
toxicity is maintained at 2 weeks of treatment; outstanding BMP-7
gene therapy reduced toxic effect of doxazosin. These data are
consistent with other study that have shown the protective effect
of BMP-7 to hepatocytes, with improved liver function and
regeneration (53).
Hepatocyte proliferation is also related to changes
in the expression of regulatory genes involved in ECM remodeling
(53,54). In this regard, Arendt et al
demonstrated that MMP-13 and TIMP-1 are differentially expressed
after TGF-β deprivation (55).
Besides, previous studies had shown that the N-Myc
downstream-regulated gene 2 abrogates the profibrogenic TGF-β
activity by diminution of the expression of TIMP-2 mRNA (56). We demonstrated that down-expression
of TIMP-2 mRNA correlate with reduction of collagen type I
deposition in BMP-7 and doxazosin treated groups, however
combinatory therapies were not as efficient as doxazosin alone.
In conclusion, the current study indicates that
BMP-7 diminish the deposition of collagen type I by hampering the
activation-proliferation of HSCs, and likely increasing hepatocyte
proliferation. Although BMP-7 gene therapy did not improve the
antifibrotic effect of doxazosin, it reduced its toxic effect in
the liver. The combination of gene and drug therapies may not favor
the increment of therapeutic effects, but its application can
reduce adverse effects of conventional drugs.
Acknowledgements
The present study was supported by the grants (nos.
241312 and 134487) awarded by the National Council of Science and
Technology (CONACYT), and PIBB15-10N from the Autonomous University
of Aguascalientes. LRAM is a Postdoctoral Fellow awarded by CONACYT
(grant no. 176507). Authors are grateful to Beatriz Orozco for
excellent technical assistance and animal care.
References
|
1
|
Mokdad AA, Lopez AD, Shahraz S, Lozano R,
Mokdad AH, Stanaway J, Murray CJ and Naghavi M: Liver cirrhosis
mortality in 187 countries between 1980 and 2010: A systematic
analysis. BMC Med. 12:1452014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Muir AJ: Understanding the complexities of
cirrhosis. Clin Ther. 37:1822–1836. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ramos-Lopez O, Martinez-Lopez E, Roman S,
Fierro NA and Panduro A: Genetic, metabolic and environmental
factors involved in the development of liver cirrhosis in Mexico.
World J Gastroenterol. 21:11552–11566. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ibañez P, Solis N, Pizarro M, Aguayo G,
Duarte I, Miquel JF, Accatino L and Arrese M: Effect of losartan on
early liver fibrosis development in a rat model of nonalcoholic
steatohepatitis. J Gastroenterol Hepatol. 22:846–851. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Friedman SL: Mechanisms of hepatic
fibrogenesis. Gastroenterology. 134:1655–1669. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Garbuzenko DV, Arefyev NO and Belov DV:
Mechanisms of adaptation of the hepatic vasculature to the
deteriorating conditions of blood circulation in liver cirrhosis.
World J Hepatol. 8:665–672. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang F, Liu S, Du T, Chen H, Li Z and Yan
J: NF-κB inhibition alleviates carbon tetrachloride-induced liver
fibrosis via suppression of activated hepatic stellate cells. Exp
Ther Med. 8:95–99. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shen M, Chen K, Lu J, Cheng P, Xu L, Dai
W, Wang F, He L, Zhang Y, Chengfen W, et al: Protective effect of
astaxanthin on liver fibrosis through modulation of TGF-β1
expression and autophagy. Mediators Inflamm. 2014:9545022014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gu L, Deng WS, Sun XF, Zhou H and Xu Q:
Rapamycin ameliorates CCl4-induced liver fibrosis in mice through
reciprocal regulation of the Th17/Treg cell balance. Mol Med Rep.
14:1153–1161. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Garcíade León Mdel C, Montfort I, Montes E
Tello, López Vancell R, García A Olivos, González Canto A,
Nequiz-Avendaño M and Pérez-Tamayo R: Hepatocyte production of
modulators of extracellular liver matrix in normal and cirrhotic
rat liver. Exp Mol Pathol. 80:97–108. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kang M, Zhao L, Ren M, Deng M and Li C:
Zinc mediated hepatic stellate cell collagen synthesis reduction
through TGF-β signaling pathway inhibition. Int J Clin Exp Med.
