Introduction
Human infertility is a major worldwide issue
affecting 15% of couples of reproductive age. Human infertility is
associated with genetic and non-genetic causes. Chromosomal
abnormalities and gene mutations are frequently found in infertile
men, particularly in those with a low sperm count, and also in
women with low quality oocytes (1–3).
Gametogenesis is a complex biological process that involves
producing cells for sexual reproduction via both meiotic and
mitotic cell division steps. The feature of meiosis is the
reduction of the DNA content by half through two cell divisions,
resulting in germ cells. At the molecular level, spermatogenesis is
highly organized and involves the expression and interaction of
numerous overexpressed specific genes. Abnormal translocation
carriers may result in disorders of gene expression, meiotic
arrest, failure of spermatogenesis and infertility (4–6).
During this process, proteins and protein interactions may have a
universal role in gametogenesis. Scaffold proteins act as important
modulators of a variety of physiological functions based on protein
interactions. Ran-binding protein M (RanBPM), also termed Ranbp9,
is a scaffold protein belonging to the ran-binding proteins
(ranBPs) (7). RanBPM is a
multimodular protein containing a consensus SPRY domain, a CRA
domain, a lissencephaly type-1-like homology (LisH) motif, a
C-terminal to LisH (CTLH) domain and a proline-rich SH3-binding
module. SPRY domains are protein-protein interaction modules that
were initially discovered in the ryanodine receptor (8,9). The
CRA motif is also involved in protein-protein interactions
(10), whereas the LisH motif is
involved in protein dimerization and microtubule binding (11–13).
The CTLH domain function is unknown and is frequently found
adjacent to the LisH domain in proteins involved in microtubule
dynamics, cell migration, nucleokinesis and chromosome segregation,
and has been previously identified in various proteins associated
with RanBPM (14).
RanBPM is component of a large protein complex in
which it functions as an adaptor scaffold protein (15–17).
The full-length RanBPM cDNA was determined and predicted to encode
a protein of 729 amino acids. The human sequence shares 95%
sequence identity with the mouse RanBPM sequence. RanBPM was
identified and mapped to human chromosome 6. It is a member of the
RAS superfamily of proteins. The identification of the different
domains of RanBPM may provide important clues in understanding the
underlying molecular mechanisms of this protein in human
infertility. Several previous studies have also suggested that
RanBPM interacts with various proteins and participates in numerous
cellular processes including neuronal morphogenesis (18–20),
cell growth and cell migration signalling (21–24),
regulation of gene transcription (18,25),
apoptosis and apoptotic pathways (21,26,27).
RanBPM-deficient mice study revealed a role for the protein in
gametogenesis, and RanBPM is essential for normal gonad
development, as both male and female RanBPM−/− mice are
sterile (7). However, additional
defects resulting from RanBPM deficiency remain to be investigated.
Although RanBPM has a role in mouse gametogenesis, the exact
expression pattern, cellular localization and physiological
function in human testis and ovaries remain unclear.
In the present study, it was demonstrated that
RanBPM was highly expressed in human testis and ovary tissue, and
differentially expressed during spermatogenesis and oogenesis. In
addition, the RanBPM protein was localized in both the nucleus and
cytoplasm of germ cells, especially primary spermatocytes and
follicle cells. RanBPM was found to interact with a variety of
functional proteins in germ cells, suggesting multiple roles in the
regulation of germ cell proliferation during human spermatogenesis
and oogenesis. These results provide additional support to expand
the understanding of infertility in men, and future studies on
spermatogenesis in individuals that are RanBPM mutation carriers
may improve the understanding of the underlying mechanisms of
fertility.
Materials and methods
Tissue collection and ethics
statement
Testicular and ovary tissue samples were obtained
from men with obstructive azoospermia and infertile women, at the
Peking University Third Hospital. The muscle, heart and kidney
tissues were also obtained from men (aged 25–38 years) at the
Peking University Third Hospital. All experiments were performed in
accordance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals issued by the Peking University
Third Hospital in Beijing. All participants gave their written
informed consent, and the study was approved by the Ethics
Commission of the Medical Faculty of the Peking University Third
Hospital. The protocol was approved by the Institutional Care and
Use Committee of the Peking University Third Hospital (protocol no.
2013SZ021). From August to September 2013, 15 patients (aged 25
weeks of fetal-38 years old) and 9 patients (aged 25 weeks of
fetal-38 years old) agreed to further tests. The participants or
close relatives signed consent forms permitting the collection and
use of all samples used in this study. Chemicals were obtained from
Invitrogen; Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
RNA isolation and semi-quantitative
polymerase chain reaction (PCR)
Total RNA was extracted from human testicular tissue
and ovaries using an RNeasy Mini kit (QiagenGmbH, Hilden, Germany)
and treated with DNase I (Roche Applied Science, Penzberg, Germany)
according to the manufacturer's protocol (28–30).
First-strand cDNA was generated by reverse transcription using the
Superscript III first-strand synthesis system (Invitrogen; Thermo
Fisher Scientific, Inc.) and random hexamer oligonucleotides.
