Introduction
As a major public health problem, the incidence of
asthma is on the rise worldwide (1,2).
This condition is accompanied by airflow obstruction. Airway
remodelling is one of the major characteristics of asthma, and has
attracted many attentions due to its critical role in asthma
disease (3). Bronchial epithelium,
as a physical mediator between the internal milieu and the external
environment, is exposed to many environmental factors (i.e. air
pollutants, allergens and environmental chemicals), therefore
leading remodelling, which is considered as a response to cell and
tissue damage (4,5). Many efforts have been done to combat
asthma, however, only a few strategies proved to be effective in
preventing/alleviating this condition. Therefore, it is necessary
to develop new methods for asthma treatment. Benzo[a]pyrene (Bap)
is a common environmental chemical carcinogen, which has strong
carcinogenicity, mutagenicity and teratogenicity (6). The atmospheric Bap mainly comes from
wood burning (7). Lung epidermal
cells are the main media, via which some harmful substances are
absorbed by human bodies. Research reported that Bap is able to
induce pulmonary epithelial injury (8). Thus, Bap will be used to help set up
the bronchial epithelium injury model in the present study.
The balance of redox state is considered to
contribute to multiple cellular events (9). Emerging evidences have suggested that
reactive oxygen species (ROS) are the mediator for the chronic
airway diseases, which includes asthma (10,11).
ROS is a byproduct of cell respiration, and it can be removed by
antioxidant enzymes under normal conditions. However, if the
oxidant/antioxidant balance is disrupted, excessive ROS oxidation
metabolites will accumulate in cells, and therefore inducing
oxidative stress and creating impairments to the cells (12). The predominant consequence of
oxidative stress is the cell apoptosis (13). Thus, strategies targeting at the
oxidative stress and apoptosis of bronchial epithelial cells may be
effective to control tissue injury in the treatment of asthma.
Various molecules and factors, such as phosphatase
and tensin homolog (PTEN), focal adhesion kinase (FAK) and
phosphatidylinositol 3-kinase (PI3K)/Akt, are associated with cell
survival (14,15). PTEN has generally been recognized
as a tumor suppressor (16), and
it may be a gatekeeper in acute lung injury and fibrosis (17). Moreover, PI3K/Akt, as an important
signaling pathway that regulates cell survival, is possibly
negatively regulated by PTEN (18). Furthermore, FAK is critical to the
integration of various signals (19), which are of significance to the
cell survival and apoptosis (15,20).
FAK is reported to be the upstream of PI3K/Akt signaling pathway as
well (21). Therefore, these
signals may be the possible molecule targets in the treatment of
bronchial epithelial cell injury.
Annexin A1 (ANXA1), which originally believed to
inhibit phospholipase activity (22), has been reported to be able to
modulate various cellular functions in multiple cell types
(23,24). Protective roles of ANXA1 have been
observed (25,26). A recent study demonstrated that
ANXA1 is able to accelerate the healing of gastric ulcers that
caused by indomethacin (27).
However, to the best of our knowledge, the regulatory role of ANXA1
in bronchial epithelium injury and the potential mechanisms have
not been validated. Thus, the main scope of this study was to
determine the effect(s) of ANXA1 on bronchial epithelial cell
injury in vitro.
Materials and methods
Cell culture and cell viability
assay
Human bronchial epithelial cells BEAS-2B (CRL-9609;
ATCC, Manassas, VA, USA) (passage numbers 60–80) were maintained in
Dulbecco's Modified Eagle's Medium (DMEM; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) that containing with 10% fetal bovine
serum, and, 1% streptomycin/penicillin (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C with and 5% CO2 at 37°C. CCK-8
method (Beyotime Institute of Biotechnology, Haimen, China) was
adopted to measure the cell viability. The cells were seeded at a
density of 2.5×103 cell/well. After 24 h, 10 µl CCK-8
solutions were added into the incubator and then maintained with
the cells at 37°C for 4 h. The absorbance 450 nm was measured by a
microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Cell grouping and transfection
The cell grouping were as follows: i) Control group,
normal cells; ii) empty vector group, cells that were transfected
with negative vectors; iii) NXA1 group: cells that were transfected
with ANXA1 over-expression vectors; iv) Bap group, cells that were
treated with Bap (64 µM for 6 h, as stated in the results
section); v) emp+Bap, Cells that were transfected with negative
vectors, and then treated with 64 µM Bap for 6 h), Anx+Bap: Cells
that were transfected with ANXA1 vector and then treated with 64 µM
Bap for 6 h. E.V.=empty vector, Bap=benzo[a]pyrene. Lipofectamine
LTX™ Reagent (Invitrogen; Thermo Fisher Scientific, Inc.) was used
for cell transfection. The p-Receiver-M01/ANXA1 vector for
over-expression were ordered from GeneCopoeia (Rockville, MD, USA).
The expression vector p-Receiver-M01 served as negative control
vector. The cells were cultured in serum-free medium overnight.
Lipofectamine Reagent and DNA solution (vector ANXA1 or negative
control) were gently mixed. After being incubation for 30 min, the
Lipofectamine-DNA complex was added into cells, and mixed by
rocking the plate back and forth. Subsequently, the plate was
incubated in an incubator. Then the cells in each group were
treated with or without Bap. The cells were pretreated with 500 nM
SF1670 (MedchemExpress, South Brunswick, NJ, USA) at 37°C for 30
min to inhibit the PTEN pathway, following steps described in a
previous study (28).
Apoptosis estimation
Flow cytometry (FCM) assay was performed to assess
cell apoptosis using FITC/Annexin V Apoptosis detection kit I (BD
Biosciences, Franklin Lakes, NJ, USA), according to the
manufacturer's instructions. The cells were incubated with
FITC/Annexin V and PI at room temperature in the dark for
cellstaining. The cells were fixed with paraformaldehyde (0.5%),
and then they were loaded on a FACStar Plus flow cytometer (BD
Biosciences).
ROS content detection
To evaluate the intracellular ROS levels, the cells
were stained with 10 µM fluorescent dye 2,7-dichlorofluorescein
diacetate (DCFH-DA) dye (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) at 37°C for 15 min in the dark. DCFH-DA can be transformed
into fluorescent DCF by the presence of ROS. The fluorescent
intensity indicated the ROS levels. The DCF fluorescence signals
were detected by a fluorescence plate reader (BioTek Instruments,
Inc., Winooski, VT, USA).
Oxidative stress measurement
The cells were centrifuged and the supernatant was
collected. Following the manufacturer's instructions, the
activities of superoxide dismutase (SOD; S0101), glutathione
peroxidases (GPX; S0056), malondialdehyde (MDA; S0131) and lactic
dehydrogenase (LDH; C0016) were assessed by available commercial
kits (Beyotime Institute of Biotechnology). A microplate reader
(Bio-Rad Laboratories, Inc.) was used to measure the absorbance
when appropriate.
