Introduction
Bronchial asthma is a chronic inflammatory
respiratory disease that seriously affects human health (1). It develops from a chronic airway
inflammation that involves eosinophils, lymphocytes, mast cells,
neutrophils and other inflammatory cells and cell components
(2–4). Approximately 300 million people
worldwide currently suffer from asthma. The World Health
Organization sponsored the World Asthma Day (the first Tuesday in
May each year) to remind the public awareness of the disease and
strengthen its prevention and treatment. At present, many
asthma-related issues remain poorly understood. Therefore, it
remains very important to investigate the pathogenesis of asthma
and to look for new preventive and therapeutic targets in
asthma.
High mobility group box 1 protein (HMGB1) is a
highly conserved nuclear protein that can be used as an
immunomodulatory factor and an inflammatory factor to participate
in airway inflammation (5).
Intranuclear HMGB1 mainly regulates DNA recombination, replication,
repair and transcription (6).
Extranuclear HMGB1 can be actively released by immune cells
including monocytes/macrophages under the stimulation of
inflammatory factors such as lipopolysaccharide, tumor necrosis
factor (TNF)-α, and interleukin (IL)-1 and it can be passively
released by necrotic cells in the early stage of tissue injury and
necrosis (7–9). Extranuclear HMGB1, as an important
endogenous proinflammatory factor and an inflammatory mediator,
participates in the pathological process of various diseases such
as sepsis, pneumonia, and arthritis (10–12).
Toll-like receptor-4 (TLR4) is an important immune
pattern recognition receptor that controls innate and adaptive
immune responses and plays an important role in initiating and
regulating airway inflammation (13,14).
TLR4 is the main receptor of HMGB1 (15). Nuclear factor (NF)-κB is located on
the downstream TLR4 signaling pathway and is also in a pivotal
position to regulate the immune response, cell proliferation and
differentiation (16). Therefore,
HMGB1/TLR4/NF-κB signaling pathway is an important part of
immunoregulatory processes. It is also considered an important
pathological mechanism underlying asthma. Regulation and
intervention of any part of the signaling pathway may affect the
occurrence and development of asthma.
Vitamin D (VD) is reportedly to be involved in many
aspects of allergic and autoimmune diseases, exhibiting a
remarkable immunoregulatory effect (17,18).
There is evidence that VD receptor (VDR) gene is a new
susceptibility gene for asthma (19). VD exhibits biological effects via
binding to intracellular VDR, and its immunoregulatory effects on
asthma has become an increasing area of interest. A previous study
has demonstrated that 1,25-(OH)2D3 can
downregulate the expression of MHC class II molecules and
co-stimulatory molecules on antigen-presenting cell surface,
prevent antigen presentation and T cell immune response, inhibit
the expression of IL-4, interferon-γ, and IL-5, and thereby
alleviate airway inflammation (20). In this study, we investigated the
role of VD in a mouse model of asthma, and the mechanism of action
of HMGB1/TLR4/NF-κB in asthma, providing a theoretical basis for
clinical application of targeted drug therapy and development of
new drugs against asthma.
Materials and methods
Animals and grouping
Healthy Balb/c mice, weighing 20–22 g, were purposed
by Laboratory Animal Center, China Medical University, China. The
experiments were approved by China Medical University Institutional
Animal Care and Use Committee (IACUC; no. 20167642). Mice were
randomly divided into six groups with 8 mice in each group: Normal
control, asthma, asthma + high-dose VD (HVD), asthma + medium-dose
VD (MVD), asthma + low-dose VD (LVD), asthma + VD + 40 µg/kg/d
E5564 (EVD). Mice in the LVD group were daily administered a
mixture of 0.1 µg/ml/20 g VD solution via the tail vein. Mice in
the MVD group were identically administered a mixture of 0.4
µg/ml/20 g VD solution per day. Mice in the HVD group were
identically injected with a mixture of a mixture of 1 µg/ml/20 g VD
solution per day. Mice in the normal control group were daily
injected with equal amounts of normal saline via the tail vein.
Establishment of mouse models of
asthma
Mice in all groups with the exception of normal
control group were intraperitoneally administered 0.2 ml antigen
solution (50 µg ovalbumin + 0.15 ml 10% Al (OH)3 + 0.05
ml normal saline). Mice were immunized on experimental days 1, 8
and 15. On day 21, mice were placed in a closed container and
sprayed with 5% ovalbumin and normal saline, once a day, 45 min
each time, for 7 successive days. In the normal control group, only
equal amount of normal saline was used to spray the mice. Mice in
the HVD, MVD, and LVD groups were injected with VD solution via the
tail vein 30 min before irritation.
Sample collection
On the day of asthma model establishment and on day
28 of intervention, blood samples were collected from mouse vein.