Primers were designed based on sequences in the human genome
database. Subsequent PCR analysis was performed with ExTaqHot-Start
DNA polymerase (cat. no. RR006A; Takara Biotechnology Co., Ltd.,
Dalian, China). All experiments were performed semi-quantitatively
at three different escalation cycles, and representative images are
shown in the results. For PCR, reaction mixture was first heated at
94°C for 2 min. 24 cycles were then carried out with the following
parameters: Denaturing at 94°C for 30 sec, annealing at 58°C for 30
sec, extension at 72°C for 45 sec. Reaction was finished with a
final extension at 72°C for 5 min.
The primer pairs: Forward,
5′-AAGGTCGACACATGAATAGACTACCAGGTTGG-3′ and reverse,
5′-CGCAAGCTTTTCAAATCAGCAGAGCTAGTC-3′, were used to detect the mRNA
expression levels of RanBPM. The products of PCR were separated by
1.5% agarose gel electrophoresis and stained with ethidium bromide
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Relative amounts
of cDNA were normalized against β-actin (forward,
5′-GATGGTGGGAATGGGTCAGAA-3′ and reverse,
5′-TGGCGTGAGGGAGAGCATAGC-3′).
Protein extract preparation and
western blotting
Ovary and testicular tissue samples were homogenised
and lysed in radioimmunoprecipitation assay lysis buffer [50
mMTris-HCl (pH 7.5), 150 mMNaCl, 2 mM EDTA, 1% Triton X-100, 1%
sodium deoxycholate, 0.1% SDS and protease inhibitors], as
previously described (31). Human
tissues were homogenized in lysis buffer containing 20 mMTris-HCl
(pH 8.0), 150 mM NaCl, 2 mM EDTA (pH 8.0) and 1% Nonidet-P40
(NP-40). After centrifuged at 12,000 × g for 30 min at 4°C,
supernatants were diluted with sample buffer A from NE-PER™ Nuclear
and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific,
Inc.) containing 0.5% NP-40, 1 mM DTT and 1X protease inhibitor
(PI) and boiled. A total of 10 µg protein was separated by 7.5%
SDS-PAGE and transferred to a nitrocellulose (cat. no. 10401196;
Sigma-Aldrich; Merck KGaA) or polyvinylidene difluoride (cat. no.
RPN303F; GE Healthcare, Chicago, IL, USA) membranes. Membranes were
blocked at room temperature with 5% (w/v) skimmed milk for 30 min.
After 5 washes with 0.1% PBS with Tween-20, membranes were
incubated with primary antibodies at room temperature for 1 h.
Rabbit anti-RanBPM (1:2,000; cat. no. ab64275; Abcam, Cambridge,
UK) was used as primary antibody, after 3 washes with 0.1% PBST,
horseradish peroxidase-conjugated (1:2,000; cat. no. N100; Thermo
Fisher Scientific, Inc) was incubated for 1 h at room temperature.
β-actin antibody (1:5,000; cat. no. ab8226; Abcam) was used as a
loading control. Protein expression levels were analysed using
Image-Pro plus software version 5.1 (Media Cybernetics, Inc.,
Rockville, MD, USA) and ECL reagent (cat. no. RPN2209; GE
Healthcare).
Generation of antibodies and
immunofluorescence
Immunohistochemistry was performed using standard
procedures, as previously described (32,33).
Bouin's fixed human testicular tissue and ovaries were dehydrated
and embedded in paraffin at room temperature. Human testis and
ovary tissue were fixed immediately in 4% paraformaldehyde in PBS
for 15 min at room temperature. Then, 8 µm sections of testicular
tissue and ovaries were incubated in citrate buffer for 30 min at
99°C for antigen retrieval and incubated in 1%
H2O2 for 10 min at room temperature.
Non-specific binding was blocked with 20% normal goat serum in TBS
for 30 min to eliminate background. Sections were washed 3 times, 5
min each, in TBS (0.05 M Tris, 0.15 M NaCl, pH 7.6) plus 0.1%
Tween-20 (TBST). After deparaffinization and rehydration, the
sections were incubated with a rabbit anti-RanBPM antibody (1:50;
cat. no. ab64275; Abcam) in TBS plus 5% horse serum or 5%
pre-immune rabbit serum (used instead of the primary antibody as a
negative control) for 1 h at room temperature. Sections were washed
3 times, 5 min each in TBST between the incubation with antibodies.
A secondary antibody (1:300; cat. no. a2556; monoclonal mouse
anti-rabbit IgG; Sigma-Aldrich; Merck KGaA) and a third antibody
(1:300; cat. no. A0216; HRP-labelledgoat anti-mouse IgG; Beyotime
Institute of Biotechnology, Haimen, China) in TBS plus 5% horse
serum, was added to the sections for 45 min at room temperature.
After washing the section in TBST, the protein of interest was
revealed using an VECTASTAINABC kit (cat. no. PK-6101; Vector
laboratories, Ltd., Burlingame, CA, USA) for 30 min and
diaminobenzidine substrate solution (cat. no. P0018; Beyotime).
Sections were examined under a laser confocal microscope (Zeiss
GmbH, Jena, Germany).
