Introduction
Alzheimer's disease (AD) is a neurodegenerative
disorder with pathological hallmarks including senile plaques
induced by abnormal deposition of amyloid-β (Aβ) and
neurofibrillary tangles (NFTs) formed by aberrantly hyperphorylated
tau protein. It is well known that Aβ has a critical role in the
pathogenesis of AD, as the accumulation of Aβ leads to
neuroinflammation and induces neuronal damage (1). One of the most important mechanisms
of Aβ accumulation in the brain is transshipment imbalances through
the blood-brain barrier (BBB) (2),
and two primary influx and efflux receptors have been implicated in
this process: Receptor for advanced glycation endproducts (RAGE)
and lipoprotein receptor-related protein-1 (LRP-1) (3–5).
RAGE acts as an important transporter via regulating influx of
circulating Aβ into the brain (6),
whereas LRP1 mediates the efflux of brain-derived Aβ into the
circulation via the BBB (7).
Furthermore, RAGE is a multi-ligand receptor of the immunoglobulin
superfamily of cell surface molecules, with RAGE ligands consisting
of advanced glycation end products and Aβ proteins (8). It has been indicated that these
ligand-receptor interactions are able to activate receptor-mediated
signaling pathways, including the mitogen-activated protein kinase
(MAPK) and nuclear factor (NF)-κB pathway (9), and subsequently promote the
production of pro-inflammatory cytokines, such as interleukin
(IL)-1β, IL-6 and tumor necrosis factor (TNF)-α (10,11).
Therefore, RAGE/MAPK, the NF-κB signaling pathway and the
downstream inflammatory response may be a potential therapeutic
target for Aβ-induced brain damage.
Lingguizhugan decoction (LGZG) is composed of Poria
(the dried fungus nucleus of Poria cocos (Schw.) Wolf),
Cinnamomi ramulus (the dried twigs of Cinnamomum cassia (L.)
J. Presl), Atractylodis macrocephalae rhizoma (the fried and dried
rhizome of Atractylodes macrocephala Koidz) and Glycyrrhizae
radix et rhizoma (the dried roots and rhizome of Glycyrrhizae
uralensis Fisch.), and is widely used in Traditional Chinese
Medicine. The formula originates from Treatise on Febrile Diseases
(12), and has been used for the
treatment of AD, inflammatory injury and fluid retention (13). It has previously been demonstrated
that LGZG inhibits the levels of pro-inflammatory cytokines
produced by Aβ1−42 induced BV-2 microglia cells
(14). These findings indicate the
potential of LGZG in the treatment of AD; however, the molecular
mechanism remains unclear. Therefore, the aim of the present study
was to investigate the effect of LGZG on Aβ-induced AD rats.
Materials and methods
Animals
All experiments and animal care protocols in the
present study were performed in accordance with the Guide for the
Care and Use of Laboratory Animals (15). Ethical approval for these protocols
was granted by the Medical Ethics Committee of Nanjing University
of Traditional Chinese Medicine [Nanjing, China; approval no.
ACU-06(20151113)].
Male Sprague-Dawley rats (n=56; 8 week-old, weight,
200–250 g) were purchased from Shanghai SLAC Laboratory Animal Co.,
Ltd. (Shanghai, China) and allowed 1 week to adapt to the
laboratory environment prior to experiments. Rats were housed at
50±10% humidity, 24±2°C and a 12-h light/dark cycle with ad
libitum access to standard chow and water.
Rats were randomly divided into seven experimental
groups: Control group, sham group (saline + sterile distilled
water), model group (Aβ1-42 + sterile distilled water), LGZG 4.8
group (Aβ1-42 + 4.8 g/kg LGZG), LGZG 2.4 group (Aβ1-42 + 2.4 g/kg
LGZG), LGZG 1.2 group (Aβ1-42 + 1.2 g/kg LGZG) and positive control
group [Aβ1-42 + 2 mg/kg donepezil hydrochloride (DNP)]. Each group
contained 8 rats.
