Introduction
Rheumatoid arthritis (RA) is a systemic and chronic
inflammatory disease affected by genetic and environmental factors,
which leads to joint destruction and dysfunction. Individuals with
RA frequently possess autoantibodies against citrullinated
peptides, thus indicating that peptidyl arginine deiminase type IV
(PADI4) may be associated with RA. It has previously been reported
that PADI4 haplotypes associated with susceptibility to RA increase
the production of citrullinated peptides as autoantigens, leading
to an increased risk of developing RA (1).
PADI4 is localized in the cytoplasm of monocytes, T
and B cells, neutrophils, eosinophils and natural killer cells, and
can translocate to the nucleus upon cell activation (2). PADI4 is involved in the
post-translational conversion of arginine residues to citrulline.
Therefore, it has been suggested that increased translation of
variant PADI4 mRNA promotes citrullinated peptide production, which
acts as an autoantigen and causes marked adaptive immune responses
(3). To date, the association
between the PADI4 gene and the severity of RA has been reflected
with regards to the correlation between PADI4 haplotypes (or
alleles) and serum titer (or positivity) of the anti-cyclic
citrullinated peptides antibody (ACPA) (4–6).
However, PADI4 may serve an additional role in the occurrence and
development of RA, in addition to its role in ACPA formation
(7).
A substantial increase in the number of resident
synovial cells is an important pathophysiological characteristic in
patients with RA (8). One of the
key components of invasive synovium is RA fibroblast-like
synoviocytes (RA-FLS), which are associated with the development
and progression of destructive joint inflammation, and are
resistant to receptor-mediated apoptosis at numerous levels,
alongside the alteration in mitochondrial pathway-mediated
apoptosis, another characteristic of RA (9,10).
In addition to apoptosis, autophagy is another important mechanism
that regulates cell survival and homeostasis. Numerous mechanisms
that contribute to reduced apoptosis have been extensively studied,
including autophagy, which is modulated by apoptosis (11,12).
RA-FLS are resistant to apoptosis, which is induced by apoptotic
stimuli; however, the role of apoptosis, as well as autophagy, in
the regulation of RA-FLS remains unclear. The present study aimed
to provide an overview regarding the participation of PADI4 in the
apoptosis of RA-FLS, and its role in the hypoxia-induced autophagy
of RA-FLS.
Materials and methods
Ethical approval
The present study was approved by the Ethics
Committee of Tongji University (Shanghai, China). Written informed
consent was obtained from all participants prior to the study.
Patients and tissue samples
All the specimens were obtained from patients
admitted to Shanghai East Hospital in China from February 2016 to
December 2016. Synovial tissues were obtained from 8 patients with
RA, including 2 males and 6 females (age range, 48–74 years; median
age, 58 years). In addition, synovial tissues were also obtained
from 6 patients with osteoarthritis (OA; 3 males and 3 females; age
range, 48–77 years; median age, 60 years), who underwent knee
arthroscopic or replacement surgery, were recruited to the present
study (13). The tissue samples
were immediately processed for immunohistochemical analysis. All
patients fulfilled the American College of Rheumatology criteria
for the diagnosis of RA and OA.
Animal model
A total of 18 inbred female rats (Sprague-Dawley;
age, 7–8 weeks) weighing ≥150 g were purchased from Shanghai SLAC
Laboratory Animal Co., Ltd., (Shanghai, China) and were allowed to
acclimate for ≥1 week prior to experimentation. The rats were
housed in an animal facility and were maintained at 25°C (humidity,
60–70%) under a 12 h light/dark cycle with ad libitum access
to food and water. Bovine type II collagen (CII; Chondrex, Inc.,
Redmond, WA, USA) was used to induce RA, which was dissolved in
0.05 mol/l acetic acid to a final concentration of 2.0 mg/ml, and
was vortexed overnight at 4°C. Subsequently, CII was emulsified
with complete Freund's adjuvant (Chondrex, Inc.) at a ratio of 1:1.
The rats were immunized with 300 µl CII emulsion by subcutaneous
injection into the tail root. On day 7, the rats received a
subcutaneous booster injection (300 µl) into the tail; the primary
injection site was avoided. After 28 days following the induction
of the RA model via CII, the rats were sacrificed. All procedures
that involved animals were performed in accordance with the
institutional animal welfare guidelines of Tongji University
(14). The rats were divided into
the following groups: i) Control group, in which RA was not induced
(n=6) and were treated with saline and an ii) RA group, in which RA
was induced via CII (n=12). Rats were examined three times per
week.
Cell lines and reagents
Synovial tissues were obtained from the rats; N-FLS
were obtained from the control group and RA-FLS were obtained from
the RA group. Synovial tissues were minced into pieces of 2 to 3 mm
in size and incubated with 1640 medium (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% FBS in a
humidified atmosphere containing 5% CO2, which was
changed every 3–5 days, and non-adherent tissue pieces were
carefully removed. FLS from synovial tissues in the rat model were
cultured in Dulbecco's modified Eagle's medium (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(FBS; Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C in a
humidified atmosphere containing 5% CO2. FLS were grown
over 4–6 passages. Subsequently, FLS were cultured in 1%
O2 for 48 h to induce autophagy and in 20% O2
for normal conditions.
Adenovirus production and transient
transfection
The short hairpin (sh)RNA sequences of PADI4 were
designed using Oligoengine 2.0 software (Oligoengine, Seattle, WA,
USA) and were verified by nucleotide BLAST searches (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE=BlastSearch).
The candidate sequence and the scrambled sequence with no
significant homology are listed in Table I. The shRNA sequence or coding
sequence (NM_012387.2) of PADI4 was cloned into pHBAd (Shanghai
GenecChem Co., Ltd., Shanghai, China) or GV314 adenovirus vectors
(Shanghai GeneChem Co., Ltd.) using BamHI/AgeI
(Shanghai GeneChem Co., Ltd.), respectively. FLS were seeded
(1×104 cells/well) in 96-well plates overnight. A total
of 1 µg green fluorescent protein (GFP)-microtubule-associated
protein light chain 3 (LC3) expressing plasmids, along with 1 µg
GV314-PADI4 (Ad-PADI4) or 1 µg pHBAd-shPADI4 (sh-PADI4) adenovirus
vectors, were transiently transfected into rat normal FLS (N-FLS)
or RA-FLS, respectively using the Fugene HD transfection reagent
(cat. no. 04709705001; Roche Diagnostics, Indianapolis, IN, USA),
according to the manufacturer's protocol. Cells were transfected
with GFP-LC3, along with Ad-PADI4 or sh-PADI4 adenovirus vectors,
for 8 h. At the end of each experiment, autophagy was detected by
counting the percentage of GFP-LC3-expressing FLS under a
fluorescence microscope (IX71; excitation wavelength was between
460–550 nm; Olympus Corporation, Tokyo, Japan). The cells
transfected with GFP-LC3 expressing plasmids along with blank GV314
(Ad-Vector) or pHBAd-scramble shRNA (sh-Vector) adenovirus vectors
were used as negative controls. A minimum of 200 cells were counted
in each sample. The experiment was conducted in triplicate.
 | Table I.Sequences of PADI4-short hairpin
RNA. |
Table I.