Results
RanBPM mRNA and protein expression
levels in human testes and ovaries
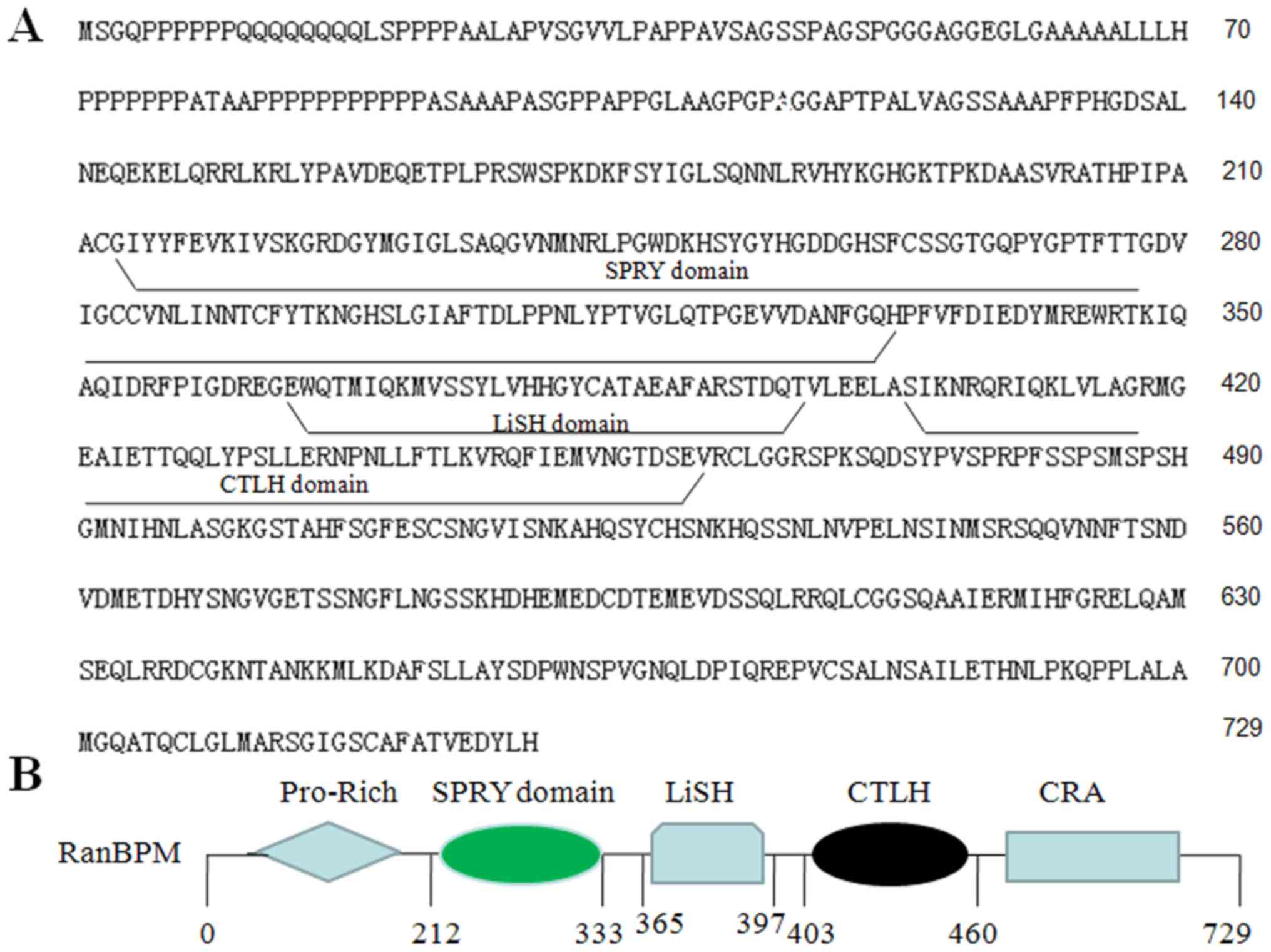
Human RanBPM encodes a protein of 729 amino acids
and consists of a CTLH domain, LisH domain, a SPRY domain, a
pro-rich domain and a CRA domain (Fig.