8:20463–20471. 2015.PubMed/NCBI
|
|
12
|
Hellerbrand C: Molecular targets for
antifibrotic therapy in liver disease: Using magic bullets for
crossfire rather than a one-sided shotgun attack. Gut.
63:1039–1041. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Muñoz-Ortega MH, Llamas-Ramírez RW,
Romero-Delgadillo NI, Elías-Flores TG, Tavares-Rodríguez Ede J,
Campos-Esparza Mdel R, Cervantes-García D, Muñoz-Fernández L,
Gerardo-Rodríguez M and Ventura-Juárez J: Doxazosin treatment
attenuates carbon tetrachloride-induced liver fibrosis in hamsters
through a decrease in transforming growth factor β secretion. Gut
Liver. 10:101–108. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ceral J and Solar M: Doxazosin: Safety and
efficacy in the treatment of resistant arteria hypertension. Blood
Press. 18:74–77. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tahmatzopoulos A, Rowland RG and Kyprianou
N: The role of alpha-blockers in the management of prostate cancer.
Expert Opin Pharmacother. 5:1279–1285. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mazzolini G, Alfaro C, Sangro B, Feijoó E,
Ruiz J, Benito A, Tirapu I, Arina A, Sola J, Herraiz M, et al:
Intratumoral injection of dendritic cells engineered to secrete
interleukin-12 by recombinant adenovirus in patients with
metastatic gastrointestinal carcinomas. J Clin Oncol. 23:999–1010.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hahn GV, Cohen RB, Wozney JM, Levitz CL,
Shore EM, Zasloff MA and Kaplan FS: A bone morphogenetic protein
subfamily: Chromosomal localization of human genes for BMP5, BMP6,
and BMP7. Genomics. 14:759–762. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Andrews PW, Damjanov I, Berends J, Kumpf
S, Zappavigna V, Mavilio F and Sampath K: Inhibition of
proliferation and induction of differentiation of pluripotent human
embryonal carcinoma cells by osteogenic protein-1 (or bone
morphogenetic protein-7). Lab Invest. 71:243–251. 1994.PubMed/NCBI
|
|
19
|
Midorikawa Y, Ishikawa S, Iwanari H,
Imamura T, Sakamoto H, Miyazono K, Kodama T, Makuuchi M and
Aburatani H: Glypican-3, overexpressed in hepatocellular carcinoma,
modulates FGF2 and BMP-7 signaling. Int J Cancer. 103:455–465.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Knight PG and Glister C: TGF-beta
superfamily members and ovarian follicle development. Reproduction.
132:191–206. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Genander M, Cook PJ, Ramsköld D, Keyes BE,
Mertz AF, Sandberg R and Fuchs E: BMP signaling and its pSMAD1/5
target genes differentially regulate hair follicle stem cell
lineages. Cell Stem Cell. 15:619–633. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su YH, Cai HB, Ye ZY and Tan WS: BMP-7
improved proliferation and hematopoietic reconstitution potential
of ex vivo expanded cord blood-derived CD34(+) cells. Hum Cell.
28:14–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Beck SE, Jung BH, Fiorino A, Gomez J,
Rosario ED, Cabrera BL, Huang SC, Chow JY and Carethers JM: Bone
morphogenetic protein signaling and growth suppression in colon
cancer. Am J Physiol Gastrointest Liver Physiol. 291:G135–G145.
