Introduction
Diabetic nephropathy (DN) is a complication of
diabetes. It is estimated that ~33% of patients with diabetes
develop end-stage kidney disease (ESKD), which may further progress
to DN (1–3). Diabetic kidney disease (DKD) is the
primary cause of kidney failure. DKD is frequently identified as a
degenerative disease of the glomerulus and occurs due to
alterations of the intrarenal metabolism and structure (4,5).
Recently, the incidence rate of DKD has increased (6). In addition, type 2 diabetes
(T2D)-ESKD has an unfavorable 5-year survival rate at <30%
(7).
Pathogenesis of DKD is multifactorial and
complicated. Inflammation and oxidative stress are identified as
the two critical regulators in DKD. Nuclear factor, erythroid 2
like 2 (Nrf2) is a transcription factor that mediates the
expression levels of antioxidant genes. Bardoxolone methyl, which
may activate the expression of Nrf2, was determined to increase the
estimated glomerular filtration rate in patients with DKD (8). Previous studies also suggested
inflammation as a major pathophysiological mechanism for DKD
progression (9,10).
Although clinical treatments for DKD have not been
progressed, several effective agents and biological molecules have
been identified. For example, metformin extracted from herbal
medicines, may be a useful agent for the control of DKD, as it has
hypoglycemic characteristics and renoprotective activities
(11). Other herbal medicines have
also been identified, such as curcumin and glycosides (12). A recent study revealed that the
JAK-STAT pathway was associated with the development of DKD and
gene expression of JAK2 was increased in glomerular
podocytes and JAK2 inhibitor had an evident reduced effect on
albuminuria, and may be used as a novel therapy of DKD (13).
MicroRNAs (miRNAs) have significant roles in the
regulation of gene expression and cellular activities. A previous
study determined that miR-192 was elevated in DKD via the mediation
of renal fibrosis (14).
Additional important miRNAs for DKD progression have also been
previously identified, including miR-21, miR-29c, miR-200b,
miR-216a and miR-377 (15).
However, comprehensive understanding of the
pathogenesis of DKD is still unclear. A previous study established
a microarray profile, GSE30122, identifying the crucial genes in
DKD samples and highlighted several key molecules that were
differentially expressed in glomeruli and the diabetic tubili,
including cell division cycle 42 and vascular endothelial growth
factor. Inflammation-associated pathways were primarily enriched
for these genes (16). Although
they performed expression validations of these genes, potential
interactions between the differentially-expressed genes (DEGs),
particularly at the protein level, were not investigated further.
Therefore, Tang et al (5)
used this expression profile to perform the weighted gene
co-expression network analysis and identified 10 novel potential
therapeutic targets for DKD, including ETS proto-oncogene 1,
transcription factor, lipopolysaccharide induced TNF factor,
nuclear factor, erythroid derived 2, retinoic acid receptor, γ and
signal transducer and activator of transcription 5A (5). However, regulation from miRNAs was
not taken into consideration. Therefore, the present study
re-analyzed the dataset of GSE30122, and performed a series of
bioinformatics analyses. Following the DEG selection in glomeruli
and tubili, the overlapping DEGs were selected to perform
enrichment analysis and protein-protein interaction (PPI) network
analysis. Additionally, a sub-network was further extracted from
the PPI network. It is of note that the present study identified
the miRNAs that may target these DEGs and constructed an integrated
miRNA-target regulatory network. Using these comprehensive
analyses, the present study aimed to provide novel insight into the
pathogenesis of DKD and find novel therapeutic biomarkers,
particularly miRNAs.
Materials and methods
Data resources
The expression profile GSE30122 (16) was downloaded in the Gene Expression
Omnibus (GEO, www.ncbi.nlm.nih.gov/geo) database, was used in the
current study. The dataset consisted of 35 glomeruli and 34 tubuli
tissue samples. The glomeruli samples were from 9 diabetic human
kidney patients (DKD-G group) and 26 healthy controls (G-control
group) and the tubuli samples were from 10 DKD patients (DKD-T
group) and 24 healthy controls (T-control group). The platform used
was Affymetrix Human Genome U133A 2.0 array (GPL571 HG-U133A_2;
Affymetrix; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Data pretreatment and differential
analysis
Raw data in the format of CEL was obtained and the
Affy package (version 1.48.0) (17) in R (version 3.3.1) was used to
perform background correction and quantiles normalization. After
these pretreatments, differentially-expressed genes (DEGs) between
DKD-G and G-control groups, and between DKD-T and T-control groups
were identified, respectively, using non-paired t-test in the limma
(www.bioconductor.org/packages/release/bioc/html/limma.html)
package (18). The thresholds for
significant DEG selection were log |fold-change|≥1 and P<0.05.
In addition, the pheatmap (cran.r-project.org/web/packages/pheatmap/index.html)
software was utilized to draw the clustering heatmap of these DEGs
(19). Then the overlapped DEGs in
the two comparisons were selected.
Enrichment analysis of the overlapped
DEGs
The function and pathway enrichment analyses of the
overlapped DEGs were performed using the online tool of Database
for Annotation, Visualization and Integration Discovery (DAVID;
david.abcc.ncifcrf.gov), in combination
with the gene ontology (GO; www.geneontology.org) and the Kyoto Encyclopedia of
Genes and Genomes (KEGG, http://www.genome.jp/kegg/pathway.html) databases.
Statistical significance was determined based on hypergeometric
test and the cut-off values for significant GO terms and pathway
terms were gene count (number of enriched genes in a specific
function or a pathway) was ≥2 and P<0.05.
Construction of the PPI network and
sub-network
To explore potential interactions of the overlapped
DEGs at the protein level, a PPI network was constructed based on
the Search Tool for the Retrieval of Interacting Genes (STRING;
string-db.org) database (20) and visualized using Cytoscape
(version 3.2.0, http://cytoscape.org/) (21). Parameters for the network
establishment were: The species was Homo sapiens and the PPI
score (indicating medium confidence) was ≥0.4. A node was defined
as the protein product of a DEG in the network and it was required
that all nodes in the network were DEGs.