Enzyme linked immunosorbent assay
(ELISA)
Active Caspase-3 Quantikine ELISA kit (R&D
Systems, Inc., Minneapolis, MN, USA) was used to detect the
activity of active caspase-3. To explain further, the cells were
collected and re-suspended. Then, the samples were added into each
well and then incubated at room temperature for 1 h. After being
washed, the substrate solution was added into the wells and then
incubated for 30 min at room temperature. Finally, the stop
solution was added into each well. The absorbance was read at 450
nm using a microplate reader (Bio-Rad Laboratories, Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated from the harvested cells
using TRizol regent (Invitrogen; Thermo Fisher Scientific, Inc.),
following the manufacturer's protocols. The integrity of RNA was
tested by agarose gel electrophoresis. The total RNA (1 µg) was
reverse-transcribed into cDNA using M-MLV (Promega Corporation,
Madison, WI, USA). RT-qPCR analysis was performed to detect the
relative expression of target genes. The amplification was
conducted on ABI Prism 7,500 Sequence detection instrument using
SYBR Premix TaqTM II kit (Takara Bio, Inc., Otsu, Japan). The
2−∆∆Cq method was used for expression quantification
(29). The primers used were as
follows: Cyclin D1 forward: 5′-CCCTCGGTGTCCTACTTCAA-3′; Cyclin D1
reverse: 5′-CTTAGAGGCCACGAACATGC-3′; B-cell lymphoma 2 (Bcl-2)
forward: 5′-CACACACACACATTCAGGCA-3′; Bcl-2 reverse:
5′-GGCAATTCCTGGTTCGGTTT-3′; Bcl-2-associated X protein (Bax)
forward: 5′-TGGCCTCCTTTCCTACTTCG-3′; Bax reverse:
5′-AAAATGCCTTTCCCCGTTCC-3′; β-actin forward: 5′CCC GCG AGC ACA GCT
TCT TTG3′; β-actin reverse: 5′ACATGCCGGAGCCGTTGTCGAC3′.
Western blot analysis
The lysed cells were centrifuged at 12,000 g
at 4°C for 10 min. The protein concentration was determined by BCA
Protein Assay Kit (Bio-Rad Laboratories, Inc.). The proteins were
denatured by being heated in boiling water for 5 min. The proteins
were then separated on SDS-PAGE gel by electrophoresis. After being
transferred onto PVDF membrane, the proteins were blocked with
skimm milk for 2 h at room temperature. Primary antibodies were
incubated with PVDF membrane overnight at 4°C. Information of
primary antibodies used in the experiment was as follows:
Anti-ANXA1 (ab19830, 1:2,000), anti-Cyclin D1 (ab134175, 1:10,000),
anti-Bax (ab32503, 1:1,000), anti-Bcl-2 (ab692, 1:500), anti-PTEN
(ab170941, 1:4,000), anti-FAK (ab76496, 1:1,000), anti-Akt1/2
(ab182729, 1:5,000), anti-p-Akt (ser473) (ab81283, 1:5,000; all
from Abcam, Cambridge, UK), PI3Kinase Class III (4263, 1:1,000; CST
Biological Reagents Co., Ltd., Shanghai, China), anti-p-PI3Kinase
Class III (Ser249) (13857, 1:1,000; CST Biological Reagents Co.,
Ltd.) and anti-β-actin (ab8226, 1:5,000; Abcam). Subsequently, the
secondary antibodies (ab205718, 1:5,000; Abcam) coupled with
horseradish peroxidase were added and interact with the primary
antibodies. The band was developed with enhanced chemiluminescence
system (GE Healthcare, Chicago, IL, USA). The grey value was read
using quantity one 4.6.2.
Statistical analysis
Data are presented as mean ± standard deviation.
Student's t-test or one-way analysis of variance followed by
Dunnett's post hoc test was performed to compare the differences
among groups by using GraphPad Prism Software 6 (GraphPad Software,
Inc., La Jolla, CA, USA), when appropriate. P<0.05 was
considered to indicate a statistically significant difference.
Results
ANXA1 improved the viability of
Bap-treated bronchial epithelial cells
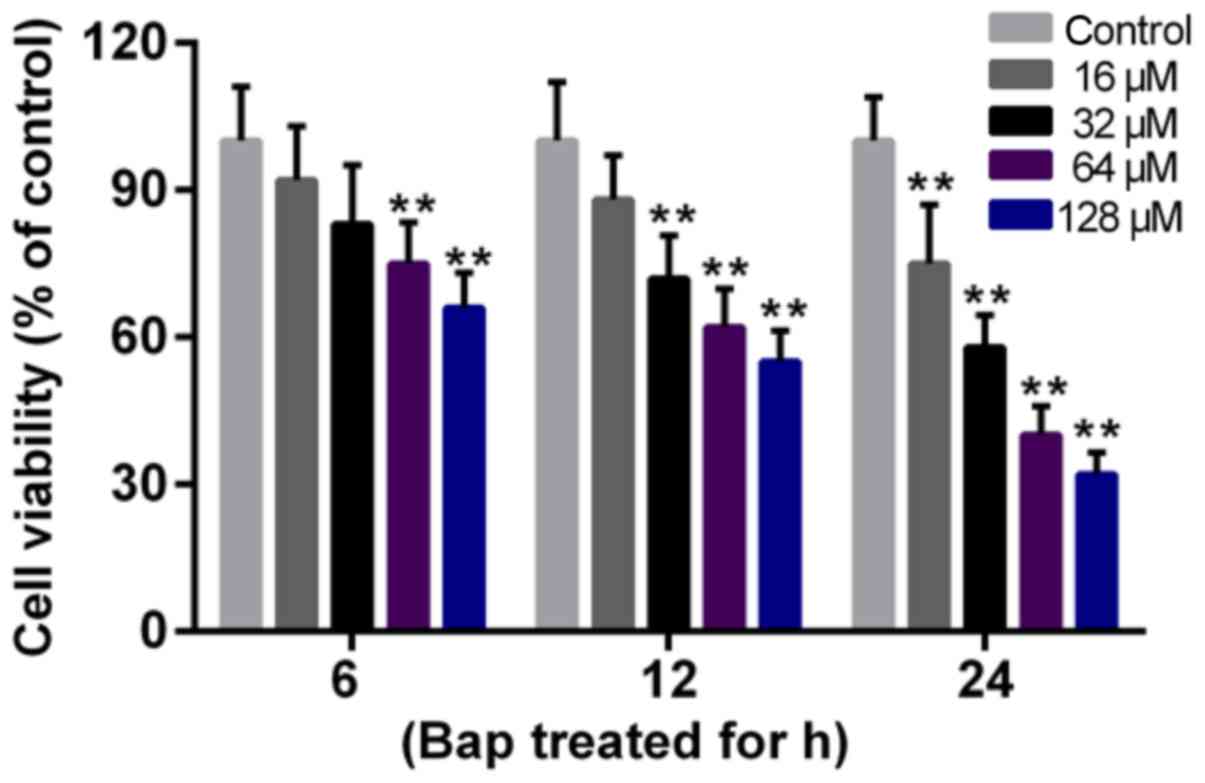
The effect of Bap on cell viability was determined
first. Data showed that the cell viability deceased gradually with
the increase of time and of the dosage of Bap. The viability began
to decline significantly after 6 h of 64 µM Bap incubation
(Fig. 1). Thus, incubating 64 µM
Bap for 6 h was selected for inducing bronchial epithelium injury.