Reagents and administration
procedures
Poria (Anhui, China), Cinnamomi ramulus (Guangdong,
China), Atractylodis macrocephalae rhizoma (Jiangsu, China), and
Glycyrrhizae radix et rhizoma (Xinjiang, China) were purchased from
Jiangsu Lianshui Pharmaceutical Co., Ltd. (Lianshui, China), and
prepared in a dry weight ratio of 4:3:2:2 for the production of
LGZG. The constituents were then mixed with ×10 the quantity of
distilled water, decocted for 2 h and again for 1 h,
pressure-filtered, concentrated using a rotating evaporator to
obtain a 1 g/ml solution and stored at 4°C prior to further use.
Aβ1-42 (cat no. 107761-42-2) and monoclonal mouse GAPDH (cat no.
9001-50-7) antibodies were obtained from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). Antibodies against LRP-1 (cat no. 64099), RAGE
(cat no. 4679), phosphorylated (p)-NF-κB (cat no. 3033), NF-κB
inhibitor α (IκBα) (cat no. 4812), p-IκBα (cat no. 2852),
extracellular signal-regulated kinase (Erk)1/2 (cat no. 9102),
p-Erk1/2 (cat no. 4370), p38 (cat no. 9212) and p-p38 (cat no.
9211), as well as horseradish peroxidase (HRP)-conjugated
anti-mouse (cat no. 7072) or anti-rabbit (cat no. 7071) IgG
secondary antibodies were obtained from Cell Signaling Technology,
Inc. (Danvers, MA, USA). Rabbit anti-rat antibodies specific
against LRP-1 and RAGE were obtained from Beyotime Institute of
Biotechnology, Haimen, China (cat no. A0208). ELISA kits for TNF-α
(cat no. JEB-13718), IL-1β (cat no. JEB-13503), IL-6 (cat no.
JEB-13729) and Aβ (cat no. JEB-15109) were purchased from Nanjing
KJI Biological Technology Development Co., Ltd. (Nanjing, China).
Other chemicals were purchased from Merck KGaA.
Following anesthesia with intraperitoneal injection
of 0.45% pentobarbital sodium (40 mg/kg), rats were secured on a
stereotaxic device. Either peptides (Aβ1-42; 10 µg; Sigma-Aldrich,
Merck KGaA) or vehicle (saline) was administered into the bilateral
hippocampus CA1, according to the rat brain in stereotaxic
coordinates (−3.0 mm posterior to bregma, ±2.0 mm lateral to
midline, and −3.0 mm dorsal to ventral dura). These were supplied
progressively within a 5-min duration, followed by a cessation
period of 5 min, as described previously (16–18).
Rats were subsequently administered with 200,000 U/ml penicillin-G
once daily for 3 days following surgery. Following the procedure,
LGZG (1.2, 2.4 and 4.8 g/kg), 2 mg/kg DNP and vehicle (sterile
distilled water) were administered orally to the rats twice daily
by gavage for 30 days.
Morris water maze test
The water-maze test was performed 25 days following
drug administration. This test was performed in a circular pool
(180 cm diameter ×60 cm height). A hidden circular platform was
submerged 2 cm below the water level in one pool quadrant. Rats
were allowed two trial sessions each day for 4 successive days with
an intermission of 5 h between trials. A video tracking system was
used to view the complete procedure. When the rat had located the
platform successfully, it was permitted to remain there for 10 sec.
Rats was physically positioned on the platform for 10 sec in cases
where they did not succeed in locating the platform within 90 sec,
wherein escape latency was marked as 120 sec. A single probe trial
was performed on the fifth day.
Hematoxylin-eosin (H&E)
staining
Rats were anesthetized with 0.45% pentobarbital
sodium, then sacrificed via perfusion transcardially with 0.1 mol/l
PBS (pH 7.4) and 4% paraformaldehyde. Sections of the hippocampi
were harvested and fixed in 4% formalin at 4°C for ≥72 h,
dehydrated in graded alcohol series, and embedded in paraffin.
Specimens were cut in 5-µm sections on a rotary microtome and
mounted on 3-aminopropyltriethoxysilane-coated glass slides, then
stained at 4°C for 20 min with H&E and light microscopic
examination for histopathological alteration was performed (Olympus
BH2; Olympus Corporation, Tokyo, Japan) as described previously
(19,20). Images were captured using a Nikon
Coolpix 990 camera (Nikon Corporation, Tokyo, Japan) with an
original magnification of ×200.