Serum was separated and ELISA assay was performed. Mouse bronchial
alveolar tissue was isolated. One portion of bronchial alveolar
tissue was fixed in 10% formaldehyde and the other portion was used
for western blot assay and reverse transcription-quantitative
polymerase chain reaction (RT-qPCR; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Measurement of airway resistance in
asthmatic mice
The airway resistance in asthmatic mice was measured
at 24 h after the last spraying. Briefly, after anesthesia with
sodium pentobarbital, a 22G indwelling needle was positioned for
tracheal intubation and it was connected with a pulmonary function
testing instrument. The mice were placed in a closed incubator to
maintain body temperature at 37°C and assisted mechanical
ventilation was given. Airway resistance (cm H2O/L/s)
was measured. The airway resistance for mice inhaling PBS was
designated as R (baseline) and the airway resistance for mice
inhaling different concentrations of methacholine was designed as R
(response) at the corresponding concentrations. The highest value
of R at each concentration of methacholine was introduced into the
following formula. The fold increase of R was used as an evaluation
index of airway resistance: Fold increase of R=[R (response)-R
(baseline)]/R (baseline).
Hematoxylin and eosin staining
Mouse bronchial alveolar tissue was dehydrated,
cleared, embedded with paraffin, sliced, stained with hematoxylin
for 5 min, washed with PBS, differentiated with hydrochloric
acid-ethanol for 3 sec, stained with eosin for 2 min, and mounted
with neutral resin. Finally, pathological change of mouse bronchial
alveolar tissue was observed under the optical microscope.
ELISA assay
Serum levels of IL-1β (SEA563Mu, CCC), IL-6
(SEA079Mu, CCC), TNF-α (SEA134Mu, CCC) and IL-10 (SEA056Mu, CCC)
were measured by ELISA assay according to kit instructions. A total
of 100 µl standard sample and 100 µl diluted sample were added to
the reaction plate and incubated at 37°C for 30 min. After washes,
100 µl tested sample was added to each well and then incubated at
37°C for 2 h. After washes, 100 µl horseradish peroxidase-labeled
secondary antibody was added to each well. Samples were incubated
at 37°C for 30 min. After washes, developer A and developer B, each
50 µl, was added. The tested sample was developed for 15 min in the
dark. A stop solution was added at 50 µl per well to terminate the
reaction. Optical density at 450 nm was read using an ELISA reader
(EXL808; BioTek Instruments, Inc., Winooski, VT, USA). A standard
curve was drawn. According to the curve equation, the concentration
of the corresponding sample was calculated.
RT-qPCT
Mouse lung tissue was thoroughly ground into a
powder and total RNA was extracted using 1 ml TRIzol (15596018;
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the reagent operating instructions. First chain DNA
was reverse transcribed. RT-qPCR was performed according to kit
instructions (204057; Qiagen GmbH, Hilden, Germany). The relative
gene expression data was analyzed with the 2−ΔΔCq
method. The primers used for RT-qPCR are listed in Table I.
 | Table I.RT-qPCR using gene primers. |
Table I.
RT-qPCR using gene primers.
| Gene | Primer (5′→3′) |
|---|
| Bax | Forward:
GCGGCGACATGGAGACAG |
|
| Reverse:
GTGTGACCCGAACCAGAAG |
| Bcl2 | Forward:
GCAGGGTGGTGGCACTGT |
|
| Reverse:
ACAATTATCAGCTGGACC |
| Caspase-3 | Forward:
CACATGGCAGACGGTGGC |
|
| Reverse:
CTGGAGTTCTCACCACTG |
| GAPDH | Forward:
AACATCGATCTCGAGGTC |
|
| Reverse:
TTCAACTGCCGCAGGGTT |
Western blot analysis
Mouse bronchial alveolar tissue was lysed using RIPA
lysate containing protease inhibitor on ice and centrifuged. The
supernatant was collected. Protein concentration was determined
using a BCA protein assay kit. Protein samples were subjected to
SDS-PAGE and then transferred onto PVDF membrane. Bax (ab32503;
Abcam, Cambridge, UK), Bcl2 (ab59348; Abcam), caspase-3 (ab13847;
Abcam), Active-caspase3 (ab47822; Abcam) HMGB1 (ab18256; Abcam),
TLR4 (ab13867; Abcam), NF-κB (ab16502; Abcam) and p-NF-κB p65
(ab86299; Abcam) antibodies were added. Protein samples were
incubated overnight at 4°C. PVDF membrane was washed with TBST.
After horseradish peroxidase labeled secondary antibody was added,
protein samples were incubated at room temperature for 2 h. Protein
bands were visualized using an ECL chemiluminescence (Bio-Rad
Laboratories, Inc.) detection kit and a gel imaging system.
Absorbance analysis was performed using Image Tools.
Statistical analysis
All data were statistically analyzed using SPSS 19.0
software and are expressed as the mean ± standard deviation. t-test
was used for pairwise comparison. One-way analysis of variance was
performed for comparisons among groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
VD improves asthma symptoms in
asthmatic mice
Compared with normal control mice, asthmatic mice
had reduced activities, were irritable, exhibited running nose,
sneezing and lackluster hair as well as deep and fast breathing,
nodding while breathing, slow action, and cyanosis in lips after
spraying. LVD group mice showed obvious runny nose and other asthma
symptoms. MVD group the asthma symptoms has improved compared with
LVD group. After treatment with HVD, the abovementioned symptoms
alleviated, and mice breathed better and slow action was improved.