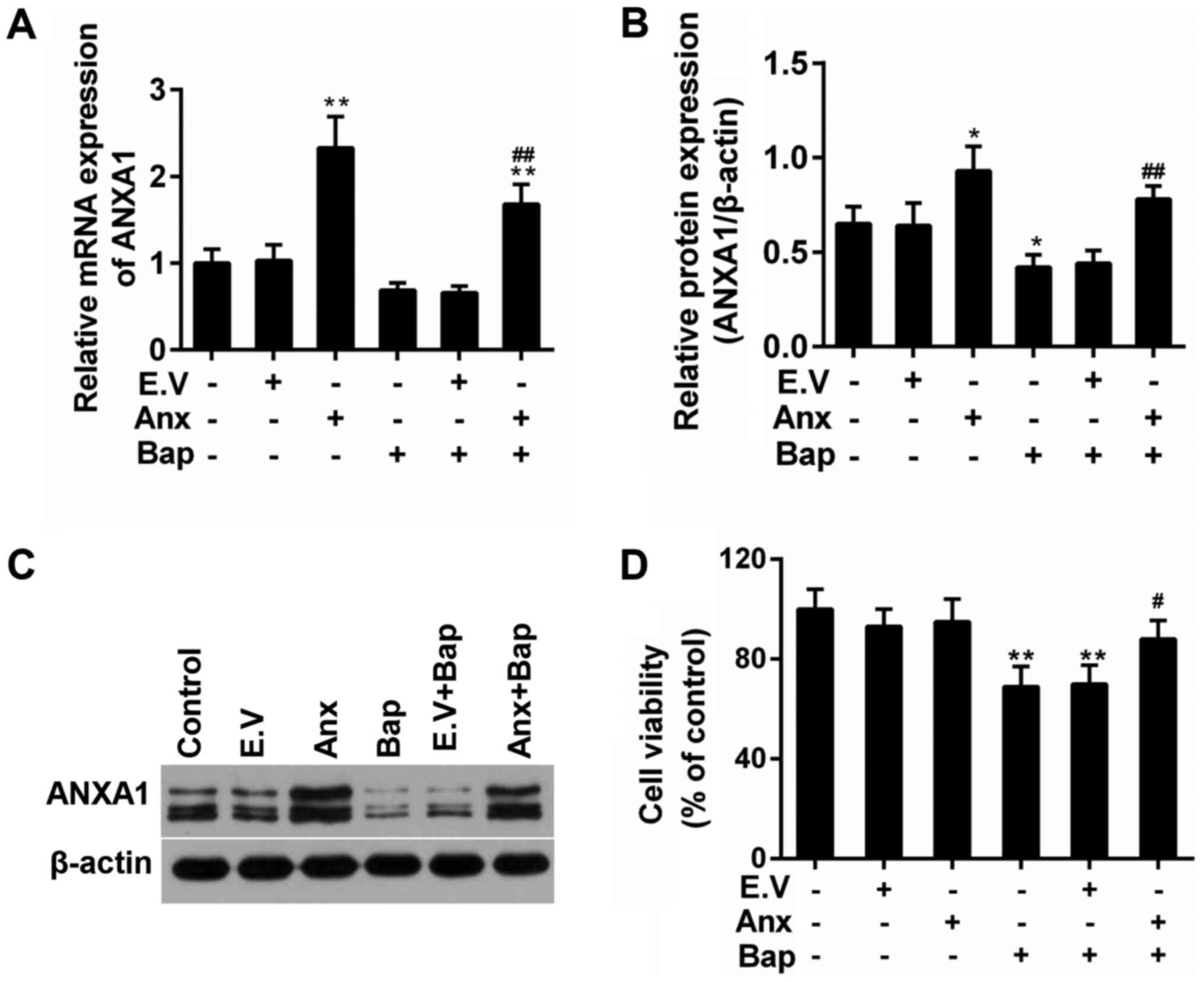
Moreover, the expression of ANXA1 was depressed by the presence of
Bap in mRNA and protein levels. However, we observed that the
over-expression of ANXA1 reversed this phenomenon (Fig. 2A-C). Furthermore, the decreased
viability of bronchial epithelial cells was recovered by ANXA1
over-expression (Fig. 2D).
ANXA1 reduced the Bap-mediated
oxidative stress in bronchial epithelial cells
ROS induction is a critical mechanism of bronchial
epithelium injury (30). The
effect of ANXA1 on ROS generation was tested. Our data indicated
that the intracellular ROS content was mitigated in ANXA1+Bap
group, compared to Bap group (Fig.
3A). Furthermore, compared to Bap group, the activity of free
radical scavenging enzymes, including superoxide dismutase (SOD)
and glutathione peroxidase (Gpx) was rescued. however, the content
of oxidative injury makers, including malondialdehyde (MDA) and
lactate dehydrogenase (LDH), was declined by ANXA1 over-expression
(Fig. 3B).
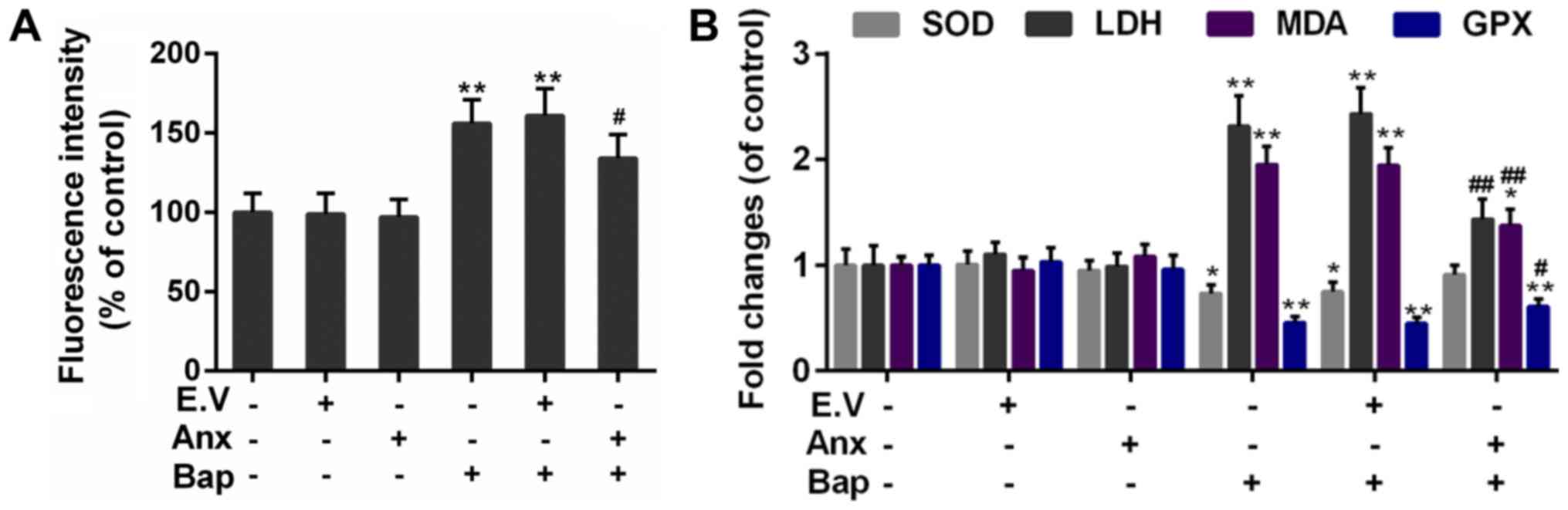 | Figure 3.Effect of ANXA1 on oxidative stress.