Immunohistochemistry of LRP-1 and
RAGE
For immunohistochemical analysis, brain tissue was
fixed in 4% paraformaldehyde at 4°C for 24 h, embedded in paraffin,
and then cut into 4 µm-sections, hydrated and incubated in 3%
hydrogen peroxide at room temperature for 10 min. The sections were
washed three times with PBS and then stored at 4°C in PBS
supplemented with 3% bovine serum albumin (cat no. B2064,
Sigma-Aldrich, Merck KGaA) for 30 min. Sections were then incubated
with antibodies against LRP-1 (1:200) or RAGE (1:200) overnight at
4°C, and a negative control was incubated with PBS. These sections
were then exposed to biotinylated universal secondary antibodies
(1:5,000; cat no. A0208; Beyotime Institute of Biotechnology) for 1
h and subsequently to streptavidin biotin horseradish peroxidase
solution (1:1,000) at room temperature for 1 h, following washing
with PBS. And subsequently stained with 3,3′-diaminobenzidine
solution for 45 sec at 37°C. Sections were counterstained with
hematoxylin at room temperature for 20 sec. Graded alcohols were
then used to dehydrate dehydrate section, and they were
subsequently fixed in neutral balsam (Sinopharm Chemical Reagent
Co., Ltd., Shanghai, China) at 60°C for 30 min. LRP-1 and RAGE
staining was evaluated using Image-Pro Plus 6.0 software (Media
Cybernetics, Inc., Rockville, MD, USA).
ELISA of TNF-α, IL-1β, IL-6, and
Aβ
Rats were decapitated 60 min after the behavioral
tests. Brains were immediately harvested, weighed and homogenized
using 0.9% ice-cold saline water. The homogenate was centrifuged at
3,000 × g for 10 min at 4°C and the supernatants were harvested and
served at 4°C prior to ELISA analysis. The expressions of TNF-α,
IL-1β, IL-6, and Aβ in brain tissues of rats were measured using
ELISA kits according to the manufacturer's instructions.
Western blot analysis
To determine the expression levels of LRP-1, RAGE,
p-NF-κB, IκBα, p-IκBα, Erk1/2, p-Erk1/2, p38, p-p38 in the brain,
western blotting was performed as described previously (20). The rat brain tissues were
homogenized (centrifuged at 14,000 × g, 4°C, 15 min), washed with
PBS, and incubated in lysis buffer (cat no. P0013; Beyotime
Institute of Biotechnology) containing protease inhibitor cocktail
(cat no. P8330, Sigma-Aldrich; Merck KGaA) to isolate brain protein
samples. A total of 30 µg protein samples were separated by 10%
SDS-PAGE gels and blotted onto nitrocellulose membranes with TBST
buffer (10 mM Tris-HCl, 150 mM NaCl, 0.1% v/v Tween-20) containing
5% nonfat milk for 1 h at room temperature. Membranes were
incubated with primary antibodies against LRP-1, RAGE, p-NF-κB,
IκBα, p-IκBα, Erk1/2, p-Erk1/2, p38, and p-p38 (1:1,000) at 4°C
overnight, with GAPDH (1:1,000) was used as loading control.
Membranes were subsequently washed three times in 0.05% Tween 20 in
TBS for 5 min and then incubated with HRP-conjugated secondary
antibodies (1:5,000) for 1 h at room temperature. Enhanced
chemiluminescence (cat no. NEL103E001EA; PerkinElmer, Inc.,
Waltham, MA, USA) was used to detect the signals. Quantity One
software version 4.62 (Bio-Rad Laboratories, Inc., Hercules, CA,
USA) was used for densitometry analysis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol reagent (cat
no. 15596026; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol and reverse transcribed to produce cDNA
using a TaqMan reverse transcription reagent kit (Thermo Fisher
Scientific, Inc.). cDNA was then amplified via PCR with the
following primers: RAGE, forward 5′-GACCACTCCTACCTATTCCT-3′ and
reverse 5′-TCCACCTTCAGGCTCAACCA-3′; LRP-1, forward
5′-CGTCACTTACATCAACAACC-3′, and reverse 5′-CAGCCATTCACATTTCTTGC-3′;
and GAPDH, forward 5′-GAGCTGAACGGGAAACTCAC-3′, and reverse
5′-GGTCTGGGATGGAAACTGTG-3′. The thermal cycling conditions included
5 min at 94°C followed by 32 cycles of 30 sec at 94°C, 45 sec at
55°C, and 45 sec at 72°C. The abundance of the mRNA expression
level was measured according to the 2−∆∆Cq method as
described previously (21).