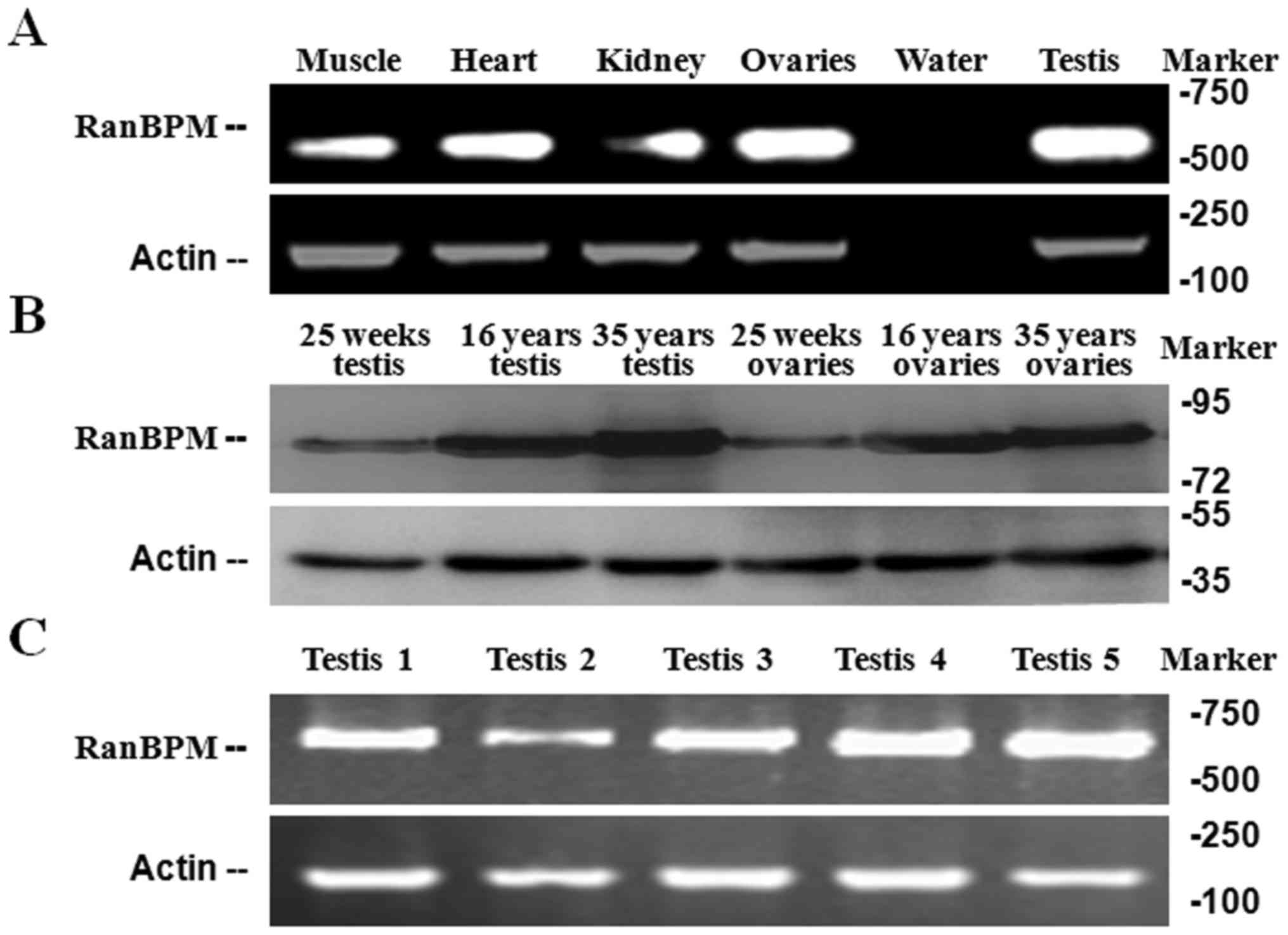
1A and B). RanBPM mRNA and protein expression was investigated
in various male and female tissues by PCR (Fig. 2A) and western blotting (Fig. 2B). RanBPM mRNA was detected in 5
tissues of adult (Fig. 2A) and 5
patients (aged 25–38 years) of testis were performed (Fig. 2C). The RanBPM protein consists of
729 amino acids and had an apparent molecular mass of 78 kDa based
on western blotting. RanBPM mRNA expression was detected in testis
tissue (Fig. 2A), and the RanBPM
protein displayed a similar expression pattern RanBPM protein
expression was not abundant in the testis shortly after birth, but
the levels were higher in adult testis (Fig. 2B), suggesting RanBPM may be
involved in male germ cell development and proliferation.
Furthermore, RanBPM protein was also abundant in adult ovaries
(Fig. 2B).
RanBPM is localized to the nucleus and
cytoplasm of testis
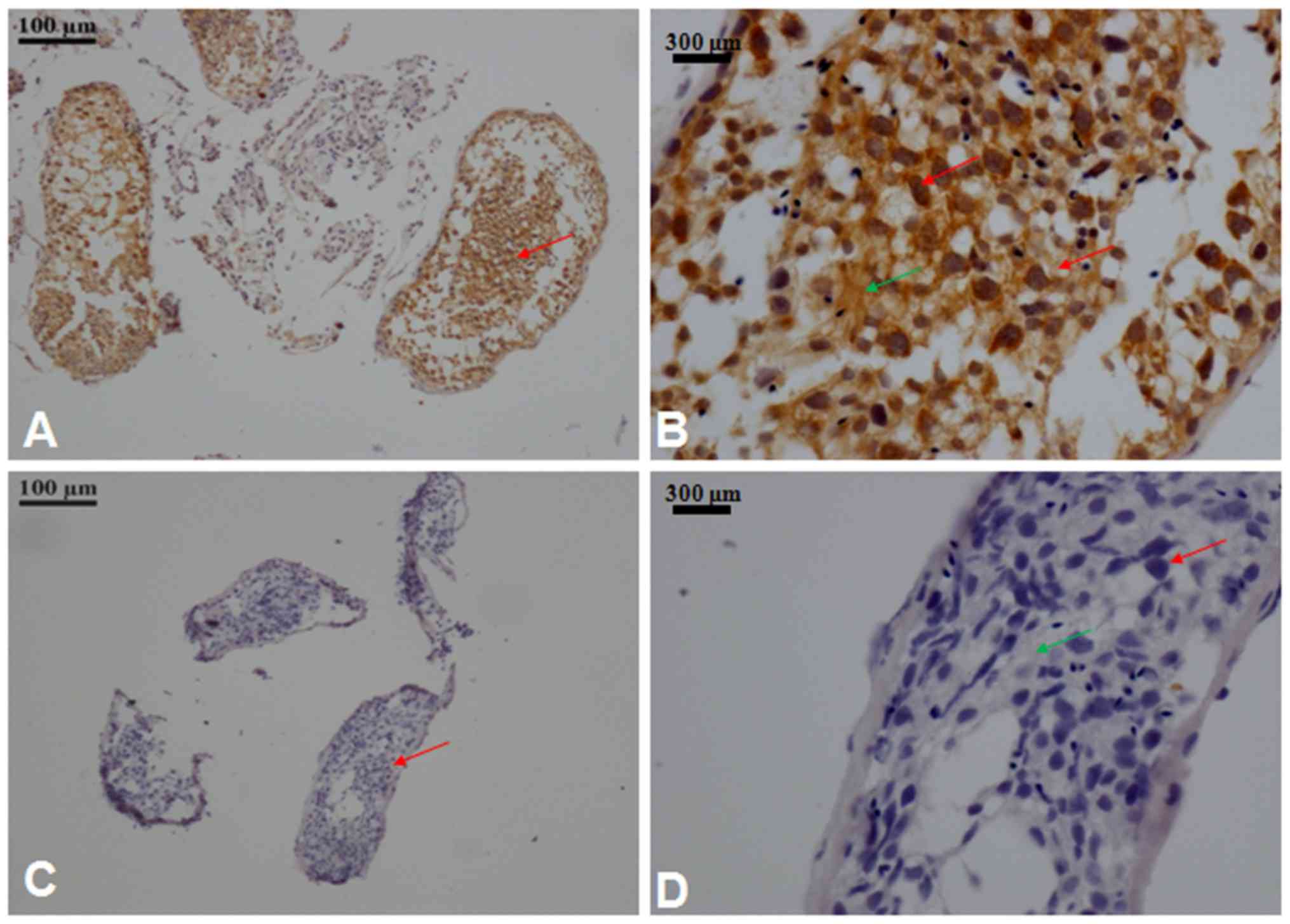
Immunostaining of RanBPM in human testis tissue
sections was performed to determine the distribution in different
germ cell stages (Fig. 3).
Immunostaining revealed the localization of RanBPM in the nucleus
and cytoplasm of germ cells. RanBPM expression was detected in both
spermatogonia and sertoli cells that support the spermatogenic
process (Fig. 3B).
RanBPM is expressed in human
ovaries
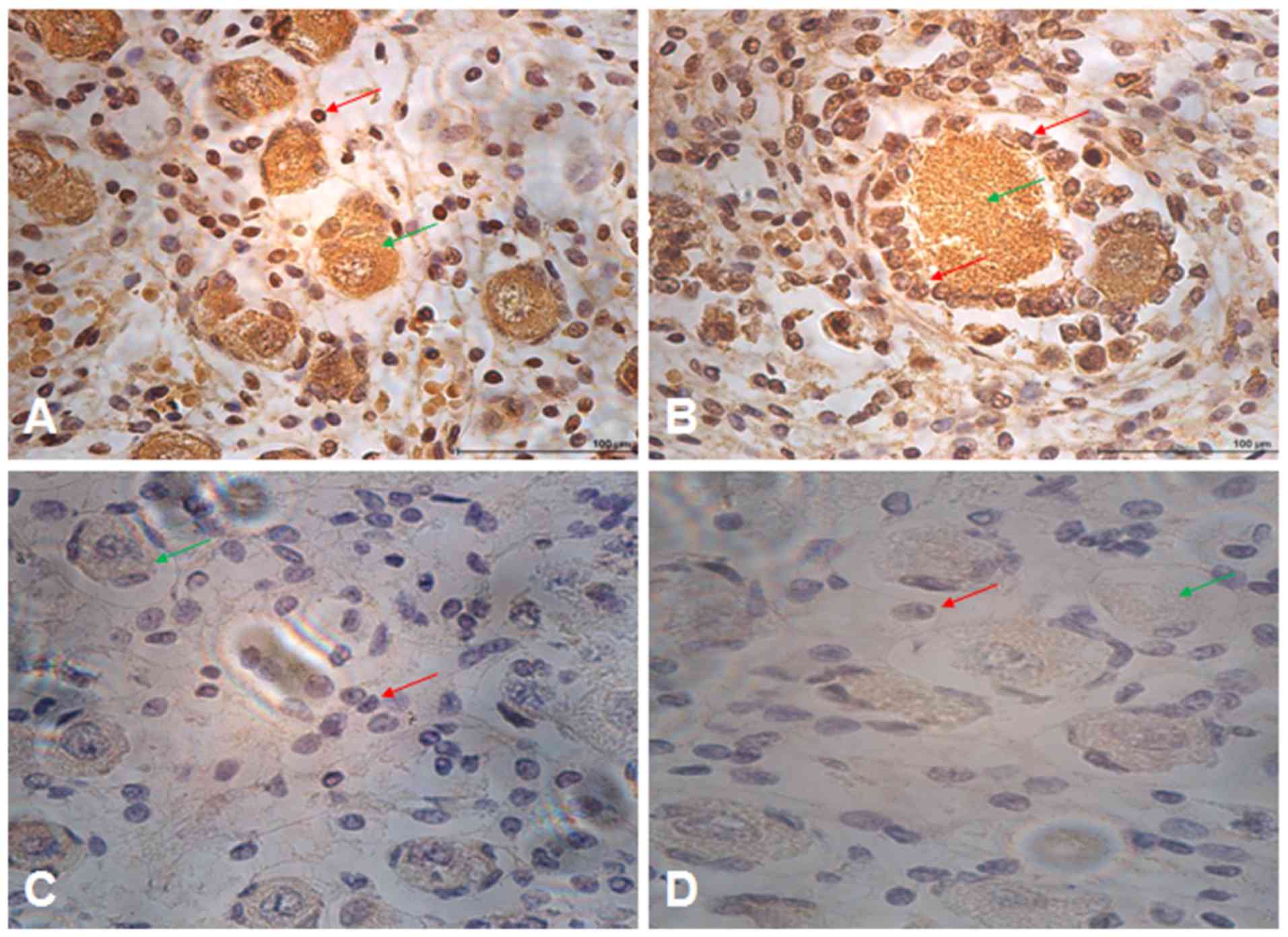
Oogenesisis a complex developmental process that
involves sophisticated regulation of gene expression in ovaries. To
investigate the expression of RanBPM in oogenesis, immunostaining
was performed on adult ovary sections (Fig. 4). The RanBPM protein was found to
be present in the primordial follicle (Fig. 4A) of ovary sections RanBPM appeared
to be localized the nucleus of follicles (Fig. 4B).
Discussion
Gametogenesis is a complex process by which diploid
cells differentiate into haploid gametes during sexual