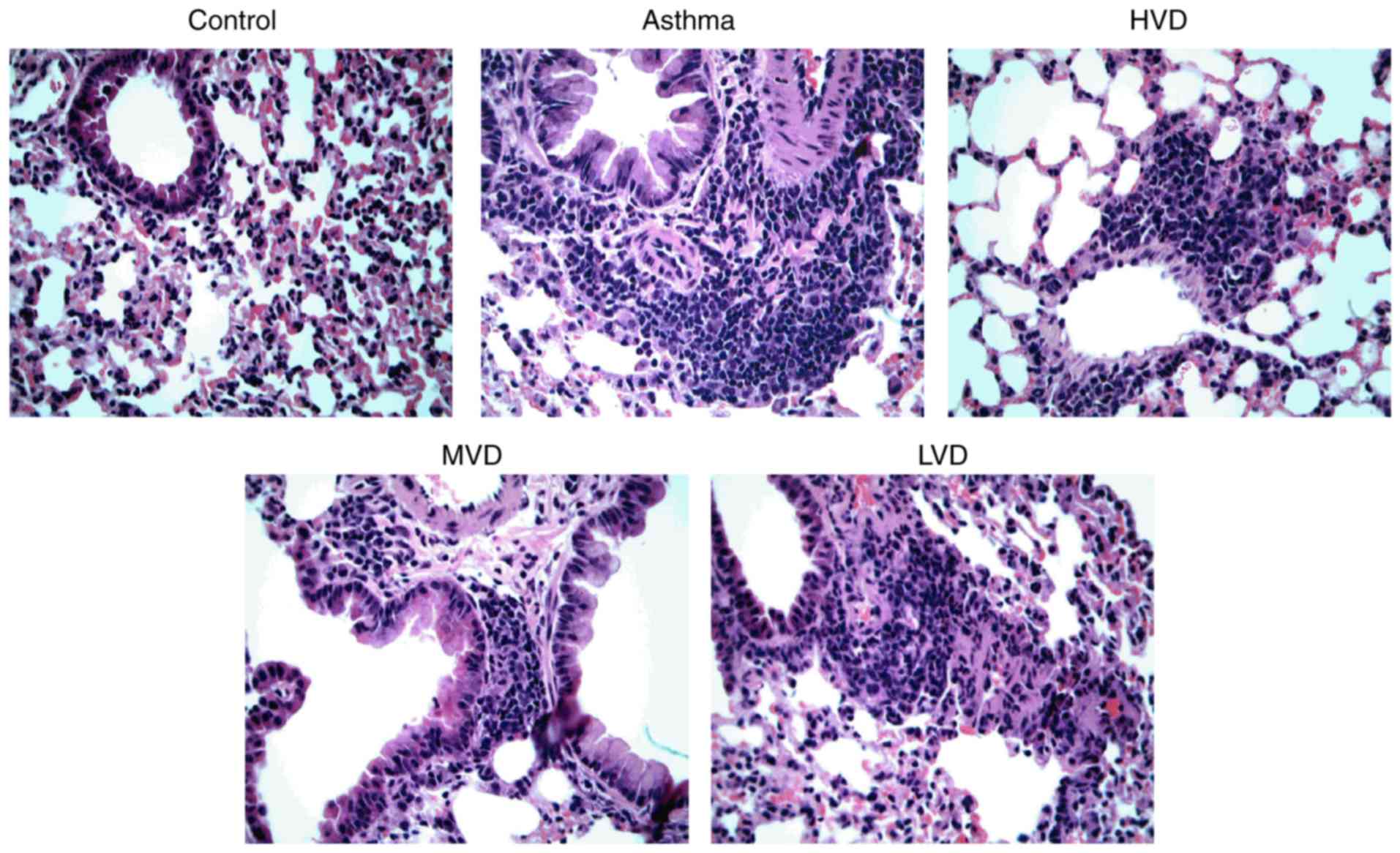
Hematoxylin and eosin staining revealed that airway injury was
obvious in asthmatic mice, a large number of inflammatory cells
were infiltrated, and epithelial cells were poorly arranged
(Fig. 1). After treatment with
HVD, the number of infiltrated inflammatory cells was decreased.
MVD and LVD group mice still have a lot of inflammatory cell
infiltration. HVD exhibited better obvious therapeutic effects than
MVD and LVD.