2016. View Article : Google Scholar
|
|
24
|
Pegorier S, Campbell GA, Kay AB and Lloyd
CM: Bone morphogenetic protein (BMP)-4 and BMP-7 regulate
differentially transforming growth factor (TGF)-beta1 in normal
human lung fibroblasts (NHLF). Respir Res. 11:852010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang P and Dressler GR: The Groucho
protein Grg4 suppresses Smad7 to activate BMP signaling. Biochem
Biophys Res Commun. 440:454–459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lim RR, Tan A, Liu YC, Barathi VA, Mohan
RR, Mehta JS and Chaurasia SS: ITF2357 transactivates Id3 and
regulate TGFβ/BMP7 signaling pathways to attenuate corneal
fibrosis. Sci Rep. 6:208412016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kaimori A, Potter J, Kaimori JY, Wang C,
Mezey E and Koteish A: Transforming growth factor-beta1 induces an
epithelial-to-mesenchymal transition state in mouse hepatocytes in
vitro. J Biol Chem. 282:22089–22101. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang S and Hirschberg R: Bone
morphogenetic protein-7 signals opposing transforming growth factor
beta in mesangial cells. J Biol Chem. 279:23200–23206. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yanagita M: Inhibitors/antagonists of
TGF-β system in kidney fibrosis. Nephrol Dial Transplant.
27:3686–3691. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang G, Zhu Z, Wang Y, Gao A, Niu P and
Tian L: Bone morphogenetic protein-7 inhibits silica-induced
pulmonary fibrosis in rats. Toxicol Lett. 220:103–108. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang SL, Yang CQ, Qi XL, Yuan M, Chang YZ,
Yang L and Gao HJ: Inhibitory effect of bone morphogenetic
protein-7 on hepatic fibrosis in rats. Int J Clin Exp Pathol.
6:897–903. 2013.PubMed/NCBI
|
|
32
|
Wang LP, Dong JZ, Xiong LJ, Shi KQ, Zou
ZL, Zhang SN, Cao ST, Lin Z and Chen YP: BMP-7 attenuates liver
fibrosis via regulation of epidermal growth factor receptor. Int J
Clin Exp Pathol. 7:3537–3547. 2014.PubMed/NCBI
|
|
33
|
Hernandez-Hurtado AA, Borrego-Soto G,
Marino-Martinez IA, Lara-Arias J, Romero-Diaz VJ, Abrego-Guerra A,
Vilchez-Cavazos JF, Elizondo-Riojas G, Martinez-Rodriguez HG,
Espinoza-Juarez MA, et al: Implant composed of demineralized bone
and mesenchymal stem cells genetically modified with AdBMP2/AdBMP7
for the regeneration of bone fractures in ovis aries. Stem Cells
Int. 2016:74038902016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kondou H, Mushiake S, Etani Y, Miyoshi Y,
Michigami T and Ozono K: A blocking peptide for transforming growth
factor-beta1 activation prevents hepatic fibrosis in vivo. J
Hepatol. 39:742–748. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Osawa Y, Oboki K, Imamura J, Kojika E,
Hayashi Y, Hishima T, Saibara T, Shibasaki F, Kohara M and Kimura
K: Inhibition of cyclic adenosine monophosphate (cAMP)-response
element-binding protein (CREB)-binding protein (CBP)/β-catenin
reduces liver fibrosis in mice. EBioMedicine. 2:1751–1758. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bravo E, D'Amore E, Ciaffoni F and Mammola
CL: Evaluation of the spontaneous reversibility of carbon
tetrachloride-induced liver cirrhosis in rabbits. Lab Anim.
46:122–128. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Maros T, Seres-Sturm L, Lakatos O,
Seres-Sturm MT and Blazsek V: Spontaneous reversibility of advanced
toxic liver cirrhosis. Acta Morphol Acad Sci Hung. 23:293–302.