Statistical analysis
Data are expressed as the mean + standard deviation.
Statistical analysis was performed using the SPSS 19.0 (IBM Corp.,
Armonk, NY, USA). Statistical differences between groups were
determined using one-way analysis of variance with Bonferroni's
multiple comparison test using software EZR version 3.1.2 (Jichi
Medical University, Tochigi, Japan; www.jichi.ac.jp/saitama-sct/SaitamaHP.files/statmedEN.html).
P<0.05 was considered to indicate a statistically significant
difference.
Results
LGZG ameliorates memory deficits of AD
rats
As presented in Fig.
1A, the mean escape latency in the Morris water maze declined
progressively in all groups throughout the training period. No
significant difference was observed between the control and
sham-operated groups. Aβ-treated rats spent significantly more
retentive time on reaching the platform vs. controls (P<0.01),
which indicated notable cognitive damage in Aβ-treated rats.
Furthermore, this increase in escape latency was significantly
ameliorated by LGZG (1.2 g/kg: day 3, P<0.05; 2.4 g/kg: day 2
and 4, P<0.05; 4.8 g/kg: day 1, P<0.05, day 2–4, P<0.01
vs. the model) and donepezil (day 1, P<0.05; day 2–4, P<0.01
vs. the model) treatment. The swim routes followed by the rats
during the second trial on day 2 and day 4 are illustrated in
Fig. 1B. The rats seemed to show
the tendency to enter all four quadrants of the pool on day 2. On
day 4, the control rats were observed to swim in the direction of
the platform, whereas Aβ-treated rats took lengthier swimming
routes. In the probe test, as shown in Fig. 1C and D, the control and sham
operated rats spent more time in the target quadrant (26.88±5.41
and 26.75±5.55 sec, respectively) and exhibited greater crossing
times (6.13±1.25 and 5.25±1.03 sec, respectively) than the model
group rats (15.13±3.04 sec, P<0.01 vs. control; 1.88±0.83 sec,
P<0.01 vs. control). When compared with the model group rats,
LGZG (4.8 and 2.4 g/kg) and donepezil treated rats exhibited
increased crossing times (P<0.01, P<0.05 and P<0.01,
respectively). As for time in the target quadrant, a noteworthy
enhancement was observed in the LGZG group at a dose of 2.4
(P<0.05) and 4.8 g/kg (P<0.01) as well as the donepezil group
(P<0.01). This data suggests that LGZG is able to improve the
memory and learning ability of the Aβ-induced model rats in the
Morris water maze test.
LGZG improves brain neuronal
damage
In the present study, no neuronal damage was
observed in the rats in the control and sham groups. However, rats
in the model group exhibited marked neuronal loss in the
hippocampus. HE staining also indicated that the number of neurons
in the LGZG (4.8 g/kg) and donepezil rats was markedly increased
vs. the model (Fig. 2). These
results indicate that LGZG has a neuroprotective effect on the
nerve cells of the brain in AD rats.
Effects of LGZG on levels of TNF-α,
IL-1β, IL-6, and Aβ1-42 in the brain
The results of ELISA analysis (Fig. 3) demonstrate the anti-inflammatory
and neuroprotective effects of LGZG in rats with AD. There was a
significant increase in the levels of TNF-α, IL-1β, IL-6, and
Aβ1-42 in the model group (P<0.01 vs. control). LGZG at doses of
4.8 (P<0.01) and 2.4 g/kg (P<0.05), as well as donepezil
(P<0.01), significantly decreased the levels of TNF-α, IL-1β,
IL-6, and Aβ1-42 compared with the model. There were significant
differences in the levels of IL-6 between the group of LGZG at a
dose of 1.2 g/kg (P<0.05) and the model group. Notably, LGZG at
1.2 g/kg demonstrated a tendency to decrease pro-inflammatory
cytokines levels (TNF-α and IL-1β) levels, but no significant
difference was observed. These findings suggest that LGZG is able
to inhibit neuroinflammation and decrease Aβ1-42 levels in the
brain of AD rats.
