Introduction
Breast cancer is the most rapidly increasing
malignancy in females worldwide (1). Due to the rising incidence of breast
cancer (2), there is a growing
concern about the matter among the general public. The critical
challenge in the treatment of patients with breast cancer is tumor
recurrence and metastasis following surgery. Therefore, early
metastatic risk assessment and treatment is necessary for
individualized therapy. Previous evidence has shown that breast
tumors are heterogeneous in vitro and in vivo
(3–5). Certain subpopulations of breast
tumors cells may possess a specific metastatic potential, and
exploring this function may provide improved understanding about
breast cancer. Metastasis is a key cause for cancer morbidity and
mortality (6,7). Metastasis involves numerous cell
processes, including proliferation, invasion and angiogenesis in
the tumor microenvironment. However, these processes have not yet
been fully elucidated.
Chemokines and chemokine receptors perform an
important role in tumor cell growth, survival, adhesion and
metastasis. Previously, stromal cell-derived factor-1 (CXCL12) and
its chemokine receptor CXCR7 were reported to regulate tumor cell
function in numerous cancers, including lung and kidney cancer
(8–12). CXCL12 is involved in a number of
important physiological steps, including vasculogenesis. Notably,
CXCL12 regulates proliferation, migration and invasion in numerous
tumor cells (13–16). In addition, CXCR7 is overexpressed
in malignant cells and vascular endothelium. CXCR7 critically
controls the cardiovascular system development in animal models.
Decreased CXCR7 expression in zebrafish embryos inhibits blood
vessel formation (17), and the
knock down of CXCR7 in mice causes early postnatal mortality as a
result of myocardial degeneration and heart vessel damage (18). Evidence from a previous study
demonstrated that CXCR7 induces tumor growth, invasion and
metastasis (19–22). Thus, it is necessary to explore the
role of CXCR7 in breast cancer progress.
It has been shown that CXCR7 promotes tumor growth
in a mouse model of lung cancer, and that expression of CXCR7
affects experimental lung metastasis (23). In addition, CXCR7 enhances the cell
adhesion, invasion and blood vessel sprout formation in
vitro, and promotes tumor growth in vivo (24,25).
Other studies have revealed that CXCR7 mediated the proliferation
and migration of tumor cells towards CXCL12 in vitro
(22,26–28).
All results proposed that CXCR7 may perform an important role in
breast cancer. Although the role of CXCL12 in the tumor is
extensively documented and CXCR4 activation signals have been
reported, the role of CXCR7 in regulating breast cancer is not
known. Thus, it is necessary to investigate the function of CXCR7
in breast cancer development.
In the present study, the effects of CXCR7 in breast
cancer invasion, migration and angiogenesis were investigated.
Materials and methods
Cell culture
The MDA-MB-231 cell line was obtained from American
Type Culture Collection (Manassas, VA, USA) and grown in Dulbecco's
modified Eagle's medium (DMEM; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), supplemented with 10% fetal
bovine serum, 5 U/ml of penicillin and 5 mg/ml of streptomycin.
HUVECs (American Type Culture Collection, Manassas, VA, USA) were
maintained in endothelial cell growth medium (PromoCell GmbH,
Heidelberg, Germany) containing endothelial cell growth supplement,
and HUVECs were used at passage 3–5.
Chemokines and reagents
Recombinant CXCL12/SDF-1α was obtained from R&D
Systems, Inc. (cat. no. 350-NS; Minneapolis, MN, USA). The CXCR7
antagonist CCX771 was obtained from ChemoCentryx, Inc. (Mountain
View, CA, USA). Calcein-AM was purchased from Sigma-Aldrich (cat.
no. 148504-34-1; Merck KGaA, Darmstadt, Germany).
Cell invasion assay
The upper surface of a modified Boyden chamber
(Corning, Inc., Corning, NY, USA) was pre-treated with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA). MDA-MB-231 cells were
treated with 5 µm CCX771 for 1 h. A total of 2×104 cells
were added to the upper Boyden chamber. Serum-free DMEM media (0.5
ml) containing CXCL12 (0–100 ng/ml) was then added to the lower
chamber for 24 h. The noninvasive cells were gently removed
following incubation. The invasive cells at the bottom of the
Matrigel were stained by Calcein-AM. The number of invasive cells
was counted under an inverted fluorescent microscope (IX51; Olympus
Corporation, Tokyo, Japan) in at least three fields (magnification,
×10).
Cell migration assays
Cell migration assays were performed using a
modified Boyden chamber (BD Biosciences). MDA-MB-231 was
pre-treated with 5 µm CCX771 for 1 h at 37°C. A total of
2×103 MDA-MB-231 cells per well were added to the upper
of the Boyden chamber. CXCL12 (100 ng/ml) was added to the lower
chamber with DMEM media. The Boyden chamber was incubated at 37°C
for 5 h. The migrated cells on the lower side of the filter were
stained by Calcein-AM and the migration of cells was quantified.
All experiments were repeated three times in three wells.
Cell adhesion assay
A cell adhesion assay was performed using the
CytoSelect™ extracellular matrix cell adhesion assay kit
(Cell BioLabs, Inc., San Diego, CA, USA), according to the
manufacturer's protocol. The 48-well plates were pre-coated with
laminin (LN) or fibronectin (FN) for 1 h at 37°C. MDA-MB-231was
pre-treated with 5 µm CCX771 and/or CXCL12 (100 ng/ml) for 24 h at
37°C. A total of 2×104 cells/well were added to the
plate for 1 h at 37°C. Non-adhesive cells were then removed using
PBS. The adhesive cells were then measured by absorbance at 560 nm
with a microplate reader. All the experiments were repeated three
times in three wells.
