Introduction
The 5-year survival rate for early-stage non-small
cell lung cancer (NSCLC) patients who are eligible for curative
surgery (1) varies from
approximately 66.3% (stage IB) to 83.9% (stage IA) (2), whereas, the 5-year survival rate of
patients with unresectable NSCLC is 5 to 17% even after
chemo-radiation therapy (3).
Current techniques, such as computed tomography, magnetic resonance
imaging, positron emission tomography, sputum cytology, and
histological examination of bronchoscopic tissues (4), are expensive or impractical for
large-scale population screening of early-stage NSCLC. Therefore,
the identification of efficient and accurate tools for early
detection is crucial.
Our previous study demonstrated that the expression
of the TRIM28 gene was high in NSCLC cell lines and tissues. TRIM28
provided a survival advantage to lung cancer cells, because TRIM28
knockdown decreased cell proliferation and inhibited cell cycle
progression in NSCLC cell lines (5). TRIM28 is an important member of the
tripartite motif superfamily which regulats embryonic development
(6), chromatin organization,
erythroblast differentiation (7),
and the DNA damage response (8–11).
Moreover, TRIM28 acts as a universal corepressor of KRAB zinc
finger proteins. Studies have shown that TRIM28 also plays a role
in the carcinogenesis and metastasis of cervical cancer (12), breast cancer (13), colorectal cancer (14), gastric cancer (15), kaposi's sarcoma (16), and ovarian cancer (17,18).
The downregulation of TRIM28 gene expression reduced the ability of
cancer stem cells to self-renew, resulting in a significant
reduction of tumor growth. The transforming growth factor-β
(TGF-β)-induced epithelial-mesenchymal transition was impaired by
shRNAi-mediated TRIM28 depletion via the acetylation and
methylation of histones on E-cadherin and N-cadherin promoters
(19). Loss of function of TRIM28
led to dysregulation of the cell cycle, cellular response to
stress, cancer cell metabolism, and inhibition of oxidative
phosphorylation (20). Despite
these well-documented biological functions of TRIM28, little is
known about the mechanisms underlying the role of TRIM28 in
NSCLC.
Serum marker detection is a high-profile topic for
auxiliary diagnosis of early NSCLC, and its advantages include easy
operation, low price, noninvasiveness, accessibility of samples,
and the ability for continuous monitoring (21). Clinical studies have examined
various indicators. Halvorsen et al (22) validated six microRNAs (miR-429,
miR-205, miR-200b, miR-203, miR-125b, and miR-34b) that were
candidate biomarkers for future screening detection, because their
abundance in the serum of NSCLC patients was significantly higher
than that in chronic obstructive pulmonary diseases (COPD) patients
and healthy volunteers. The serum laminin levels measured by
solid-phase sandwich enzyme-linked immunosorbent assay (ELISA) were
significantly higher in NSCLC patients, and this method might be
promising as a diagnostic supplement (23). The level of four serum markers
(CEA, CYFRA21-1, NSE and miR-21) was measured in 50 NSCLC patients
and 60 healthy donors, and the results indicated that serum miR-21
had the highest diagnostic value, whereas the combination of miR-21
and CYFRA21-1 improved the diagnostic efficiency for early NSCLC
(24). The serum of patients with
lung cancer was analyzed by mass spectrometry, and a total of 17
distinct predictive proteins were identified in NSCLC patients with
metastasis compared to healthy controls (25).
In this study, we observed that TRIM28 knockdown
exerted anti-tumor and pro-apoptotic activity using a nude mouse
xenograft tumor model of NSCLC. Here, using TRIM28 as the
tumor-associated antigen, we also determined the suitability of
reverse ELISA for the detection of specific antibodies in the serum
of NSCLC patients to develop an early diagnostic method and an
effective therapeutic strategy for NSCLC.
Patients and methods
Patients and samples
Serum samples from 138 NSCLC patients and 80 healthy
controls were collected between November 2012 and December 2016 in
the Affiliated Hospital of the Chengde Medical College Cancer
Center (Chengde, China). The cases were confirmed by
histopathological and clinical diagnosis. The cases involving
autoimmune disease, systemic inflammatory conditions, or the use of
immunosuppressive agents were excluded from the study. Tumor
differentiation and clinical stages were classified according to
the fifth edition of the tumor-node-metastasis classification of
the International Union Against Cancer. Patient recruitment and
serum analysis were conducted under written informed consent and
the study was approved by the Research Ethics Review Committee of
Chengde Medical College (Chengde, China).
Reverse ELISA
High-binding 96-well microtiter plates (Costar,
Cambridge, MA, USA) were coated with 200 mg/ml of TRIM28
recombinant protein (Abcam, Cambridge, UK) diluted in 0.1 M NaHCO3
(pH 8.3) overnight at 4°C. The plates were washed three times with
PBS containing Tween-20 and blocked with 1% bovine serum albumin
(BSA) for 2 h at 37°C. Serum samples were serially diluted in PBS
containing 1% BSA, and 100 µl of the diluted samples were applied
to microwells and incubated overnight at 4°C. Then, the plates were
washed three times and incubated for 2 h at 37°C in goat anti-human
HRP-conjugated secondary antibody (PharMingen, San Diego, CA, USA)
diluted in PBS. After washing, the microwells were developed using
TMB Peroxidase Substrate solution and the absorbance was read at
450 nm using a microplate autoreader.
The standard serum was assigned a value of 100 U/ml
rabbit anti-human TRIM28 PcAbs (Abcam). Positivity of TRIM28
autoantibodies for an individual patient was defined as an
absorbance value greater than two standard deviations (SD) above
the mean absorbance of non-cancer control sera.
