Introduction
Stroke is prevalent worldwide, and is among the most
frequent causes of adult disability and neurological-associated
mortality. Stroke thus affects the quality of life of the patients
and their families (1). The
majority of strokes (88%) result from a blockage of blood vessels
in the brain, which is known as ischemic stroke; many ischemic
strokes occur in the middle cerebral artery (MCA), which has been
the focus of previous animal stroke models (2). Thus, the MCA occlusion (MCAO) model
results in a cessation of blood flow and subsequent brain
infarction in the MCA territory (3); however, the causal mechanisms of
stroke require further investigation.
Toll-like receptor 4 (TLR4) expression levels are
increased in response to ischemia within MCAO mice models (4). In addition, neurological deficits
were significantly decreased in mice deficient in TLR4 when
compared with controls (5);
however, the mechanism remains unclear. TLRs are widely expressed
in innate immune cells, including macrophages and microglia in the
central nervous system, which serve a critical role in early innate
immunity. Some TLRs, including TLR2, TLR4 and TLR8, are present in
microglia, astrocytes, oligodendrocytes and neurons in the brain
(6–8). Damaged neuronal structure and
deficits in neuroregeneration are common characteristics of
post-ischemic injury in stroke. Collapsin response mediator protein
2 (CRMP2) is known to be a neuronal phosphoprotein that regulates
microtubule assembly and is involved in anterograde vesicular
transport for growth-associated molecules along neuronal
microtubules (9). Phosphorylated
CRMP2 (p-CRMP2) is unable to bind with microtubules, which in turn
inhibits microtubule polymerization and stabilization, thereby
inhibiting axonal elongation in neurons (10). Therefore, the phosphorylation of
CRMP2 serves a major role in neuroregeneration post-ischemic injury
and stroke (11). TLR4 has been
demonstrated to be involved in neural stem cell proliferation and
serves an important role in neurogenesis (12). Thus, p-CRMP2 may be a candidate for
TLR4 regulation. Rho kinase is the enzyme responsible for the
phosphorylation of proteins, which participates in microtubule
assembly and may serve an important role in the TLR4 regulation of
p-CRMP2 (13,14); however, the association between
TLR4 and p-CRMP2 in stroke conditions of an MCAO ischemic model
remains unknown.
In the present study, the mechanism of stroke in a
model of TLR4 regulation was investigated. To the best of our
knowledge, the present study is the first to report that TLR4 may
promote the phosphorylation of CRMP2 within MCAO rat models, and to
demonstrate that the effects of phosphorylation may be mediated via
the activation of Rho-kinase.
Materials and methods
Reagents and antibodies
The Rho-associated protein kinase 2 (ROCK-II)
inhibitor, Y-27632 (cat. no. 555550; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany), the TLR4 specific agonist lipopolysaccharide
(LPS; cat. no. 437628; EMD Millipore, Billerica, MA, USA), CRMP
monoclonal antibody (cat. no. ab129082; Abcam, Cambridge, UK)
anti-p-CRMP2 polyclonal antibody (cat. no. 9397; Cell Signaling
Technology, Inc., Danvers, MA, USA), anti-Rho kinase antibody (cat.
no. 9029; Cell Signaling Technology, Inc.), TLR4 monoclonal
antibody (cat. no. sc-293072; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA), TLR4-neutralizing antibody (cat. no. 16-9917;
eBioscience; Thermo Fisher Scientific, Inc., Waltham, MA, USA), and
β-actin (cat. no. sc-47778; Santa Cruz Biotechnology, Inc.) were
used.
Animals
A total of 95 adult male Sprague-Dawley rats
weighing 240–280 g (10–12 weeks old) were randomly divided into 8
groups: The normal, sham, MCAO/reperfusion, saline, LPS,
TLR4-neutralizing antibody, LPS+Y-27632 and Y-27632 groups. The
animals were housed in an animal center at a constant temperature
of 22±2°C, a relative humidity of 50±10% and a 12-h light/dark
cycle. They were allowed free access to food and water. The present
study was approved by the Ethics Committee of the Department of
Forensic Medicine, Shantou University (Shantou, Guangdong, China).
All experiments were conducted in compliance with the National
Institute of Health's Guidelines for the Care and Use of Laboratory
Animals (15).
MCAO/reperfusion
The rats in the MCAO/reperfusion, saline, LPS,
LPS+TLR4-neutralizing, TLR4-neutralizing antibody (n=10 each
group), LPS+Y-27632 and Y-27632 groups (n=5 each group) were
subjected to MCAO and reperfusion models according to previous
studies (16). Briefly, rats were
anesthetized with 4% chloral hydrate intraperitoneally (300 mg/kg),
and MCAO was conducted using an intraluminal filament introduced
via the common carotid artery (CCA). A midline surgical incision
was made to expose the right CCA, external carotid artery and
internal carotid artery (ICA). A 40-mm-long monofilament nylon
suture (~0.26 mm in diameter) was inserted into the right CCA lumen
and gently advanced into the ICA to ~18 mm. The nylon sutures were
slowly removed from the artery 2 h later. The animal's body
temperature was continually monitored and maintained at 37°C with a
homeothermic blanket throughout the surgical procedure. The animals
in the sham group (n=10) were treated similarly, with the exception
that the filament was not advanced to the origin of the MCA. No
treatment performed on the normal group. Neurological function
deficits were evaluated following reperfusion when the rats
regained consciousness according to the Longa method (17) to evaluate the success of MCAO
surgery. Rats without neurological deficits 2 h following the
induction of stroke were excluded from this study.
Intracerebroventricular injection
The intracerebroventricular injection was performed
to the saline (control), LPS, LPS+Neutral, Neutral, Y-27632 and
LPS+Y-27632 groups (n=5 in each group) according to a previous
study (18). The rats were
anesthetized and placed in a stereotaxic frame with a mouse head
holder. A stainless-steel cannula was implanted into the right
lateral cerebral ventricle according to the following coordinates:
1.0 mm posterior from the Bregma, 1.5 mm lateral from the midline
and at a depth of 4.0 mm from the skull surface (19). TLR4-neutralizing antibody [200
µg/kg (20)], LPS [0.2 mg/kg
(21)], LPS [0.2
mg/kg]+TLR4-neutralizing antibody [200 µg/kg], LPS (0.2
mg/kg)+Y-27632 [10 mg/kg (22)],
and Y-27632 [10 mg/kg] was dissolved in saline to 5 µl and injected
to each group 30 min prior to MCAO surgery. The rats in the saline
group were injected with saline at the same volume as a control.