reproduction. During this process, a large number of genes and
proteins involved in fertility are highly regulated in males and
females (31,32). Gametes are generated from diploid
germ cells through meiosis, during which homologous chromosomes
must align, pair, synapse, recombine and separate to form haploid
cells. This complicated process involves a series of highly
regulated molecular events that are dependent on complex
interactions between a number of distinct genes and proteins
(33,34). Gametogenesis is highly organized
and involves the expression and interaction of numerous genes
(28,31,33);
however, mammalian gametogenesis remains to be elucidated at the
molecular level. The targeting of candidate genes with proposed
roles in gametogenesis has provided valuable information (29,32).
For example, many genes involved in spermatogenesis in mice are
X-linked (31,33,34)
and are expressed exclusively in males.
A malfunction in the expression of genes such as
RanBPM may result in spermatogenic disorders, although the
diagnostic potential of this gene and its underlying mechanism of
action in human gametogenesis and reproduction are poorly
understood. In the present study, RanBPM was demonstrated to be
abundantly expressed in human tissues (such as kidney, heart and
muscle), especially in the nuclei of testes and ovaries. Although
RanBPM was generally localized within nuclei, its specific
localization in the cytoplasm of round spermatids and sertoli cell
suggested a possible involvement in gametogenesis in male patients.
In addition, the RanBPM protein was present in the primordial
follicle in ovary sections. PCR also revealed that RanBPM was
expressed in follicles in ovary sections. These results suggested
that RanBPM may be involved in human spermatogenesis and
oogenesis.
RanBPM is a scaffold protein belonging to the
Ran-binding protein family, members of which contain a SPRY domain,