(A) The ROS content was determined by 2,7-dichlorofluorescein
diacetate dye. (B) The activities of SOD, GPX, MDA and LDH were
regulated by ANXA1. The cells were treated with 64 µM Bap for 6 h
to establish the cell injury model. *P<0.05 and **P<0.01 vs.
control group; #P<0.05 and ##P<0.01 vs.
E.V.+Bap group. ANXA1/Anx, Annexin A1; ROS, reactive oxygen
species; SOD, superoxide dismutase; GPX, glutathione peroxidases;
MDA, malondialdehyde; LDH, lactic dehydrogenase; Bap,
benzo[a]pyrene; E.V., empty vector. |
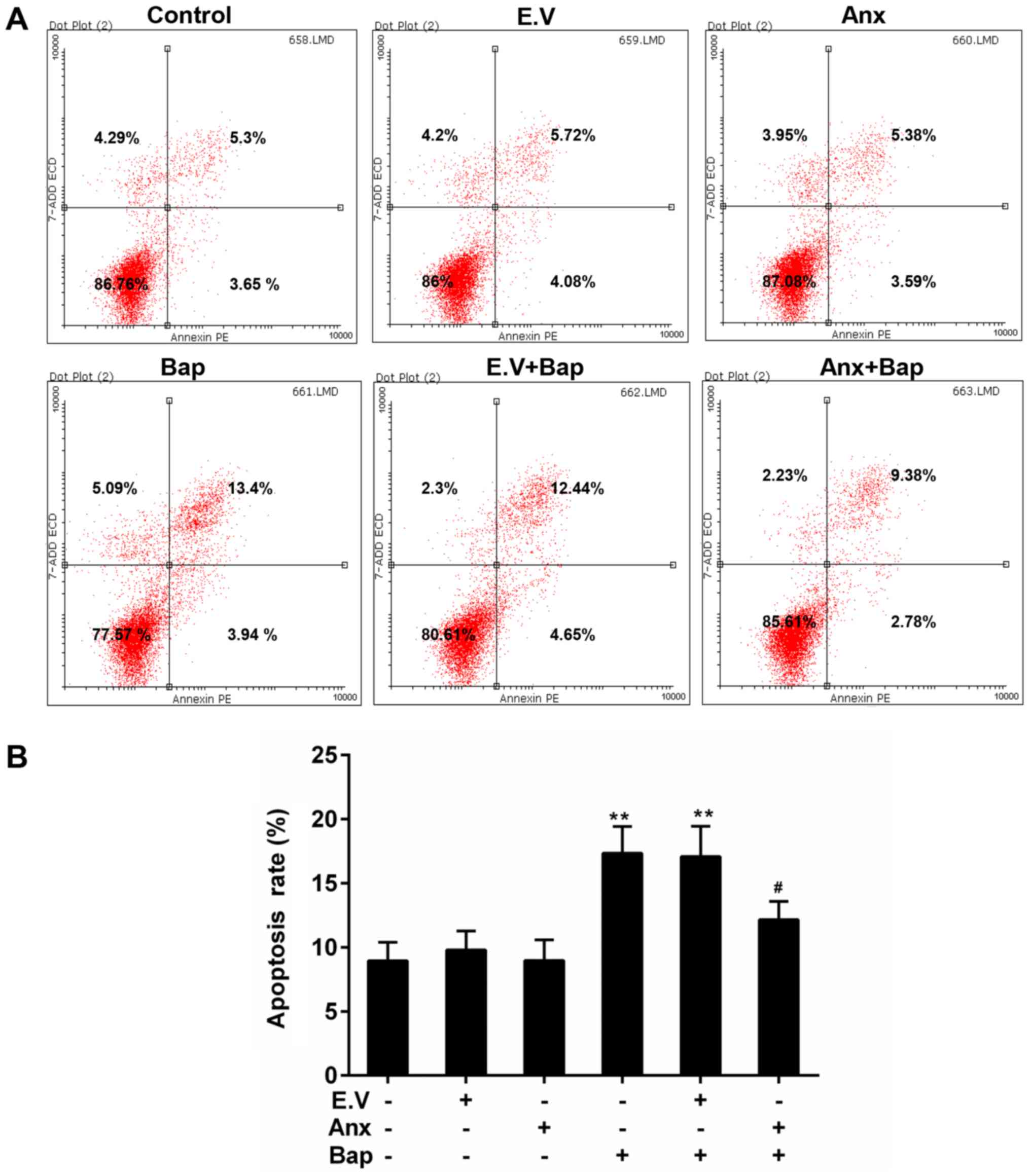
ANXA1 depressed the Bap-mediated
apoptosis in bronchial epithelial cells
Apoptosis can be triggered by oxidative stress
(31). FCM data showed that cell
apoptosis level was lower in ANXA1+Bap group than that in Bap group
(Fig. 4A and B). Moreover, the
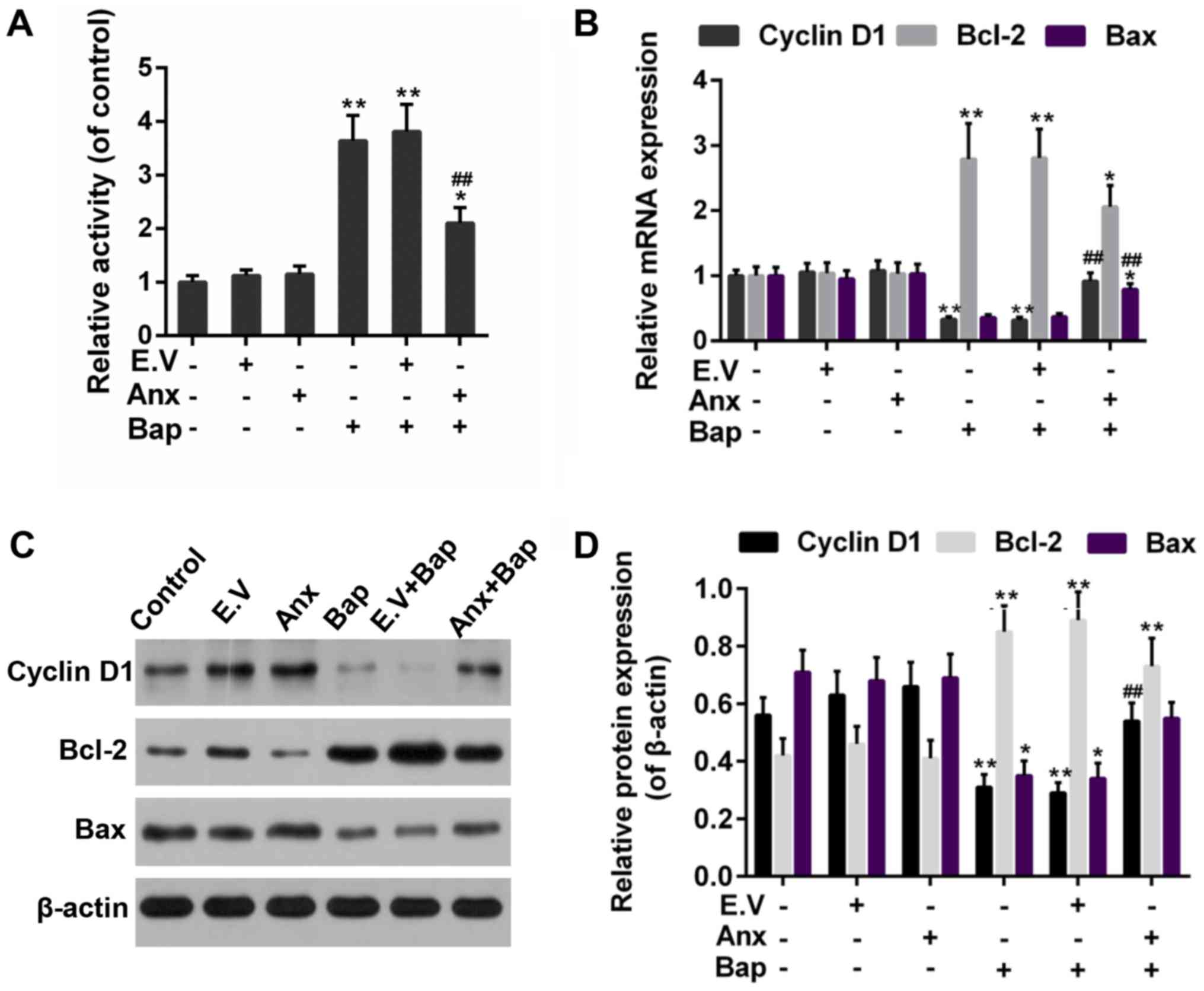
activity of caspase-3 was reduced by the over-expression of ANXA1
(Fig. 5A). In addition, Bcl-2 and
Bax, as members of the Bcl-2 family, is pro-apoptotic and
anti-apoptotic apoptosis proteins, respectively (32). Cyclin D1, as a cell cycle
regulator, is active in cell apoptosis pathways (33). Thus, the anti-apoptotic effect of