Cell culture and RNA interference
assay
The protocol was described in our previous study
(26). The human NSCLC cell line
PAa (BioVector NTCC Inc., Shanghai, China) was grown in a monolayer
in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FCS) and was maintained at 37°C in an
atmosphere of humidified air with 5% CO2. Complementary
oligonucleotides containing a short hairpin RNA (shRNA) targeting
TRIM28 were dimerized and cloned into the pLVTHM lentiviral vector
(Shanghai GeneChem Co., Ltd., Shanghai, China). The recombinant
lentiviral vector and a lentiviral package plasmid were
cotransfected into 293T cells to produce the lentiviral particles.
PAa cells infected with lentiviral particles containing GFP were
selected by flow cytometry sorting using an anti-GFP monoclonal
antibody (Abcam) for culture and expansion. The stable transfected
PAa cells were cultured for 48 h, and the expression of TRIM28 at
the mRNA and protein levels was examined by reverse
transcription-polymerase chain reaction (RT-PCR) and western blot
analysis.
Tumorigenicity in nude mice
All animal studies were approved by the Research
Ethics Review Committee of Chengde Medical College (Chengde,
China). Twenty female BALB/c nude mice (Vital River Laboratories
Co., Ltd., Beijing, China) at 4 weeks of age were divided into two
groups of 10 animals. The two groups of mice were subcutaneously
injected with PAa/control-small interfering (si)RNA and
PAa/TRIM28-siRNA cells, respectively. Mice were observed daily for
viability and changes in general health and behavior. The body
weight of the mice was monitored weekly. Tumor volume was assessed
by measuring the length and width, and a standard equation was used
for the calculation of tumor volume: Tumor volume = length ×
width2 × 0.52, and was calculated for depicting a growth
curve. Except for mice with large tumor burdens, animals were
sacrificed 4 weeks after injection by cervical dislocation. Average
tumor size was estimated by physical measurement of the excised
tumor.
Terminal deoxynucleotidyl
transferase-mediated deoxyuridine triphosphate nick-end labeling
(TUNEL) assay
Apoptosis was quantitatively analyzed by using the
in situ apoptosis detection TUNEL kit (Beyotime Institute of
Biotechnology, Beijing, China) according to the manufacturer's
instructions. Five fields were randomly chosen under a fluorescence
microscope for each slide of each section (BZ-9000; Keyence
Corporation, Osaka, Japan). The apoptosis index (AI) was calculated
as the number of apoptotic cells/total cell number ×100%. Assays
were performed in a blinded manner.
Immunohistochemistry
The tumors were collected from nude mice and
processed into 5 µm slices. Paraffin sections were stained with
hematoxylin and eosin (H&E; Beyotime Institute of
Biotechnology). Mouse immunohistochemical studies for Ki-67 were
conducted by using the streptavidin-peroxidase method (Zhongshan
Golden Bridge Biotech Co., Ltd., Beijing, China). The tissue
sections were deparaffinized with xylene and rehydrated with
decreasing concentrations of ethanol. The sample slices were
incubated overnight using a primary rabbit polyclonal antibody
against Ki-67 (dilution of 1:100; ProteinTech Group, Inc., Chicago,
IL, USA) in a humidified container at 4°C. Immunohistochemistry was
performed on a non-biotin horseradish-peroxidase detection system
according to the manufacturer's instructions. Images were acquired
using a Leica IX71 microscope.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tumor specimens using
TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer's protocol. cDNA was synthesized from 2 µg of total
RNA using a reverse transcription kit (Takara, Dalian, China). PCRs
included the following components: 100 nmol/l of each primer,
diluted cDNA templates, and SYBR-Green PCR Master mix (Applied
Biosystems, Warrington, UK), to a total volume of 20 µl. The
real-time quantitative PCR assays were conducted with an ABI Prism
7000 SDS instrument (Applied Biosystems Life Technologies, Foster
City, CA, USA). Reactions were incubated at 94°C for 4 min and then
amplified using temperature cycles of 94°C for 15 sec, 58°C for 30
sec, 72°C for 35 sec. Amplifications were carried out for 45
cycles, followed by a 3 min extension at 72°C. All reactions were
performed in triplicate and the data were converted to fold change
and normalized to the expression level of GAPDH, which
served as an endogenous control. The relative gene expression
levels were calculated using the following equation: 2 – [Ct B cell
lymphoma (Bcl)-2/Bcl-2 associated X protein (Bax) – Ct control].
The primer sequences were as follows: Bcl-2 F,-5-TAC GAG TGG GAT
GCG GGA GAT GT-3 and R,-5-CCA CCG AAC TCA AAG AAG GC-3; Bax
F,-5-CAC CAG CTC TGA GCA GA T CA-3 and R,-5-ATG TCA GCT GCC ACT CGG
A-3; GAPDH F,-5-CAA TGA CCC CTT CAT TGA CC-3 and R,-5-TGG AAG ATG
GTG ATG GGA TT-3; and p53 F,-5-GCA GTT CCT CTT CCT GCA GTA CTC-3
and R, 5-AAC CAG ACC TCA GGC GGC TCA TAG-3 (Shenggong Biotech Co.,
Ltd., Shanghai, China).
Western blot analysis
Tumor tissues were lysed in ice-cold lysis buffer
containing 50 mmol/l Tris-HCl (pH 8.0), 150 mmol/l NaCl, 0.5% NP40,
0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), plus
protease inhibitor (Calbiochem, Darmstadt, Germany). Equal amounts
of proteins were subjected to SDS-polyacrylamide gel
electrophoresis using a 12% running gel. The gel-separated proteins
were transferred to polyvinylidene difluoride membranes (Millipore,
Bedford, MA, USA), and the membranes were blocked with 0.5% milk in
PBS for 1 h at room temperature. The membranes were washed in
Tris-buffered saline Tween (TBST), incubated with rabbit monoclonal
Bcl-2, Bax or p53 antibody (dilution of 1:500; ProteinTech Group,
Inc.) overnight at 4°C, washed again in TBST, incubated with
horseradish peroxidase-conjugated goat anti-rabbit IgG for 1 h at
room temperature, washed again in TBST, and detected by
chemiluminescence using the ECL detection kit (Amersham,
Buckinghamshire, UK).