Score of a node in the network was calculated using
one of the topological properties, degree centrality, which is
based on the concept that nodes with the highest number of
interactions were significant in the network (22). Hub nodes were those with the high
degrees in the network.
Sub-networks were extracted from the PPI network to
further elucidate the most significant functional modules of the
DEGs, using the MCODE Cytoscape plug-in (23).
Integrated miRNA-target regulatory
network construction
Potential miRNAs that may target the overlapping
DEGs were identified using the WebGestal tool (version 2.0)
(24). This was also visualized
using Cytoscape software.
Results
DEGs identified in different DKD
samples
Following preprocessing of raw data, the DEGs were
identified when the DKD-G and G-control samples and DKD-T vs.
T-control samples were compared based on the aforementioned
criteria. A total of 680 or 724 DEGs were selected in DKD-G or
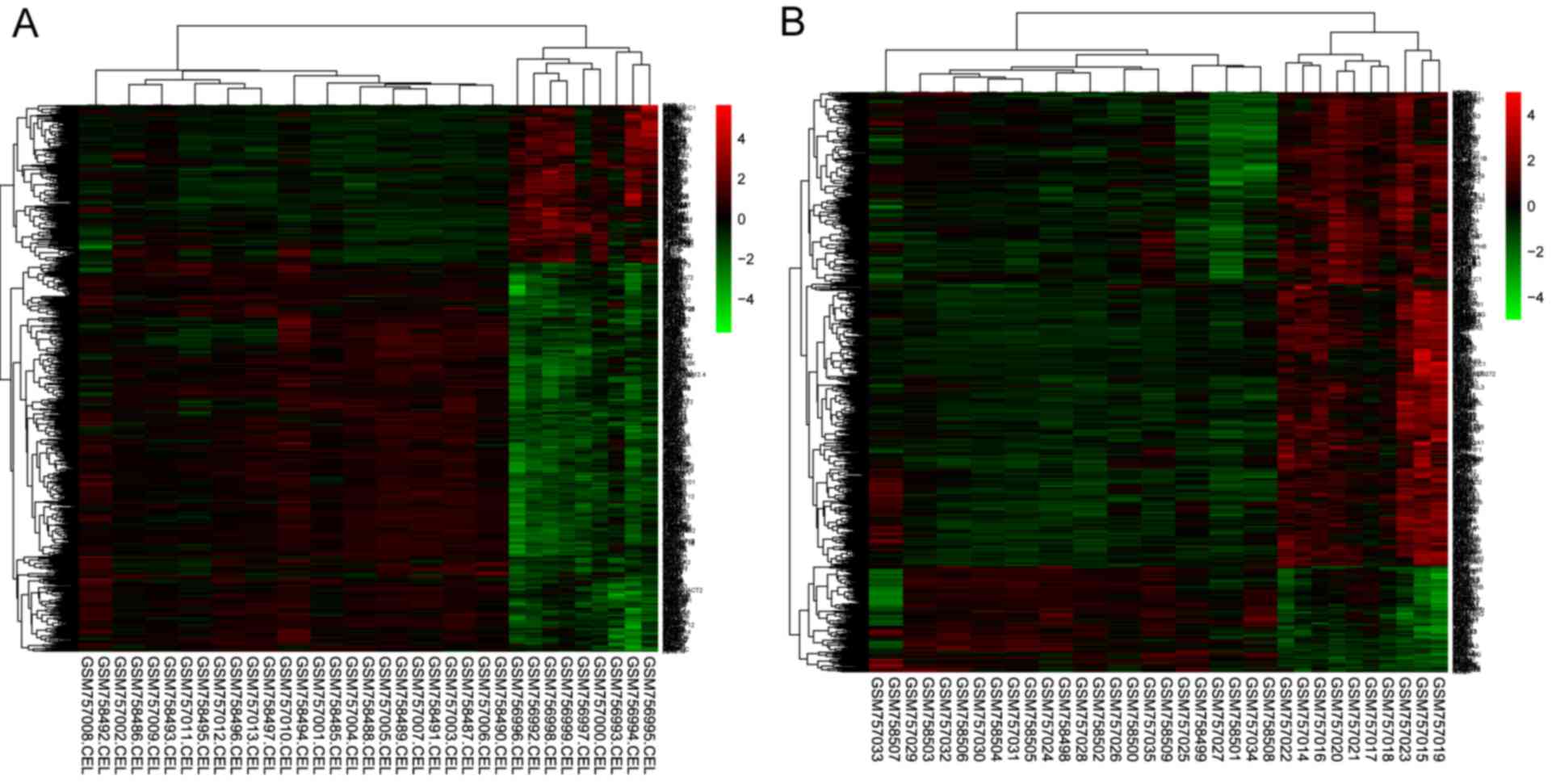
DKD-T samples, respectively. A heatmap of gene expressions were
presented in Fig. 1, which
revealed that these DEGs may distinguish the DKD-G/DKD-T samples
from the control samples.
Using overlapping analysis, a total of 139
upregulated and 28 downregulated DEGs in the two sets of DEGs were
identified.
Enrichment results of the overlapping
DEGs
The upregulated 139 DEGs were significantly enriched
in biological processes (BP), such as inflammatory/immune response,
molecular functions (MF, such as peptidase inhibitor activity and
13 pathways, including extracellular matrix (ECM)-receptor
interactions, including CD44, FN1 and collagen type
VI α3 chain (COL6A3), primary immunodeficiency, including protein
tyrosine phosphatase, receptor type, C (PTPRC), CD3d molecule and
LCK (LCK proto-oncogene, Src family tyrosine kinase) and
cytokine-cytokine receptor interaction, including C-X-C motif
chemokine receptor 4 (CXCR4), C-C motif chemokine ligand 5
(CCL5) and chemokine (C-C motif) receptor 2 (Table I), whereas the 28 downregulated
genes were primarily enriched in metabolic BP and MF, such as
carbohydrate and polysaccharide binding. No significant pathways
were enriched for the downregulated DEGs.
 | Table I.Significant pathways for the 139
overlapping upregulated differentially-expressed genes. |
Table I.