Tube formation assay
MDA-MB-231 cells were treated with 5 µm CCX771
and/or 100 ng/ml CXCL12, and the media were collected 24 h
following treatment. The wells of 96-well plates were pre-coated
with Matrigel for 1 h at 37°C. A total of 2×104 HUVECs
were seeded onto wells with the aforementioned media for 24 h.
Images were captured with a Leica Microsystems microscope (10X
objective; Leica Microsystems, Inc., Buffalo Grove, IL, USA). Only
perfectly continuous tubes between two branching points were
considered as a tube formation. At least 4 fields were tested per
well and experiments were repeated three times.
ELISA
The human VEGF Quantikine ELISA kit (R&D
Systems, Inc.) was used for ELISA. The MDA-MB-231 cells were
cultured for 24 h and the supernatant was collected by
centrifugation at 1,000 × g for 10 min at room temperature. VEGF
secretion was measured using ELISA. In brief, 50 µl of sample or
standard was added to the microplate wells at room temperature for
2 h, and 100 µl of VEGF conjugate was added at room temperature for
1 h. Microplates were washed with PBS and 200 µl of substrate
solution was added. The optical density (OD) was then read at 450
nm using an ELISA plate reader.
Cell proliferation assay
Cell proliferation was measured by MTT assay
(Promega Corporation, Madison, WI, USA). MDA-MB-231 was pre-treated
with 5 µm CCX771, and 5×104 cells/well were seeded onto
96-well dishes in DMEM containing CXCL12 (100 ng/ml) for 72 h. The
OD was measured with a microplate reader at 570 nm.
Matrigel plug assay in vivo
Aliquots of 0.5 ml of Matrigel (containing 5 µm
CCX771 and/or 100 ng/ml CXCL12) were injected subcutaneously into
the sides of BALB/c mice (all male, aged 8–12 weeks). All animal
experiments were approved by the Committee on Animal Welfare of the
Tongji University (Shanghai, China). The animals were housed at
24°C with a 12 h light/dark cycle and were checked daily for
mortality. Animals were euthanized by CO2 and the
Matrigel plugs were excised after 10 days. Matrigel plugs from the
control and drug treated groups were stained with hematoxylin and
eosin.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Statistically significance of differences between untreated control
and drug-treated cells was evaluated using Student's one-tailed
t-test. P<0.05 was considered to indicate a statistically
significant difference.
Results
CXCR7 regulates CXCL12-induced
enhancement on breast carcinoma cell invasion in vitro
CXCR7 has an important role in the invasion of
several tumors. It is therefore of interest to explore whether
CXCR7 affects breast cancer cell invasion by reducing CXCR7
expression using the CXCR7 antagonist CCX771. Cell invasion
experiments used a Matrigel invasion chamber, which is considered
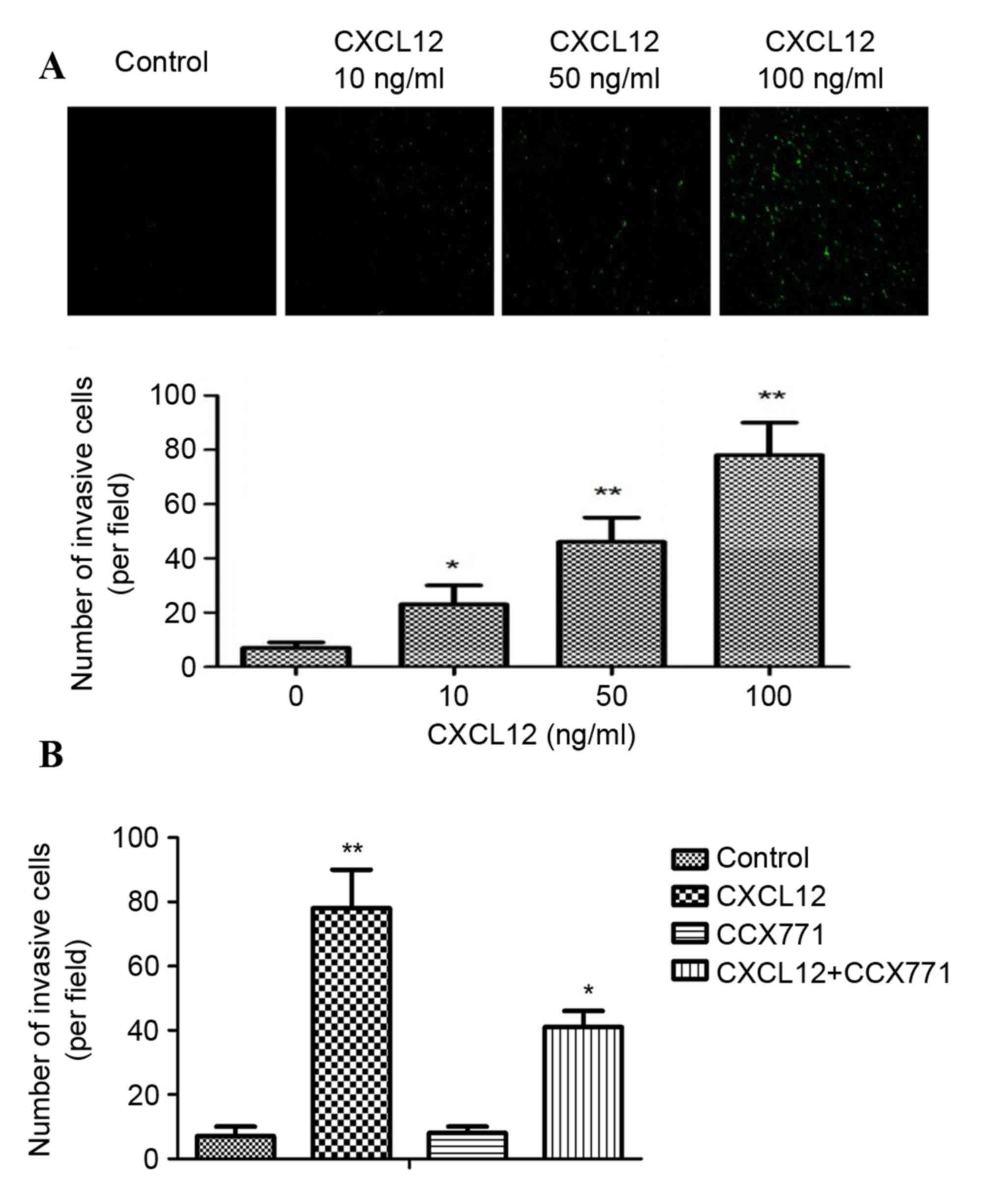
an in vitro model for metastasis research. As shown in
Fig. 1A, CXCL12 induced
significant and dose-dependent MDA-MB-231 cell invasion through
Matrigel. In addition, the inhibition of CXCR7 on MDA-MB-231 cells
reduced invasive ability compared with CXCL12 treated cells
(Fig. 1B). These data indicated
that CXCL12 induces invasive behavior of MDA-MB-231 cells, and that
inhibition of CXCR7 reduces the invasive ability of cells.