Statistical analysis
Data were presented as mean and SD. Analysis of
variance followed by Bonferroni's test was used to determine the
significant differences between groups. P<0.05 was considered
significant. All statistical analyses were conducted using software
version SPSS 17.0 (SPSS, Inc., Chicago, IL, USA).
Results
Knockdown of TRIM28 inhibited the
tumor growth in vivo
TRIM28 specific siRNA lentivirus vector and a
negative control vector (si-control) were designed and constructed
to silence TRIM28 in lung adenocarcinoma cell PAa. Nude mice
received an injection of PAa/TRIM28-siRNA cells and
PAa/control-siRNA cells for 4 weeks (5 times/week). Western blot
assay were used to confirm TRIM28 siRNA led to a marked reduction
of protein levels both in PAa cell line and tumor tissue from nude
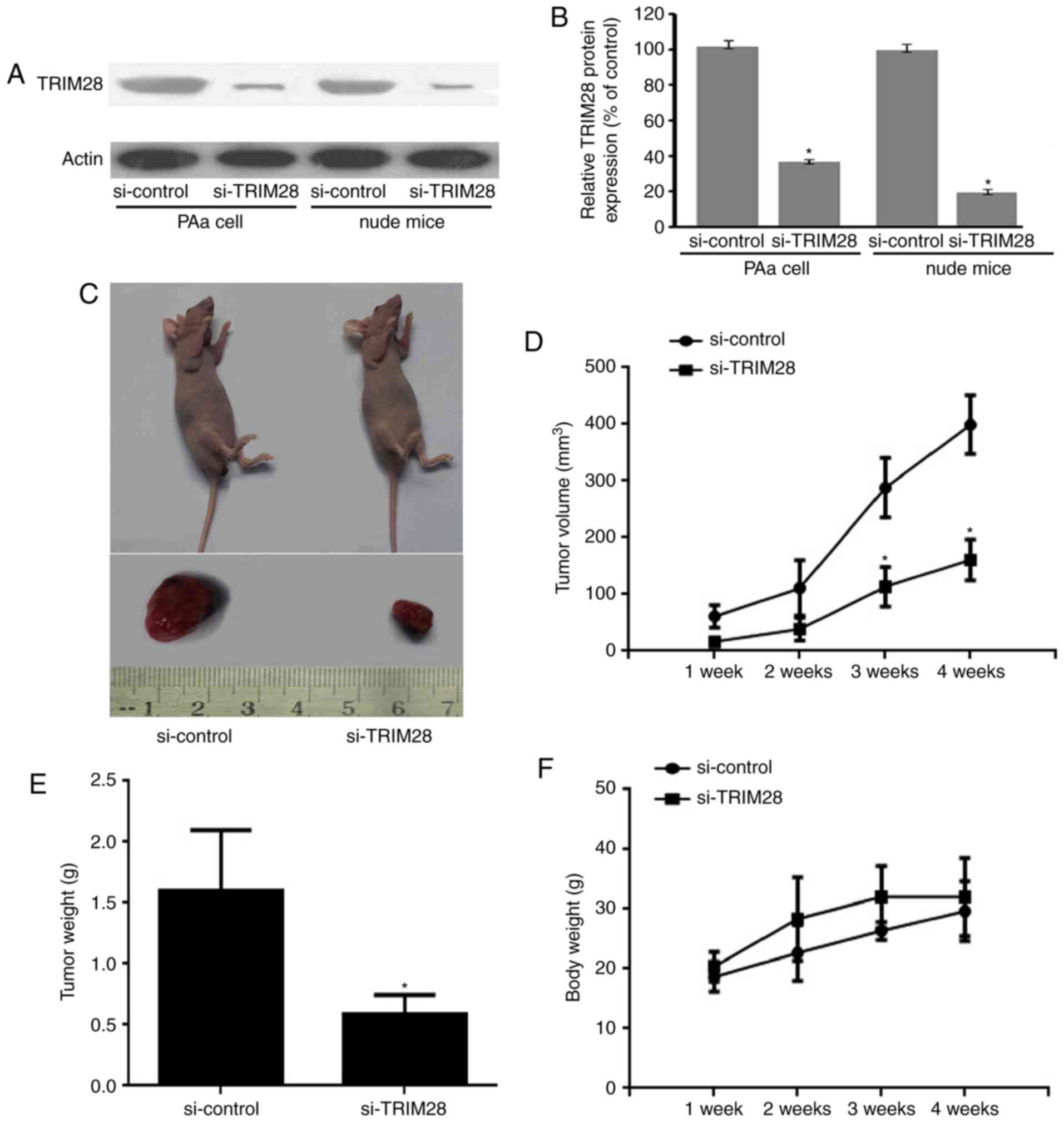
mice (P<0.001, relative to the si-control; Fig. 1A and B).
The tumor growth of nude mice was assessed during
the 4-week treatment period. TRIM28 siRNA inhibited tumor growth,
and the inhibition was significantly higher than that of
PAa/control-siRNA (Fig. 1C).
Tumors were developed within 1 week in mice injected with
PAa/control-siRNA. In the PAa/TRIM28-siRNA treated group, tumor
formation was delayed by more than 1 week compared with the control
group (Fig. 1D). Tumor size in
mice from the PAa/TRIM28-siRNA group was significantly smaller than
in the PAa/control-siRNA group (110±15 vs. 294±27 mm3 at
3 weeks, P<0.001, and 155±14 vs. 407±24 mm3 at 4
weeks, P<0.001) (Fig. 1D).