ANXA1 was further determined by testing the expression of these
apoptosis-related genes. Our data recorded that the increase
expression of Bcl-2 was alleviated, whereas the expression of Bax
and cyclin D1 was rescued by the over-expression of ANXA1 in both
mRNA and protein levels, compared to Bap group (Fig. 5B-D).
ANXA1 modulated the activity of
PTEN/FAK/PI3K/Akt signals
PTEN/PI3K/Akt is reported to be related to the lung
cell injury (34,35). And FAK is associated with cell
apoptosis (36). Thus, the effect
of ANXA1 on these factors was investigated. We observed that Bap
reduced expression of PTEN and FAK, which was recovered by ANXA1
(Fig. 6A and B). Moreover, the
results showed that the enhanced levels of phosphorylation of PI3K
and Akt (p-PI3K/p-Akt) induced by Bap were partly declined by ANXA1
(Fig. 6C and D).
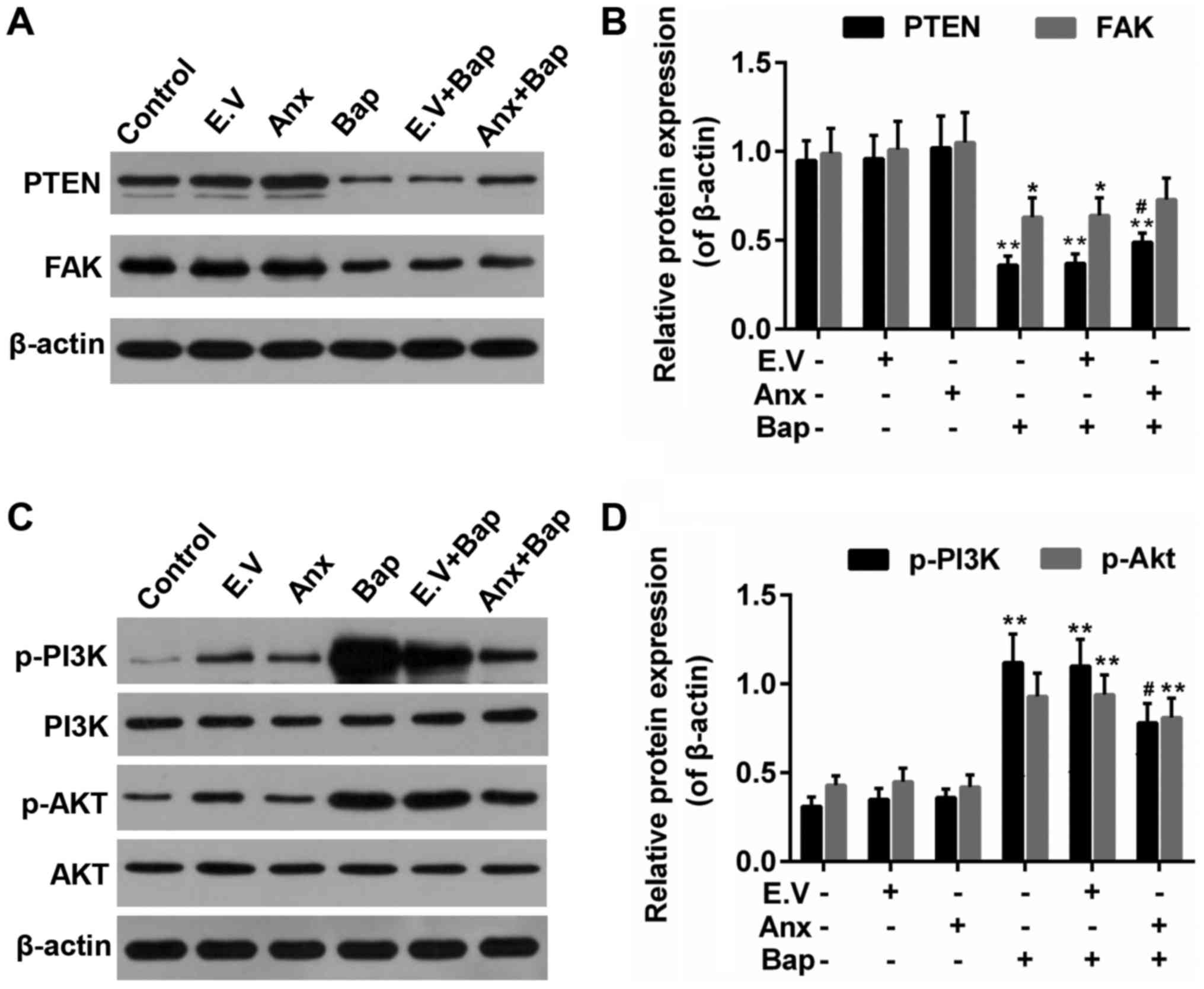 | Figure 6.Effect of ANXA1 on the activities of
PTEN, FAK and PI3K/Akt. (A) Western blotting was used to determine
the protein levels of (B) PTEN and FAK. (C) Western blotting was
also performed to determine the protein levels of (D) PI3K/Akt,
p-PI3K and p-Akt. The cells were treated with 64 µM Bap for 6 h to
establish the cell injury model. *P<0.05 and **P<0.01 vs.