Significant pathways for the 139
overlapping upregulated differentially-expressed genes.
| ID | Name | P-value | Genes |
|---|
| hsa05322 | Systemic lupus
erythematosus | 0.000136 | C1QA, HLA-DQB1,
C1QB, C7, C3, ACTN1, HLA-DPA1, HLA-DRA |
| hsa04512 | ECM-receptor
interaction | 0.001105 | CD44, TNC, COL6A3,
COL1A2, THBS2, SPP1, FN1 |
| hsa05340 | Primary
immunodeficiency | 0.00143 | PTPRC, CD3D, LCK,
IL7R, BLNK |
| hsa04514 | Cell adhesion
molecules (CAMs) | 0.001654 | HLA-DQB1, PTPRC,
SELL, CD2, VCAN, ITGB2, HLA-DPA1, HLA-DRA |
| hsa04510 | Focal adhesion | 0.006241 | RAC2, TNC, COL6A3,
COL1A2, PDGFRA, ACTN1, THBS2, SPP1, FN1 |
| hsa05310 | Asthma | 0.006949 | FCER1A, HLA-DQB1,
HLA-DPA1, HLA-DRA |
| hsa04640 | Hematopoietic cell
lineage | 0.007496 | CD3D, CD44, CD1C,
CD2, IL7R, HLA-DRA |
| hsa05020 | Prion diseases | 0.013215 | C1QA, C1QB, C7,
CCL5 |
| hsa05416 | Viral
myocarditis | 0.015647 | HLA-DQB1, RAC2,
ITGB2, HLA-DPA1, HLA-DRA |
| hsa04610 | Complement and
coagulation cascades | 0.01646 | C1QA, C1QB, C7, C3,
CFB |
| hsa04060 | Cytokine-cytokine
receptor interaction | 0.021393 | CXCR4, IL10RA,
CCR2, PDGFRA, CCL19, CXCL6, IL7R, CCL5, LTB |
| hsa04672 | Intestinal immune
network for IgA production | 0.026364 | HLA-DQB1, CXCR4,
HLA-DPA1, HLA-DRA |
| hsa04062 | Chemokine signaling
pathway | 0.036074 | DOCK2, RAC2, CXCR4,
CCR2, CCL19, CXCL6, CCL5 |
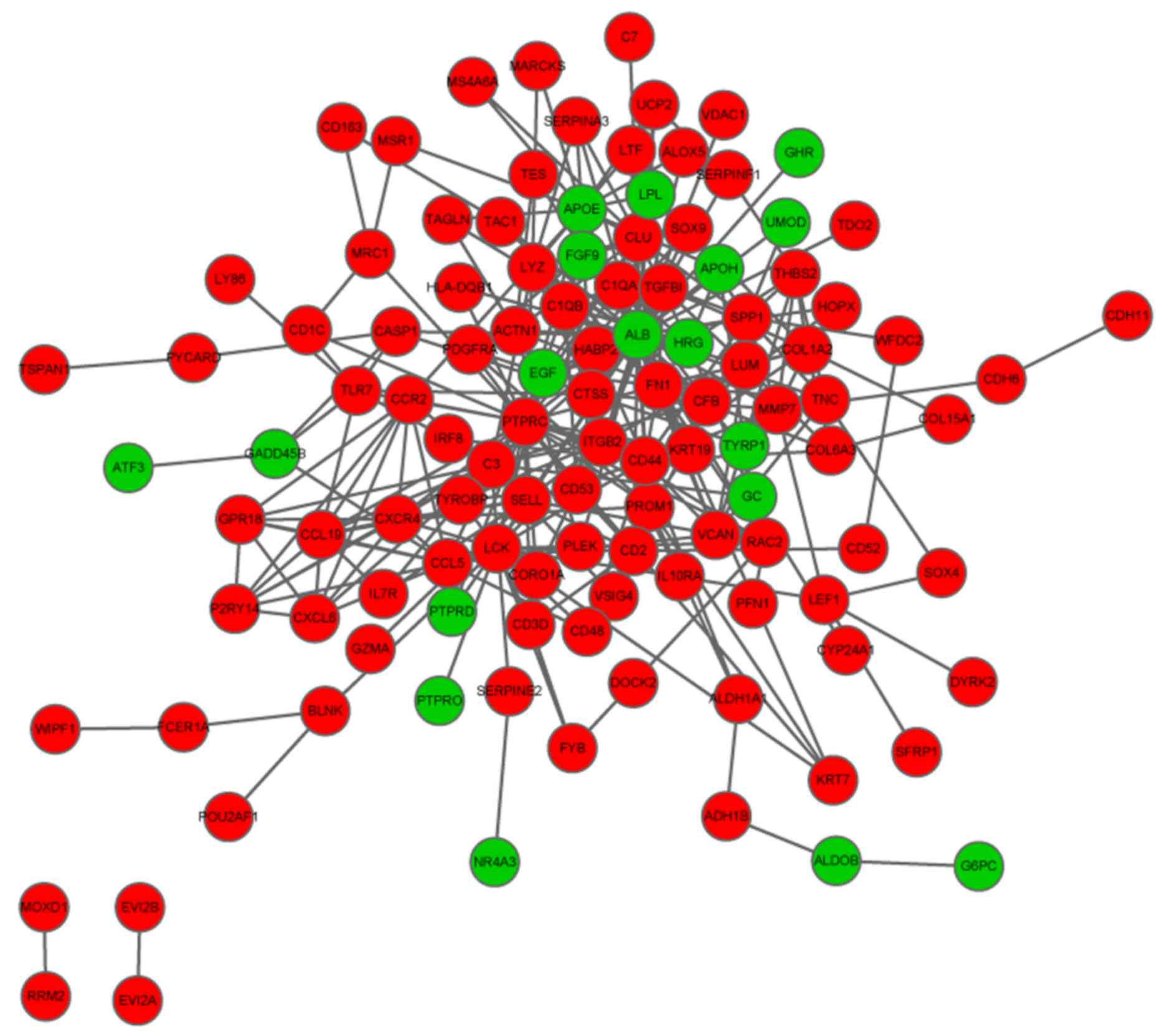
The PPI network of the overlapping
DEGs
Using the aforementioned parameters, a PPI network
was established, containing 115 nodes and 303 interactions. Top ten
nodes with high degrees were ALB (degree = 47), PTPRC (degree =
29), LCK (degree = 18), CD44 (degree = 17), C3 (degree = 16), FN1
(degree = 16), APOE (degree = 15), CLU (degree = 13), CCL5 (degree
= 11) and CXCR4 (degree = 11) (Fig.