Sequences of PADI4-short hairpin
RNA.
| Gene | Sequence
(5′→3′) |
|---|
| PADI4 |
|
| Candidate
sequence |
CCAGAUUUUGGCUAUGUAACUTT |
| Scrambled
sequence |
GACAUUGAGAGAACAUAAUUCTT |
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Briefly, total RNA was extracted from synovial
tissues and FLS using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
First-strand cDNA was synthesized from 2 µg total RNA using an AMV
reverse transcriptase kit (Fermentas; Thermo Fisher Scientific,
Inc.) for 60 min at 37°C, 5 min at 85°C and 5 min at 4°C. The
primer sequences are listed in our previous study (13). qPCR was conducted using a
SYBR-Green PCR kit (Toyobo Life Science, Osaka, Japan) with the
7300 Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The PCR cycling conditions were as follows: 95°C
for 10 min, followed by 40 cycles at 95°C for 15 sec and 60°C for
45 sec, and a final extension step of 95°C for 15 sec, 60°C for 1
min, 95°C for 15 sec and 60°C for 15 sec. β-actin mRNA expression
was used an internal control for normalization. The average
Cq values for triplicate reactions were calculated, and
relative expression was determined with the comparative
Cq method (15) using
average Cq values.
Cell viability assay
An MTT assay was performed to analyze cell
viability. Cells were plated in 96-well plates at 5×103
cells/100 ml medium overnight prior to the experiment. The cell
viability in each well was examined using MTT colorimetric assay (5
mg/ml; cat. no. M2003; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). MTT solution was then added to each well and incubated
for 3 h at 37°C; 100 µl dimethyl sulfoxide was used to dilute the
formazan crystals. The optical density value of each sample was
measured at 490 nm using a plate reader. All determinations were
carried out in sextuplicate.
Cell apoptosis assay
Cell apoptosis was performed by using Annexin V
Apoptosis Detection kit APC (eBioscience; Thermo Fisher Scientific,
Inc.). FLS (2×105 cells/well) were cultured in 6-well
plates until they reached 70–80% confluence, after which the cells
were collected by trypsinization, washed twice with ice-cold
Annexin V binding buffer, and stained with 300 µl 1X binding buffer
containing 5 µl Annexin V and 5 µl propidium iodide (PI) for 30 min
at room temperature in the dark. Subsequently, 400 µl Annexin V
binding buffer was added and the cells were analyzed using flow
cytometry (BD Biosciences, Franklin Lakes, NJ, USA), according to
the manufacturer's protocol; ≥30,000-gated events were acquired
from each sample. Early stage apoptotic cells were stained with
fluorescein isothiocyanate (FITC) Annexin V, but not PI; whereas
late stage apoptotic cells and necrotic cells were stained
positively for FITC Annexin V and PI.
H&E, immunohistochemistry (IHC)
and immunofluorescence
Briefly, synovial tissues specimens were dehydrated
and embedded in paraffin; subsequently, 4-µm tissue sections were
cut using a Leica Biosystem Rotary Microtome (Leica Microsystem
Nussloch GmbH, Wetzlar, Germany). The sections of the synovial
tissues were stained with HE (Richard Allan Scientific Co.; Thermo
Fisher Scientific, Inc.). IHC was performed using the standard
streptavidin-biotin-peroxidase complex method. The synovial tissue
sections were incubated with affinity-purified anti-PADI4 (1:100;
cat. no. ab128086; Abcam, Cambridge, UK), anti-autophagy-related
gene 5 (ATG5; 1:100; cat. no. ab109490; Abcam), anti-LC3 (1:200;
cat. no. 3868) and anti-hypoxia-inducible factor-1α (HIF-1α)
antibodies (1:50; cat. no. 79233; Cell Signaling Technology, Inc.,
Danvers, MA, USA) for 1 h at room temperature, followed by
incubation with a biotin-labeled secondary antibody (1:2,000; cat.
no. A0192; Beyotime Institute of Biotechnology, Co., Ltd., Haimen,
China) for 1 h at room temperature. Finally, slides were developed
using 3,3-diaminobenzidine (Shanghai Long Island Biotec, Co., Ltd.,
China) solution, and were counterstained with hematoxylin (BASO,
Taipei, Taiwan). The primary antibody was omitted in control
experiments. The slides were observed under an identical
lightmicroscope (IX70; Olympus Corporation) at a magnification of
×200 or ×400. The specimens were evaluated by two independent
observers who were unaware of the clinical information.
For immunofluorescence, FLS were cytospun at 1,000 ×
g for 4 min at 25°C and fixed with 4% paraformaldehyde for 30 min
at 25°C. Following permeabilization by 0.5% Triton X-100 (Beijing
Solarbio Science & Technology Co., Ltd., Beijing, China) for 10
min and blocking by 1% bovine serum albumin (Beijing Solarbio
Science & Technology Co., Ltd.) for 1 h at 25°C. Cells were
stained sequentially with the PADI4 (1:100; cat. no. ab128086;
Abcam), LC3 (1:200; cat. no. 3868; Cell Signaling Technology, Inc.)
or Beclin1 (1:200; cat. no. Ab207612; Abcam) antibody overnight at
4°C, after blocking with 2% bovine serum albumin (Sigma-Aldrich;
Merck KGaA) in PBS for 1 h. Cells were washed three times with PBS
containing 0.1% Tween-20 and were incubated for 1 h with a
corresponding FITC-conjugated secondary antibody (1:500; cat. no.
A0568; Beyotime Institute of Biotechnology, Co., Ltd.) at room
temperature. For the identification of cells with nuclear changes
typical of apoptosis, cells were stained with 0.1 mg/ml DAPI for 5
min. The samples were examined under a fluorescence microscope
(IX70; Olympus Corporation).
Statistical analysis
All data were generated without knowledge of the
clinical status of the samples, and were analyzed using GraphPad
Prism software, version 5 (GraphPad Software, Inc., La Jolla, CA,
USA). Student's t-test was performed for the analysis of paired and
unpaired samples. One-way analysis of variance and a multiple
comparisons test (Least Significant Difference test) among three
groups were performed. P<0.05 was considered to indicate a
statistically significant difference.
Results
PADI4 promotes the proliferation of
FLS in human RA via hypoxia-induced autophagy
In order to investigate the effects of PADI4 on the
growth of human FLS in RA, immunostaining was performed in synovial
tissues obtained from patients with RA and OA. As presented in
Fig. 1A, PADI4 was overexpressed
in RA-FLS, and the proliferation of FLS increased in RA-FLS
compared with N-FLS. These results indicated that PADI4 may affect
the growth of RA-FLS in patients with RA, compared with in patients
with OA.