Furthermore, the treatment with PAa/TRIM28-siRNA significantly
decreased tumor weight (0.71±0.28 g) compared with
PAa/control-siRNA (1.67±0.81 g, P<0.001; Fig. 1E). None of the evaluated mice lost
body weight during the experimental period (Fig. 1F).
TRIM28 siRNA stimulated apoptosis in a
nude mouse xenograft tumor model
We used histology to evaluate the effect of TRIM28
siRNA on tumor growth in the nude mice. H&E staining of tumor
tissues demonstrated that microvessels filled with erythrocytes
were abundant in tumor tissues from mice injected with
PAa/control-siRNA cells; however, a few microvessels were observed
in tumors treated with PAa/TRIM28-siRNA (Fig. 2A, upper panel). Cell proliferation
was detected by Ki-67 staining. The number of cells that stained
positive for Ki-67 in tumor tissues from mice treated with
PAa/TRIM28-siRNA (4.13±1.42%) was significantly smaller than in
mice injected with PAa/control-siRNA cells (14.23±2.49%, P=0.013;
Fig. 2A, middle panel, and
Fig. 2B). The degree of apoptosis
was analyzed by conducting a TUNEL assay. The knockdown of TRIM28
significantly induced apoptosis compared to the control group
(Fig. 2A, lower panel). The
percentage of total nuclei in TUNEL-positive cells in mice injected
with PAa/TRIM28-siRNA and PAa/control-siRNA was 21.67±2.58 and
5.08±1.89%, respectively (P<0.001; Fig. 2A, lower panel, and Fig. 2C).
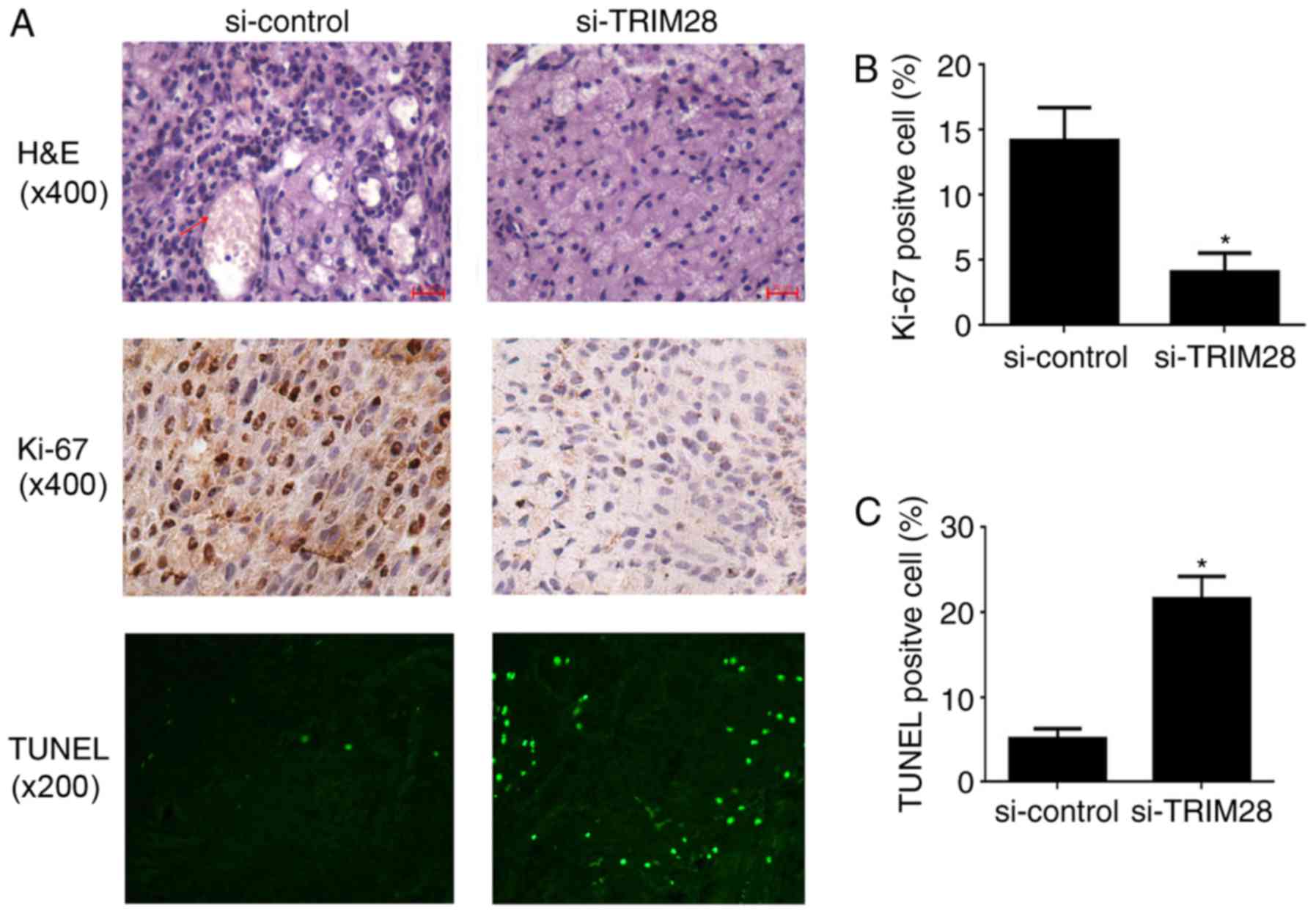 | Figure 2.Histology and immunohistochemistry of
tumor tissues from xenografted nude mice. (A) Top, H&E
staining. As indicated by the red arrow, the red-stained material
in tumors from the control group is erythrocytes and the adjacent
structures are blood vessels, which are rarely observed in tumors
from PAa/TRIM28-siRNA treated mice (magnification, ×400). Middle,
immunostaining for Ki-67 (magnification, ×400). Bottom, TUNEL
staining (magnification, ×200). (B) The percentages of
Ki-67-positive nuclei was determined on four slides for each group
and expressed as mean ± SD. (C) Analysis of the degree of apoptosis
in tumors from mice. *P<0.001 compared to the si-control
group. H&E, hematoxylin and eosin; si, small interfering;
TUNEL, terminal deoxynucleotidyl transferase-mediated deoxyuridine
triphosphate nick-end labeling; SD, standard deviation. |
Effects of TRIM28 knockdown on the
expression levels of Bcl-2, Bax, and p53
Tumors were removed from mice after treatment, and
the expression levels of the anti-apoptotic gene Bcl-2 and
pro-apoptotic genes Bax and p53 were measured by real-time PCR and
western-blotting. The expression of Bcl-2 was downregulated
(Fig. 3A, D and E) whereas the
expression of Bax (Fig. 3B, D and
E) and p53 (Fig. 3C-E) was
upregulated at both the mRNA and protein levels in the tumor
tissues from mice injected the PAa/TRIM28-siRNA cells compared to
the levels in the control-siRNA group (P<0.001). These
results suggest that the knockdown of TRIM28 expression in nude
mice models caused differential expression of pro- and
anti-apoptotic genes.