VD attenuates airway resistance in
asthmatic mice
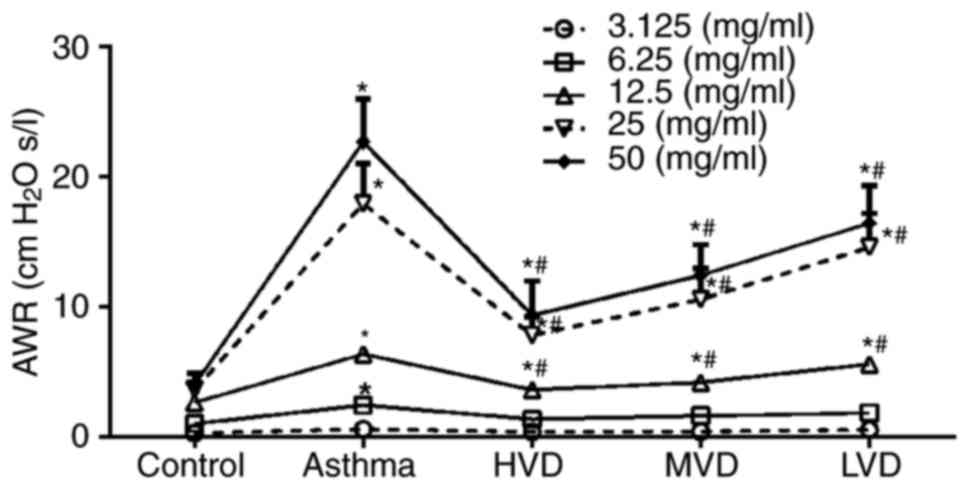
After spayed with different concentrations of
methacholine, the airway resistance of asthmatic mice was obviously
increased, but after treatment with HVD, airway resistance of
asthmatic mice was significantly decreased (P<0.05, vs. asthma
group) (Fig. 2). Airway resistance
of asthmatic in LVD group had no significantly decreased, and in
MVD group airway resistance of asthmatic mice was decreased, but it
was no significantly improved compared with HVD. All these results
suggest that VD can obviously attenuate the airway resistance of
asthmatic mice.
VD reduces inflammatory response in
asthmatic mice
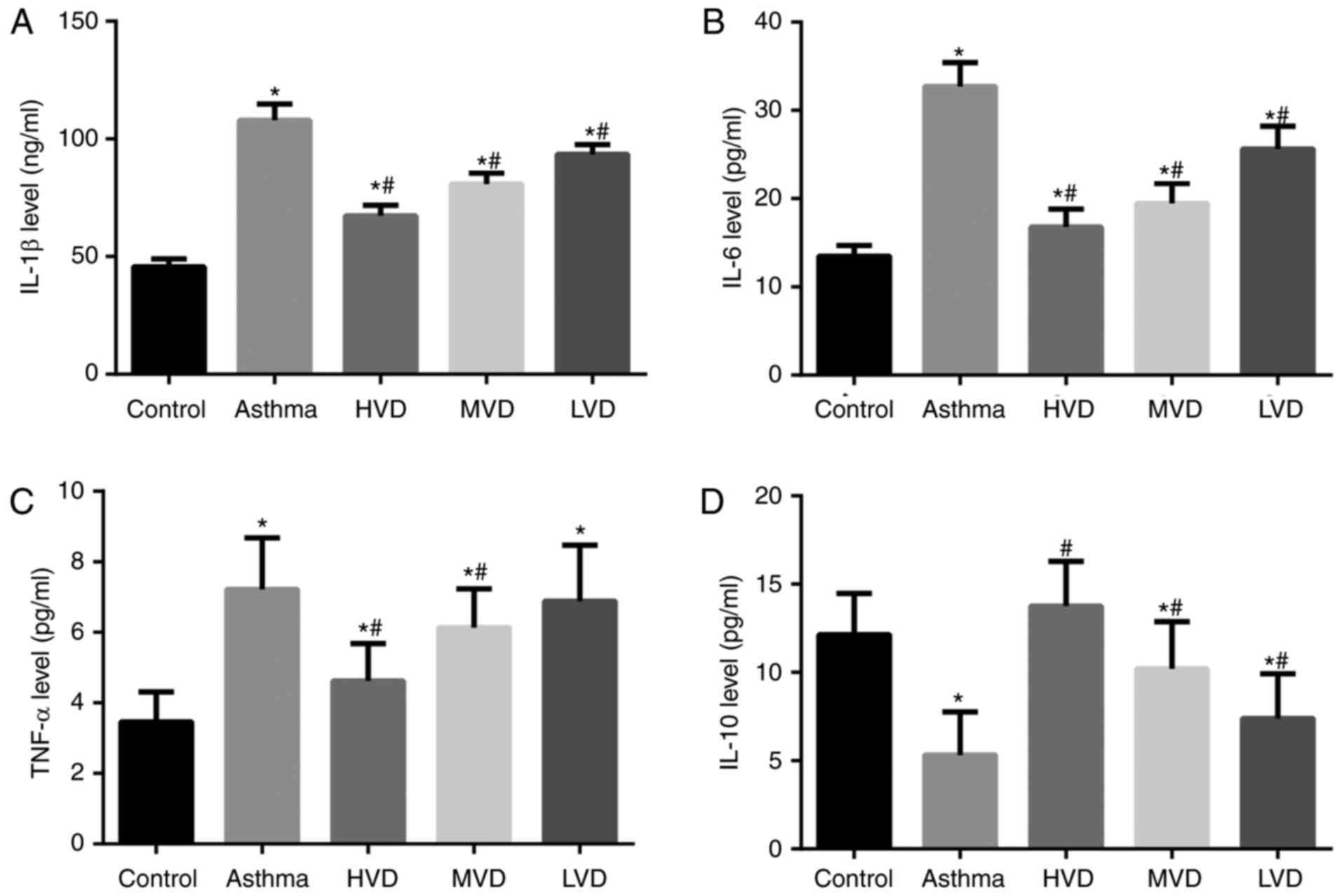
ELISA results showed that serum levels of IL-1β,
IL-6, TNF-α were significantly increased (Fig. 3A-C) and IL-10 was significantly
decreased in the asthma group than in the normal control group
(P<0.05) (Fig. 3D). After VD
treatment, serum levels of IL-1β, IL-6, TNF-α were significantly
decreased and IL-10 was significantly increased in the HVD group
than in the asthma group (P<0.05). These results suggest that
HVD can effectively reduce the inflammatory response in asthmatic
mice.