CRA domain, LisH motif and CTLH domain. Scaffold proteins act as
important modulators of a variety of protein-mediated physiological
functions. RanBPM is involved in the regulation of the immune
system (35). RanBPM is associated
with cell cycle proteins and regulates the progression of the cell
cycle. Additionally, RanBPM may inhibit protein ubiquitination and
acts as a tumour suppressor. For example, it interacts with the Met
tyrosine kinase receptor and in doing so facilitates activation of
the RAS-extracellular regulated kinase pathway (36). In addition, RanBPM has been
associated with citron kinase, possibly affecting the rate of
mitosis during the production of pyramidal neurons (19). It also binds to the kelch repeat
protein muscle in and affects cell morphology (23). In the immune system, it interacts
with the 2-integrin lymphocyte function-associated antigen-1
(35), the neural cell adhesion
molecule L1, and the neurotrophic receptor, tropomyosin receptor
kinase B (37,38). Human sperm membrane protein-1 and
RanBPM complex localize to the microtubule-organizing centre and
interact with tubulins, and thereby may modulate microtubule
assembly and/or activity (39).
RanBPM has been demonstrated to interact with >50 different
proteins, consistent with a number of important roles in multiple
human tissues.
RanBPM is a scaffold protein due to its ability to
associate with a wide range of proteins (37). In the present study, RanBPM was
localized to the cytoplasm of round spermatids and the nuclei of
sertoli cells. Therefore, it appears to be a nuclear protein in
spermatocytes, but a cytoplasmic protein in round spermatids,
suggesting a role in spermatid cell proliferation. In conclusion,
the present study examined the expression and localization of
RanBPM in human testis and ovary tissues. The results may suggest
that RanBPM has an involvement in spermatogenesis and oogenesis.