Detection of TRIM28 serum
autoantibodies in NSCLC patients
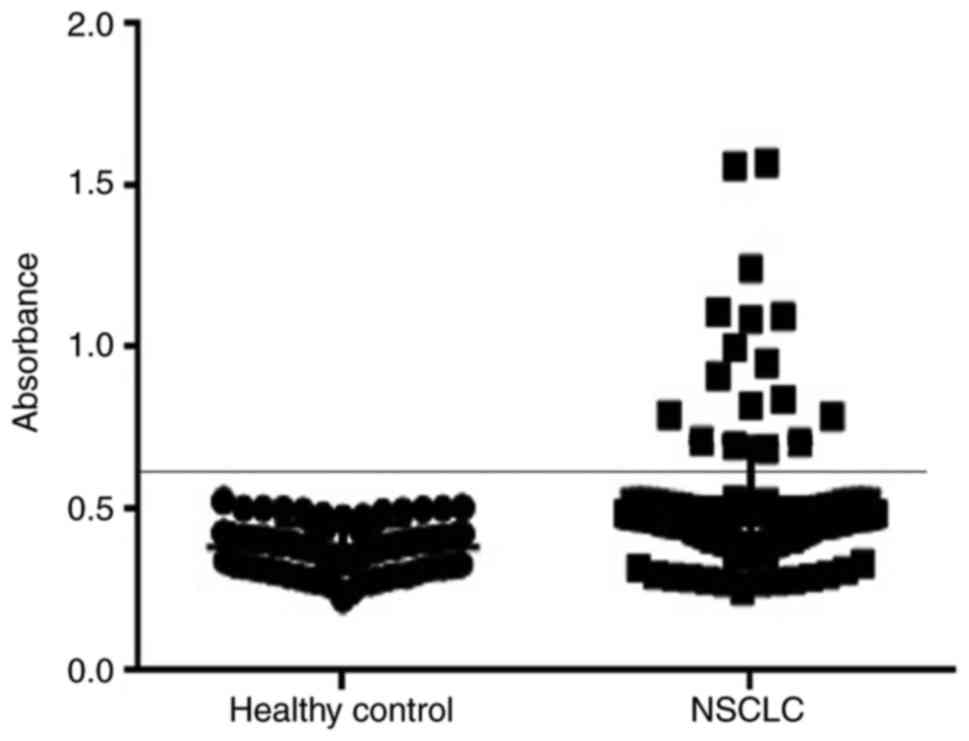
In our study, 138 NSCLC patients and 80 healthy
donors were recruited. Autoantibodies to TRIM28 were detected in
the sera of 17 NSCLC patients but in none of the healthy controls
(P=0.001), corresponding to a sensitivity of 12.32% (17/138) and a
specificity of 100% (Fig. 4). Of
note, antibodies to TRIM28 were detected in 14.43% (14/97) of
patients with early stage I and II disease and 7.32% (3/41) of
patients with more advanced disease (stage III and IV). The
positive detection rate of TRIM28 was non-significantly higher in
adenocarcinomas (18.97%, 11/58) than in squamous epithelial
carcinoma (9.80%, 4/51) and large cell lung cancer (6.90%, 2/29)
(Table I). The relationship
between the detection of TRIM28 antibodies and the clinical outcome
of NSCLC patients was investigated by conducting a 70-month
follow-up of 97 early-stage patients. The incidence of recurrence
and/or metastasis was significantly higher (P=0.001) in the
patients with TRIM28 antibodies (85.7%, 12 of 14) than in those
without TRIM28 antibodies (39.8%, 33 of 83). Of note was that among
these 12 patients with TRIM28 antibodies, 10 died during
follow-up.
 | Table I.Patient characteristics and the
positive rate of TRIM28 serum autoantibodies in 138 non-small lung
cancer patients. |
Table I.