CXCR7 regulates CXCL12-induced
enhancement on breast carcinoma cell migration in vitro
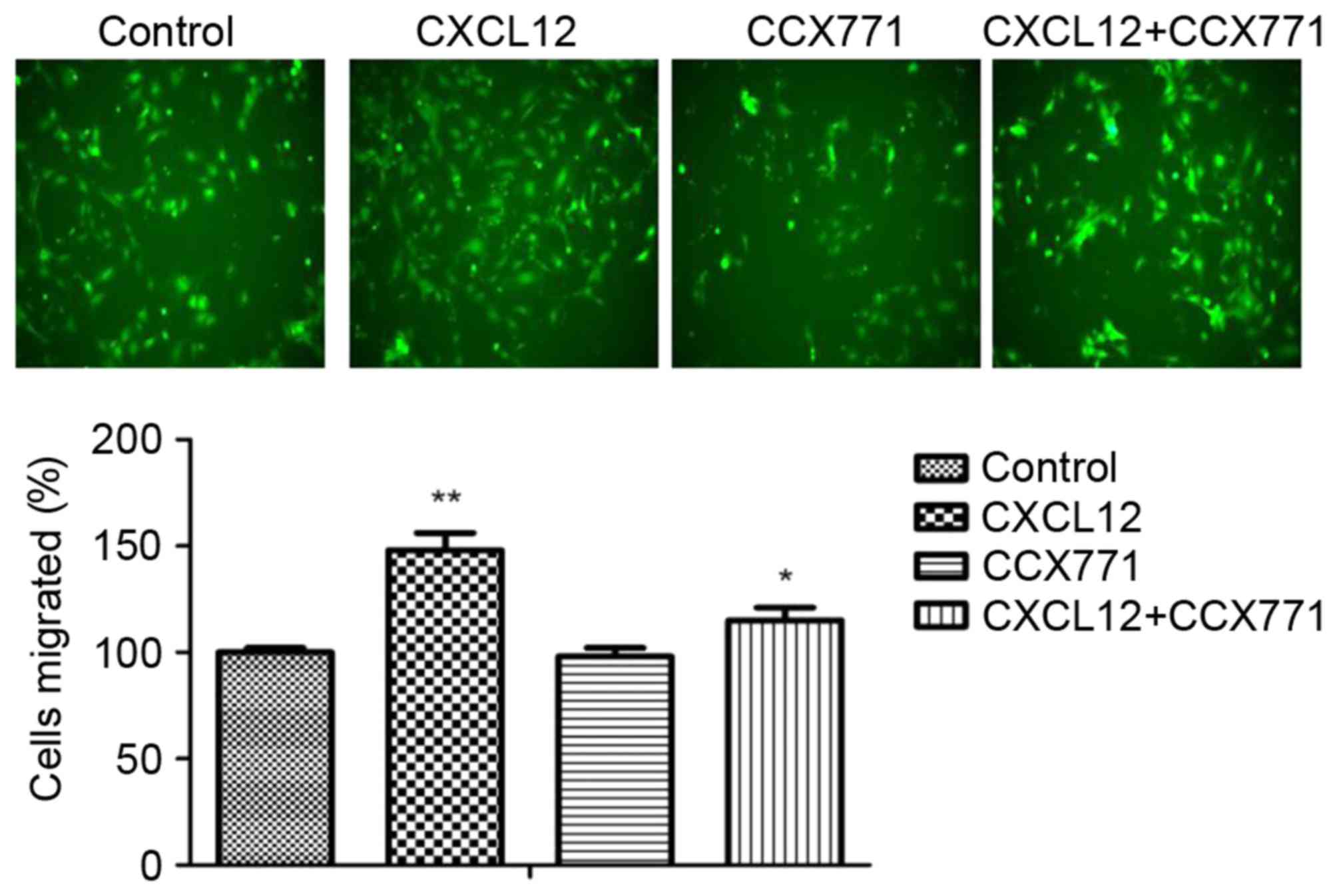
The enhanced migration behavior of tumor cells
decided their metastatic phenotype. To determine whether CXCR7
affects breast cancer cells migration, a Transwell migration assay
was performed. CXCL12 significantly increased MDA-MB-231 cell
migration capability, and the migration cells were markedly reduced
in the presence of CCX771 (Fig.