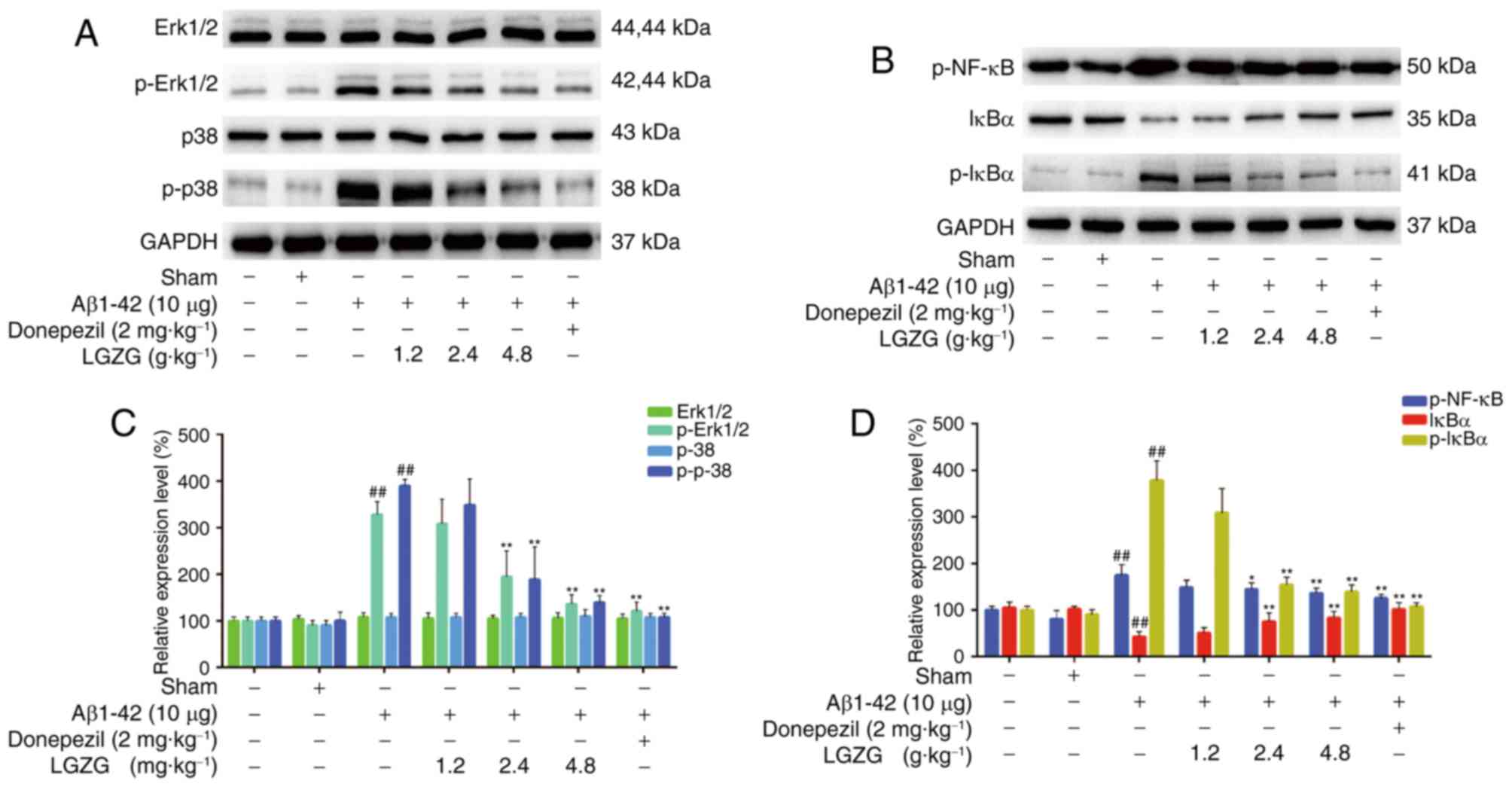
LGZG modulates the expression of MAPK
and the NF-κB pathway in the brain
To evaluate the expression of MAPK and the NF-κB
pathway, the protein levels in the brain of the rats were analyzed
using western blot analysis. As presented in Fig. 4, Aβ1-42 administration
significantly increased the protein levels of p-Erk, p-p38, p-NF-κB
and p-IκBα (P<0.01 vs. control) in rats, which indicated
activation of MAPK and the NF-κB pathway in AD rats. However, LGZG
treatment significantly ameliorated the expression levels of
p-Erk1/2, and p-p38 at 2.4 and 4.8 g/kg (P<0.01) in the brain of
AD rats, compared with the model. Furthermore, the protein levels
of p-NF-κB and p-IκBα at 2.4 (p-NF-κB, P<0.05; p-IκBα,
P<0.01) and 4.8 g/kg (P<0.01) LGZG were also significantly
downregulated compared with the model. These results suggested that
LGZG is able to inhibit MAPK and NF-κB signaling in the brain of AD
rats.