Patient characteristics and the
positive rate of TRIM28 serum autoantibodies in 138 non-small lung
cancer patients.
|
| NSCLC |
|
|---|
|
|
|
|
|---|
|
Characteristics | n | positive rate
(%) | Healthy control
(n) |
|---|
| Age, years |
|
|
|
|
>60 | 74 | 9
(12.16) | 45 |
|
<60 | 64 | 8
(12.50) | 35 |
| Sex |
|
|
|
|
Male | 88 | 11 (12.50) | 32 |
|
Female | 50 | 6
(12.00) | 48 |
| Pathology |
|
|
|
|
SCC | 51 | 4 (9.80) |
|
|
Ade | 58 | 11 (18.97) |
|
|
LCLC | 29 | 2 (6.90) |
|
| Clinical
stages |
|
|
|
| I and
II | 97 | 14 (14.43) |
|
| III and
IV | 41 | 3 (7.32) |
|
| Tumor
differentiation |
|
|
|
|
Good | 36 | 4
(11.11) |
|
|
Moderate | 62 | 9
(14.52) |
|
|
Poor | 40 | 4
(10.00) |
|
Discussion
TRIM28 is an important regulator of carcinogenesis
in several neoplasms. Addison et al (13) showed that TRIM28 was overexpressed
in breast tumors and the depletion of TRIM28 in breast cancer cell
lines slowed cell proliferation and inhibited tumor growth and
metastasis in mice xenografts models. Bojkowska et al
(27) demonstrated that
liver-specific TRIM28 knockout induced strikingly sexually
dimorphic phenotypic disturbances, and developed male-specific
steatosis and secondary hepatic tumors. The expression of the
TRIM28 gene was significantly higher in gastric cancer tissues and
the proliferation rate of cancer and resistance to anoikis were
decreased after the knockdown of TRIM28 in gastric cancer cell
lines (15). Here, we also showed
that TRIM28 knockdown significantly inhibited NSCLC tumor growth in
nude mice. These results suggest that TRIM28 promoted tumor
progression and provided a survival advantage to tumor cells.
Therefore, the knockdown of TRIM28 may be a therapeutic target for
several tumors. In our study, the H&E staining of histology
sections suggests that TRIM28 knockdown may play a role in
antiangiogenesis, which may be a putative mechanism of inhibition
of tumor formation. However, further studies are necessary to
elucidate the underlying mechanisms.
The present study showed that the knockdown of
TRIM28 significantly induced apoptosis compared to the control
group (P<0.001). Moreover, TRIM28 knockdown increased the
expression of Bax and p53 and decreased the expression of Bcl-2 at
both the gene and protein levels. Wang et al (28) demonstrated that TRIM28 interacted
with MDM2, which stimulated the formation of the p53-HDAC1 complex.
Furthermore, the knockout of TRIM28 promoted the acetylation and
transcriptional activity of p53, sensitized the response of p53 to
DNA damage, and increased apoptosis. A study showed that TRIM28
regulated apoptosis induced by the E2F1 signaling pathway by
binding to E2F1 transcription factors in a retinoblastoma
protein-independent manner and inhibiting E2F1 activity (29). Another study suggests that TRIM28
plays a role in other signaling pathways associated with apoptosis
in tumorigenesis. Kamitani et al (30) found that TRIM28 associated with
endogenous STAT1 repressed the interferon-mediated signaling
pathway. A siRNA-mediated reduction in TRIM28 expression enhanced
interferon-induced STAT1-dependent IRF-1 gene expression. Tsuruma
et al (31) found that
endogenous TRIM28 associated with endogenous STAT3 in vivo.
The siRNA-mediated reduction in TRIM28 expression enhanced
interleukin-6-induced STAT3-dependent transcription and gene
expression and the marked accumulation of STAT3 phosphorylated on
Ser727 in the nucleus. Although these studies suggested that TRIM28
may play a role in many signal pathways of apoptosis, the mechanism
underlying this role are not clear.
In our study, the subcutaneous xenograft model in
nude mice was set up to demonstrate that the silencing of TRIM28
expression by a siRNA lentivirus vector inhibited the growth of
NSCLC. If the further roles of TRIM28 are to be explored,
influencing metastasis in particular, a clinically relevant
orthotopic model of lung cancer put forth by the recent researches
(32–34) is needed. Human lung cancer cell
lines were genetically transformed to express fluorescent protein.
Tumors were grown subcutaneously for each cell line and harvested
and minced for surgical orthotopic implantation on the lung of nude
mice. Tumor growth and metastasis were measured by fluorescence
imaging (34).
Several studies have shown that TRIM28 may be a
valuable biomarker for tumor diagnosis and prognosis prediction.
Wang et al (35)
demonstrated that TRIM28 was overexpressed in human hepatoma cell
lines and hepatocellular cancer (HCC) tumor tissues. Moreover, the
abnormal expression of TRIM28 was strongly correlated with the
clinical stage and tumor size and thus this molecule might be a
predictor of poor prognosis in HCC patients. The correlation
between TRIM28 expression and clinicopathological features was
studied in ovarian cancer (17).
The results of multivariate analysis showed that TRIM28
overexpression was an independent predictor of cancer and was
correlated with clinical stage, pathological grade, metastases and
poor survival. Wang et al (36) used RT-qPCR to detect the expression
levels of TRIM28 in peripheral blood karyocytes of gastric cancer
patients. The results revealed significant correlations between
TRIM28, TNM tumor stages, and distant metastases. Using a cohort of
colorectal cancer patients and non-cancerous controls, Hector et
al (37) demonstrated that
autoantibodies to TRIM28 were detectable in the sera of several
colorectal cancer patients compared to controls.
Using nested real-time PCR, we measured the
expression levels of the TRIM28 gene in the circulating cancer
cells of these patients in a previous study (5). Here, autoantibodies to TRIM28 were
determined in these patients' sera, representing the measurements
at the protein level. The positive detection rates were 30.4%
(42/138) for the TRIM28 gene and 12.32% (17/138) for TRIM28
autoantibodies in 138 NSCLC patients. The expression of the TRIM28
gene was detected in the sera of all 17 patients with TRIM28
antibodies in circulating cancer cells. In turn, the expression of
TRIM28 gene and TRIM28 autoantibodies was not detected in the 80
normal controls. The detection rates were 29.9% (29/97) for TRIM28
gene and 14.43% (14/97) for TRIM28 autoantibodies in early-stage
NSCLC, which suggests that the expression of TRIM28 at both the
gene and protein levels may be valuable for early diagnosis. In
addition, our results indicate that TRIM28 may be a useful marker
for the prediction of metastasis and prognosis. The incidence of
recurrence and/or metastasis was significantly higher in the TRIM28
gene positive group (20 of 29, 69.0%, P=0.004) than in the TRIM28
gene negative group (25 of 68, 36.8%). Similarly, the rate of
recurrence and/or metastasis was higher (12 of 14, 85.7%) in
early-stage patients with TRIM28 antibodies (10 patients died in
this period) than in patients without TRIM28 antibodies (33 of 83,
39.8%).