VD reduces cell apoptosis in the lung
tissue of asthmatic mice
Western blot analysis revealed that Bax, caspase-3
and active-caspase3 expression in the lung tissue was significantly
increased, while Bcl-2 expression was significantly decreased, in
the asthma group than those in the normal control group (P<0.05;
Fig. 4A). Bax and caspase-3
expression in the lung tissue was significantly, while Bcl-2
expression was significantly increased in the HVD, MVD and LVD
groups than those in the asthma group (P<0.05; Fig. 4B). These data suggest that VD can
reduce cell apoptosis in the lung tissue of asthmatic mice.
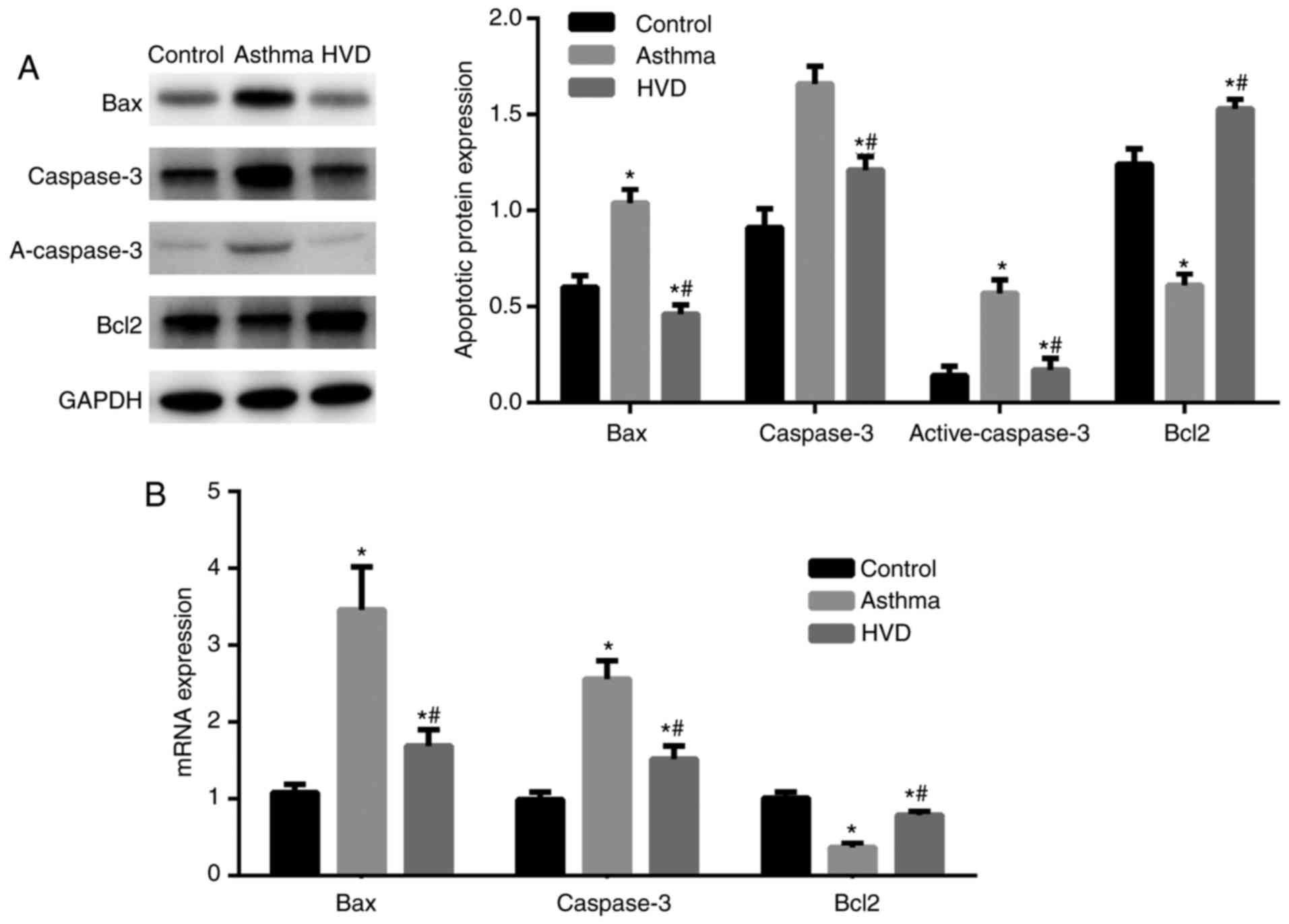 | Figure 4.After high, medium and low dose of
vitamin D (VD) were treated, the high dose of VD was choose to the
treatment dose. Apoptosis factors Bcl-2-associated X (Bax),
caspase-3, B-cell lymphoma 2 (Bcl2) expression in mice lung tissue.