RanBPM may be a gametogenesis-associated protein that is abundantly
expressed in the testis and ovary. Given its unique expression
pattern and localization, RanBPM may have multiple roles in the
regulation of gametogenesis. These results also expand on
understanding of the underlying mechanism of spermatogenesis and
oogenesis, and may aid development of treatments for gene
expression-based infertility disorders in men.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81671513), Beijing
Natural Science Foundation (grant no. 7172236), Research Fund from
China Sexology Association (grant no. A75490-03), Excellent Young
Physician Research Fund from Peking University Third Hospital
(grant no. Y75490-02) and the open project of Key Laboratory of
Male Reproductive and Genetics, National Health and Family Planning
Commission (grant no. KF201706).
References
|
1
|
Borgmann J, Tüttelmann F, Dworniczak B,
Röpke A, Song HW, Kliesch S, Wilkinson MF, Laurentino S and Gromoll
J: The human RHOX gene cluster: Target genes and functional
analysis of gene variants in infertile men. Hum Mol Genet.
15:4898–4910. 2016.
|
|
2
|
Zhuang X, Huang J, Jin X, Yu Y, Li J, Qiao
J and Liu P: Chromosome aberrations and spermatogenic disorders in
mice with Robertsonian translocation (11; 13). Int J Clin Exp
Pathol. 7:7735–7743. 2014.PubMed/NCBI
|
|
3
|
Ishizuka M, Ohtsuka E, Inoue A, Odaka M,
Ohshima H, Tamura N, Yoshida K, Sako N, Baba T, Kashiwabara S, et
al: Abnormal spermatogenesis and male infertility in testicular
zinc finger protein Zfp318-knockout mice. Dev Growth Differ.
58:600–608. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Diao R, Fok KL, Chen H, Yu MK, Duan Y,
Chung CM, Li Z, Wu H, Li Z, Zhang H, et al: Deficient human
β-defensin 1 underlies male infertility associated with poor sperm
motility and genital tract infection. Sci Transl Med.
6:249ra1082014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Miyamoto T, Tsujimura A, Miyagawa Y, Koh
E, Namiki M and Sengoku K: Male infertility and its causes in
human. Adv Urol. 2012:3845202012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ji ZY, Sha YW, Ding L and Li P: Genetic
factors contributing to human primary ciliary dyskinesia and male
infertility. Asian J Androl. 19:515–520. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Puverel S, Barrick C, Dolci S, Coppola V
and Tessarollo L: RanBPM is essential for mouse spermatogenesis and
oogenesis. Development. 138:2511–2521. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ponting C, Schultz J and Bork P: SPRY
domains in ryanodine receptors (Ca(2+)-release channels). Trends
Biochem Sci. 22:193–194. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Perfetto L, Gherardini PF, Davey NE,
Diella F, Helmer-Citterich M and Cesareni G: Exploring the
diversity of SPRY/B30.2-mediated interactions. Trends Biochem Sci.
38:38–46. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Menon RP, Gibson TJ and Pastore A: The C
terminus of fragile X mental retardation protein interacts with the
multi-domain Ran-binding protein in the microtubule-organising
centre. J Mol Biol. 343:43–53. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Emes RD and Ponting CP: A new sequence
motif linking lissencephaly, Treacher Collins and
oral-facial-digital type 1 syndromes, microtubule dynamics and cell
migration. Hum Mol Genet. 10:2813–2820. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim MH, Cooper DR, Oleksy A, Devedjiev Y,
Derewenda U, Reiner O, Otlewski J and Derewenda ZS: The structure
of the N-terminal domain of the product of the lissencephaly gene
Lis1 and its functional implications. Structure. 12:987–998. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gerlitz G, Darhin E, Giorgio G, Franco B
and Reiner O: Novel functional features of the Lis-H domain: Role
in protein dimerization, half-life and cellular localization. Cell
Cycle. 4:1632–1640. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nishitani H, Hirose E, Uchimura Y,
Nakamura M, Umeda M, Nishii K, Mori N and Nishimoto T: Full-sized
RanBPM cDNA encodes a protein possessing a long stretch of proline
and glutamine within the N-terminal region, comprising a large
protein complex. Gene. 272:25–33. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kobayashi N, Yang J, Ueda A, Suzuki T,
Tomaru K, Takeno M, Okuda K and Ishigatsubo Y: RanBPM, Muskelin,
p48EMLP, p44CTLH, and the armadillo-repeat proteins ARMC8alpha and
ARMC8beta are components of the CTLH complex. Gene. 396:236–247.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Umeda M, Nishitani H and Nishimoto T: A
novel nuclear protein, Twa1, and Muskelin comprise a complex with
RanBPM. Gene. 303:47–54. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Salemi LM, Almawi AW, Lefebvre KJ and
Schild-Poulter C: Aggresome formation is regulated by RanBPM
through an interaction with HDAC6. Biol Open. 3:418–430. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brunkhorst A, Karlén M, Shi J, Mikolajczyk
M, Nelson MA, Metsis M and Hermanson O: A specific role for the
TFIID subunit TAF4 and RanBPM in neural progenitor differentiation.