In contrast, our findings demonstrated that the
autoantibody-positive rate was only 3.72% in advanced-stage NSCLC
patients vs. 14.43% in early-stage patients. These results suggest
that tumor-associated antigens may activate a complete set of
specific and nonspecific defense mechanisms in early-stage
patients. In turn, in advanced stages, when the host defense
mechanisms against cancer are ineffective, tumor cells evade the
immune system (immune incompetent patients) and malignant cells
grow and expand in an uncontrolled manner.
In conclusion, our results suggest that the
knockdown of TRIM28 expression by lenti-siRNA/TRIM28 may inhibit
tumor growth and induce cell apoptosis in vivo. In addition,
we demonstrated the high level of specificity of TRIM28
autoantibodies in the sera of NSCLC patients. These results suggest
that TRIM28 knockdown may be a valuable approach for the treatment
of NSCLC and TRIM28 autoantibodies may act as a blood-based tumor
marker to detect early- stage NSCLC.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (81703001), Hebei Natural Science
Foundation (H2015406014), Hebei Province Talent Engineering
Training Funded Research Projects (A2016002085), University
Emphasis Subject of Hebei Province and Emphasis Subject of Chengde
Medical College
References
|
1
|
National Lung Screening Trial Research
Team, . Aberle DR, Berg CD, Black WC, Church TR, Fagerstrom RM,
Galen B, Gareen IF, Gatsonis C, Goldin J, et al: The National Lung
Screening Trial: Overview and study design. Radiology. 258:243–253.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Asamura H, Goya T, Koshiishi Y, Sohara Y,
Eguchi K, Mori M, Nakanishi Y, Tsuchiya R, Shimokata K, Inoue H, et
al: A Japanese Lung Cancer Registry study: Prognosis of 13,010
resected lung cancers. J Thorac Oncol. 3:46–52. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Aberle DR, DeMello S, Berg CD, Black WC,
Brewer B, Church TR, Clingan KL, Duan F, Fagerstrom RM, Gareen IF,
et al: Results of the two incidence screenings in the National Lung
Screening Trial. N Engl J Med. 369:920–931. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu L, Zhao E, Li C, Huang L, Xiao L,
Cheng L, Huang X, Song Y and Xu D: TRIM28, a new molecular marker
predicting metastasis and survival in early-stage non-small cell
lung cancer. Cancer Epidemiol. 37:71–78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Messerschmidt DM, de Vries W, Ito M,
Solter D, Ferguson-Smith A and Knowles BB: Trim28 is required for
epigenetic stability during mouse oocyte to embryo transition.
Science. 335:1499–1502. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barde I, Rauwel B, Marin-Florez RM,
Corsinotti A, Laurenti E, Verp S, Offner S, Marquis J, Kapopoulou
A, Vanicek J and Trono D: A KRAB/KAP1-miRNA cascade regulates
erythropoiesis through stage-specific control of mitophagy.
Science. 340:350–353. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pfeifer GP: Protein phosphatase PP4: Role
in dephosphorylation of KAP1 and DNA strand break repair. Cell
Cycle. 11:2590–2591. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
White DE, Negorev D, Peng H, Ivanov AV,
Maul GG and Rauscher FJ III: KAP1, a novel substrate for PIKK
family members, colocalizes with numerous damage response factors
at DNA lesions. Cancer Res. 66:11594–11599. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gilmore-Hebert M, Ramabhadran R and Stern
DF: Interactions of ErbB4 and Kap1 connect the growth factor and
DNA damage response pathways. Mol Cancer Res. 8:1388–1398. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hu C, Zhang S, Gao X, Gao X, Xu X, Lv Y,
Zhang Y, Zhu Z, Zhang C, Li Q, et al: Roles of kruppel-associated
box (KRAB)-associated co-repressor KAP1 ser-473 phosphorylation in
DNA damage response. J Biol Chem. 287:18937–18952. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin LF, Li CF, Wang WJ, Yang WM, Wang DD,
Chang WC, Lee WH and Wang JM: Loss of ZBRK1 contributes to the
increase of KAP1 and promotes KAP1-mediated metastasis and invasion
in cervical cancer. PLoS One. 8:e730332013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Addison JB, Koontz C, Fugett JH, Creighton
CJ, Chen D, Farrugia MK, Padon RR, Voronkova MA, McLaughlin SL,
Livengood RH, et al: KAP1 promotes proliferation and metastatic
progression of breast cancer cells. Cancer Res. 75:344–355. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fitzgerald S, Sheehan KM, O'Grady A, Kenny
D, O'Kennedy R, Kay EW and Kijanka GS: Relationship between
epithelial and stromal TRIM28 expression predicts survival in
colorectal cancer patients. J Gastroenterol Hepatol. 28:967–974.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yokoe T, Toiyama Y, Okugawa Y, Tanaka K,
Ohi M, Inoue Y, Mohri Y, Miki C and Kusunoki M: KAP1 is associated
with peritoneal carcinomatosis in gastric cancer. Ann Surg Oncol.