The injection procedure continued for 10 min, and the microinjector
remained for 5 min prior to withdrawal. The bone wound was closed
with bone wax, and then the rats underwent subsequent MCAO surgery.
Following 48 h post-MCAO establishment, the animals were
sacrificed, and the brains were removed immediately for further
study.
Immunofluorescence
Rats were anesthetized and perfused with PBS and 4%
buffered paraformaldehyde. Following collection of brain tissue,
the right cortex was post-fixed in 4% paraformaldehyde for 24 h at
4°C, embedded in paraffin and coronally sectioned at 4 µm. Sections
were permeabilized with 1% Triton X-100 for 2 h, then blocked with
10% goat serum (Beyotime Institute of Biotechnology, Shanghai,
China) in PBS with 0.3% Triton X-100 for 30 min at room
temperature. Sections were then incubated with anti-TLR4 (1:2,000),
anti-ROCK-II (1:1,000), anti-p-CRMP2 (1:1,000) or saline at 4°C
overnight. Following washing with PBS, sections were incubated with
the secondary antibodies [tetramethylrhodamine- (1:100, Goat
anti-Rabbit; cat no. T-2769; Thermo Fisher Scientific, Inc.) or
fluorescein isothiocyanate-conjugated (1:500, Goat anti-mouse; cat
no. A16079; Thermo Fisher Scientific, Inc.)] for 1 h at 37°C.
Finally, DAPI was employed for nuclei staining for 10 min at room
temperature. Saline was used as a negative control instead of
primary antibody. Images were obtained via fluorescence microscopy
(magnification, ×200). The Image Pro-Plus 6.0 Software (Media
Cybernetics, Inc., Rockville, MD, USA) enabled computer-controlled
image acquisition and processing. The relative optical density was
calculated, and the relative protein expression was then
determined.
Western blotting
Brain tissue was homogenized with lysis buffer (1%
Triton X-100, 1 mM EDTA in 1X PBS, pH 7.4) containing 200 µM
phenylmethylsulfonyl fluoride. Homogenates were centrifuged at
15,000 × g for 20 min at 4°C, and the supernatants were isolated.
Protein concentration was determined using a Bicinchoninic Acid
protein assay kit. Protein of the samples (40 µg) was separated
using 10% SDS-PAGE and transferred to nitrocellulose membranes. The
membranes were blocked in 5% nonfat milk for 1 h at 37°C, followed
by an overnight incubation at 4°C with primary antibodies
(anti-TLR4 1:1,000, anti-total CRMP2 1:1,000, anti-p-CRMP2 1:1,500,
anti-ROCK2 1:1,000, anti-β-actin 1:2,000). Following Tris-buffered
saline and 0.1% Tween-20 (TBST) washing and incubation with a
secondary antibody (cat. no. 7074, anti-rabbit, 1:5,000; cat. no.
7076 anti-mouse, 1:5,000; Cell Signaling Technology, Inc.) for 1 h
at 37°C, protein bands were visualized using a chemiluminescence
detection kit (cat no. P0018F; Beyotime Institute of Biotechnology,
Shanghai, China). β-actin was used as an endogenous loading
control. Relative protein expression levels were reflected by the
band density of the target proteins relative to β-actin. The
Quantity One software version 4.4 (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) was used to analyze the integrated absorbance
value of the band densities.
Modified neurological severity score
(mNSS)
To further clarify whether Rho-kinase acts on the
neurological deficits induced by LPS, neurological function was
evaluated using the mNSS at 24 h, 2 and 7 days, and 2 weeks
post-MCAO in the MCAO, LPS and LPS+Y-27632 groups (n=5 each group).
The mNSS is a composite of motor, sensory, balance and reflex
tests, and is graded on a scale from 0 to 18 (normal score, 0;
maximal deficit score, 18). A higher score indicates more severe
neurological behavioral impairments (23).
Statistical analysis
All experiments were repeated three times. Data were
presented as the mean ± standard deviation. All data were analyzed
using one-way analysis of variance followed by Fisher's exact test
for post-hoc analyses. P<0.05 was considered to indicate
statistically significant difference. Statistical analysis was
performed using SPSS 18.0 software (SPSS, Inc., Chicago, IL,
USA).