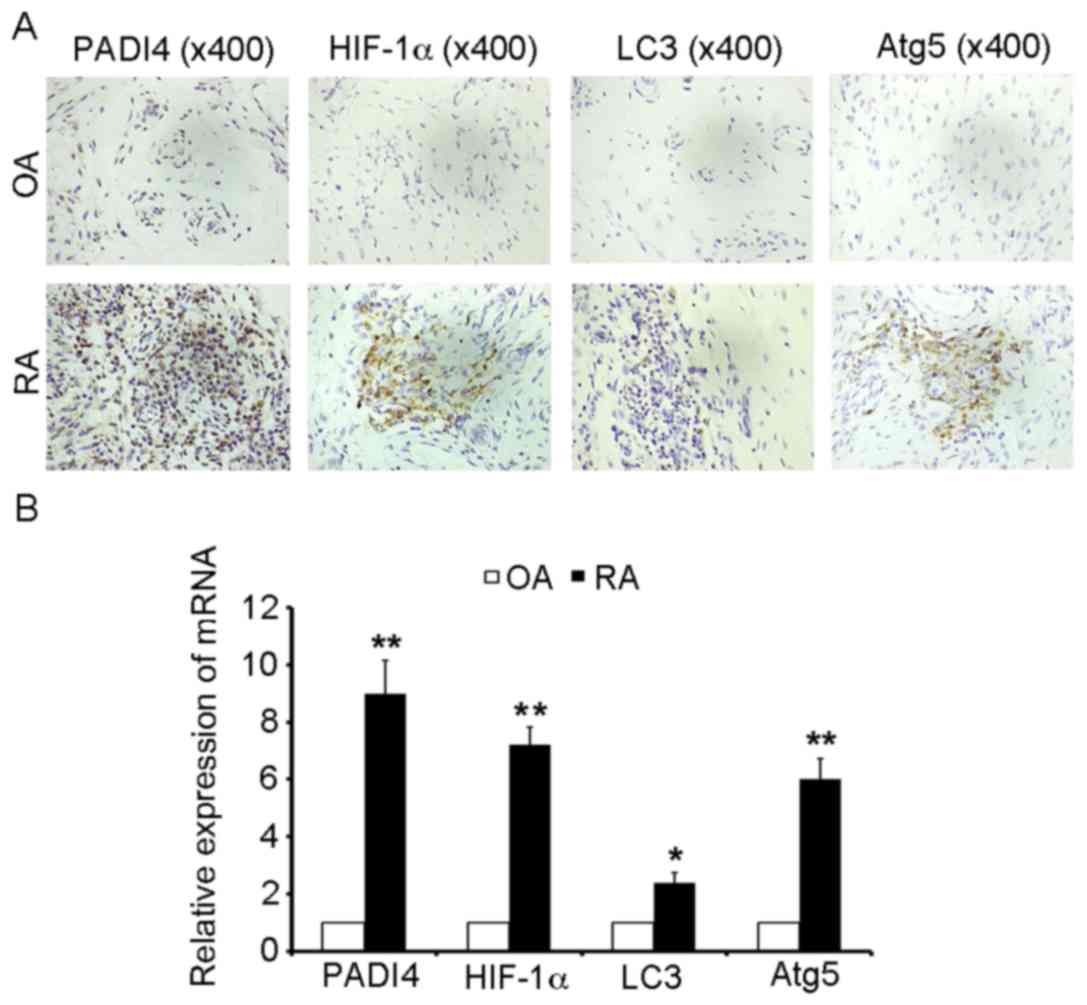 | Figure 1.PADI4 and autophagy marker expression
in arthritic synovial tissues from patients with RA and OA. (A)
Representative images of immunohistochemical staining of PADI4,
HIF-1α, LC3 and Atg5 in patients with RA and OA (magnification,
×400). (B) Relative mRNA expression levels of PADI4, HIF-1α, LC3
and Atg5 in arthritic synovial tissues from patients with RA and
OA. Data represent three independent experiments with presented as
mean ± standard deviation. *P<0.01; **P<0.001. Atg5,
autophagy-related gene 5; HIF-1α, hypoxia-inducible factor-1α; LC3,
microtubule-associated protein light chain 3; OA, osteoarthritis;
PADI4, peptidyl arginine deiminase type IV; RA, rheumatoid
arthritis. |
Since apoptosis is often associated with autophagy,
the present study aimed to determine whether PADI4 could induce
impaired apoptosis of RA-FLS through hypoxia-induced autophagy.
HIF-1α is an important inducible factor responsible for cellular
adaptation to low oxygen tension, and is able to induce autophagy;
therefore, the present study determined whether HIF-1α was involved
in hypoxia-induced autophagy. As shown in Fig. 1A, HIF-1α was overexpressed in
RA-FLS compared with in OA. In order to analyze the mechanism
underlying the autophagic process, IHC was used to monitor and
evaluate autophagic activity. LC3 and Atg5 are specific markers of
autophagy initiation; therefore, LC3 and Atg5 expression were
characterized in resected synovial specimens obtained from patients
with RA. The results revealed that LC3 and Atg5 were overexpressed
in the RA samples, compared with in the OA tissues (Fig. 1A). Furthermore, it was demonstrated
that the mRNA expression levels of PADI4 and markers of autophagy
(HIF-1α, LC3 and Atg5) were increased in patients with RA compared
with in patients with OA (Fig.
1B). Together, these results suggested that PADI4 may be
involved in the response of FLS to hypoxia-induced autophagy.
PADI4 promotes the proliferation of
FLS in RA in vivo
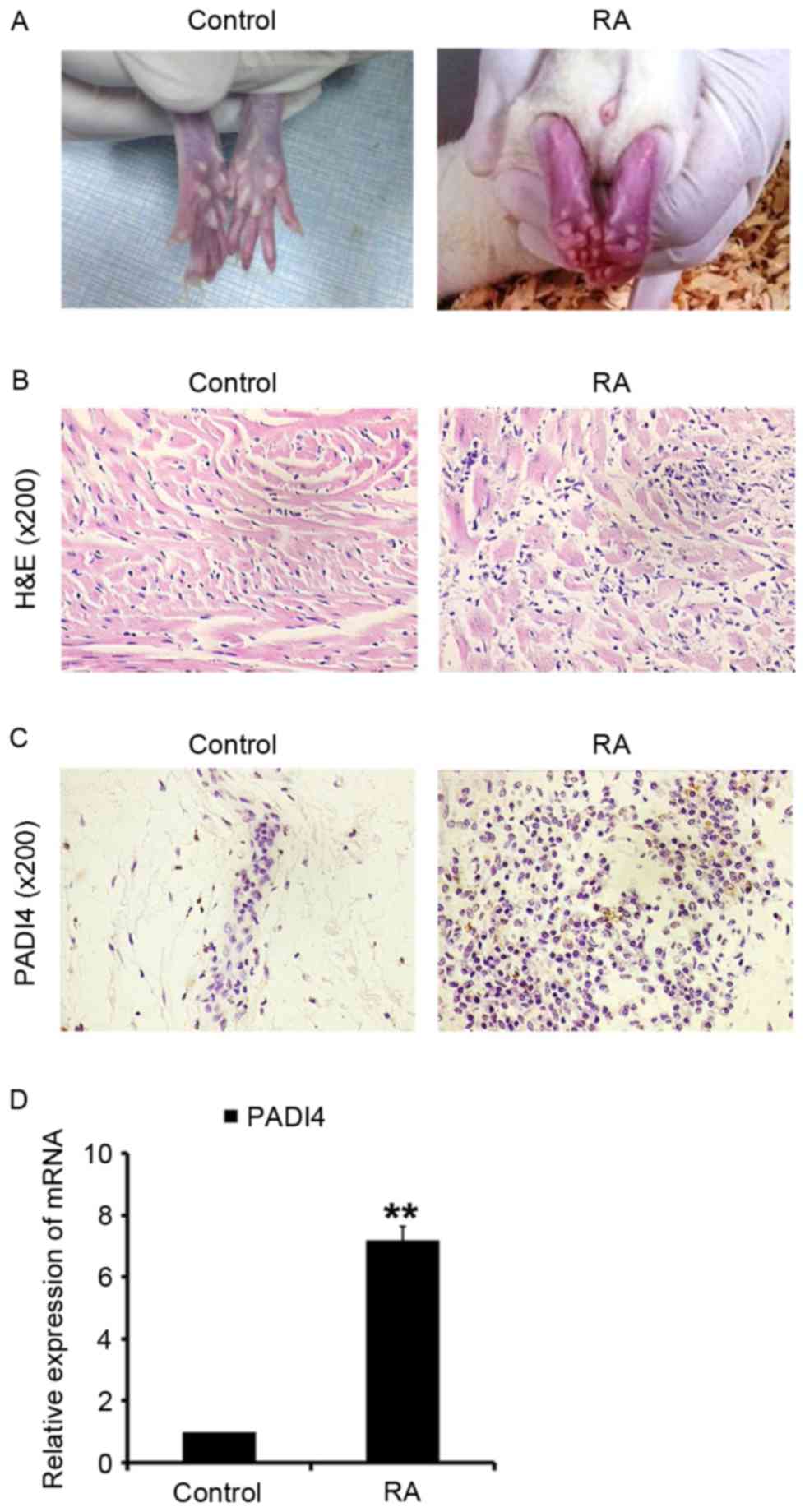