Effect of LGZG on the expression
levels of RAGE and LRP-1 in the brain
RAGE and LRP-1 are essential in controlling Aβ
balance in the brain (21). In the
present study, the expression of RAGE and LRP-1 in the brain of the
rats were analyzed by western blot analysis, RT-qPCR and
immunohistochemistry. Western blot analysis (Fig. 5A) and RT-qPCR (Fig. 5B) indicated an increased expression
of LRP-1, and a decreased expression of RAGE in the brains of LGZG
rats compared with model rats. As presented in Fig. 6, the mean densities of LRP-1 were
lower (Fig. 6A and C), whereas the
mean densities of RAGE were significantly higher (Fig. 6B and D) in the brain of model rats
compared with controls. When compared with the model group, LGZG
significantly downregulated the reactive expression levels of RAGE
and upregulated the reactive expression levels of LRP-1 (2.4 g/kg,
P<0.05; 4.8 g/kg, P<0.01).
Discussion
AD is one of the most profound neurodegenerative
disease in elderly people, which affects cognition, behavior and
function (22). In the present
study, Aβ administration significantly decreased the ability of
learning and memory and induced neuropathological changes in rats,
which is in accordance with the results of the previous study
(23). Accordingly, it was
demonstrated in the present study that LGZG treatment significantly
ameliorated learning and memory and Aβ-induced pathologic changes
in rats. These results above suggested that the administration of
LGZG exhibits neuroprotective effects in AD.
It was recently verified that Aβ accumulation has a
key role in the mechanism of neuron damage, and learning and memory
function in AD (24). Aβ levels
increase significantly in rats following Aβ injection, which
indicated the accumulation of Aβ in this model. One explanation is
the enhanced circulation in brain transportation and the impaired
brain to-blood transportation of Aβ across the BBB, which have been
observed in AD patients and rat models (25,26).
In accordance with these reports, the present data demonstrated
that RAGE, which is a primary transporter of Aβ across the BBB into
the brain, was upregulated following Aβ administration. Conversely,
LRP-1, which is the transporter of Aβ out of the brain, was
downregulated in AD rats. Furthermore, these changes were
ameliorated by treatment with LGZG. Therefore, these results
indicated that LGZG treatment decreased the accumulation of Aβ in
AD rats, possibly by regulating the transport receptors of Aβ.
In addition, RAGE ligation by Aβ is able to activate
multiple signaling pathways, including MAPK and the NF-κB pathway
(9). In the present study, it was
reported that AD rats exhibited significantly higher
phosphorylation levels of MAPK and NF-κB, which is consistent with
previous studies (10).
Subsequently, marked increases of pro-inflammatory cytokines were
observed in this animal model. However, previous in vitro
study have revealed the anti-inflammatory effect of LGZG (−14). In
accordance with these studies, the present results demonstrated
that LGZG significantly inhibited the activation of MAPK and NF-κB,
and reduced the Aβ-induced elevated levels of TNF-α, IL-1β and IL-6
in AD rats, which suggested that the anti-inflammatory effect may
contribute to the protective effects of LGZG in AD rats. These
findings indicated that LGZG attenuates Aβ-induced AD partially via
suppressing the activation of RAGE/MAPK and the NF-κB pathway.