Results
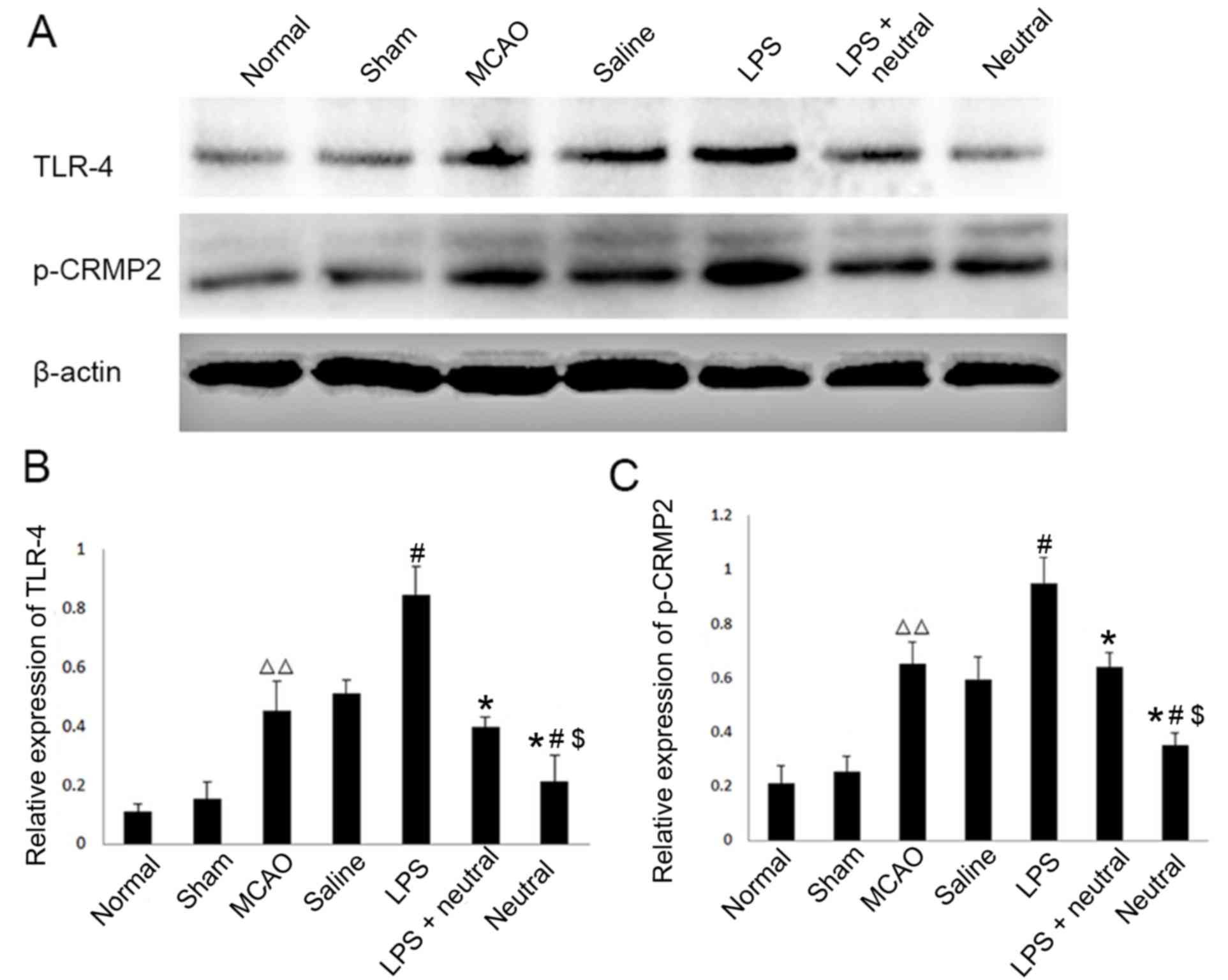
Expression levels of TLR4 and p-CRMP2
are significantly increased in MCAO rats as observed via western
blotting
The results of western blotting demonstrated that
the expression levels of TLR4 and p-CRMP2 were significantly
increased in the cortex of the MCAO group (TLR4, 0.453±0.098;
p-CRMP2, 0.653±0.078) when compared with the normal (TLR4,
0.112±0.023; P=0.0037; p-CRMP2, 0.211±0.064; P=0.0082) and sham
groups (TLR4, 0.153±0.056; P=0.0074; p-CRMP2, 0.253±0.056;
P=0.009). The TLR4-specific agonist LPS also significantly
increased the expression levels of TLR4 (0.8456±0.098, P=0.037) and
p-CRMP2 (0.948±0.098, P=0.024) in the cortex when compared with the
saline group. The application of the TLR4-neutralizing antibody
treatment suppressed the increased expression of TLR4 and p-CRMP2
induced by LPS induction, as observed in the reduction of TLR4
(0.396±0.034, P=0.029) and p-CRMP2 (0.640±0.054, P=0.031)
expression levels in the LPS+TLR4-neutralizing antibody group when
compared with the LPS group. When LPS was absent, the expression
levels of TLR4 (0.210±0.082, P=0.038) and p-CRMP2 (0.352±0.049,
P=0.017) were significantly decreased compared with LPS+
TLR4-neutralizing antibody group (Fig.
1).
Expression levels of TLR4 and p-CRMP2
are significantly increased in MCAO rats as observed by
immunofluorescence
The immunofluorescence results revealed that MCAO
treatment significantly increased the TLR4 (0.378±0.054; Fig. 2) and p-CRMP2 (0.540±0.044, Fig. 3) positive fluorescence in the
cortex compared with the normal group (TLR4: 0.162±0.027, P=0.075;
p-CRMP2: 0.173±0.039, P=0.003). Treatment with LPS also
significantly increased the intensity of TLR4 (0.726±0.068) and
p-CRMP2-positive (0.803±0.081) fluorescence in the cortex when
compared with the saline group (TLR4: 0.441±0.056, P=0.047;
p-CRMP2: 0.537±0.030, P=0.038). Application of the
TLR4-neutralizing antibody treatment significantly suppressed the
increased TLR4 and p-CRMP2-positive fluorescence induced by LPS
induction, as demonstrated by the decrease in TLR4 (0.604±0.050,
P=0.026) and p-CRMP2-positive fluorescence (0.620±0.093, P=0.008)
in the LPS+TLR4-neutralizing antibody group when compared with the
LPS group. The expression of TLR4 (0.412±0.059, P=0.007) and
p-CRMP2 (0.347±0.038, P=0.005) were significantly decreased
compared with the LPS+ TLR4-neutralizing antibody group (Figs. 2 and 3).