Data collected from control group, asthma group, HVD group.
Compared with control group, *P<0.05; Compared with asthma
group, #P<0.05. (A) Bax, caspase-3, Bcl2 expression by western
blotting. (B) Bax, caspase-3, Bcl2 mRNA expression by RT-qPCR. |
VD reduces inflammatory response in
asthmatic mice through HMGB1/TLR4/NF-κB signaling pathway
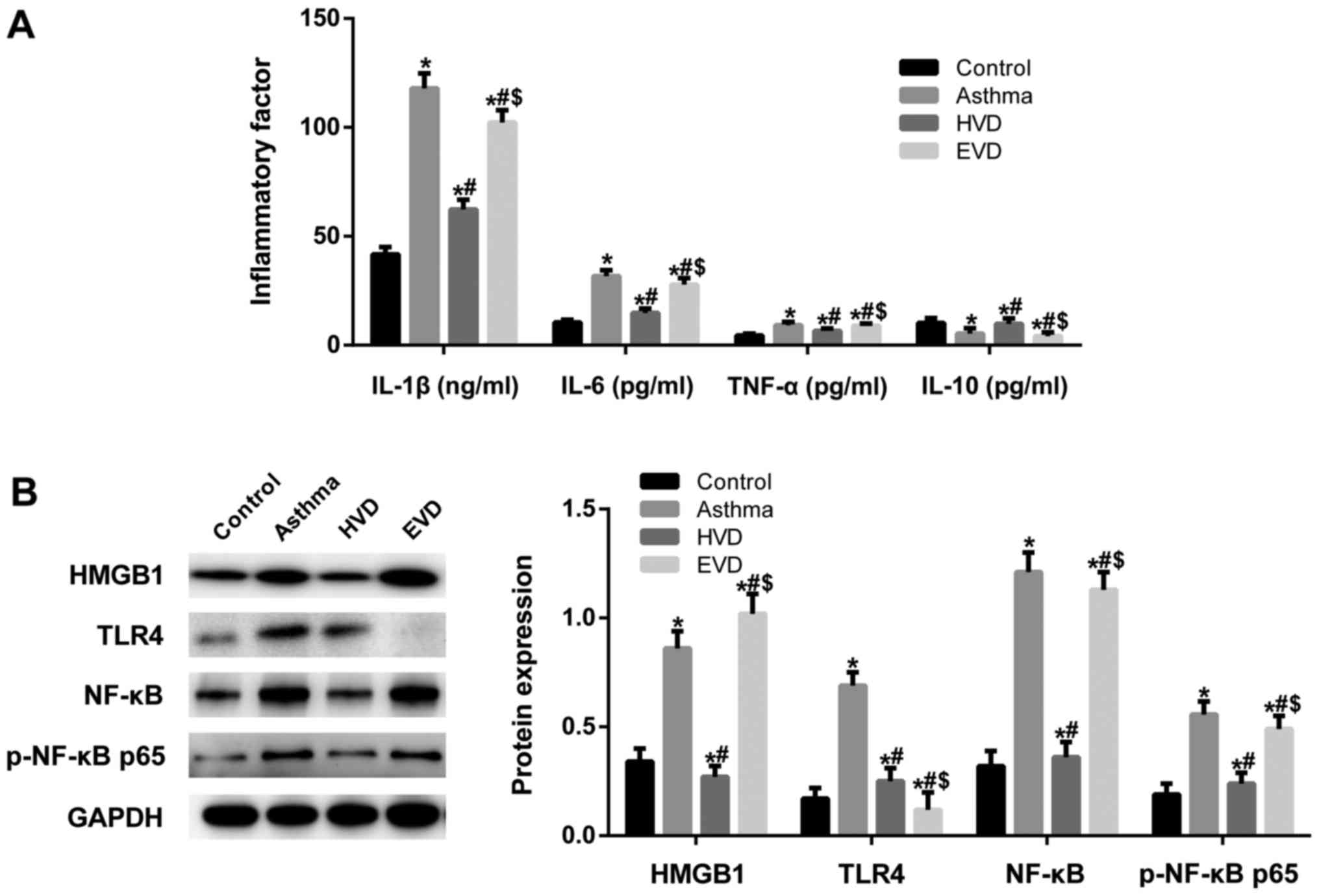
To investigate the mechanism of action of VD, we
used TLR4 inhibitor E5564. ELISA results showed that after TLR4 was
inhibited, the protective effects of VD disappeared (Fig. 5A). Western blot analysis revealed
that HMGB, TLR4, NF-κB and p-NF-κB p65 expression levels in the
lung tissue were significantly increased in the asthma group than
those in the normal control group (P<0.05; Fig. 5B). HMGB, TLR4, NF-κB and p-NF-κB
p65 expression levels were significantly decreased in the HVD, MVD
and LVD groups than those in the asthma group (P<0.05). After
the expression of TGF-β1 receptor was inhibited, HMGB, TLR4, NF-κB
and p-NF-κB p65 expression levels in the lung tissue were
significantly increased compared to those in the VD-treated groups
(P<0.05). These data suggest that VD reduces inflammatory
response in asthmatic mice through HMGB1/TLR4/NF-κB signaling
pathway.