2).
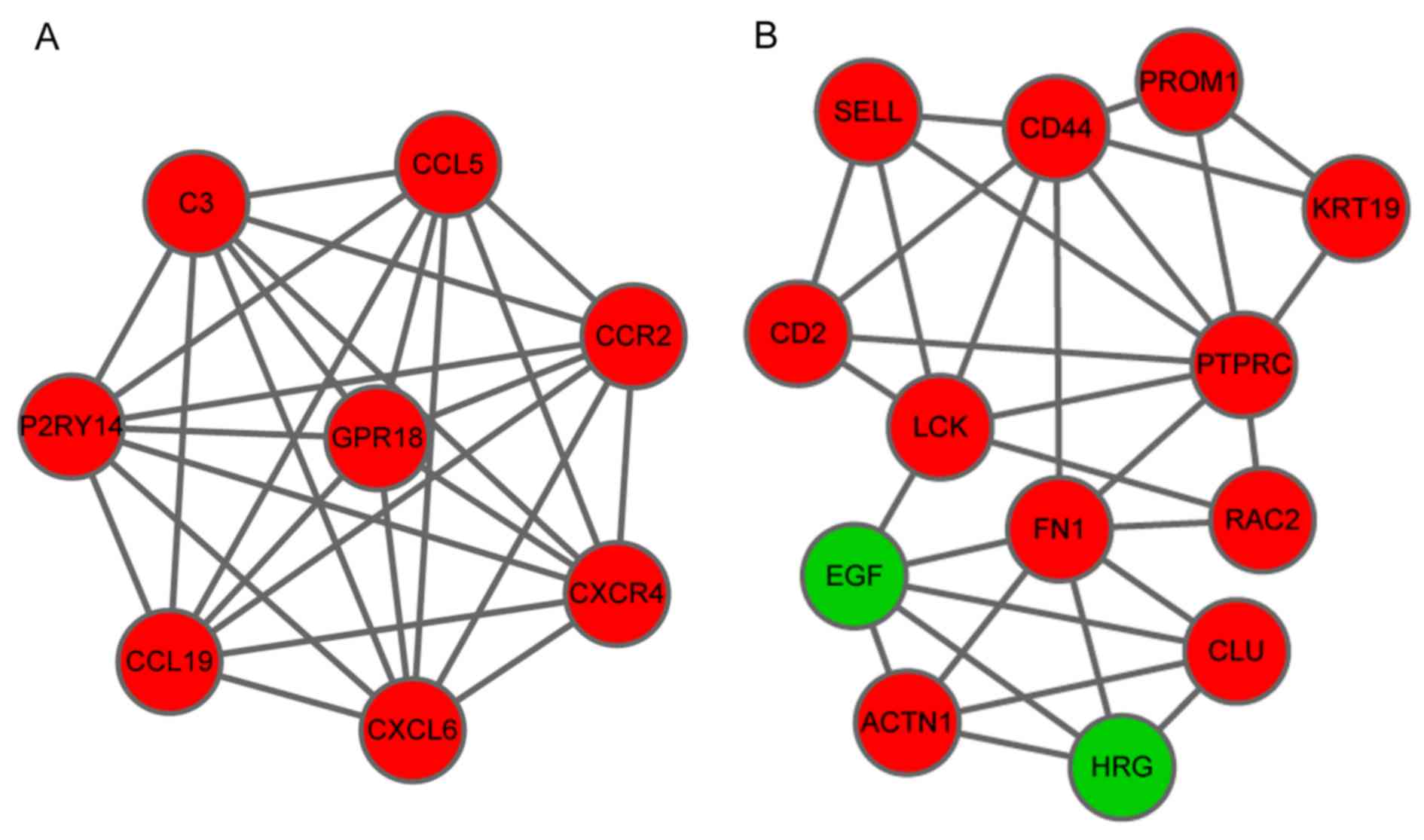
Sub-network analysis obtained two significant module
networks, A and B. Module A contained 13 nodes and 31 interactions
and Module B consisted of 8 nodes and 28 interactions. It is of
note that the majority of the of the top ten nodes identified in
the PPI network were also present in these two modules (Fig. 3).