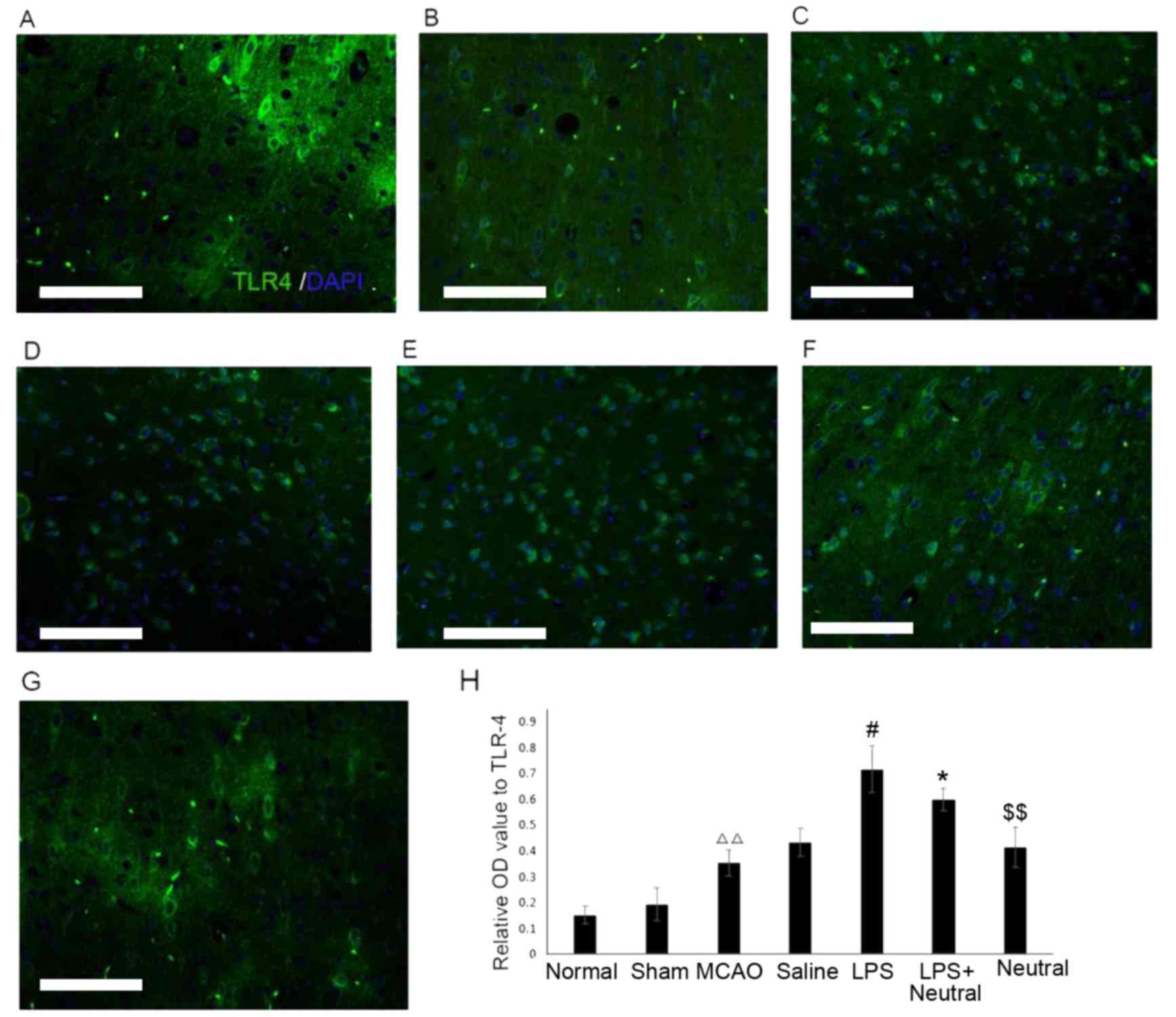 | Figure 2.Expression levels of TLR4 in the
cortex via immunofluorescence (×200). TLR4-positive expression in
the cortex in the (A) Normal, (B) Sham, (C) MCAO, (D) Saline, (E)
LPS, (F) LPS+TLR4-neutralizing antibody and (G) TLR4-neutralizing
antibody groups. (H) Quantitative analysis of the OD of
TLR4-positive cells in the cortex. Data are expressed as the mean ±
standard deviation (n=5). Scale bar=100 µm. ∆∆P<0.01
vs. the normal group; #P<0.05 vs. the MCAO group;
*P<0.05 vs. the LPS group; $$P<0.01 vs. the
LPS+Neutral group. OD, optical density; LPS, lipopolysaccharide;
MCAO, middle cerebral artery occlusion and reperfusion group;
Normal, rats that did not undergo MCAO; LPS+Neutral, rats treated
with LPS and TLR-neutralizing antibody prior to MCAO; Neutral, rats
treated with TLR-neutralizing antibody only prior to MCAO; Sham,
rats that underwent surgery without MCAO; Saline, rats administered
an intracerebroventricular injection of saline prior to MCAO; TLR4,
Toll-like receptor 4. |
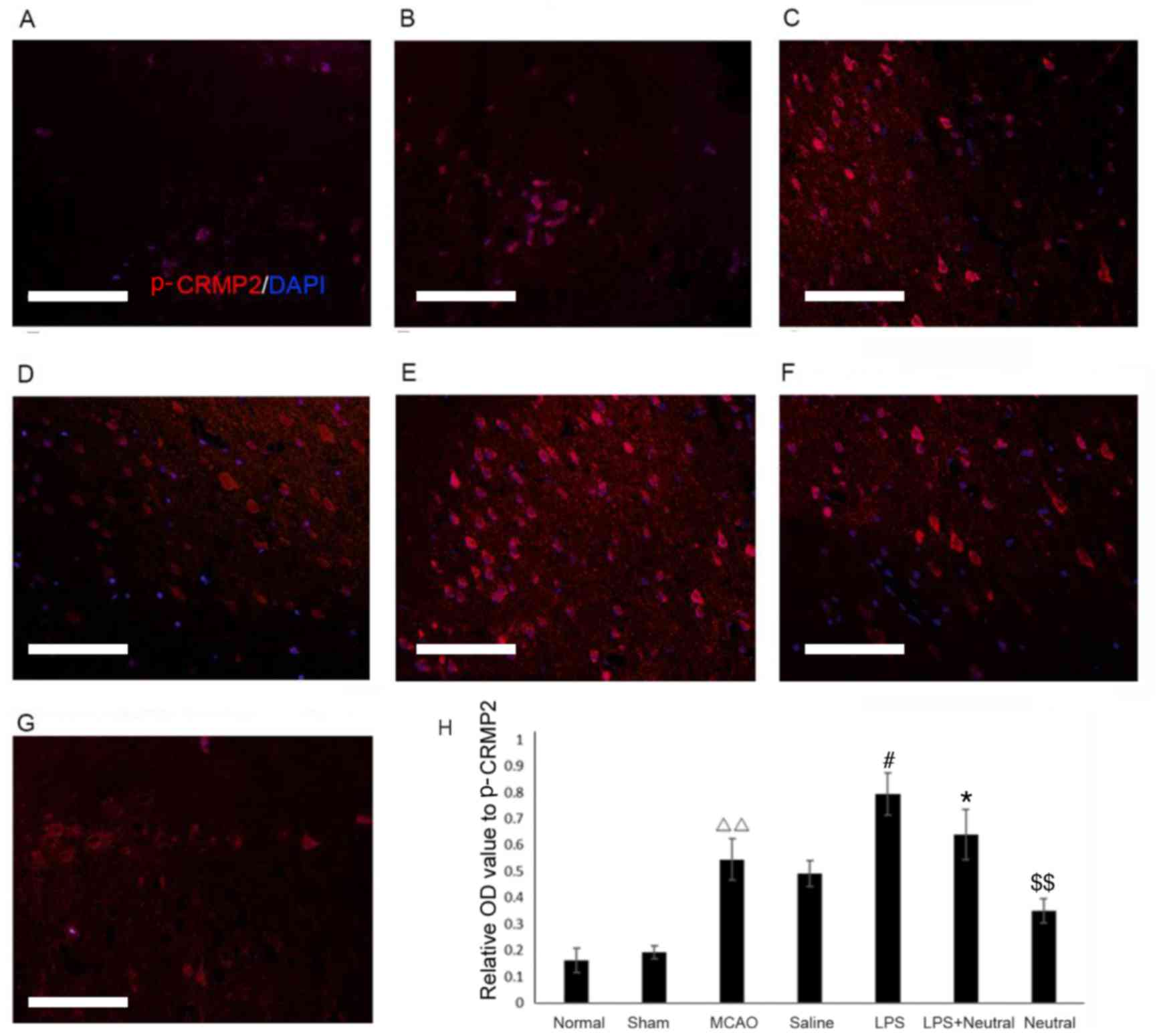 | Figure 3.Phosphorylation expression of CRMP2 in
the cortex via immunofluorescence (magnification, ×200). The
p-CRMP2-positive expression in the cortex of the (A) normal, (B)
sham, (C) MCAO, (D) saline, (E) LPS, (F) LPS+TLR4-neutralizing
antibody and (G) TLR4-neutralizing antibody treatment groups. (H)
Quantitative analysis of the OD of TLR4-positive cells in the
cortex relative to the OD value. Data are expressed as the mean ±
standard deviation (n=5). Scale bar=100 µm. ∆∆P<0.01
vs. the normal group; #P<0.05 vs. the MCAO group;
*P<0.05 vs. the LPS group; $$P<0.01 vs. the
LPS+neutral group. OD, optical density; p-CRMP2, phosphorylated
collapsin response mediator protein 2; TLR, Toll-like receptor;