2). The present results indicated that CXCL12 induces
MDA-MB-231 cell migration behavior and that inhibition of CXCR7
reduces the cell migration ability.
CXCR7 regulates CXCL12-induced
enhancement on breast carcinoma cell adhesion in vitro
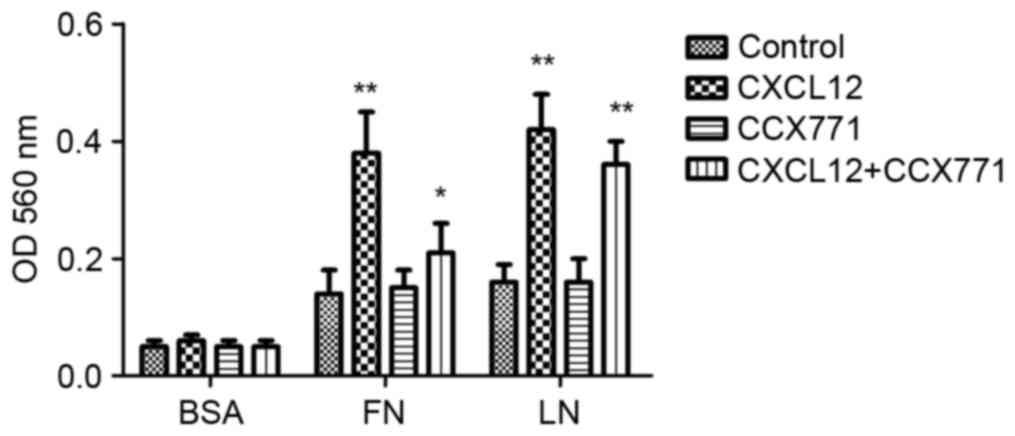
Tumor cell adhesion is a key process in tumor
invasion. To examine the mechanism by which CXCR7 affects
MDA-MB-231 cell adhesion to LN or FN, a cell adhesion assay was
performed. As shown in Fig. 3,
CXCL12-treated MDA-MB-231 cells demonstrated increased cell
adhesion to LN or FN. However, CXCR7 inhibition using CCX771 in
MDA-MB-231 cells showed significantly reduced ability of adhesion
to LN or FN. These results indicated that CXCL12 increases adhesive
ability of MDA-MB-231 cells and that inhibiting of CXCR7 decreased
adhesive ability.
CXCR7 regulates CXCL12-induced
enhancement of breast carcinoma cell angiogenic processes in
vitro
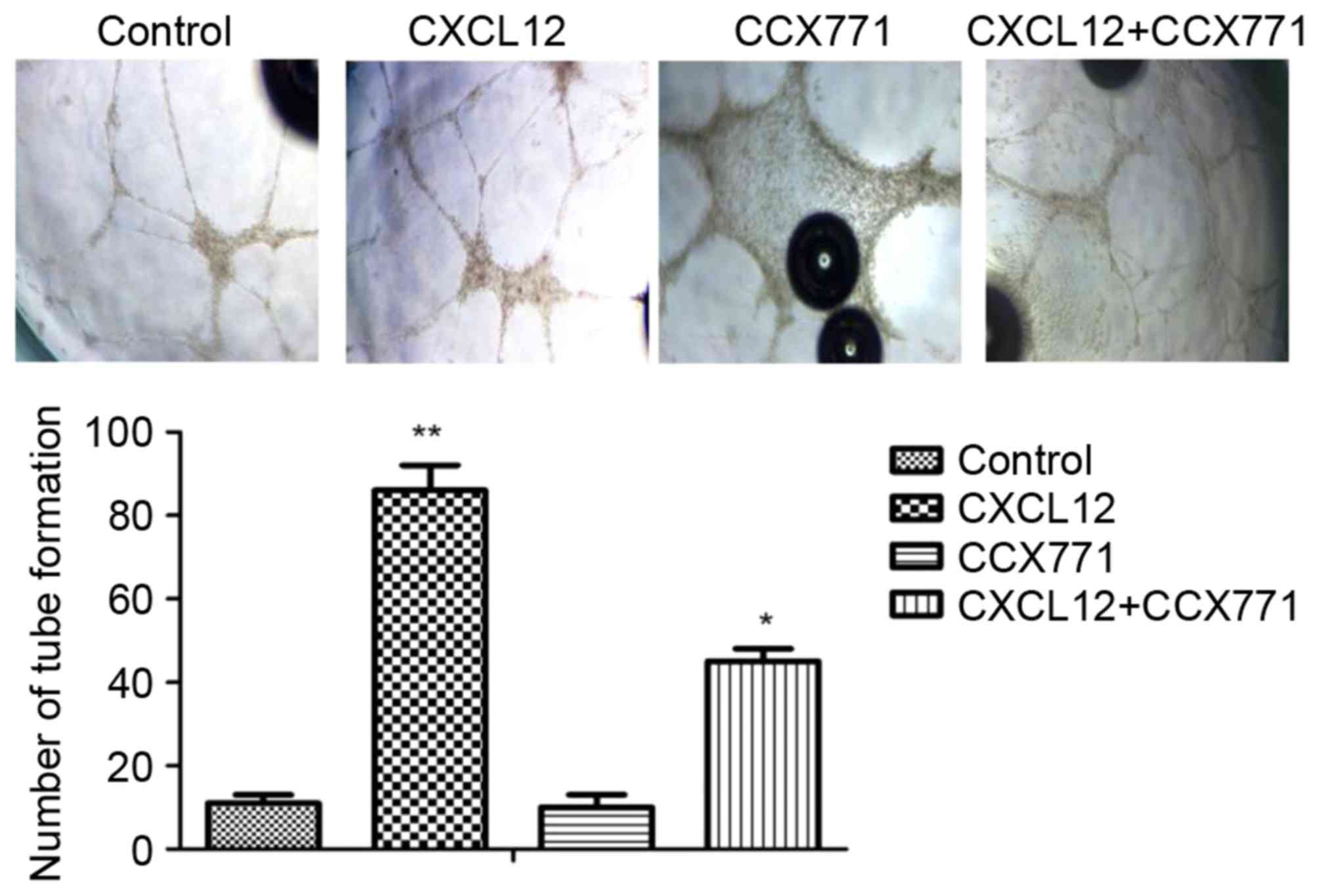
To explore whether CXCR7 mediates tube formation
in vitro, the Matrigel tube formation assay was used. The
amount of tube formation was quantified. As shown in Fig. 4, the CXCL12-treated MDA-MB-231
cells induced HUVECs form capillary-like structures within 24 h. By
contrast, MDA-MB-231 cells treated with CCX771 markedly inhibited
tube formation. Therefore, CXCL12 increased angiogenic processes of
MDA-MB-231 cells, whereas inhibition of CXCR7 decreased angiogenic
processes.