control group; #P<0.05 vs. E.V.+Bap group. ANXA1/Anx,
Annexin A1; PTEN, phosphatase and tensin homolog; FAK, focal
adhesion kinase; PI3K, phosphatidylinositol 3-kinase; Akt, protein
kinase B; p-, phosphorylated; Bap, benzo[a]pyrene; E.V., empty
vector. |
Inhibition of PTEN reversed the
anti-apoptotic effect of ANXA1
Subsequently, an inhibitor of PTEN-SF1670, was
adopted to further confirm the role of PTEN in the anti-apoptotic
effect that conferred by ANXA1. The results showed that the
apoptosis mediated by Bap was mitigated by over-expression of
ANXA1, whereas SF1670 reversed the anti-apoptotic effect of ANXA1
(Fig. 7A). Moreover, the WB data
indicated that the enhanced expression of PTEN and FAK was
decreased by SF1670. And the decreased expression of p-PI3K/p-Akt
was augmented by SF1670 (Fig. 7B).
The work model relate to the current study was shown in Fig. 8.
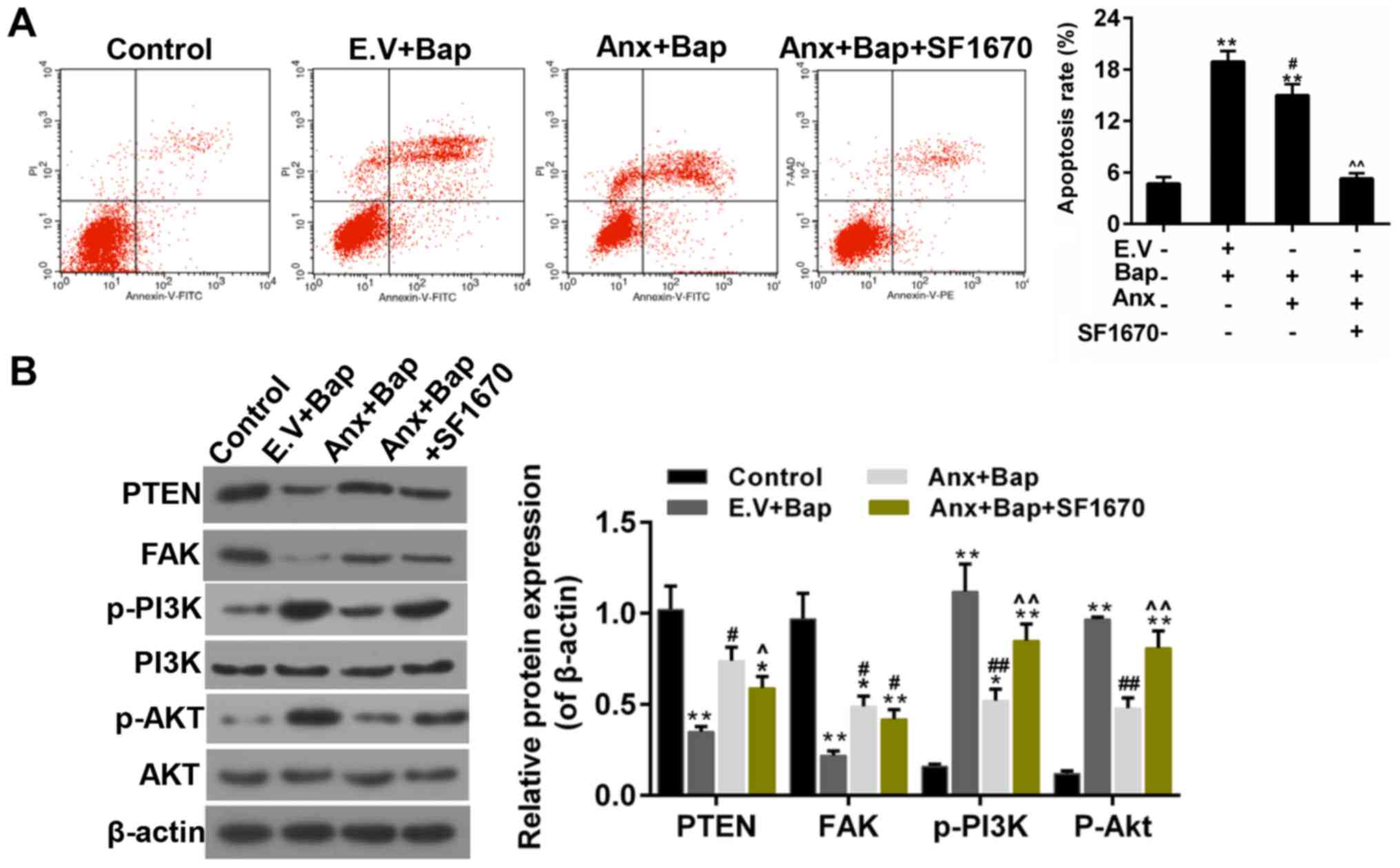 | Figure 7.SF1670 reverses the effect of ANXA1.
(A) The levels of apoptosis were estimated by flow cytometry assay.