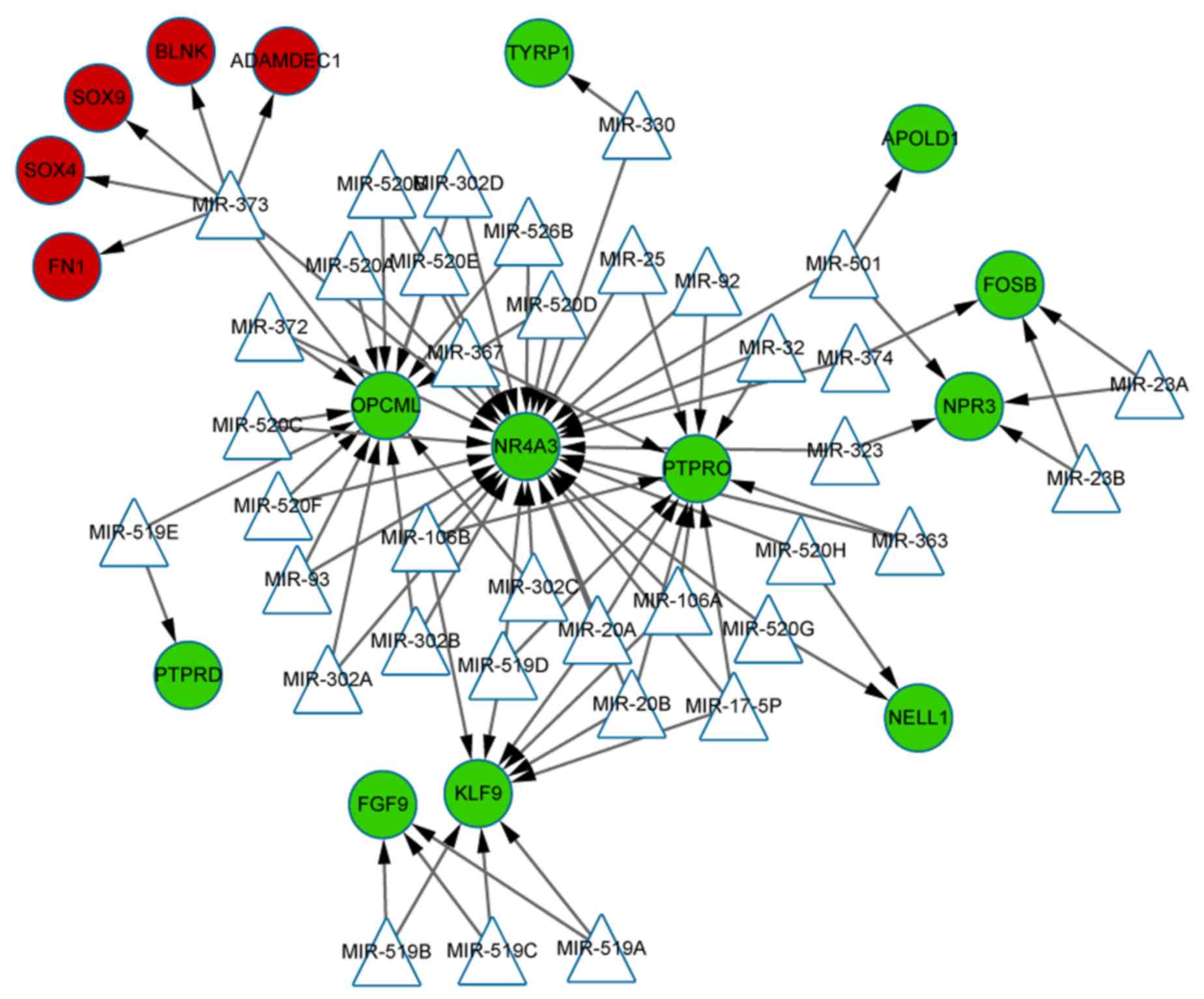
Integrated miRNA-target regulatory
network
The miRNAs that may regulate the DEGs were
identified based on the up and downregulation expressions. As
indicated in this integrated miRNA-target network (Fig. 4), only one miRNA, miR-373 was
predicted to target the upregulated DEGs, such as ADAM like decysin
1, SRY-box 4, FN1, B-cell linker and SRY-box 9. Conversely,
the network consisted of 11 downregulated DEGs, which were targeted
by 37 miRNAs. Among them, OPCML, nuclear receptor subfamily
4 group A member 3 (NR4A3), protein tyrosine phosphatase,
receptor type O (PTPRO) and Kruppel like factor 9
(KLF9) were identified as the four predominant genes with
high topological scores, which suggested that they may be the
targets of multiple miRNAs. It is of note that miR-17-5p, miR-20a
and miR-106a were all predicted to simultaneously target the genes
of NR4A3, PTPRO and KLF9.
Discussion
The present study re-analyzed the dataset of
GSE30122 and identified several important genes that were
differentially expressed in both of glomeruli and tubuli DKD
samples, including CD44, FN1, CCL5 and CXCR4. All were predominant
nodes in the PPI network and were associated with pathways like
ECM-receptor interaction and cytokine-cytokine receptor
interaction. It is of note that the present study identified
important miRNAs, including miR-17-5p, miR-20a and miR-106a and the
overlapping DEGs such as NR4A3, PTPRO and KLF9
were all predicted as target genes for the three miRNAs.
A hallmark of chronic kidney disease (CDK) is the
accumulation of ECM in glomeruli and tubular interstitium, which
may lead to glomerulosclerosis and tubulointerstitial fibrosis.
Previous studies have demonstrated that transforming growth
factor-β (TGF-β) is a key regulator in renal fibrosis (25–27)
and its action is dependent on the activation of a protein
complexes in ECM. Considering the close relationship of DKD and
CDK, it is possible that the ECM-receptor interaction pathway is a
significantly enriched pathway of the overlapped DEGs and may be
important in DKD pathogenesis. CD44, also termed CD44 Antigen or
ECMR-III, is a major cell-surface receptor for ECM and hyaluronate
(28). A previous study reported
that the protein of CD44 was downregulated in patients with kidney
transplants (29). The proximal
tubule (PTC) was identified to be important in renal interstitial
fibrosis in DN and CD44 is the receptor of hyaluronic acid in the
process of PTC hyaluronan synthesis (30). FN1 is a glycoprotein in the ECM and
has been associated with DKD metabolism (31). As a long non-coding RNA (lncRNA),
Pvt1 oncogene (PVT1) has been reported to affect the renal
metabolism and is proposed as a regulator of DN (31). ECM degradation has an active role
in promoting the glomerular fibrosis, whereas PVT1 inhibited the
degradation of ECM proteins in the glomerulus. In mesangial cells,
knockdown of this lncRNA may lead to an evident reduction of FN1
(31,32). A previous study confirmed the
deregulation of FN1 in DN, and demonstrated the reduced expression
level of this gene is regulated by the miR-1207-5p (33). These findings collectively support
the results of the current study that CD44 and FN1 were important
genes in DKD that enriched in the ECM-receptor interaction pathway
and were highlighted in the PPI network. It is possible that the
two genes may have a significant effect on DKD progression via
involvement in this ECM-associated pathway.