Mol Cell Neurosci. 29:250–258. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chang Y, Paramasivam M, Girgenti MJ,
Walikonis RS, Bianchi E and LoTurco JJ: RanBPM regulates the
progression of neuronal precursors through M-phase at the surface
of the neocortical ventricular zone. Dev Neurobiol. 70:1–15.
2010.PubMed/NCBI
|
|
20
|
Togashi H, Schmidt EF and Strittmatter SM:
RanBPM contributes to Semaphorin3A signaling through plexin-A
receptors. J Neurosci. 26:4961–4969. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kramer S, Ozaki T, Miyazaki K, Kato C,
Hanamoto T and Nakagawara A: Protein stability and function of p73
are modulated by a physical interaction with RanBPM in mammalian
cultured cells. Oncogene. 24:938–944. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hafizi S, Gustafsson A, Stenhoff J and
Dahlbäck B: The Ran binding protein RanBPM interacts with Axl and
Sky receptor tyrosine kinases. Int J Biochem Cell Biol.
37:2344–2356. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Valiyaveettil M, Bentley AA, Gursahaney P,
Hussien R, Chakravarti R, Kureishy N, Prag S and Adams JC: Novel
role of the muskelin-RanBP9 complex as a nucleocytoplasmic mediator
of cell morphology regulation. J Cell Biol. 182:727–739. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zou Y, Lim S, Lee K, Deng X and Friedman
E: Serine/threonine kinase Mirk/Dyrk1B is an inhibitor of
epithelial cell migration and is negatively regulated by the Met
adaptor Ran-binding protein M. J Biol Chem. 278:49573–49581. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Poirier MB, Laflamme L and Langlois MF:
Identification and characterization of RanBPM, a novel coactivator
of thyroid hormone receptors. J Mol Endocrinol. 36:313–325. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bai D, Chen H and Huang BR: RanBPM is a
novel binding protein for p75NTR. Biochem Biophys Res Commun.
309:552–557. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mikolajczyk M, Shi J, Vaillancourt RR,
Sachs NA and Nelson M: The cyclin-dependent kinase 11(p46) isoform
interacts with RanBPM. Biochem Biophys Res Commun. 310:14–18. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhuang XJ, Shi YQ, Xu B, Chen L, Tang WH,
Huang J, Lian Y, Liu P and Qiao J: SLX2 interacting with BLOS2 is
differentially expressed during mouse oocyte meiotic maturation.
Cell Cycle. 13:2231–2237. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhuang XJ, Tang WH, Feng X, Liu CY, Zhu
JL, Yan J, Liu DF, Liu P and Qiao J: Trim27 interacts with Slx2, is
associated with meiotic processes during spermatogenesis. Cell
Cycle. 15:2576–2584. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhuang XJ, Lu YQ, Zhang M, Lu SS and Lu
KH: Microisolation and microcloning of bovine X-chromosomes for
identification of sorted buffalo (Bubalus bubalis) spermatozoa.
Anim Reprod Sci. 126:32–36. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhuang XJ, Tang WH, Liu CY, Zhu JL, Feng
X, Yan J, Lian Y, Liu P and Qiao J: Identification and
characterization of Xlr5c as a novel nuclear localization protein
in mouse germ cells. PLoS One. 10:e01300872015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shi YQ, Liao SY, Zhuang XJ and Han CS:
Mouse Fem1b interacts with and induces ubiquitin-mediated
degradation of Ankrd37. Gene. 485:153–159. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhuang XJ, Hou XJ, Liao SY, Wang XX, Cooke
HJ, Zhang M and Han C: SLXL1, a novel acrosomal protein, interacts
with DKKL1 and is involved in fertilization in mice. PLoS One.
6:e208662011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shi YQ, Zhuang XJ, Xu B, Hua J, Liao SY,
Shi Q, Cooke HJ and Han C: SYCP3-like X-linked 2 is expressed in
meiotic germ cells and interacts with synaptonemal complex central
element protein 2 and histone acetyltransferase TIP60. Gene.
527:352–359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Denti S, Sirri A, Cheli A, Rogge L,
Innamorati G, Putignano S, Fabbri M, Pardi R and Bianchi E: RanBPM
is a phosphoprotein that associates with the plasma membrane and
interacts with the integrin LFA-1. J Biol Chem. 279:13027–13034.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang D, Li Z, Messing EM and Wu G:
Activation of Ras/Erk pathway by a novel MET-interacting protein
RanBPM. J Biol Chem. 277:36216–36222. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Murrin LC and Talbot JN: RanBPM, a
scaffolding protein in the immune and nervous systems. J
Neuroimmune Pharmacol. 2:290–295. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yin YX, Sun ZP, Huang SH, Zhao L, Geng Z
and Chen ZY: RanBPM contributes to TrkB signaling and regulates
brain-derived neurotrophic factor-induced neuronal morphogenesis
and survival. J Neurochem. 114:110–121. 2010.PubMed/NCBI
|
|
39
|
Tang X, Zhang J, Cai Y, Miao S, Zong S,
Koide SS and Wang L: Sperm membrane protein (hSMP-1) and RanBPM
complex in the microtubule-organizing centre. J Mol Med (Berl).
82:383–388. 2004. View Article : Google Scholar : PubMed/NCBI
|