LPS, lipopolysaccharide; MCAO, middle cerebral artery occlusion and
reperfusion group; Normal, rats that did not undergo MCAO; LPS+
neutral, rats treated with LPS and TLR-neutralizing antibody prior
to MCAO; Neutral, rats treated with TLR-neutralizing antibody only
prior to MCAO; Saline, rats administered an intracerebroventricular
injection of saline prior to MCAO; Sham, rats that underwent
surgery without MCAO. |
Rho kinase mediates the role of TLR4
in the phosphorylation of CRMP2
The present study further investigated whether Rho
kinase is involved in the increased levels of TLR4-induced CRMP2
phosphorylation. The Rho kinase inhibitor Y-27632 was
intracerebroventricularly injected into the MCAO rat models, and
the expression of TLR4, ROCK-II and p-CRMP2 were detected via
western blotting (Fig. 4). TLR4
protein expression levels exhibited no significant differences
between the LPS (0.457±0.087) and LPS+Y-27632 groups (0.588±0.092,
P=0.702), which suggested that the activation of Rho kinase may not
affect the expression of TLR4. However, ROCK-II and p-CRMP2
expression levels were significantly reduced in the LPS+Y-27632
(ROCK-II, 0.347±0.085, P=0.016; p-CRMP2, 0.568±0.054, P=0.031) and
Y-27632 (ROCK-II, 0.201±0.050, P=0.004; p-CRMP2, 0.333±0.046,
P=0.022) groups when compared with the LPS group (ROCK-II,
0.832±0.104; p-CRMP2, 0.732±0.098). Notably, Y-27632 treatment
alone decreased p-CRMP2, ROCK-II and TLR4 expression levels when
compared with the LPS+Y-27632 group (P<0.05; Fig. 4).
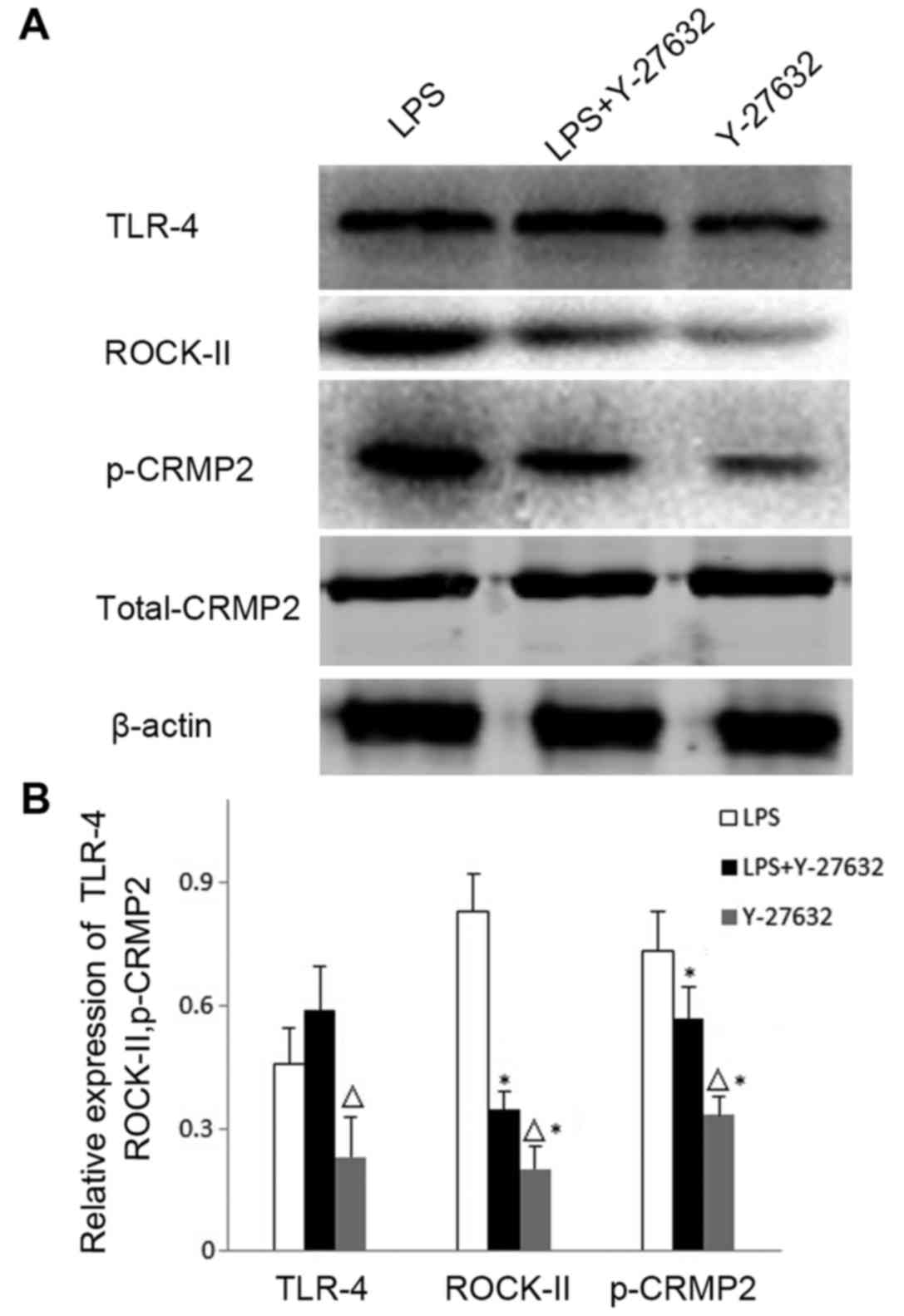 | Figure 4.Expression levels of TLR4, ROCK-II and
p-CRMP2 in the brain cortex as detected by western blotting. (A)
Western blotting was performed to reveal the expression levels of
TLR4, the activated Rho-kinase inhibitor ROCK-II and p-CRMP2 in the