Discussion
In this study, asthma mouse model was established,
data showed that HVD greatly attenuated the inflammatory response
of bronchial alveolar cells and reduced apoptosis. This effect of
VD is achieved through the HMGB1/TLR4/NF-κB signaling pathway.
Bronchial asthma is a common chronic respiratory
inflammatory disease that occurs mostly in the autumn and is
characterized by repeated and recurrent attacks and refractory
treatment. Glucocorticoids are still the most effective drugs
against bronchial asthma (21).
But they do not really benefit asthma patients and only reduced the
symptoms associated with asthma. Moreover, glucocorticoids have
many adverse reactions and possess a number of health risks
(22). Strong evidence exists that
VD participates in the development of allergic and autoimmune
diseases. Sadeghi et al found that VD can downregulate the
expression of TLRs in the monocytes and reduce the production of
pro-inflammatory cytokines such as TNF-α (23). Farhangi et al reported that
VD effectively reduced the oxidative stress and inflammatory
response of adipose tissue in a rat model of high fat diet-induced
obesity (24). Our results
revealed that VD treatment greatly reduced the airway inflammatory
response and cellular apoptosis in the lung tissue of a mouse model
of asthma.
The pathological mechanism of bronchial asthma is
very complex and inflammation is the pathological basis. HMGB1 as a
nucleoprotein was discovered through purification in the 1970s
together with chromosomal DNA.
Intranuclear HMGB1 mainly regulates DNA replication,
repair and transcription. When cells are stimulated by
inflammation, nuclear membrane permeability was increased, HMGB1
exudes into the cytoplasm, and then released into the extracellular
environment by neutrophils, monocytes/macrophages, and other
inflammatory cells, act as an extracellular signal transporter
through binding to other receptors on the cell surface to exert
pro-inflammatory effects (25).
TLR4 is an important pattern recognition receptor
and it is highly expressed on immune cells including macrophages,
neutrophils, and dendritic cells (26). TLR4 mainly functions to mediate the
maturation of dendritic cells, releases interleukin-12, promotes
the synthesis and release of cytokines, contributing to production
of inflammatory response (27).
TLR4 signaling is a process that activates NF-κB, initiates
inflammatory mediators-related gene transcription, and promotes the
production of inflammatory factors through myeloid differentiation
proteins (MyD88)-dependent and -independent signaling pathway
(28). HMGB1 can act as an
endogenous ligand, binds to TLR4, activates downstream NF-κB, and
induces immune response and inflammatory response (29). Lee et al found that HMGB1,
TLR2 and TLR4 expression in the lung tissue was obviously increased
in a mouse model of asthma-induced by ovalbumin (30). After anti-HMGB1 treatment, airway
inflammation, mucous membrane formation, collagen deposition, and
TLR2 and TLR4 expression were significantly reduced. Our results
showed that after inhibition of TLR4, the anti-inflammatory effects
of VD disappeared, presuming that VD may compete with HMGB1 to bind
to TLR4, thus inhibiting the pro-inflammatory effects of HMGB1.
Our results showed that VD reduced the inflammatory
response and apoptosis in lung tissue in asthmatic mice through
HMGB1/TLR4/NF-κB signaling pathway. This provides a theoretical
basis for clinical application of targeted drug therapy and
development of new drugs against asthma.
Acknowledgements
This study was supported by the Liaoning provincial
clinical capacity-building project LNCCC-C02-2015.
References
|
1
|
Vora AC: Bronchial asthma. J Assoc
Physicians India. 62 3 Suppl:S5–S6. 2014.
|
|
2
|
Yu HY, Li XY, Cai ZF, Li L, Shi XZ, Song
HX and Liu XJ: Eosinophil cationic protein mRNA expression in
children with bronchial asthma. Genet Mol Res. 14:14279–14285.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ciepiela O, Ostafin M and Demkow U:
Neutrophils in asthma-a review. Respir Physiol Neurobiol.
209:13–16. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Erokhina IL, Voronchikhin PA, Okovityĭ SV
and Emel'ianova OI: Reaction of population of pulmonary mast cells
in rat bronchial asthma under the effect of β-adrenoreceptor
antagonists. Tsitologiia. 55:472–474. 2013.(In Russian). PubMed/NCBI
|
|
5
|
Yang H, Antoine DJ, Andersson U and Tracey
KJ: The many faces of HMGB1: Molecular structure-functional
activity in inflammation, apoptosis, and chemotaxis. J Leukoc Biol.
93:865–873. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Smolarczyk R, Cichoń T, Jarosz M and Szala
S: HMGB1-its role in tumor progression and anticancer therapy.
Postepy Hig Med Dosw (Online). 66:913–920. 2012.(In Polish).
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kamo N, Ke B, Ghaffari AA, Shen XD,
Busuttil RW, Cheng G and Kupiec-Weglinski JW: ASC/caspase-1/IL-1β
signaling triggers inflammatory responses by promoting HMGB1
induction in liver ischemia/reperfusion injury. Hepatology.
58:351–362. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang T, Wei XY, Liu B, Wang LJ and Jiang
LH: Effects of propofol on lipopolysaccharide-induced expression
and release of HMGB1 in macrophages. Braz J Med Biol Res.