The biology of cytokines is often complicated; they
serve a variety of roles via interaction with expressed receptors,
to trigger signaling pathways and subsequent cytokine-driven
outcomes (34). Similar to the ECM
pathway, the cytokine-cytokine receptor interaction pathway is also
important in DN based on a network-centric analysis (35). In addition, this pathway is
significantly enriched for upregulated genes in T2D (36). A previous study determined that
mediated by Jak-STAT, this pathway may interact with TGF-β
signaling, which as aforementioned, has a role in renal fibrosis
(37). All these findings support
the significant role of this pathway in DKD. Chemokines have been
identified to promote the progression of DKD. For example, the
pro-inflammatory chemokine (C-C motif) ligand 12 contributed to the
glomerulosclerosis in mice with T2D and blocking its expression had
a protective effect on DKD (38).
The inflammatory CCL5 is expressed in various cell types, including
fibroblasts and renal tubular epithelial cells. Upregulated CCL5
was previously identified in the kidney and its expression was
directly associated with concentration of proteinuria in renal
tubular cells (1). As one of the
CXC chemokine receptors (CXCRs), CXCR4 is specific for stromal
cell-derived factor-1. The CXCR4 gene was identified to be
increased in injured kidneys (39), and was associated with the
cytokine-cytokine receptor interaction pathway in various diseases
(40,41). In the present study, CCL5 and CXCR4
were predominant in the PPI network and enriched in the
cytokine-cytokine receptor interaction pathway, supporting the
involvement of chemokines and the receptors in DKD progression.
In terms of the miRNAs, the miR-17~92 cluster is the
focus in renal disease (42). A
previous study validated that miR-17-5p and miR-20a were associated
with nephron progenitor proliferation and they are upregulated in
polycystic kidney disease (43).
Additionally, miR-20a is upregulated by TGF-β signaling and
implicated in tumorigenesis (44).
As mentioned previously, TGF-β has an important role in renal
fibrosis, and it may be inferred that the TGF-β-induced miR-20a may
also be important in DKD pathogenesis. MiR-106 has also been
previously implicated in diabetic retinopathy (45), however, the role in renal diseases
has not been identified. These findings suggested the importance of
the three miRNAs, miR-17-5p, miR-20a and miR-106, in DKD
progression.
NR4A belongs to the nuclear transcription factor
family that act as key regulators of cytokines. NR4A is suggested
to have an important role in renal inflammation (46). As one of the NR4A members,
NR4A3 was confirmed to be differentially expressed in the
renal medulla of hypertensive patients, compared with normotensive
controls (46). A previous study
confirmed that in cell transformation, NR4A3 is a target of
miR-17-5p (47). This provides a
foundation of the targeting relationship between NR4A3 and
miR-17-5p in DKD, as predicted in the integrated network of the
present study. In addition, NR4A3 may be a novel target of
miR-20a and miR-106, based on the predictions of the current
study.
PTPRO encodes for the protein tyrosine
phosphatase receptor type O, a member of the R3 subtype family.
Dysregulation of this protein leads to childhood-onset nephrotic
syndrome (48), and antibodies to
this protein lead to an increase in the glomerular albumin
permeability (49). The
Wnt/β-catenin (Ctnnb1) pathway is important to DKD and genes or
proteins in this pathway have been upregulated in glomeruli and
podocytes. It is of note that, in heterozygous
NPHS2Cre/Ctnnb1FloxE3/WT mice, expression of
PTPRO was significantly reduced (50), suggesting that this gene may
participate in the Ctnnb1 pathway. It has been previously
established that miR-17~92 cluster members have oncogenic function
in human cancers and PTPRO acts as a tumor suppressor. The
evidence provided by Xu et al (51) confirmed that PTPRO was
regulated by the miR-17~92 cluster in tumor (48). A previous review that focused on
the role of miRNA in the pathogenesis of DN indicated that
PTPRO is predicted a target of miR-25 (52). Based on our study, PTPRO may
be a novel target of miR-17-5p, miR-20a and miR-106a.
KLF9 was upregulated and identified as a
biomarker of DN by integrating three microarray datasets (53). However, its function in DKD remains
to be elucidated. A previous study reported that KLF9 was
upregulated by oxidative stress and could promote this stress
induced cell death (54). It has
been previously identified that as a target of miR-106 under
thyroid hormone treatment (55).
In the present study, KLF9 was predicted as the target gene
of miR-17-5p, miR-20a and miR-106a.
Lack of the experimental validation is a limitation
of the present study, particularly the validation of miRNA-target
regulations these will be performed in future studies. The present
study is a valuable contribution to the prediction of important
miRNAs and the reveal of pathological mechanisms in DKD.
In conclusion, the present study identified several
important genes in DKD, which may be involved in pathways, such as
ECM-receptor and cytokine-cytokine receptor interactions. Three
potential miRNAs biomarkers were identified, including miR-17-5p,
miR-20a and miR-106a, with the predicted targets of NR4A3,
PTPRO and KLF9. However, additional validation is
required to confirm these findings.
Glossary
Abbreviations
Abbreviations:
|
DKD
|
diabetic kidney disease
|
|
DN
|
diabetic nephropathy
|
|
ESKD
|
end-stage kidney disease
|
|
DKD
|
diabetic kidney disease
|
|
PPI
|
protein-protein interaction
|
|
BP
|
biological process
|
|
MF
|
molecular function
|
|
ECM
|
extracellular matrix
|
|
CDK
|
chronic kidney disease
|
|
PTC
|
proximal tubule
|
References
|
1
|
Navarro-González JF, Mora-Fernández C,
Muros de Fuentes M and García-Pérez J: Inflammatory molecules and
pathways in the pathogenesis of diabetic nephropathy. Nat Rev
Nephrol. 7:327–340. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reidy K, Kang HM, Hostetter T and Susztak
K: Molecular mechanisms of diabetic kidney disease. J Clin Invest.
124:2333–2340. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nasri H: On the occasion of the world
diabetes day 2013; diabetes education and prevention; a nephrology
point of view. J Renal Inj Prev. 2:31–32. 2013.PubMed/NCBI
|
|
4
|
Xiao X, Ma B, Dong B, Zhao P, Tai N, Chen
L, Wong FS and Wen L: Cellular and humoral immune responses in the
early stages of diabetic nephropathy in NOD mice. J Autoimmun.