LPS, LPS+Y-27632 and Y-27632 groups. (B) Quantitative analysis of
TLR4, ROCK-II and p-CRMP2 expression levels relative to β-actin in
the cortex from (A). Data are expressed as the mean ± standard
deviation (n=5). ∆P<0.05 vs. the LPS+Y-27632 group;
*P<0.05 vs. the LPS group. LPS, lipopolysaccharide; p-CRMP2,
phosphorylated collapsin response mediator protein 2; ROCK-II,
Rho-associated protein kinase 2; Y-27632, rats treated with the
ROCK-II inhibitor Y-27632; LPS+Y-27632, rats treated with LPS and
Y-27632; TLR4, Toll-like receptor 4. |
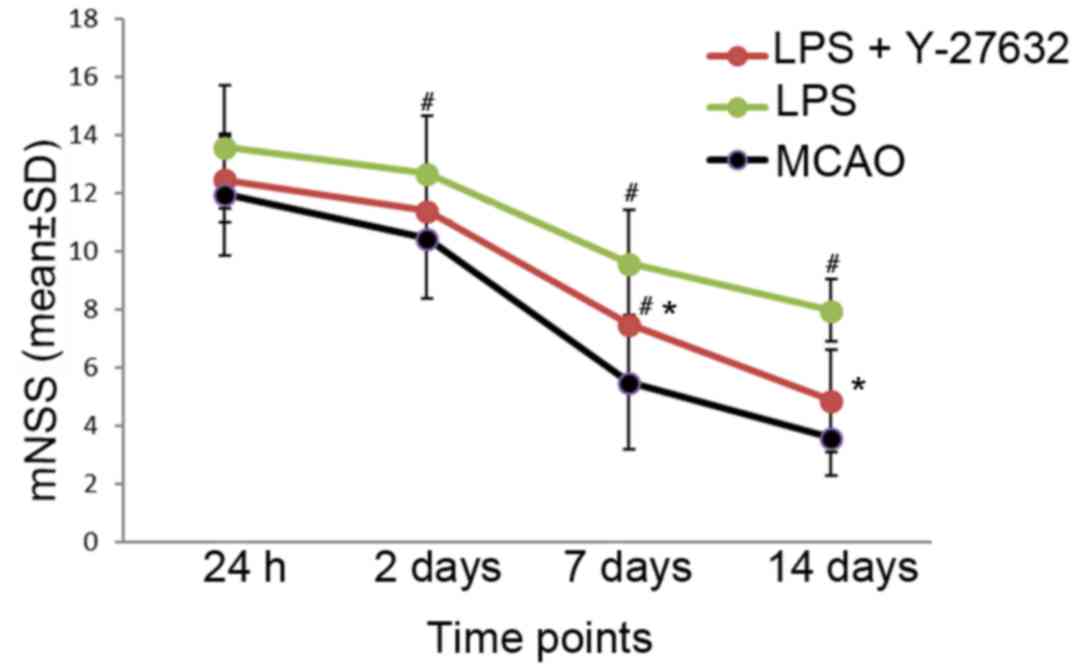
Neurological function is reversed by
Rho kinase inhibition
The mNSS score was used to evaluate the neurological
function following MCAO surgery. To study the effects of LPS and
Rho kinase inhibition on neurological function, LPS or LPS+Y-27632
was injected into rats at several time points including 24 h, 2 and
7 days, and 2 weeks post-MCAO to determine the mNSS score. The
results of the present study revealed no significant differences in
the mNSS score between the groups 24 h post-MCAO (MCAO:
11.945±2.094; LPS: 12.997±1.483; LPS+Y-27632 12.456±2.123,
P=0.866). Compared with in the MCAO group, the mNSS score in the
LPS group was significantly increased at 2 days (12.654±2.006,
P=0.044), 7 days (9.567±1.843, P=0.039) and 14 days (7.945±1.046,
P=0.027) following MCAO surgery, which means more severe
neurological deficits compared with the MCAO group. In addition,
Rho kinase inhibition may reverse the increased mNSS score
exhibited by the LPS group at 7 days (7.457±2.075, P=0.025) and 14
days (4.845±1.763, P=0.018) post-MCAO, as demonstrated by the
significant decrease in the mNSS score observed with the
administration of Y-27632 when compared with the LPS group
(Fig. 5).