17:821–828. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang L, Zhu C, Guo Y, Wei F, Lu J, Qin J,
Banerjee S, Wang J, Shang H, Verma SC, et al: Inhibition of KAP1
enhances hypoxia-induced Kaposi's sarcoma-associated herpesvirus
reactivation through RBP-Jκ. J Virol. 88:6873–6884. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cui Y, Yang S, Fu X, Feng J, Xu S and Ying
G: High levels of KAP1 expression are associated with aggressive
clinical features in ovarian cancer. Int J Mol Sci. 16:363–377.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu M, Fu X, Cui Y, Xu S, Xu Y, Dong Q and
Sun L: Expression of KAP1 in epithelial ovarian cancer and its
correlation with drug-resistance. Int J Clin Exp Med.
8:17308–17320. 2015.PubMed/NCBI
|
|
19
|
Chen L, Muñoz-Antonia T and Cress WD:
Trim28 contributes to EMT via regulation of E-cadherin and
N-cadherin in lung cancer cell lines. PLoS One. 9:e1010402014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Czerwińska P, Shah PK, Tomczak K, Klimczak
M, Mazurek S, Sozańska B, Biecek P, Korski K, Filas V, Mackiewicz
A, et al: TRIM28 multi-domain protein regulates cancer stem cell
population in breast tumor development. Oncotarget. 8:863–882.
2017.PubMed/NCBI
|
|
21
|
Du ZY, Shi MH, Ji CH and Yu Y: Serum
pleiotrophin could be an early indicator for diagnosis and
prognosis of non-small cell lung cancer. Asian Pac J Cancer Prev.
16:1421–1425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Halvorsen AR, Bjaanæs M, LeBlanc M, Holm
AM, Bolstad N, Rubio L, Peñalver JC, Cervera J, Mojarrieta JC,
López-Guerrero JA, et al: A unique set of 6 circulating microRNAs
for early detection of non-small cell lung cancer. Oncotarget.
7:37250–37259. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tas F, Bilgin E, Tastekin D, Erturk K and
Duranyildiz D: Clinical significance of serum laminin levels in
patients with lung cancer. Biomed Rep. 4:485–488. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun M, Song J, Zhou Z, Zhu R, Jin H, Ji Y,
Lu Q and Ju H: Comparison of serum microRNA21 and tumor markers in
diagnosis of early non-small cell lung cancer. Dis Markers.
2016:38231212016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sreseli RT, Binder H, Kuhn M, Digel W,
Veelken H, Sienel W, Passlick B, Schumacher M, Martens UM and
Zimmermann S: Identification of a 17-protein signature in the serum
of lung cancer patients. Oncol Rep. 24:263–270. 2010.PubMed/NCBI
|
|
26
|
Liu L, Xiao L, Liang X, Chen L, Cheng L,
Zhang L, Wu X, Xu Q and Ma C: TRIM28 knock-down increases
sensitivity to etoposide by upregulating E2F1 in non-small cell
lung cancer. Oncol Rep. 37:3597–3605. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bojkowska K, Aloisio F, Cassano M,
Kapopoulou A, de Sio Santoni F, Zangger N, Offner S, Cartoni C,
Thomas C, Quenneville S, et al: Liver-specific ablation of
Krüppel-associated box-associated protein 1 in mice leads to
male-predominant hepatosteatosis and development of liver adenoma.
Hepatology. 56:1279–1290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang C, Ivanov A, Chen L, Fredericks WJ,
Seto E, Rauscher FJ III and Chen J: MDM2 interaction with nuclear
corepressor KAP1 contributes to p53 inactivation. EMBO J.
24:3279–3290. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang C, Rauscher FJ III, Cress WD and Chen
J: Regulation of E2F1 function by the nuclear corepressor KAP1. J
Biol Chem. 282:29902–29909. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kamitani S, Ohbayashi N, Ikeda O, Togi S,
Muromoto R, Sekine Y, Ohta K, Ishiyama H and Matsuda T: KAP1
regulates type I interferon/STAT1-mediated IRF-1 gene expression.
Biochem Biophys Res Commun. 370:366–370. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tsuruma R, Ohbayashi N, Kamitani S, Ikeda
O, Sato N, Muromoto R, Sekine Y, Oritani K and Matsuda T: Physical
and functional interactions between STAT3 and KAP1. Oncogene.
27:3054–3059. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rashidi B, Yang M, Jiang P, Baranov E, An
Z, Wang X, Moossa AR and Hoffman RM: A highly metastatic Lewis lung
carcinoma orthotopic green fluorescent protein model. Clin Exp
Metastasis. 18:57–60. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rashidi B, Moossa AR and Hoffman RM:
Specific route mapping visualized with GFP of single-file streaming
contralateral and systemic metastasis of Lewis lung carcinoma cells
beginning within hours of orthotopic implantation [correction of
implantion]. J Cell Biochem. 114:1738–1743. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yano S, Zhang Y, Miwa S, Kishimoto H,
Urata Y, Bouvet M, Kagawa S, Fujiwara T and Hoffman RM: Precise
navigation surgery of tumours in the lung in mouse models enabled
by in situ fluorescence labelling with a killer-reporter
adenovirus. BMJ Open Respir Res. 2:e0000962015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang Y, Jiang J, Li Q, Ma H, Xu Z and Gao
Y: KAP1 is overexpressed in hepatocellular carcinoma and its
clinical significance. Int J Clin Oncol. 21:927–933. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang YY, Li L, Zhao ZS and Wang HJ:
Clinical utility of measuring expression levels of KAP1, TIMP1 and
STC2 in peripheral blood of patients with gastric cancer. World J
Surg Oncol. 11:812013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hector S, Chen H, Kijanka G, Murray F and
Prehn JH: A reverse-ELISA for the detection of TRIM28/KAP1 serum
autoantibodies in colorectal cancer patients. Acta Oncol.
51:394–396. 2012. View Article : Google Scholar : PubMed/NCBI
|