Inflammation of the ankles was quantified daily by
measuring ankle diameter using the standard caliper method from
disease onset to day 28 after the initiation of arthritis (Fig. 2A). At the end of 4 weeks, the
animals were sacrificed; the left and right femurs were dissected
from the soft tissue, and fixed in 10% natural buffered formalin
for analysis (Fig. 2B). The
effects of PADI4 in vivo were evaluated after the initiation
of arthritis. PADI4 was overexpressed in synovial tissues in the RA
group rats (Fig. 2C and D). In
addition, it was observed that RA-FLS exhibited increased
proliferation in the RA group (Fig. 3A
and B). These results suggested that PADI4 may have a similar
effect on the growth of RA-FLS in rats, similar to in patients with
RA.
In vitro PADI4 promotes the
proliferation of FLS through hypoxia-induced autophagy
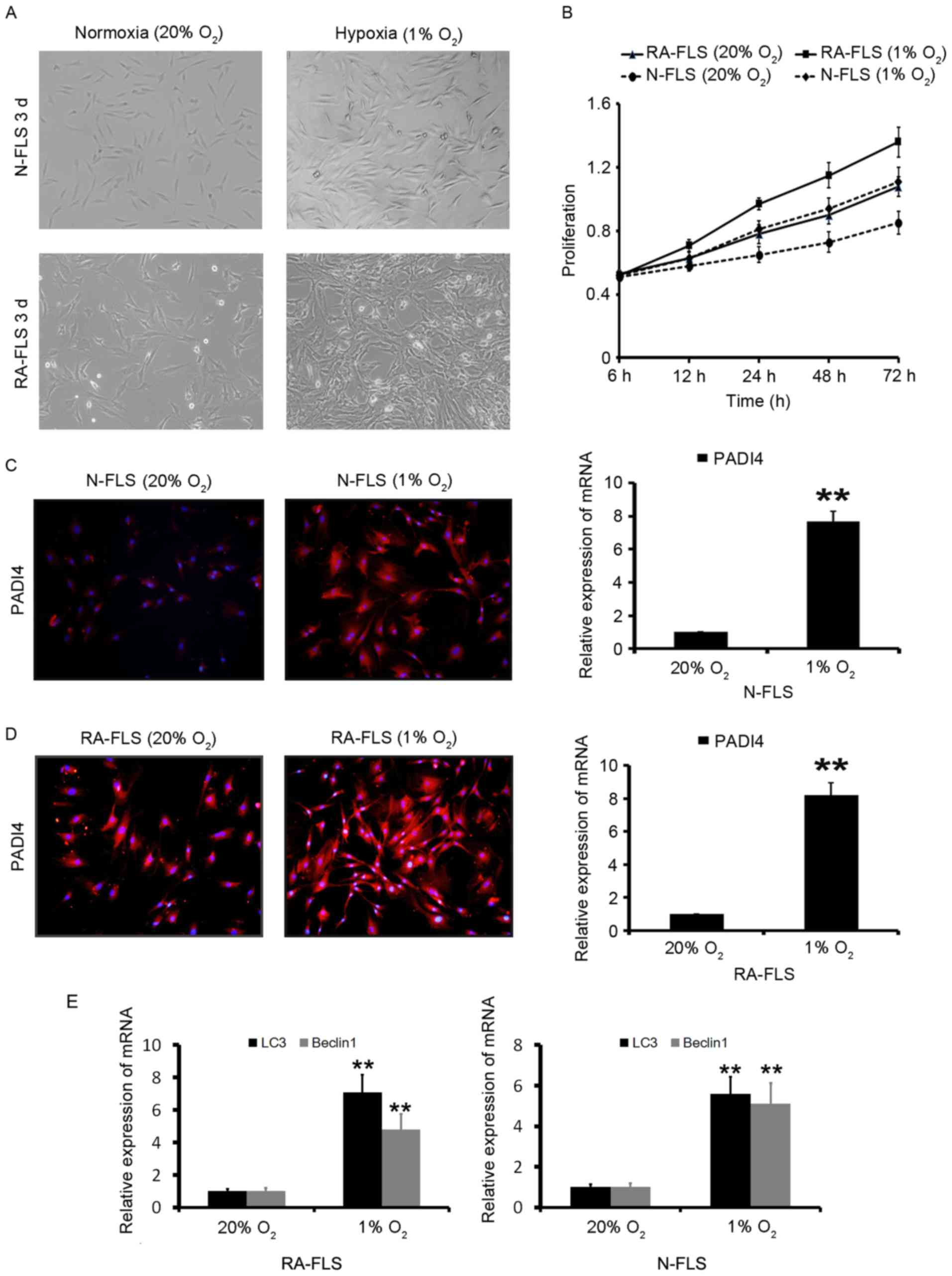
FLS in 1% O2 were observed to markedly
proliferate in a time-dependent manner (Fig. 3A and B). These results indicated
that hypoxia significantly increased RA-FLS proliferation compared
with N-FLS proliferation. The effects of 20 and 1% O2 on
PADI4 expression in vitro were evaluated. As shown in
Fig. 3C and D, PADI4 was
overexpressed in FLS under hypoxia compared with in cells under
normoxia. These results suggested that PADI4 may promote the
proliferation of RA-FLS through hypoxia.
Hypoxia-induced autophagy in RA-FLS was subsequently
analyzed. The results demonstrated that autophagy was significantly
increased in the first 24 h of hypoxia, and was further increased
after 48 h of hypoxia (data not shown). The induction of autophagy
was identified by two well-established measurements of autophagy;
the enhancement of Beclin1, a component of the class III
phosphatidylinositol 3-kinase complex essential for autophagosome
formation, and LC3 (16) as
presented in Fig. 3E. In order to
investigate the effects of PADI4 on hypoxia-induced autophagy in
FLS, N-FLS transfected with Ad-PADI4 and RA-FLS transfected with
PADI4 shRNA were cultured under hypoxia for 5 days.