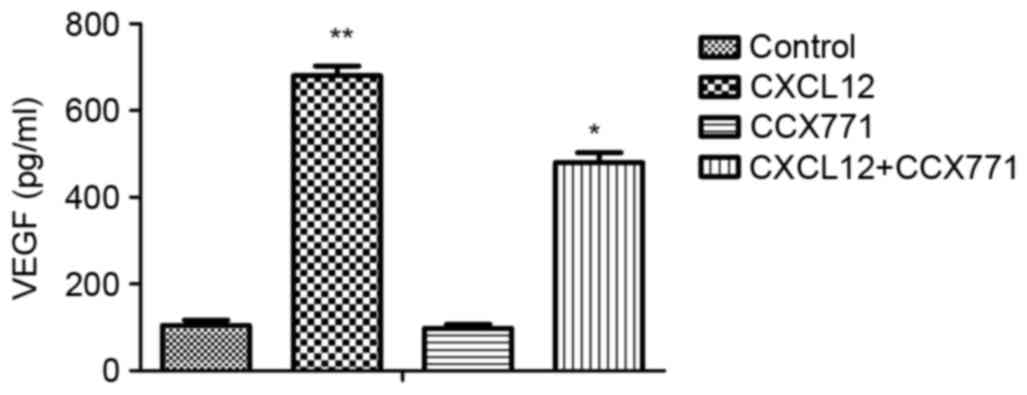
CXCR7 regulates the secretion of
VEGF
To explore the possible effects of CXCR7 on the
regulation of proangiogenic factor secretion, ELISA was performed.
The supernatant from MDA-MB-231 cells treated with CCX771 and/or
CXCL12 were tested. The results demonstrated that CXCL12 increased
VEGF level and blocking of CXCR7 significantly decreased the VEGF
level in MDA-MB-231 cells (Fig.
5).
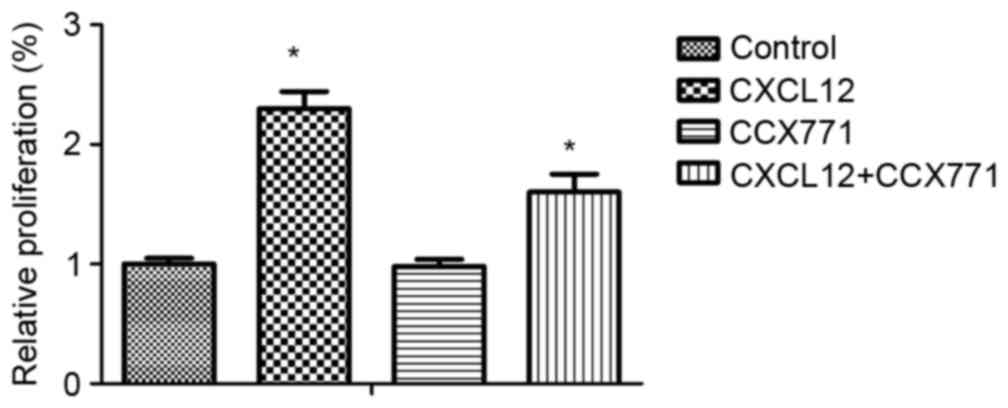
CXCR7 regulates CXCL12-induced
enhancement of breast carcinoma cell proliferation in vitro
To explore the possible effects of CXCR7 on breast
cancer cell proliferation, MDA-MB-231 cells were treated with
CCX771 and/or CXCL12, and proliferation rates were determined. The
results demonstrated that CXCL12 increased MDA-MB-231 cell
proliferation, and blocking of CXCR7 significantly decreased
MDA-MB-231 cell proliferation (Fig.
6).