1975.PubMed/NCBI
|
|
40
|
Muriel P, Moreno MG, Hernández Mdel C,
Chávez E and Alcantar LK: Resolution of liver fibrosis in chronic
CCl4 administration in the rat after discontinuation of treatment:
Effect of silymarin, silibinin, colchicine and trimethylcolchicinic
acid. Basic Clin Pharmacol Toxicol. 96:375–380. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chávez E, Segovia J, Shibayama M, Tsutsumi
V, Vergara P, Castro-Sánchez L, Salazar EP, Moreno MG and Muriel P:
Antifibrotic and fibrolytic properties of celecoxib in liver damage
induced by carbon tetrachloride in the rat. Liver Int. 30:969–978.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lv P, Meng Q, Liu J and Wang C:
Thalidomide accelerates the degradation of extracellular matrix in
rat hepatic cirrhosis via down-regulation of transforming growth
factor-β1. Yonsei Med J. 56:1572–1581. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xu F, Liu C, Zhou D and Zhang L:
TGF-β/SMAD pathway and its regulation in hepatic fibrosis. J
Histochem Cytochem. 64:157–167. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hao ZM, Cai M, Lv YF, Huang YH and Li HH:
Oral administration of recombinant adeno-associated virus-mediated
bone morphogenetic protein-7 suppresses CCl(4)-induced hepatic
fibrosis in mice. Mol Ther. 20:2043–2051. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tandon A, Sharma A, Rodier JT, Klibanov
AM, Rieger FG and Mohan RR: BMP7 gene transfer via gold
nanoparticles into stroma inhibits corneal fibrosis in vivo. PLoS
One. 8:e664342013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sugimoto H, Grahovac G, Zeisberg M and
Kalluri R: Renal fibrosis and glomerulosclerosis in a new mouse
model of diabetic nephropathy and its regression by bone
morphogenic protein-7 and advanced glycation end product
inhibitors. Diabetes. 56:1825–1833. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hou F, Liu R, Liu X, Cui L, Wen Y, Yan S
and Yin C: Attenuation of liver fibrosis by herbal compound 861 via
upregulation of BMP-7/Smad signaling in the bile duct ligation
model rat. Mol Med Rep. 13:4335–4342. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gallego-Delgado J, Lazaro A, Gomez-Garre
D, Osende JI, Gonzalez-Rubio ML, Herraiz M, Manzarbeitia F, Fortes
J, Fernandez-Cruz A and Egido J: Long-term organ protection by
doxazosin and/or quinapril as antihypertensive therapy. J Nephrol.
19:588–598. 2006.PubMed/NCBI
|
|
49
|
Perlini S, Palladini G, Ferrero I, Tozzi
R, Fallarini S, Facoetti A, Nano R, Clari F, Busca G, Fogari R and
Ferrari AU: Sympathectomy or doxazosin, but not propranolol, blunt
myocardial interstitial fibrosis in pressure-overload hypertrophy.
Hypertension. 46:1213–1218. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Pawluczyk IZ, Patel SR and Harris KP: The
role of the alpha-1 adrenoceptor in modulating human mesangial cell
matrix production. Nephrol Dial Transplant. 21:2417–2424. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Oben JA, Roskams T, Yang S, Lin H, Sinelli
N, Torbenson M, Smedh U, Moran TH, Li Z, Huang J, et al: Hepatic
fibrogenesis requires sympathetic neurotransmitters. Gut.
53:438–445. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Oben JA, Roskams T, Yang S, Lin H, Sinelli
N, Li Z, Torbenson M, Huang J, Guarino P, Kafrouni M and Diehl AM:
Sympathetic nervous system inhibition increases hepatic progenitors
and reduces liver injury. Hepatology. 38:664–673. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sugimoto H, Yang C, LeBleu VS, Soubasakos
MA, Giraldo M, Zeisberg M and Kalluri R: BMP-7 functions as a novel
hormone to facilitate liver regeneration. FASEB J. 21:256–264.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jirouskova M, Zbodakova O, Gregor M,
Chalupsky K, Sarnova L, Hajduch M, Ehrmann J, Jirkovska M and
Sedlacek R: Hepatoprotective effect of MMP-19 deficiency in a mouse
model of chronic liver fibrosis. PLoS One. 7:e462712012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Arendt E, Ueberham U, Bittner R, Gebhardt
R and Ueberham E: Enhanced matrix degradation after withdrawal of
TGF-beta1 triggers hepatocytes from apoptosis to proliferation and
regeneration. Cell Prolif. 38:287–299. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yang J, Zheng J, Wu L, Shi M, Zhang H,
Wang X, Xia N, Wang D, Liu X, Yao L, et al: NDRG2 ameliorates
hepatic fibrosis by inhibiting the TGF-β1/Smad pathway and altering
the MMP2/TIMP2 ratio in rats. PLoS One. 6:e277102011. View Article : Google Scholar : PubMed/NCBI
|