32:85–93. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tang W, Gao Y, Li Y and Shi G: Gene
networks implicated in diabetic kidney disease. Eur Rev Med
Pharmacol Sci. 16:1967–1973. 2012.PubMed/NCBI
|
|
6
|
Harjutsalo V and Groop PH: Epidemiology
and risk factors for diabetic kidney disease. Adv Chronic Kidney
Dis. 21:260–266. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Palmer ND, Ng MC, Hicks PJ, Mudgal P,
Langefeld CD, Freedman BI and Bowden DW: Evaluation of candidate
nephropathy susceptibility genes in a genome-wide association study
of African American diabetic kidney disease. PLoS One.
9:e882732014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Himmelfarb J and Tuttle KR: New therapies
for diabetic kidney disease. N Engl J Med. 369:2549–2550. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Anderberg RJ, Meek RL, Hudkins KL, Cooney
SK, Alpers CE, Leboeuf RC and Tuttle KR: Serum amyloid a and
inflammation in diabetic kidney disease and podocytes. Lab Invest.
95:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Perez-Gomez MV, Sanchez-Niño MD, Sanz AB,
Zheng B, Martín-Cleary C, Ruiz-Ortega M, Ortiz A and
Fernandez-Fernandez B: Targeting inflammation in diabetic kidney
disease: Early clinical trials. Expert Opin Investig Drugs.
25:1045–1058. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nasri H and Rafieiankopaei M: Protective
effects of herbal antioxidants on diabetic kidney disease. J Res
Med Sci. 19:82–83. 2014.PubMed/NCBI
|
|
12
|
Giuliani F, Di Maio M, Colucci G and
Perrone F: Conventional chemotherapy of advanced pancreatic cancer.
Curr Drug Targets. 13:795–801. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brosius FC, Tuttle KR and Kretzler M: JAK
inhibition in the treatment of diabetic kidney disease.
Diabetologia. 59:1624–1627. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kato M, Zhang J, Wang M, Lanting L, Yuan
H, Rossi JJ and Natarajan R: MicroRNA-192 in diabetic kidney
glomeruli and its function in TGF-beta-induced collagen expression
via inhibition of E-box repressors. Proc Natl Acad Sci U S A.
104:pp. 3432–3437. 2007; View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chung ACK: MicroRNAs in diabetic kidney
disease. Adv Exp Med Biol. 888:253–269. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Woroniecka KI, Park AS, Mohtat D, Thomas
DB, Pullman JM and Susztak K: Transcriptome analysis of human
diabetic kidney disease. Diabetes. 60:2354–2369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kauffmann A, Gentleman R and Huber W:
arrayQualityMetrics - a bioconductor package for quality assessment
of microarray data. Bioinformatics. 25:415–416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Smyth GK: Limma: Linear Models for
Microarray Data. Springer; New York, NY: 2005
|
|
19
|
Kolde R: Pheatmap: Pretty Heatmaps.
2015.
|
|
20
|
Szklarczyk D, Franceschini A, Kuhn M,
Simonovic M, Roth A, Minguez P, Doerks T, Stark M, Muller J, Bork
P, et al: The STRING database in 2011: Functional interaction
networks of proteins, globally integrated and scored. Nucleic Acids
Res. 39:D561–D568. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Smoot ME, Ono K, Ruscheinski J, Wang PL
and Ideker T: Cytoscape 2.8: New features for data integration and
network visualization. Bioinformatics. 27:431–432. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Latora V and Marchiori M: A measure of
centrality based on network efficiency. New J Phys. 9:2007.
View Article : Google Scholar
|
|
23
|
Dagan T, Artzy-Randrup Y and Martin W:
Modular networks and cumulative impact of lateral transfer in
prokaryote genome evolution. Proc Natl Acad Sci USA. 105:pp.
10039–10044. 2008; View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang J, Duncan D, Shi Z and Zhang B:
WEB-based GEne SeT AnaLysis Toolkit (WebGestalt): Update 2013.
Nucleic Acids Res. 41:W77–W83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
López-Hernandez FJ and López-Novoa JM:
Role of TGF-β in chronic kidney disease: An integration of tubular,
glomerular and vascular effects. Cell Tissue Res. 347:141–154.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao J, Wang L, Cao AL, Jiang MQ, Chen X,
Wang Y, Wang YM, Wang H, Zhang XM and Peng W: HuangQi decoction
ameliorates renal fibrosis via tgf-βsmad signaling pathway in vivo
and in vitro. Cell Physiol Biochem. 38:1761–1774. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pezzolesi MG, Skupien J, Mychaleckyj JC,
Warram JH and Krolewski AS: Insights to the genetics of diabetic
nephropathy through a genome-wide association study of the GoKinD
collection. Semin Nephrol. 30:126–140. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Alldinger S, Gröters S, Miao Q, Fonfara S,
Kremmer E and Baumgörtner W: Roles of an extracellular matrix (ECM)
receptor and ECM processing enzymes in demyelinating canine
distemper encephalitis. Dtsch Tierarztl Wochenschr. 113:151–152.
2006.PubMed/NCBI
|
|
29
|
Mullen W, Delles C and Mischak H: EuroKUP
COST action: Urinary proteomics in the assessment of chronic kidney
disease. Curr Opin Nephrol Hypertens. 20:654–661. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Phillips AO and Steadman R: Diabetic
nephropathy: The central role of renal proximal tubular cells in
tubulointerstitial injury. Histol Histopathol. 17:247–252.
2002.PubMed/NCBI
|
|
31
|
Alvarez ML and Distefano JK: Functional
characterization of the plasmacytoma variant translocation 1 gene
(PVT1) in diabetic nephropathy. PLoS One. 6:e186712011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alvarez ML and Distefano JK: The role of
non-coding RNAs in diabetic nephropathy: Potential applications as
biomarkers for disease development and progression. Diabetes Res
Clin Pract. 99:1–11. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alvarez ML, Khosroheidari M, Eddy E and
Kiefer J: Role of microRNA 1207-5P and its host gene, the long
non-coding RNA Pvt1, as mediators of extracellular matrix
accumulation in the kidney: Implications for diabetic nephropathy.