Donepezil is an acetylcholinesterase/cholinesterase
inhibitor, which is considered as the standard treatment of AD
(27). Donepezil has been
demonstrated to significantly ameliorate memory-associated
behavioral deficits and decrease Aβ production (28). In the present study, it was
demonstrated that the protective effects of LGZG on learning and
memory deficits, as well as neuronal impairment were comparable to
those of donepezil. Previous studies have reported that donepezil
has cholinergic side effects (29)
and induces sleep disturbances (30) in patients with AD. Notably, no
obvious side effects or toxic reactions were observed in the groups
treated with LGZG. Therefore, the present study suggests that LGZG
may be considered for treatment of AD.
In summary, the present study demonstrated the
protective effect of LGZG in AD rats. LGZG promoted the ability of
learning and memory, and reduced neuron damage and inflammation in
an Aβ-induced AD model in rats. The mechanism may be associated
with the regulation of Aβ transportation, and inhibition of
RAGE/MAPK and NF-κB signaling by LGZG. Therefore, the present study
elucidates the potential pharmacological application of LGZG for
the treatment of AD.
Acknowledgements
The present study was financially supported by the
National Natural Science Foundation of China (grant no. 81503485)
and the Natural Science Foundation of Jiangsu Province (grant no.
BK20161047).
References
|
1
|
Ezra A, Rabinovich-Nikitin I,
Rabinovich-Toidman P and Solomon B: Multifunctional effect of human
serum albumin reduces Alzheimer's disease related pathologies in
the 3xTg mouse model. J Alzheimers Dis. 50:175–188. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Keaney J, Walsh DM, O'Malley T, Hudson N,
Crosbie DE, Loftus T, Sheehan F, McDaid J, Humphries MM, Callanan
JJ, et al: Autoregulated paracellular clearance of amyloid-β across
the blood-brain barrier. Sci Adv. 1:e15004722015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xi YD, Li XY, Ding J, Yu HL, Ma WW, Yuan
LH, Wu J and Xiao R: Soy isoflavone alleviates Aβ1-42-induced
impairment of learning and memory ability through the regulation of
RAGE/LRP-1 in neuronal and vascular tissue. Curr Neurovasc Res.
10:144–156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Do TM, Dodacki A, Alata W, Calon F,
Nicolic S, Scherrmann JM, Farinotti R and Bourasset F:
Age-dependent regulation of the Blood-brain barrier Influx/Efflux
equilibrium of amyloid-β peptide in a mouse model of Alzheimer's
disease (3xTg-AD). J Alzheimers Dis. 49:287–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Matsumoto K, Chiba Y, Fujihara R, Kubo H,
Sakamoto H and Ueno M: Immunohistochemical analysis of transporters
related to clearance of amyloid-β peptides through
blood-cerebrospinal fluid barrier in human brain. Histochem Cell
Biol. 144:597–611. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Deane R, Singh I, Sagare AP, Bell RD, Ross
NT, LaRue B, Love R, Perry S, Paquette N, Deane RJ, et al: A
multimodal RAGE-specific inhibitor reduces amyloid beta-mediated
brain disorder in a mouse model of Alzheimer disease. J Clin
Invest. 122:1377–1392. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Erickson MA, Hartvigson PE, Morofuji Y,
Owen JB, Butterfield DA and Banks WA: Lipopolysaccharide impairs
amyloid β efflux from brain: Altered vascular sequestration,
cerebrospinal fluid reabsorption, peripheral clearance and
transporter function at the blood-brain barrier. J
Neuroinflammation. 9:1502012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ko SY, Ko HA, Chu KH, Shieh TM, Chi TC,
Chen HI, Chang WC and Chang SS: The possible mechanism of advanced
glycation end products (AGEs) for Alzheimer's disease. PLoS One.
10:e01433452015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang X, Yu S, Hu JP, Wang CY, Wang Y, Liu
HX and Liu YL: Streptozotocin-induced diabetes increases amyloid
plaque deposition in AD transgenic mice through modulating
AGEs/RAGE/NF-kB pathway. Int J Neurosci. 124:601–608. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lv C, Wang L, Liu X, Yan S, Yan SS, Wang Y
and Zhang W: Multi-faced neuroprotective effects of geniposide
depending on the RAGE-mediated signaling in an Alzheimer mouse
model. Neuropharmacology. 89:175–184. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Di BB, Li HW, Li WP, Shen XH, Sun ZJ and
Wu X: Pioglitazone inhibits high glucose-induced expression of
receptor for advanced glycation end products in coronary artery
smooth muscle cells. Mol Med Rep. 11:2601–2607. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xiong MQ: Theory on Exogenous Febrile
Disease. China Press of Traditional Chinese Medicine; Beijing:
2007
|
|
13
|
Yu B, Zhou C, Zhang J, Ling Y, Hu Q, Wang
Y and Bai K: Latest study on the relationship between pathological
process of inflammatory injury and the syndrome of spleen
deficiency and fluid retention in Alzheimer's disease. Evid Based
Complement Alternat Med. 2014:7435412014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sang F: Experimental research on the
mechanism of Alzheimer's disease. J Tradit Chin Med. 6:685–687.