(B) Western blotting was performed to determine the expressions of
PTEN, FAK, PI3K/Akt, p-PI3K and p-Akt. The cells were treated with
64 µM Bap for 6 h to establish the cell injury model. *P<0.05
and **P<0.01 vs. control group; #P<0.05 and
##P<0.01 vs. E.V.+Bap group; ^P<0.05, ^^P<0.01
vs. ANXA1+BaP group. ANXA1/Anx, Annexin A1; PTEN, phosphatase and
tensin homolog; FAK, focal adhesion kinase; PI3K,
phosphatidylinositol 3-kinase; Akt, protein kinase B; p-,
phosphorylated; Bap, benzo[a]pyrene; E.V., empty vector. |
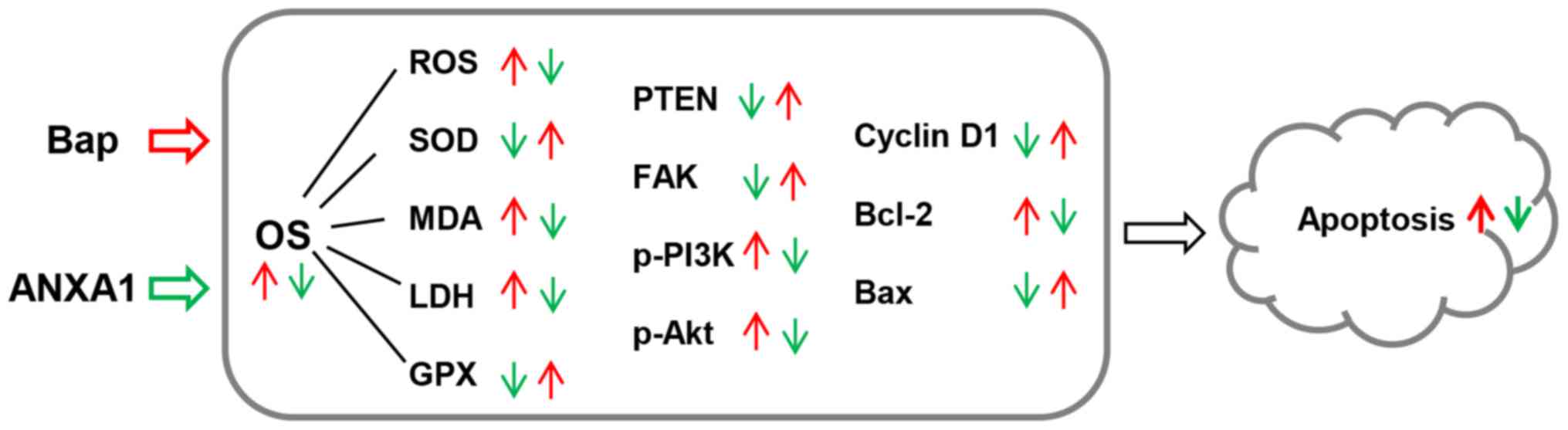 | Figure 8.The model applied in the present
study. Red arrows indicate the effect of Bap, and green arrows
indicate the effect of ANXA1. Bap, benzo[a]pyrene; OS, oxidative
stress; ANXA1, Annexin A1; PTEN, phosphatase and tensin homolog;
FAK, focal adhesion kinase; PI3K, phosphatidylinositol 3-kinase;
Akt, protein kinase B; p-, phosphorylated; SOD, superoxide
dismutase; GPX, glutathione peroxidases; MDA, malondialdehyde; LDH,
lactic dehydrogenase; ROS, reactive oxygen species; Bcl-2, B-cell
lymphoma 2; Bax, Bcl-2-associated X protein. |
Discussion
In environmental pollutants and cigarette smoke,
many toxic agents, of which Bap is a type, produce harmful effects
on human beings, especially on the respiratory tract (37,38).
Bap was employed to induce the bronchial epithelium injury in this
study. Consistent with the previous study, it was obvious that the
cytotoxic effect of Bap was augmented with the increasing of
concentration and the progress of incubation time. This study
investigated oxidative stress and apoptosis in human bronchial
epithelial cells during the incubation with Bap and the expression
of AnxA1. The data suggested that the ANXA1 exerted a protective
role in Bap-induced bronchial epithelium injury. This outcome was
in line with the increased histopathologic mucosal injury exhibited
in AnxA1-deficient animals (39).
Several mechanisms have been put forward to
interrupt the cell injury in airways, among which intracellular
induction of ROS is a type. ROS are molecules, which have a strong
cytotoxic capacity. Oxidative stress occurs when the ROS accumulate
in living cells (12). Our data
showed that the incubation of Bap remarkably increased the ROS
formation in bronchial epithelial cells and that the ROS level was
declined by the ANXA1 over-expression. Oxidative stress is caused
by oxidant/antioxidant disorder. The over-generation of ROS will
lead to enzymatic and non-enzymatic alterations and the
dys-regulation of respiratory airways (40). Our data recorded an increased
activity of SOD and Gpx as well as a decreased content of MDA and
LDH. Since the imbalance of oxidant/antioxidant enzyme was
ameliorated by ANXA1, our research data were in accordance to the
results from a previous study, in which the oxidative inactivation
and the mucosal wound repair was shown to be promoted by ANXA1 in
mucosal wound repair (41).
Activation of the apoptosis pathway is closely
related to increased ROS production (42). The cellular caspase-3 activity was
enhanced subsequent to being incubated with Bap, following
oxidative stress in this study. Moreover, the apoptosis was reduced
by the over-expression of ANXA1 via decreasing the expression Bcl-2
and increasing the expression of Bax and cyclin D1. It has been
previously suggested that ANXA1 exerted its protective role by
repressing the apoptosis. Consistently, it was believed that
excessive apoptosis is another mechanism of cell injury due to
oxidative stress in airway epithelial cells (43).
PTEN, as a well-known tumor suppressor, can
counterbalance the activation of PI3K/Akt (44). The important role of PTEN in
resistance to lung cell injury has been demonstrated before
(45). FAK is associated with both
PTEN and PI3K/Akt based on current knowledge (46). To illustrate the molecular
mechanism, the activity of PTEN, FAK, PI3K/Akt was examined. Our
data indicated that PTEN and FAK were deactivated by Bap, while
their expressions were rescued by the ANXA1 over-expression. A
previous study have shown that the antisense oligonucleotides of
FAK could depress cell viability (47). Thus, in this study, the
anti-apoptotic role of FAK was recognized once more. Furthermore,
the hyperactivation of PI3K/Akt caused by Bap was partially blunted
by ANXA1. This result was compatible with a publication in which
the inactivation of Akt definitively ameliorated ALI and airway
remodelling, and retained the integrity of alveolar epithelial
cells (17). Nevertheless,
researches also pointed out that PI3K-Akt signaling pathway was
related to anti-apoptotic functions (48,49).
These results appeared to be controversial. However, it is not
surprising that PI3K-Akt signaling pathway may exert different
roles in the cells, and this may ascribe to different cell type and
different study model. In addition, the specific inhibition of PTEN
that caused by SF1670 reversed the anti-apoptotic effect of ANXA1,
and such a phenomenon suggested that the activation of PTEN was
necessary for the protective effect of ANXA1. Moreover, SF1670
reversed the effect of ANXA1 on the expression of PTEN, FAK and
p-PI3K/p-Akt. All these data showed that PTEN/FAK/PI3K/Akt pathway
was the downstream signaling of ANXA1, and this pathway was
participated in Bap induced apoptosis.
Taken together, as the work model shown, the
over-expression of ANXA1 suppressed Bap-induced oxidative stress
and blunted activation of PI3K/Akt. ROS play important roles in the
signal transduction pathways that regulate cell proliferation
(50,51). It was implied that the
anti-apoptotic role of ANXA1 may ascribe in part to a
redox-dependent mechanism (52).
Thus, ANXA1 might have the function to prevent bronchial epithelium
injury. In addition, if the protective role of ANXA1 can be
confirmed by in vivo studies, ANXA1 may have promising
therapeutic benefits for the option of asthma.
In summary, our results demonstrated that ANXA1
could sharply reduce the oxidative stress by declining ROS extent
and modulating the level of SOD, Gpx, MDA and LDH. Additionally,
ANXA1 inhibited apoptosis via regulating the expression of Bcl-2,
Bax and cyclin D1. The underlying mechanisms is most likely to be
associated with the the activation of PTEN/FAK and the inhibition
of p-PI3K/p-Akt. The current study inspired a potential therapeutic
strategy for asthma patients.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated and/or analyzed during this study
are included in this published article.
Authors' contributions
YC wrote the main manuscript. YC and SY designed the
study and performed the experiments. YC performed data analysis.