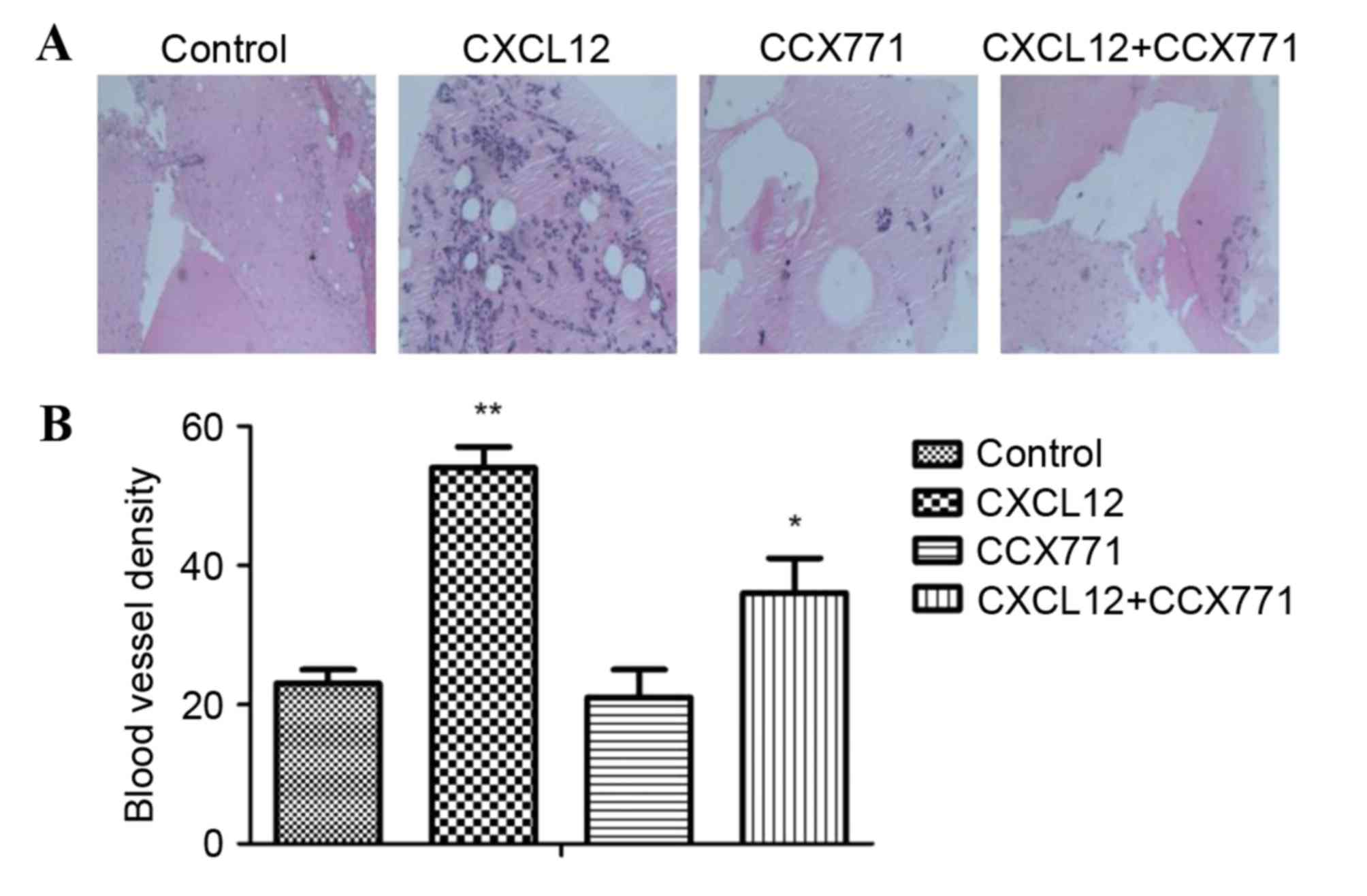
CXCR7 regulates angiogenic activity in
vivo
The new vessel formation was analyzed by Matrigel
plug assays in C57BL/6 mice. Matrigel plugs containing CXCL12 were
filled with an abundance of cellular infiltration, demonstrating
that vascular elements were formed in the Matrigel. By contrast,
the vessel formation was significantly reduced by CCX771 treatment,
demonstrating the anti-angiogenic activity in vivo (Fig. 7).
Discussion
Increasing evidence has demonstrated that CXCL12 and
its receptors are involved in cancer cellular proliferation,
invasion, metastasis and angiogenesis (29,30).
CXCR7 overexpression has been identified in numerous human cancers,
including breast carcinoma cells (31,32).
However, the role of CXCR7 in regulating breast carcinoma cells is
unclear. To evaluate the role of CXCR7 in breast carcinoma cells,
the MDA-MB-231 cell line was selected as an in vitro model.
In the present study, CXCL12 enhanced MDA-MB-231 cell invasion,
migration, adhesion and angiogenesis. The present study also
indicated that CXCR7 regulated breast carcinoma cell invasion,
migration adhesion and angiogenesis. In the present study, the
molecular mechanisms and signaling pathways in breast carcinoma
cells were not demonstrated. Thus, additional studies exploring
CXCR7-activated pathways are required.
In the present study, CXCL12 was revealed to promote
MDA-MB-231cell invasion and migration. It was also examined whether
CXCR7 could affect MDA-MB-231cell invasion and migration ability by
invasion assay and Transwell assays. The present results revealed
that CCX771 decreased cell migration and invasion. Tumor cell
contact with basement membranes is an essential step for invasion.
The previous study demonstrated that tumor cells adhere to
endothelial cells through regulation of CXCR7 (33,34).