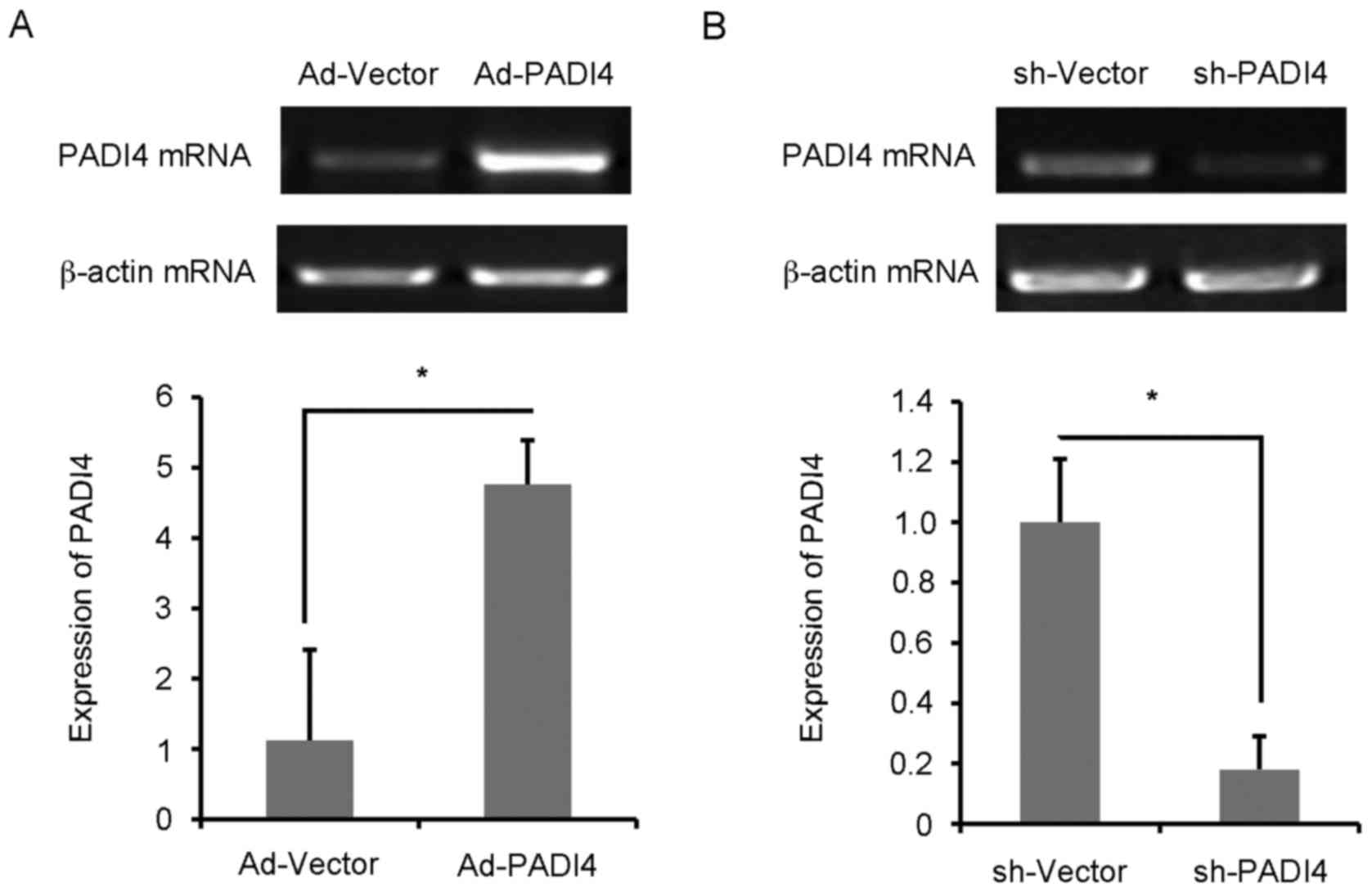
FLS with Ad-PADI4 or sh-PADI4 transfection were
examined by RT-qPCR (Fig. 4A and
B). Ad-PADI4 transfection significantly increased the mRNA
expression levels of PADI4 in N-FLS and sh-PADI4 transfection
significantly decreased the mRNA expression levels of PADI4 in
RA-FLS. FLS with GFP-LC3 plasmid transfection were examined under
hypoxia using a fluorescent microscope (Fig. 5A). RA-FLS with PADI4 shRNA
exhibited a diffuse expression pattern of GFP-LC3, whereas N-FLS
with Ad-PADI4 revealed a punctate pattern of GFP-LC3, indicating
the formation of autophagosomes. Furthermore, a large number of
N-FLS transfected with Ad-PADI4 exhibited increased LC3 and Beclin1
expression, compared with RA-FLS transfected with PADI4 shRNA
(Fig. 5B-E). These data suggested
that ectopic expression of PADI4, induced by transfection of N-FLS
with PADI4 expression plasmid, may be involved in hypoxia-induced
autophagy. Consistent with these results, PADI4 inactivation
partially inhibited hypoxia-induced autophagy.