Discussion
The present study demonstrated that the expression
levels of TLR4 and p-CRMP2 were significantly increased in the
cortex of rats with MCAO or LPS induction but were suppressed by
TLR4-neutralizing antibody treatment or Rho kinase inhibition. Rho
kinase specific inhibition reversed the aggravated neurological
deficits induced by LPS. The findings of the present study
suggested that TLR4 may promote the phosphorylation of CRMP2 via
the activation of the Rho-kinase signaling pathway in MCAO, thereby
contributing to the pathological mechanism of TLR4 in ischemia and
stroke.
TLRs are one of the major components of the innate
immune system. Recent studies have demonstrated that TLRs are
widely expressed in immune-associated cell types (24,25),
which are also located in non-immune cells, including neurons,
microglia and epithelial cells (26). TLR expression levels have been
reported to be significantly increased in neurons in response to
ischemic conditions (5,7). TLR4 inhibition may prevent neuronal
death in MCAO rat models (4);
however, the mechanism involved has not been clarified. In the
present study, the expression levels of TLR4 were significantly
increased following MCAO and the TLR4-specific agonist LPS
significantly promoted the expression of TLR4, whereas the
TLR4-neutralizing antibody reduced the expression levels of TLR4 in
MCAO rats. In addition, the results of the present study indicated
that neurological function may be inhibited when the expression
levels of TLR4 are increased by LPS. These results suggested that
TLR4 may serve an important role in the pathogenesis of MCAO.
CRMP2 is a multifunctional adaptor protein that
binds cytoskeletal partners to serve a crucial role in microtubule
dynamics, neurite outgrowth and retraction, neural differentiation
and neurotransmitter release, as well as other important
neurophysiological functions (27). CRMP2 phosphorylation by Rho-kinase
destabilizes microtubule bundles in the growth cone, in which the
cell loses its ability to promote microtubule assembly, ultimately
leading to growth cone collapse and the inhibition of
neuroregeneration (28,29). Wang et al (30) reported that the expression levels
of p-CRMP2 were notably elevated in MCAO rats and induced severe
neurological deficits. In the present study, the expression levels
of p-CRMP2 in the cortex were significantly increased
post-MCAO.
Additionally, the results of the present study
demonstrated that the activation of TLR4 by LPS significantly
promoted the expression levels of p-CRMP2, whereas the inhibition
of TLR4 by TLR4-neutralizing antibody significantly reduced the
expression of p-CRMP2. These results suggested that TLR4 may
regulate the phosphorylation of CRMP2 in MCAO rats. To further
investigate the signaling pathway underlying TLR4 regulation
associated with the phosphorylation of CRMP2, LPS and the specific
Rho-kinase inhibitor, Y-27632, were administered to the brains
prior to ischemic injury in the present study. Western blotting
revealed that Y-27632 had no effect on the increased expression of
TLR4 induced by LPS; however, the expression levels of TLR4,
ROCK-II and p-CRMP2 were significantly suppressed by Y-27632 only
treatment. These results indicated that the phosphorylation of
CRMP2 may be activated by TLR4, which was suppressed following the
inhibition of Rho kinase activation. The negative control (normal)
was not included in this western blotting experiment, which may
present a limitation of the present study.
The results of the present study demonstrated that
TLR4 promoted the phosphorylation of CRMP2 via the activation of
Rho-kinase. Additionally, the deterioration of neurological
deficits associated with LPS intervention may be alleviated by the
suppression of Rho-kinase and p-CRMP2. This suggests that the
neurological impairments caused by TLR4 may be mediated by
Rho-kinase and p-CRMP2.
However, additional experiments are required to
support the conclusions of the present study. For example, further
investigation of the direct interaction between p-CRMP and
Rho-kinase, except for intervention with specific inhibitors, is
required. In addition, it is also essential that histopathological
analysis, such as Evans Blue/hematoxylin and eosin staining is
conducted to study the levels of apoptosis/necrosis in neuronal
cells and further evaluate the brain damage, which may support the
results of behavioral neurological testing conducted in the present
study.
In conclusion, the present study demonstrated that
TLR4 may promote the phosphorylation of CRMP2 in MCAO rats, which
may have been mediated via the activation of Rho-kinase. This helps
to further clarify the pathogenesis of TLR in stroke; modulation of
TLR4 could be a potential target to limit secondary post-stroke
brain damage in future clinical applications.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Department of
Education, Guangdong Government under the Top-tier University
Development Scheme for Research and Control of Infectious Diseases
(grant no. 2015064), National Natural Science Foundation Council of
China (grant nos. 81072508 and 81501634, Natural Science Foundation
of Shandong Province (grant no. ZR2014HQ018), Project of Shandong
Province Higher Educational Science and Technology Program (no.
J17KA240), China Postdoctoral Science Foundation (no. 2017M612701)
and The Special Project of Technical Innovation about Social and
People's Livelihood in Chongqing (no. cstc2015shmszx0017).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XY and XL conceived the idea of the study and
designed research; LL and JF analysed the data; CD and XL
interpreted the results; MD wrote the paper, raised the animals and
performed the western blot protocol; all authors performed
research, discussed the results and revised the manuscript.
Ethics approval and consent to
participate
The present study and experimental protocol was
established, according to the ethical guidelines of the Helsinki
Declaration and was approved by the Ethics Committee of Department
of Forensic Medicine, Shantou University (Shantou, China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mensah GA, Norrving B and Feigin VL: The
global burden of stroke. Neuroepidemiology. 45:143–145. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Herson PS and Traystman RJ: Animal models
of stroke: Translational potential at present and in 2050. Future
Neurol. 9:541–551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sierra C, Coca A and Schiffrin EL:
Vascular mechanisms in the pathogenesis of stroke. Curr Hypertens
Rep. 13:200–207. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu X, Wen Z, Zhao N, Xu X, Wang F, Gao J,
Jiang Y and Liu X: MicroRNA-1906, a Novel regulator of toll-like
receptor 4, ameliorates ischemic injury after experimental stroke
in mice. J Neurosci. 37:10498–10515. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tang SC, Arumugam TV, Xu X, Cheng A,
Mughal MR, Jo DG, Lathia JD, Siler DA, Chigurupati S, Ouyang X, et
al: Pivotal role for neuronal Toll-like receptors in ischemic brain
injury and functional deficits. Proc Natl Acad Sci USA.