The present data demonstrated that CXCL12 induces MDA-MB-231 cell
adhesion to FN and LN. Increased cell-matrix adhesion may induce
tumor cell metastasis. In addition, CCX771 was revealed to inhibit
adhesion of MDA-MB-231 cells to LN or FN. Therefore, the present
findings indicated that CXCR7 is involved in CXCL12-induced
cell-matrix adhesion of MDA-MB-231 cells.
Angiogenesis is a complex process, involving cell
migration, proliferation and tube formation. Tumor cell survival
and proliferation relies on angiogenesis (30). CXCR7 was identified to be involved
in tube formation in vitro, which may induce tumor growth
(35). Although CXCL12-stimulated
VEGF expression has been reported in various cells, to the best of
our knowledge, CXCL12-regulated VEGF secretion in breast carcinoma
cells has not been studied yet. In the present study, CXCL12 was
found to increase VEGF secretion. Furthermore, the present data
also demonstrated that inhibition of CXCR7 decreased VEGF level and
tube formation, indicating that CXCR7 may be involved in
angiogenesis in breast carcinoma.
The present findings indicated that CXCR7 regulates
multiple processes in breast carcinoma. CXCL12 regulated breast
carcinoma invasion, adhesion, migration, angiogenesis and VEGF
secretion. Therefore, the effects of CXCR7 on cell invasion,
adhesion, migration, tube formation in vitro and Matrigel
plug assay in vivo were examined. The present study provided
mechanistic evidence that CXCR7 may affect breast carcinoma
progression by numerous mechanisms, including adhesion, invasion
and angiogenesis. The present data demonstrated that CCX771
significantly inhibited invasion, adhesion, migration and
angiogenesis in vitro and in vivo. Although the
present study demonstrated the importance of CXCR7 in breast
carcinoma, the signaling pathway of CXCR7 in tumor progression was
not fully established. Greater attention to the function of CXCR7
in cancer is expected in the future. Thus, our studies elucidating
the CXCR7 may be a novel target for tumor therapy.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81271694, 81301157,
81301310 and 31301523) and the Science and Technology Commission of
Shanghai Municipality (grant nos. 11411951500 and 12nm0502200).
References
|
1
|
Ennour-Idrissi K, Maunsell E and Diorio C:
Telomere length and breast cancer prognosis: A systematic review.
Cancer Epidemiol Biomarkers Prev. 26:3–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dhennin-Duthille I, Gautier M, Faouzi M,
Guilbert A, Brevet M, Vaudry D, Ahidouch A, Sevestre H and
Ouadid-Ahidouch H: High expression of transient receptor potential
channels in human breast cancer epithelial cells and tissues:
Correlation with pathological parameters. Cell Physiol Biochem.
28:813–822. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pieta B, Chmaj-Wierzchowska K and Opala T:
Life style and risk of development of breast and ovarian cancer.
Ann Agric Environ Med. 19:379–384. 2012.PubMed/NCBI
|
|
4
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan L, Strasser-Weippl K, Li JJ, St Louis
J, Finkelstein DM, Yu KD, Chen WQ, Shao ZM and Goss PE: Breast
cancer in China. Lancet Oncol. 15:e279–e289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Christofori G: Cancer: Division of labour.
Nature. 446:735–736. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Podsypanina K, Du YC, Jechlinger M,
Beverly LJ, Hambardzumyan D and Varmus H: Seeding and propagation
of untransformed mouse mammary cells in the lung. Science.
321:1841–1844. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Akashi T, Koizumi K, Tsuneyama K, Saiki I,
Takano Y and Fuse H: Chemokine receptor CXCR4 expression and
prognosis in patients with metastatic prostate cancer. Cancer Sci.
99:539–542. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu TP, Shen H, Liu LX and Shu YQ: The
impact of chemokine receptor CXCR4 on breast cancer prognosis: A
meta-analysis. Cancer Epidemiol. 37:725–731. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wald O, Shapira OM and Izhar U:
CXCR4/CXCL12 axis in non small cell lung cancer (NSCLC) pathologic
roles and therapeutic potential. Theranostics. 3:26–33. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Miao Z, Luker KE, Summers BC, Berahovich
R, Bhojani MS, Rehemtulla A, Kleer CG, Essner JJ, Nasevicius A,
Luker GD, et al: CXCR7 (RDC1) promotes breast and lung tumor growth
in vivo and is expressed on tumor-associated vasculature. Proc Natl
Acad Sci USA. 104:pp. 15735–15740. 2007; View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang J, Shiozawa Y, Wang J, Wang Y, Jung
Y, Pienta KJ, Mehra R, Loberg R and Taichman RS: The role of
CXCR7/RDC1 as a chemokine receptor for CXCL12/SDF-1 in prostate
cancer. J Biol Chem. 283:4283–4294. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Marchesi F, Monti P, Leone BE, Zerbi A,
Vecchi A, Piemonti L, Mantovani A and Allavena P: Increased
survival, proliferation, and migration in metastatic human
pancreatic tumor cells expressing functional CXCR4. Cancer Res.
64:8420–8427. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sutton A, Friand V, Brulé-Donneger S,
Chaigneau T, Ziol M, Sainte-Catherine O, Poiré A, Saffar L, Kraemer
M, Vassy J, et al: Stromal cell-derived factor-1/chemokine (C-X-C
motif) ligand 12 stimulates human hepatoma cell growth, migration,
and invasion. Mol Cancer Res. 5:21–33. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Burns JM, Summers BC, Wang Y, Melikian A,
Berahovich R, Miao Z, Penfold ME, Sunshine MJ, Littman DR, Kuo CJ,
et al: A novel chemokine receptor for SDF-1 and I-TAC involved in
cell survival, cell adhesion, and tumor development. J Exp Med.