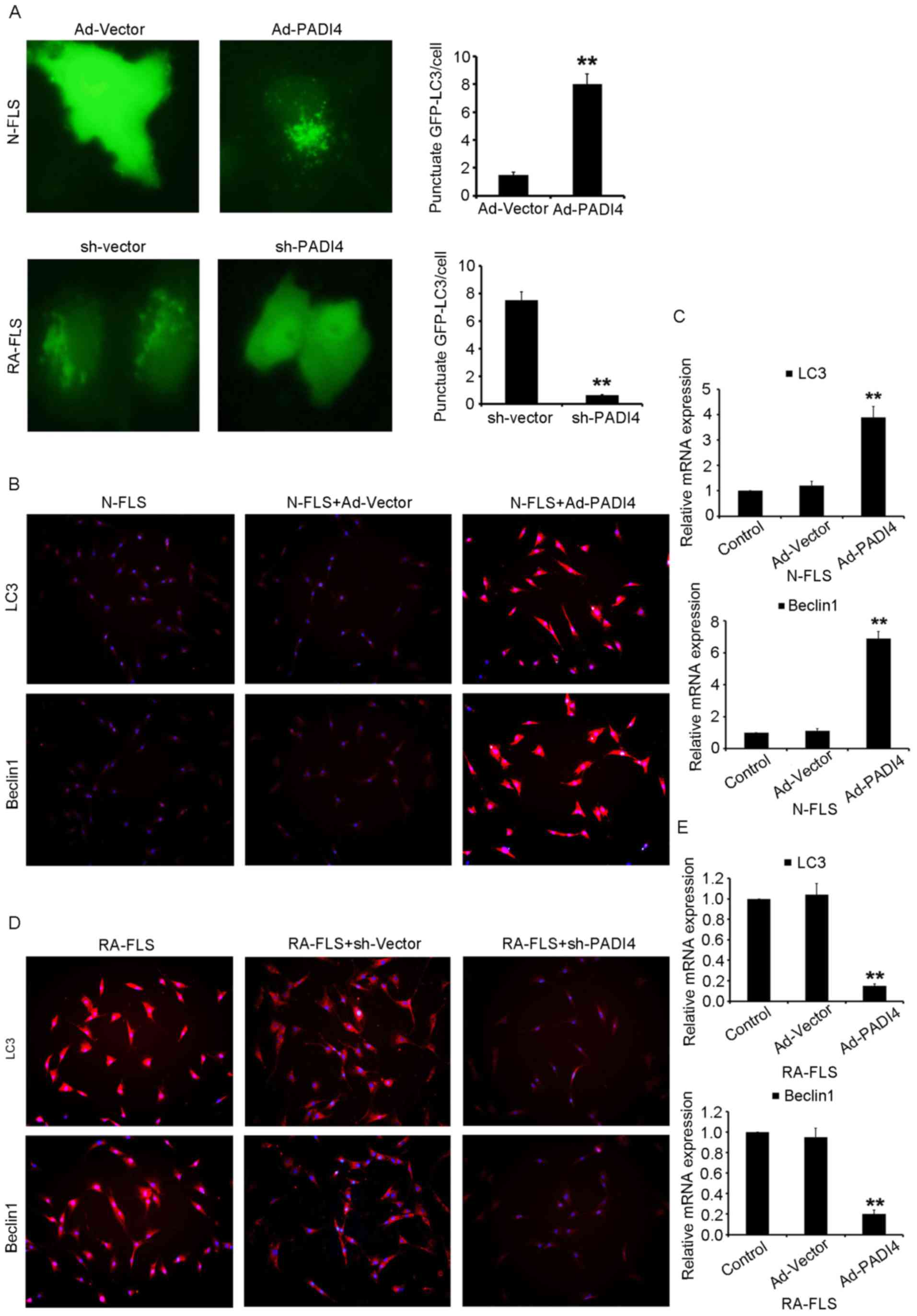 | Figure 5.Inactivation of PADI4 partially
inhibits hypoxia-induced autophagy. (A) Representative images
(magnification, ×1,000) of GFP-LC3 expression patterns in FLS
transfected with Ad-PADI4 orsh-PADI4 under hypoxia (1%
O2) for 3 days. FLS transfected with GFP-LC3 plasmid
were detected. Expression of LC3 and Beclin1 detected using (B)
immunofluorescence (magnification, ×400) and (C) qPCR in N-FLS
transfected with Ad-PADI4 under hypoxia for 3 days. Expression of
LC3 and Beclin1 detected by (D) immunofluorescence (magnification,
×400) and (E) qPCR in RA-FLS transfected with sh-PADI4 under
hypoxia for 3 days. Data represent three independent experiments
with presented as mean ± standard deviation. **P<0.001. Ad,
adenovirus; FLS, fibroblast-like synoviocytes; GFP, green
fluorescent protein; LC3, microtubule-associated light chain 3;
PADI4, peptidyl arginine deiminase type IV; RA, rheumatoid
arthritis; sh, short hairpin RNA. |
PADI4 inactivation induces RA-FLS
apoptosis
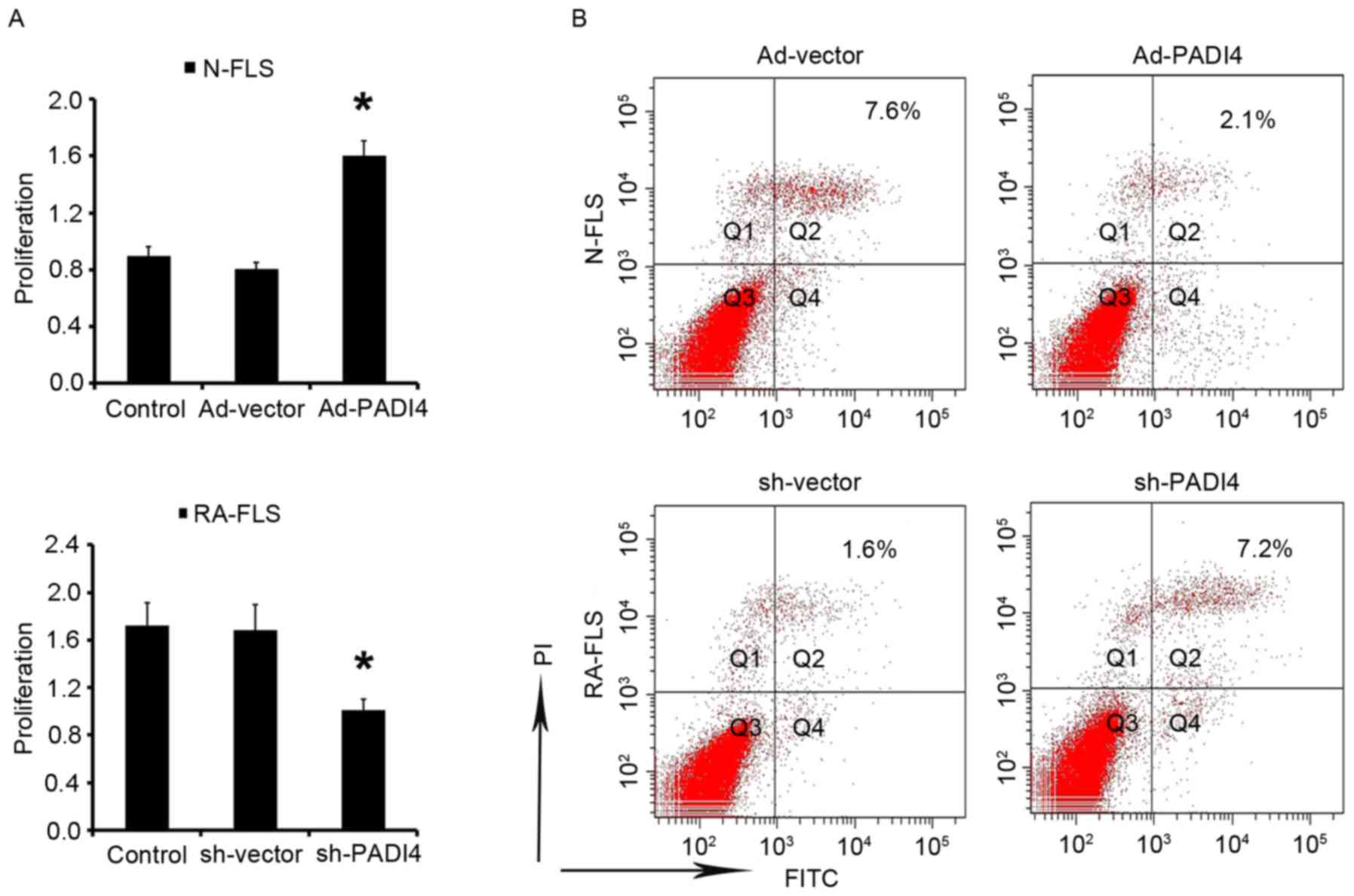
The results of the present study indicated that FLS
proliferation is increased under hypoxia. Subsequently, the study
aimed to determine the effects of PADI4 on FLS under hypoxia. After
being cultured under hypoxia for 3 days N-FLS transfected with
Ad-PADI4 exhibited significant proliferation compared with N-FLS
transfected with Ad-vector; whereas the growth of RA-FLS with PADI4
shRNA was suppressed compared with RA-FLS transfected with
sh-vector (Fig. 6A). Apoptosis of
FLS was analyzed by flow cytometry. The results demonstrated that
apoptosis was enhanced in RA-FLS transfected with PADI4 shRNA, thus
suggesting that PADI4 is an important factor in the inhibition of
RA-FLS apoptotic death (Fig. 6B).