104:13798–13803. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tang SC, Yeh SJ, Li YI, Wang YC, Baik SH,
Santro T, Widiapradja A, Manzanero S, Sobey CG, Jo DG, et al:
Evidence for a detrimental role of TLR8 in ischemic stroke. Exp
Neurol. 250:341–347. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Olson JK and Miller SD: Microglia initiate
central nervous system innate and adaptive immune responses through
multiple TLRs. J Immunol. 173:3916–3924. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lehnardt S, Lehmann S, Kaul D, Tschimmel
K, Hoffmann O, Cho S, Krueger C, Nitsch R, Meisel A and Weber JR:
Toll-like receptor 2 mediates CNS injury in focal cerebral
ischemia. J Neuroimmunol. 190:28–33. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fukata Y, Itoh TJ, Kimura T, Ménager C,
Nishimura T, Shiromizu T, Watanabe H, Inagaki N, Iwamatsu A, Hotani
H and Kaibuchi K: CRMP-2 binds to tubulin heterodimers to promote
microtubule assembly. Nat Cell Biol. 4:583–591. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yoshimura T, Kawano Y, Arimura N, Kawabata
S, Kikuchi A and Kaibuchi K: GSK-3beta regulates phosphorylation of
CRMP-2 and neuronal polarity. Cell. 120:137–149. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gim SA, Sung JH, Shah FA, Kim MO and Koh
PO: Ferulic acid regulates the AKT/GSK-3β/CRMP-2 signaling pathway
in a middle cerebral artery occlusion animal model. Lab Anim Res.
29:63–69. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ye Y, Xu H, Zhang X, Li Z, Jia Y, He X and
Huang JH: Association between toll-like receptor 4 expression and
neural stem cell proliferation in the hippocampus following
traumatic brain injury in mice. Int J Mol Sci. 15:12651–12664.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Julian L and Olson MF: Rho-associated
coiled-coil containing kinases (ROCK): Structure, regulation, and
functions. Small GTPases. 5:e298462014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoneda A, Multhaupt HA and Couchman JR:
The rho kinases I and II regulate different aspects of myosin II
activity. J Cell Biol. 170:443–453. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
National Research Council, . Guide for the
care and use of laboratory animals. 8th edition. National Academy
Press; Washington, DC: 2011, https://grants.nih.gov/grants/olaw/guide-for-the-care-and-use-of-laboratory-animals.pdfPubMed/NCBI
|
|
16
|
Arumugam TV, Salter JW, Chidlow JH,
Ballantyne CM, Kevil CG and Granger DN: Contributions of LFA-1 and
Mac-1 to brain injury and microvascular dysfunction induced by
transient middle cerebral artery occlusion. Am J Physiol Heart Circ
Physiol. 287:H2555–H2560. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xiong X, Barreto GE, Xu L, Ouyang YB, Xie
X and Giffard RG: Increased brain injury and worsened neurological
outcome in interleukin-4 knockout mice after transient focal
cerebral ischemia. Stroke. 42:2026–2032. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Iwasa T, Matsuzaki T, Tungalagsuvd A,
Munkhzaya M, Kawami T, Kato T, Kuwahara A, Yasui T and Irahara M:
Effects of ovariectomy on the inflammatory responses of female rats
to the central injection of lipopolysaccharide. J Neuroimmunol.
277:50–56. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma Y, Zhang X, Bao H, Mi S, Cai W, Yan H,
Wang Q, Wang Z, Yan J, Fan GC, et al: Toll-like receptor (TLR) 2
and TLR4 differentially regulate doxorubicin induced cardiomyopathy
in mice. PLoS One. 7:e407632012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gong G, Bai S, Wu W, Hu L, Liu Y, Niu J,
Dai X, Yin L and Wang X: Lrg participates in lipopolysaccharide
preconditioning-induced brain ischemia injury via TLR4 signaling
pathway. J Mol Neurosci. 54:20–26. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rikitake Y, Kim HH, Huang Z, Seto M, Yano
K, Asano T, Moskowitz MA and Liao JK: Inhibition of rho kinase
(ROCK) leads to increased cerebral blood flow and stroke
protection. Stroke. 36:2251–2257. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sugawara T, Ayer R, Jadhav V and Zhang JH:
A new grading system evaluating bleeding scale in filament
perforation subarachnoid hemorrhage rat model. J Neurosci Methods.
167:327–334. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kawai T and Akira S: The role of
pattern-recognition receptors in innate immunity: Update on
toll-like receptors. Nat Immunol. 11:373–384. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Medzhitov R and Janeway CA Jr: Innate
immunity: The virtues of a nonclonal system of recognition. Cell.
91:295–298. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Doi Y, Mizuno T, Maki Y, Jin S, Mizoguchi
H, Ikeyama M, Doi M, Michikawa M, Takeuchi H and Suzumura A:
Microglia activated with the toll-like receptor 9 ligand CpG
attenuate oligomeric amyloid {beta} neurotoxicity in in vitro and
in vivo models of Alzheimer's disease. Am J Pathol. 175:2121–2132.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hensley K, Venkova K, Christov A, Gunning
W and Park J: Collapsin response mediator protein-2: An emerging
pathologic feature and therapeutic target for neurodisease
indications. Mol Neurobiol. 43:180–191. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Brown M, Jacobs T, Eickholt B, Ferrari G,
Teo M, Monfries C, Qi RZ, Leung T, Lim L and Hall C:
Alpha2-chimaerin, cyclin-dependent Kinase 5/p35, and its target
collapsin response mediator protein-2 are essential components in
semaphorin 3A-induced growth-cone collapse. J Neurosci.
24:8994–9004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Arimura N, Ménager C, Kawano Y, Yoshimura
T, Kawabata S, Hattori A, Fukata Y, Amano M, Goshima Y, Inagaki M,
et al: Phosphorylation by rho kinase regulates CRMP-2 activity in
growth cones. Mol Cell Biol. 25:9973–9984. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang T, Wu X, Yin C, Klebe D, Zhang JH and
Qin X: CRMP-2 is involved in axon growth inhibition induced by RGMa
in vitro and in vivo. Mol Neurobiol. 47:903–913. 2013. View Article : Google Scholar : PubMed/NCBI
|