203:2201–2213. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mazzinghi B, Ronconi E, Lazzeri E,
Sagrinati C, Ballerini L, Angelotti ML, Parente E, Mancina R, Netti
GS, Becherucci F, et al: Essential but differential role for CXCR4
and CXCR7 in the therapeutic homing of human renal progenitor
cells. J Exp Med. 205:479–490. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Perlin JR and Talbot WS: Signals on the
move: Chemokine receptors and organogenesis in zebrafish. Sci STKE.
2007:pe452007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gerrits H, van Ingen Schenau DS, Bakker
NE, van Disseldorp AJ, Strik A, Hermens LS, Koenen TB,
Krajnc-Franken MA and Gossen JA: Early postnatal lethality and
cardiovascular defects in CXCR7-deficient mice. Genesis.
46:235–245. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Q, Zhang A, Tao C, Li X and Jin P: The
role of SDF-1-CXCR4/CXCR7 axis in biological behaviors of adipose
tissue-derived mesenchymal stem cells in vitro. Biochem Biophys Res
Commun. 441:675–680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zheng K, Li HY, Su XL, Wang XY, Tian T, Li
F and Ren GS: Chemokine receptor CXCR7 regulates the invasion,
angiogenesis and tumor growth of human hepatocellular carcinoma
cells. J Exp Clin Cancer Res. 29:312010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Singh RK and Lokeshwar BL: The
IL-8-regulated chemokine receptor CXCR7 stimulates EGFR signaling
to promote prostate cancer growth. Cancer Res. 71:3268–3277. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hao M, Zheng J, Hou K and Wang J, Chen X,
Lu X, Bo J, Xu C, Shen K and Wang J: Role of chemokine receptor
CXCR7 in bladder cancer progression. Biochem Pharmacol. 84:204–214.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xue TC, Chen RX, Han D, Chen J, Xue Q, Gao
DM, Sun RX, Tang ZY and Ye SL: Down-regulation of CXCR7 inhibits
the growth and lung metastasis of human hepatocellular carcinoma
cells with highly metastatic potential. Exp Ther Med. 3:117–123.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kollmar O, Rupertus K, Scheuer C, Nickels
RM, Haberl GC, Tilton B, Menger MD and Schilling MK: CXCR4 and
CXCR7 regulate angiogenesis and CT26.WT tumor growth independent
from SDF-1. Int J Cancer. 126:1302–1315. 2010.PubMed/NCBI
|
|
25
|
Bauerle KT, Schweppe RE, Lund G, Kotnis G,
Deep G, Agarwal R, Pozdeyev N, Wood WM and Haugen BR: Nuclear
factor κB-dependent regulation of angiogenesis, and metastasis in
an in vivo model of thyroid cancer is associated with secreted
interleukin-8. J Clin Endocrinol Metab. 99:E1436–E1444. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qin Y, Zhou Z, Zhang F, Wang Y, Shen B,
Liu Y, Guo Y, Fan Y and Qiu J: Induction of regulatory B-cells by
mesenchymal stem cells is affected by SDF-1α-CXCR7. Cell Physiol
Biochem. 37:117–130. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou SM, Zhang F, Chen XB, Jun CM, Jing X,
Wei DX, Xia Y, Zhou YB, Xiao XQ, Jia RQ, et al: miR-100 suppresses
the proliferation and tumor growth of esophageal squamous cancer
cells via targeting CXCR7. Oncol Rep. 35:3453–3459. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Inaguma S, Riku M, Ito H, Tsunoda T, Ikeda
H and Kasai K: GLI1 orchestrates CXCR4/CXCR7 signaling to enhance
migration and metastasis of breast cancer cells. Oncotarget.
6:33648–33657. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Young B, Purcell C, Kuang YQ, Charette N
and Dupré DJ: Superoxide dismutase 1 regulation of CXCR4-mediated
signaling in prostate cancer cells is dependent on cellular
oxidative state. Cell Physiol Biochem. 37:2071–2084. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang S, Wang Y, Chen M, Sun L, Han J,
Elena VK and Qiao H: CXCL12 methylation-mediated epigenetic
regulation of gene expression in papillary thyroid carcinoma. Sci
Rep. 7:440332017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen Y, Teng F, Wang G and Nie Z:
Overexpression of CXCR7 induces angiogenic capacity of human
hepatocellular carcinoma cells via the AKT signaling pathway. Oncol
Rep. 36:2275–2281. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guo JC, Li J, Zhou L, Yang JY, Zhang ZG,
Liang ZY, Zhou WX, You L, Zhang TP and Zhao YP: CXCL12-CXCR7 axis
contributes to the invasive phenotype of pancreatic cancer.
Oncotarget. 7:62006–62018. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wurth R, Barbieri F, Bajetto A, Pattarozzi
A, Gatti M, Porcile C, Zona G, Ravetti JL, Spaziante R and Florio
T: Expression of CXCR7 chemokine receptor in human meningioma cells
and in intratumoral microvasculature. J Neuroimmunol. 234:115–123.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Totonchy JE, Osborn JM, Botto S, Clepper L
and Moses AV: Aberrant proliferation in CXCR7+ endothelial cells
via degradation of the retinoblastoma protein. PloS One.
8:e698282013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Barbieri F, Thellung S, Würth R, Gatto F,
Corsaro A, Villa V, Nizzari M, Albertelli M, Ferone D and Florio T:
Emerging targets in pituitary adenomas: Role of the CXCL12/CXCR4-R7
System. Int J Endocrinol. 2014:7535242014. View Article : Google Scholar : PubMed/NCBI
|