PLoS One. 8:e774682013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sedger LM, Seddiki N and Ranasinghe C:
Cytokines and cytokine receptors as immunotherapeutics: Humble
beginnings and exciting futures. Cytokine Growth Factor Rev.
25:351–353. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ntemka A, Iliadis F, Papanikolaou N and
Grekas D: Network-centric analysis of genetic predisposition in
diabetic nephropathy. Hippokratia. 15:232–237. 2011.PubMed/NCBI
|
|
36
|
Skov V, Knudsen S, Olesen M, Hansen ML and
Rasmussen LM: Global gene expression profiling displays a network
of dysregulated genes in non-atherosclerotic arterial tissue from
patients with type 2 diabetes. Cardiovasc Diabetol. 11:1475–2840.
2012. View Article : Google Scholar
|
|
37
|
Heinzel A, Perco P, Mayer G, Oberbauer R,
Lukas A and Mayer B: From molecular signatures to predictive
biomarkers: Modeling disease pathophysiology and drug mechanism of
action. Front Cell Dev Biol. 2:372014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Darisipudi MN, Kulkarni OP, Sayyed SG, Ryu
M, Migliorini A, Sagrinati C, Parente E, Vater A, Eulberg D,
Klussmann S, et al: Dual blockade of the homeostatic chemokine
CXCL12 and the proinflammatory chemokine CCL2 has additive
protective effects on diabetic kidney disease. Am J Pathol.
179:116–124. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Long DA, Norman JT and Fine LG: Restoring
the renal microvasculature to treat chronic kidney disease. Nat Rev
Nephrol. 8:244–250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu J, Long Z, Cai H, Du C, Liu X, Yu S and
Wang Y: High expression of WISP1 in colon cancer is associated with
apoptosis, invasion and poor prognosis. Oncotarget. 7:49834–49847.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Valsecchi R, Coltella N, Belloni D,
Ponente M, Ten Hacken E, Scielzo C, Scarfò L, Bertilaccio MT,
Brambilla P, Lenti E, et al: HIF-1α regulates the interaction of
chronic lymphocytic leukemia cells with the tumor microenvironment.
Blood. 127:1987–1997. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Patel V, Williams D, Hajarnis S, Hunter R,
Pontoqlio M, Somlo S and Iqarashi P: miR-17~92 miRNA cluster
promotes kidney cyst growth in polycystic kidney disease. Proc Natl
Acad Sci USA. 110:pp. 10765–10770. 2013; View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Phua YL and Ho J: MicroRNAs in the
pathogenesis of cystic kidney disease. Curr Opin Pediatr.
27:219–226. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Butz H, Rácz K, Hunyady L and Patócs A:
Crosstalk between TGF-β signaling and the microRNA machinery.
Trends Pharmacol Sci. 33:382–393. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Moura J, Børsheim E and Carvalho E: The
role of MicroRNAs in diabetic complications-special emphasis on
wound healing. Genes (Basel). 5:926–956. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Marques FZ, Campain AE, Tomaszewski M,
Zukowska-Szczechowska E, Yang YH, Charchar FJ and Morris BJ: Gene
expression profiling reveals renin mRNA overexpression in human
hypertensive kidneys and a role for microRNAs. Hypertension.
58:1093–1098. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cloonan N, Brown MK, Steptoe AL, Wani S,
Chan WL, Forrest AR, Kolle G, Gabrielli B and Grimmond SM: The
miR-17-5p microRNA is a key regulator of the G1/S phase cell cycle
transition. Genome Biol. 9:R1272008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ozaltin F, Ibsirlioglu T, Taskiran EZ,
Baydar DE, Kaymaz F, Buyukcelik M, Kilic BD, Balat A, Iatropoulos
P, Asan E, et al: Disruption of PTPRO causes childhood-onset
nephrotic syndrome. Am J Hum Genet. 89:139–147. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Charba DS, Wiggins RC, Goyal M, Wharram
BL, Wiggins JE, McCarthy ET, Sharma R, Sharma M and Savin VJ:
Antibodies to protein tyrosine phosphatase receptor type O (PTPro)
increase glomerular albumin permeability (P(alb)). Ajp Renal
Physiology. 297:F138–F144. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kato H, Gruenwald A, Suh JH, Miner JH,
Barisoni-Thomas L, Taketo MM, Faul C, Millar SE, Holzman LB and
Susztak K: Wnt/β-catenin pathway in podocytes integrates cell
adhesion, differentiation and survival. J Biol Chem.
286:26003–26015. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xu X, Hong Y, Kong C, Xu L, Tan J, Liang
Q, Huang B and Lu J: Protein tyrosine phosphatase receptor-type O
(PTPRO) is co-regulated by E2F1 and miR-17-92. FEBS Lett.
582:2850–2856. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Conserva F, Pontrelli P, Accetturo M and
Gesualdo L: The pathogenesis of diabetic nephropathy: Focus on
microRNAs and proteomics. J Nephrol. 26:811–820. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jiang ZS, Jia HX, Xing WJ, Han CD, Wang J,
Zhang ZJ and Qu W: Investigation of several biomarkers associated
with diabetic nephropathy. Exp Clin Endocrinol Diabetes. 123:1–6.
2015.PubMed/NCBI
|
|
54
|
Zucker SN, Fink EE, Bagati A, Mannava S,
Bianchi-Smiraglia A, Bogner PN, Wawrzyniak JA, Foley C, Leonova KI,
Grimm MJ, et al: Nrf2 amplifies oxidative stress via induction of
Klf9. Mol Cell. 53:916–928. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Dong H, You SH, Williams A, Wade MG, Yauk
CL and Thomas Zoeller R: Transient maternal hypothyroxinemia
potentiates the transcriptional response to exogenous thyroid
hormone in the fetal cerebral cortex before the onset of fetal
thyroid function: A messenger and microrna profiling study. Cereb
Cortex. 25:1735–1745. 2015. View Article : Google Scholar : PubMed/NCBI
|