These data suggested that PADI4 inactivation may be involved in
hypoxia-induced autophagy, and may induce RA-FLS apoptosis.
Discussion
RA is a chronic and complex autoimmune disease with
a complex etiology, which leads to generalized bone loss, cartilage
erosion and increased fracture risk. Although the causes of RA
remain unclear, it has been reported to be attributed to genetics,
autoimmunity and lifestyle-associated factors (17). RA-FLS serve a crucial role in
producing cytokines that are responsible for inflammation, and
proteases that may result in cartilage destruction. RA-FLS are
associated with the initiation and perpetuation of RA, in which
impaired apoptosis of RA-FLS is pivotal; however, the molecular
mechanism underlying reduced apoptosis remains to be elucidated
(10).
The PADI4 gene, which encodes the PADI4 enzyme, has
an important role in protein citrullination, which is a key event
underlying the pathogenesis of RA. PADI4 is involved in gene
regulation through the citrullination of histones, which
contributes to the generation of ACPA specific substrates, and is
itself a target of autoantibodies in RA (2). PADI4 polymorphisms represent a
significant risk factor for RA not only in Asian populations, but
also in populations of European descent (18,19).
Furthermore, increased transcription of PADI4 has been detected in
the synovial membrane of RA (20).
Our previous results indicated that citrullinated vimentin
significantly increased the expression of PADI4 in cultured RA-FLS
(21). The present study
demonstrated that PADI4 protein expression was elevated in human
RA-FLS. Subsequently, the study aimed to determine whether PADI4
could be associated with the impaired apoptosis of RA-FLS.
Hypoxia is a key regulator of angiogenesis and
inflammation in RA (22). HIF has
been reported to induce adaptive responses to hypoxic stress by
activating a large number of genes that are responsible for oxygen
delivery, angiogenesis, cell proliferation, cell differentiation
and metabolism (23). The present
study revealed that HIF-1α protein expression was elevated in human
RA-FLS, thus indicating that hypoxia may be a factor promoting the
proliferation of RA-FLS. In addition, it has been reported that
synovial tissue was hypoxic in patients with RA compared with in
noninflamed synovium from patients without RA (24). Autophagy has been reported to be
induced by hypoxia (25). Hypoxic
stress, which is mediated through HIF-1α, is a strong signal that
initiates the autophagic process (26).
HIF-1α has been reported to serve a critical role in
the regulation of hypoxia-induced angiogenesis (27). In addition, autophagy modulates
apoptosis; it has previously been demonstrated that autophagy
inhibits apoptosis through the degradation of proapoptotic
proteins, including caspases (28). In the present study, PADI4
overexpression in RA rat model may have promoted the proliferation
of RA-FLS.
It has been hypothesized that inhibition of
autophagy promotes apoptosis in cancer cells with intact apoptotic
signaling pathways (29).
Furthermore, enhanced autophagy was associated with elevated levels
of HIF-1α. In the present study, IHC revealed that LC3 and Atg5
were overexpressed in resected synovial specimens obtained from
patients with RA compared with in tissues obtained from patients
with OA. These findings indicated that hypoxia-induced autophagy
may serve an important role in synovial tissues.
The number of RA-FLS that originated from the RA rat
model markedly increased in the presence of 1% O2, but
not in normal oxygen conditions. In addition, numerous stressors,
including severe hypoxia or oxidative stress could induce
activation of autophagy in synovial tissues obtained from patients
with RA (30). In the present
study, after being cultured under hypoxic conditions, a marked
increase in PADI4 expression was observed in cultured RA-FLS.
Furthermore, a significant increase in Beclin1 and LC3 expression
was detected in cells with ectopic overexpression of PADI4.
Conversely, decreased Beclin1 and LC3 expression was detected in
cells transfected with PADI4 shRNA. Increased apoptosis of RA-FLS
was also observed in response tosh-PADI4 transfection, thus
indicating that suppression of RA-FLS may be associated with PADI4
inhibition. The in vivo and in vitro results of the
present study suggested that PADI4 may be closely associated with
hypoxia-induced autophagy.
In conclusion, these results revealed that
inactivation of hypoxia-induced autophagy via the knockdown of
PADI4 may contribute to increased apoptosis of RA-FLS. Therefore,
further studies that investigate the role of PADI4 in
hypoxia-induced autophagy may provide novel information regarding
the mechanism underlying the impaired apoptosis of RA-FLS. The
present study also proposed a novel mechanism underlying the close
association between hypoxia, autophagy and inflammation in RA, and
supported the concept that autophagic inhibition may be of
therapeutic benefit in RA; however, the present study had some
shortcomings. The number of patients enrolled was insufficient and
the effects of PADI4 on the growth of RA-FLS were determined using
immunostaining only. Therefore, the findings of the present study
should be further investigated in future studies.
Acknowledgements
The present study was supported by the Natural
Science Foundation of China (grant nos. 81373203 and 81372212), the
Shanghai Municipal Science and Technology Commission (grant no.
12XD1404300) and the Natural Science Foundation of Jiangsu (grant
nos. BK2011251 and BL2013012).
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
RA
|
rheumatoid arthritis
|
|
FLS
|
fibroblast-like synoviocytes
|
|
PADI4
|
peptidyl arginine deiminase type
IV
|
|
OA
|
osteoarthritis
|
|
ACPA
|
anti-cyclic citrullinated peptides
antibody
|
|
IHC
|
immunohistochemistry
|
|
LC3
|
microtubule-associated protein light
chain 3
|
|
Atg5
|
autophagy-related gene 5
|
|
HIF-1α
|
hypoxia-inducible factor-1α
|
References
|
1
|
Ikari K, Yano K, Yoshida S, Taniguchi A,
Yamanaka H and Momohara S: Response to ‘Peptidyl arginine deiminase
type IV (PADI4) haplotypes interact with shared epitope regardless
of anti-cyclic citrullinated peptide antibody or erosive joint
status in rheumatoid arthritis: A case control study’. Arthritis
Res Ther. 16:4222014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dong S, Zhang Z and Takahara H:
Estrogen-enhanced peptidylarginine deiminase type IV gene (PADI4)
expression in MCF-7 cells is mediated by estrogen
receptor-alpha-promoted transfactors activator protein-1, nuclear
factor-Y, and Sp1. Mol Endocrinol. 21:1617–1629. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yamada R, Suzuki A, Chang X and Yamamoto
K: Citrullinated proteins in rheumatoid arthritis. Front Biosci.