2011.
|
|
15
|
National Research Council: Guide for the
Care and Use of Laboratory Animals: Eighth EditionGuide for the
Care & Use of Laboratory Animals. The National Academies Press;
pp. 1072–1073. 2010
|
|
16
|
Maurice T, Lockhart BP and Privat A:
Amnesia induced in mice by centrally administered beta-amyloid
peptides involves cholinergic dysfunction. Brain Res. 706:181–193.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cheng YF, Wang C, Lin HB, Li YF, Huang Y,
Xu JP and Zhang HT: Inhibition of phosphodiesterase-4 reverses
memory deficits produced by Aβ25-35 or Aβ1-40 peptide in rats.
Psychopharmacology (Berl). 212:181–191. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kwon SH, Lee HK, Kim JA, Hong SI, Kim SY,
Jo TH, Park YI, Lee CK, Kim YB, Lee SY and Jang CG: Neuroprotective
effects of Eucommia ulmoides, Oliv. Bark on amyloid beta
(25–35)-induced learning and memory impairments in mice. Neurosci
Lett. 487:123–127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Klunk WE, Jacob RF and Mason RP:
Quantifying amyloid beta-peptide (Abeta) aggregation using the
Congo red-Abeta (CR-abeta) spectrophotometric assay. Anal Biochem.
266:66–76. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Eisele YS, Obermüller U, Heilbronner G,
Baumann F, Kaeser SA, Wolburg H, Walker LC, Staufenbiel M,
Heikenwalder M and Jucker M: Peripherally applied Abeta-containing
inoculates induce cerebral beta-amyloidosis. Science. 330:980–982.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Golden HL, Agustus JL, Nicholas JM, Schott
JM, Crutch SJ, Mancini L and Warren JD: Functional neuroanatomy of
spatial sound processing in Alzheimer's disease. Neurobiol Aging.
39:154–164. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu B: Study on the relationship between
Alzheimer's disease and the syndrome of spleen deficiency and fluid
retention based on Lingguizhugantang's intervention of Aβ-induced
inflammatory injury (unpublished PhD dissertation). Nanjing
University of TCM. 2015.
|
|
24
|
Harrington KD, Lim YY, Gould E and Maruff
P: Amyloid-beta and depression in healthy older adults: A
systematic review. Aust N Z J Psychiatry. 49:36–46. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jeynes B and Provias J: Evidence for
altered LRP/RAGE expression in Alzheimer lesion pathogenesis. Curr
Alzheimer Res. 5:432–437. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Deane R, Bell RD, Sagare A and Zlokovic
BV: Clearance of amyloid-beta peptide across the blood-brain
barrier: Implication for therapies in Alzheimer's disease. CNS
Neurol Disord Drug Targets. 8:16–30. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Christensen DD: Higher-dose (23 mg/dayay)
donepezil formulation for the treatment of patients with
moderate-to-severe Alzheimer's disease. Postgrad Med. 124:110–116.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nordberg A: Mechanisms behind the
neuroprotective actions of cholinesterase inhibitors in Alzheimer
disease. Alzheimer Dis Assoc Disord. 20 2 Suppl 1:S12–S18. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rogers SL, Farlow MR, Doody RS, Mohs R and
Friedhoff LT: A 24-week, double-blind, placebo-controlled trial of
donepezil in patients with Alzheimer's disease. Donepezil Study
Group. Neurology. 50:136–145. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Agboton C, Mahdavian S, Singh A, Ghazvini
P, Hill A and Sweet A: Impact of nighttime donepezil administration
on sleep in the older adult population: A retrospective study.
Mental Health Clinician. 4:257–259. 2014. View Article : Google Scholar
|