48:286–291. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Willenbrock S, Braun O, Baumgart J, Lange
S, Junghanss C, Heisterkamp A, Nolte I, Bullerdiek J and Murua
Escobar H: TNF-α induced secretion of HMGB1 from non-immune canine
mammary epithelial cells (MTH53A). Cytokine. 57:210–220. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fink MP: HMGB1 as a drug target in
staphylococcal pneumonia. Crit Care. 18:1312014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang WJ, Yin SJ and Rong RQ: PKR and HMGB1
expression and function in rheumatoid arthritis. Genet Mol Res.
14:17864–17870. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang H, Ward MF and Sama AE: Targeting
HMGB1 in the treatment of sepsis. Expert Opin Ther Targets.
18:257–268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu X, Yu Y, Eugene Chin Y and Xia Q: The
role of acetylation in TLR4-mediated innate immune responses.
Immunol Cell Biol. 91:611–614. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pascual M, Fernández-Lizarbe S and Guerri
C: Role of TLR4 in ethanol effects on innate and adaptive immune
responses in peritoneal macrophages. Immunol Cell Biol. 89:716–727.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wan Z, Zhang X, Peng A, He M, Lei Z and
Wang Y: TLR4-HMGB1 signaling pathway affects the inflammatory
reaction of autoimmune myositis by regulating MHC-I. Int
Immunopharmacol. 41:74–81. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kang N, Hai Y, Yang J, Liang F and Gao CJ:
Hyperbaric oxygen intervention reduces secondary spinal cord injury
in rats via regulation of HMGB1/TLR4/NF-κB signaling pathway. Int J
Clin Exp Pathol. 8:1141–1153. 2015.PubMed/NCBI
|
|
17
|
Sperl A and Klimek L: Role of vitamin D in
allergic diseases: Current research status. HNO. 63:352–356.
2015.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Agmon-Levin N, Theodor E, Segal RM and
Shoenfeld Y: Vitamin D in systemic and organ-specific autoimmune
diseases. Clin Rev Allergy Immunol. 45:256–266. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maalmi H, Sassi FH, Berraies A, Ammar J,
Hamzaoui K and Hamzaoui A: Association of vitamin D receptor gene
polymorphisms with susceptibility to asthma in Tunisian children: A
case control study. Hum Immunol. 74:234–240. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Amento EP: Vitamin D and the immune
system. Steroids. 49:55–72. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
EmeI'yanov AV: The therapeutic potential
of inhalation glucocorticoids in patients with bronchial asthma.
Klin Med (Mosk). 93:23–29. 2015.(In Russian).
|
|
22
|
Oray M, Abu Samra K, Ebrahimiadib N, Meese
H and Foster CS: Long-term side effects of glucocorticoids. Expert
Opin Drug Saf. 15:457–465. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sadeghi K, Wessner B, Laggner U, Ploder M,
Tamandl D, Friedl J, Zügel U, Steinmeyer A, Pollak A, Roth E, et
al: Vitamin D3 down-regulates monocyte TLR expression and triggers
hyporesponsiveness to pathogen-associated molecular patterns. Eur J
Immunol. 36:361–370. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Farhangi MA, Mesgari-Abbasi M, Hajiluian
G, Nameni G and Shahabi P: Adipose tissue inflammation and
oxidative stress: The ameliorative effects of vitamin D.
Inflammation. Jul 3–2017.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang D, Shi Y, Kang R, Li T, Xiao W, Wang
H and Xiao X: Hydrogen peroxide stimulates macrophages and
monocytes to actively release HMGB1. J Leukoc Biol. 81:741–747.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ottow MK, Klaver EJ, van der Pouw Kraan
TC, Heijnen PD, Laan LC, Kringel H, Vogel DY, Dijkstra CD, Kooij G
and van Die I: The helminth Trichuris suis suppresses TLR4-induced
inflammatory responses in human macrophages. Genes Immun.
15:477–486. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim HS and Chung DH: TLR4-mediated IL-12
production enhances IFN-γ and IL-1β production, which inhibits
TGF-β production and promotes antibody-induced joint inflammation.
Arthritis Res Ther. 14:R2102012. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu HT, Bian C, Yuan JC, Chu WH, Xiang X,
Chen F, Wang CS, Feng H and Lin JK: Curcumin attenuates acute
inflammatory injury by inhibiting the TLR4/MyD88/NF-κB signaling
pathway in experimental traumatic brain injury. J
Neuroinflammation. 11:592014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qiao JY, Song L, Zhang YL and Luan B:
HMGB1/TLR4/NF-κB signaling pathway and role of vitamin D in
asthmatic mice. Zhongguo Dang Dai Er Ke Za Zhi. 19:95–103. 2017.(In
Chinese). PubMed/NCBI
|
|
30
|
Lee CC, Lai YT, Chang HT, Liao JW, Shyu
WC, Li CY and Wang CN: Inhibition of high-mobility group box 1 in
lung reduced airway inflammation and remodeling in a mouse model of
chronic asthma. Biochem Pharmacol. 86:940–949. 2013. View Article : Google Scholar : PubMed/NCBI
|