10:54–64. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suzuki A, Yamada R, Chang X, Tokuhiro S,
Sawada T, Suzuki M, Nagasaki M, Nakayama-Hamada M, Kawaida R, Ono
M, et al: Functional haplotypes of PADI4, encoding citrullinating
enzyme peptidylarginine deiminase 4, are associated with rheumatoid
arthritis. Nat Genet. 34:395–402. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zavala-Cerna MG, Gonzalez-Montoya NG, Nava
A, Gamez-Nava JI, Moran-Moguel MC, Rosales-Gomez RC,
Gutierrez-Rubio SA, Sanchez-Corona J, Gonzalez-Lopez L,
Davalos-Rodriguez IP and Salazar-Paramo M: PADI4 haplotypes in
association with RA Mexican patients, a new prospect for antigen
modulation. Clin Dev Immunol. 2013:3836812013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bang SY, Han TU, Choi CB, Sung YK, Bae SC
and Kang C: Peptidyl arginine deiminase type IV (PADI4) haplotypes
interact with shared epitope regardless of anti-cyclic
citrullinated peptide antibody or erosive joint status in
rheumatoid arthritis: A case control study. Arthritis Res Ther.
12:R1152010. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suzuki T, Ikari K, Yano K, Inoue E, Toyama
Y, Taniguchi A, Yamanaka H and Momohara S: PADI4 and HLA-DRB1 are
genetic risks for radiographic progression in RA patients,
independent of ACPA status: Results from the IORRA cohort study.
PLoS One. 8:e610452013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartok B and Firestein GS: Fibroblast-like
synoviocytes: Key effector cells in rheumatoid arthritis. Immunol
Rev. 233:233–255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bottini N and Firestein GS: Duality of
fibroblast-like synoviocytes in RA: Passive responders and
imprinted aggressors. Nat Rev Rheumatol. 9:24–33. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li H and Wan A: Apoptosis of rheumatoid
arthritis fibroblast-like synoviocytes: Possible roles of nitric
oxide and the thioredoxin 1. Mediators Inflamm. 2013:9534622013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gu X, Gu B, Lv X, Yu Z, Wang R, Zhou X,
Qiao W, Mao Z, Zuo G, Li Q, et al: 1, 25-dihydroxy-vitamin D3 with
tumor necrosis factor-alpha protects against rheumatoid arthritis
by promoting p53 acetylation-mediated apoptosis via Sirt1 in
synoviocytes. Cell Death Dis. 7:e24232016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang L, Zhang HY, Gao B, Shi J, Huang Q,
Han YH, Hu YQ, Lu WG, Zhao ZJ, Liu BH, et al: Tetramethylpyrazine
protects against glucocorticoid-induced apoptosis by promoting
autophagy in mesenchymal stem cells and improves bone mass in
glucocorticoid-induced osteoporosis rats. Stem Cells Dev.
26:419–430. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fan L, Wang Q, Liu R, Zong M, He D, Zhang
H, Ding Y and Ma J: Citrullinated fibronectin inhibits apoptosis
and promotes the secretion of pro-inflammatory cytokines in
fibroblast-like synoviocytes in rheumatoid arthritis. Arthritis Res
Ther. 14:R2662012. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin TN, He YY, Wu G, Khan M and Hsu CY:
Effect of brain edema on infarct volume in a focal cerebral
ischemia model in rats. Stroke. 24:117–121. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Matsunaga K, Saitoh T, Tabata K, Omori H,
Satoh T, Kurotori N, Maejima I, Shirahama-Noda K, Ichimura T, Isobe
T, et al: Two Beclin 1-binding proteins, Atg14L and Rubicon,
reciprocally regulate autophagy at different stages. Nat Cell Biol.
11:385–396. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kremer JM: Rheumatoid arthritis: New EULAR
guidelines for RA: A job well done. Nat Rev Rheumatol. 10:6–8.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hou S, Gao GP, Zhang XJ, Sun L, Peng WJ,
Wang HF, Ge XJ, Huang W and Sun YH: Erratum to: PADI4 polymorphisms
and susceptibility to rheumatoid arthritis: A meta-analysis. Mod
Rheumatol. 23:612013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Iwamoto T, Ikari K, Nakamura T, Kuwahara
M, Toyama Y, Tomatsu T, Momohara S and Kamatani N: Association
between PADI4 and rheumatoid arthritis: A meta-analysis.
Rheumatology (Oxford). 45:804–807. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chang X, Zhao Y, Sun S, Zhang Y and Zhu Y:
The expression of PADI4 in synovium of rheumatoid arthritis.
Rheumatol Int. 29:1411–1416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Reyes-Castillo Z, Palafox-Sánchez CA,
Parra-Rojas I, Martínez-Bonilla GE, del Toro-Arreola S,
Ramírez-Dueñas MG, Ocampo-Bermudes G and Muñoz-Valle JF:
Comparative analysis of autoantibodies targeting peptidylarginine
deiminase type 4, mutated citrullinated vimentin and cyclic
citrullinated peptides in rheumatoid arthritis: Associations with
cytokine profiles, clinical and genetic features. Clin Exp Immunol.
182:119–131. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Konisti S, Kiriakidis S and Paleolog EM:
Hypoxia-a key regulator of angiogenesis and inflammation in
rheumatoid arthritis. Nat Rev Rheumatol. 8:153–162. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brouwer E, Gouw AS, Posthumus MD, van
Leeuwen MA, Boerboom AL, Bijzet J, Bos R, Limburg PC, Kallenberg CG
and Westra J: Hypoxia inducible factor-1-alpha (HIF-1alpha) is
related to both angiogenesis and inflammation in rheumatoid
arthritis. Clin Exp Rheumatol. 27:945–951. 2009.PubMed/NCBI
|
|
24
|
Muz B, Khan MN, Kiriakidis S and Paleolog
EM: Hypoxia. The role of hypoxia and HIF-dependent signalling
events in rheumatoid arthritis. Arthritis Res Ther. 11:2012009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marin JJG, Lozano E and Perez MJ: Lack of
mitochondrial DNA impairs chemical hypoxia-induced autophagy in
liver tumor cells through ROS-AMPK-ULK1 signaling dysregulation
independently of HIF-1α. Free Radic Biol Med. 101:71–84. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang Z, Yang M, Wang Y, Wang L, Jin Z,
Ding L, Zhang L, Zhang L, Jiang W, Gao G, et al: Autophagy
regulates the apoptosis of bone marrow-derived mesenchymal stem
cells under hypoxic condition via AMP-activated protein
kinase/mammalian target of rapamycin pathway. Cell Biol Int.
40:671–685. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park SY, Jang WJ, Yi EY, Jang JY, Jung Y,
Jeong JW and Kim YJ: Melatonin suppresses tumor angiogenesis by
inhibiting HIF-1alpha stabilization under hypoxia. J Pineal Res.
48:178–184. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Feng LX, Sun P, Mi T, Liu M, Liu W, Yao S,
Cao YM, Yu XL, Wu WY, Jiang BH, et al: Agglutinin isolated from
Arisema heterophyllum Blume induces apoptosis and autophagy in A549
cells through inhibiting PI3K/Akt pathway and inducing ER stress.
Chin J Nat Med. 14:856–864. 2016.PubMed/NCBI
|
|
29
|
Ouyang L, Shi Z, Zhao S, Wang FT, Zhou TT,
Liu B and Bao JK: Programmed cell death pathways in cancer: A
review of apoptosis, autophagy and programmed necrosis. Cell
Prolif. 45:487–498. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kato M, Ospelt C, Gay RE, Gay S and Klein
K: Dual role of autophagy in stress-induced cell death in
rheumatoid arthritis synovial fibroblasts. Arthritis Rheumatol.
66:40–48. 2014. View Article : Google Scholar : PubMed/NCBI
|