All authors read and approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Anandan C, Nurmatov U, van Schayck OC and
Sheikh A: Is the prevalence of asthma declining? Systematic review
of epidemiological studies. Allergy. 65:152–167. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Worldwide variation in prevalence of
symptoms of asthma, allergic rhinoconjunctivitis, and atopic
eczema: ISAAC. The International Study of Asthma and Allergies in
Childhood (ISAAC) Steering Committee. Lancet. 351:1225–1232. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kay AB, Phipps S and Robinson DS: A role
for eosinophils in airway remodelling in asthma. Trends Immunol.
25:477–482. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Capra V and Rovati GE: Rosuvastatin
inhibits human airway smooth muscle cells mitogenic response to
eicosanoid contractile agents. Pulm Pharmacol Ther. 27:10–16. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Al-Muhsen S, Johnson JR and Hamid Q:
Remodeling in asthma. J Allergy Clin Immunol. 128:451–464. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Uno S and Makishima M: Benzo[a]pyrene
toxicity and inflammatory disease. Curr Rheumatol Rev. 5:266–271.
2009. View Article : Google Scholar
|
|
7
|
Dong JI and Bozzelli JW: Benzo(a)pyrene
levels in several indoor environments with kerosene heaters and
wood-burning fireplaces. Chemosphere. 18:1829–1836. 1989.
View Article : Google Scholar
|
|
8
|
Qamar W, Khan R, Khan AQ, Rehman MU,
Lateef A, Tahir M, Ali F and Sultana S: Alleviation of lung injury
by glycyrrhizic acid in benzo(a)pyrene exposed rats: Probable role
of soluble epoxide hydrolase and thioredoxin reductase. Toxicology.
291:25–31. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Burhans WC and Heintz NH: The cell cycle
is a redox cycle: Linking phase-specific targets to cell fate. Free
Radic Biol Med. 47:1282–1293. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Michaeloudes C, Sukkar MB, Khorasani NM,
Bhavsar PK and Chung KF: TGF-β regulates Nox4, MnSOD and catalase
expression, and IL-6 release in airway smooth muscle cells. Am J
Physiol Lung Cell Mol Physiol. 300:L295–L304. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kirkham P and Rahman I: Oxidative stress
in asthma and COPD: Antioxidants as a therapeutic strategy.
Pharmacol Ther. 111:476–494. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schieber M and Chandel NS: ROS function in
redox signaling and oxidative stress. Curr Biol. 24:R453–R462.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kannan K and Jain SK: Oxidative stress and
apoptosis. Pathophysiology. 7:153–163. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Keniry M and Parsons R: The role of PTEN
signaling perturbations in cancer and in targeted therapy.
Oncogene. 27:5477–5485. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Frisch SM, Vuori K, Ruoslahti E and
Chan-Hui PY: Control of adhesion-dependent cell survival by focal
adhesion kinase. J Cell Biol. 134:793–799. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tamura M, Gu J, Matsumoto K, Aota S,
Parsons R and Yamada KM: Inhibition of cell migration, spreading,
and focal adhesions by tumor suppressor PTEN. Science.
280:1614–1617. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Miyoshi K, Yanagi S, Kawahara K, Nishio M,
Tsubouchi H, Imazu Y, Koshida R, Matsumoto N, Taguchi A, Yamashita
S, et al: Epithelial Pten controls acute lung injury and fibrosis
by regulating alveolar epithelial cell integrity. Am J Respir Crit
Care Med. 187:262–275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maehama T and Dixon JE: The tumor
suppressor, PTEN/MMAC1, dephosphorylates the lipid second
messenger, phosphatidylinositol 3,4,5-trisphosphate. J Biol Chem.
273:13375–13378. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Schaller MD: Signaling through the focal
adhesion kinase. Soc Gen Physiol Ser. 52:241–255. 1997.PubMed/NCBI
|
|
20
|
Levkau B, Herren B, Koyama H, Ross R and
Raines EW: Caspase-mediated cleavage of focal adhesion kinase
pp125FAK and disassembly of focal adhesions in human endothelial
cell apoptosis. J Exp Med. 187:579–586. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sonoda Y, Watanabe S, Matsumoto Y,
Aizu-Yokota E and Kasahara T: FAK is the upstream signal protein of
the phosphatidylinositol 3-kinase-Akt survival pathway in hydrogen
peroxide-induced apoptosis of a human glioblastoma cell line. J
Biol Chem. 274:10566–10570. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Goulding NJ, Godolphin JL, Sampson MB,
Maddison PJ and Flower RJ: Hydrocortisone induces lipocortin 1
production by peripheral blood mononuclear cells in vivo in man.
Biochem Soc Trans. 18:306–307. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chatterjee BE, Yona S, Rosignoli G, Young
RE, Nourshargh S, Flower RJ and Perretti M: Annexin 1-deficient
neutrophils exhibit enhanced transmigration in vivo and increased
responsiveness in vitro. J Leukoc Biol. 78:639–646. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Perretti M, Ahluwalia A, Harris JG,
Goulding NJ and Flower RJ: Lipocortin-1 fragments inhibit
neutrophil accumulation and neutrophil-dependent edema in the
mouse. A qualitative comparison with an anti-CD11b monoclonal
antibody. J Immunol. 151:4306–4314. 1993.PubMed/NCBI
|
|
25
|
D'Amico M, Di Filippo C, La M, Solito E,
McLean PG, Flower RJ, Oliani SM and Perretti M: Lipocortin 1
reduces myocardial ischemia-reperfusion injury by affecting local
leukocyte recruitment. FASEB J. 14:1867–1869. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
La M, D'Amico M, Bandiera S, Di Filippo C,
Oliani SM, Gavins FN, Flower RJ and Perretti M: Annexin 1 peptides
protect against experimental myocardial ischemia-reperfusion:
Analysis of their mechanism of action. FASEB J. 15:2247–2256. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Martin GR, Perretti M, Flower RJ and
Wallace JL: Annexin-1 modulates repair of gastric mucosal injury.
Am J Physiol Gastrointest Liver Physiol. 294:G764–G769. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Y, Prasad A, Jia Y, Roy SG, Loison F,
Mondal S, Kocjan P, Silberstein LE, Ding S and Luo HR:
Pre-treatment with phosphatase and tensin homolog deleted on
chromosome 10 (PTEN) inhibitor SF1670 augments the efficacy of
granulocyte transfusion in a clinically relevant mouse model.
Blood. 117:6702–6713. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Feng L, Le J, Lan Y, Pang W and Wang C:
The role of ROS in human bronchial epithelial cell injury caused by
gunpowder smoke. Lab Immun Clin Med. 17:27–29. 2010.
|
|
31
|
Chandra J, Samali A and Orrenius S:
Triggering and modulation of apoptosis by oxidative stress. Free
Radic Biol Med. 29:323–333. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sedlak TW, Oltvai ZN, Yang E, Wang K,
Boise LH, Thompson CB and Korsmeyer SJ: Multiple Bcl-2 family
members demonstrate selective dimerizations with Bax. Proc Natl
Acad Sci USA. 92:7834–7838. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Albanese C, D'Amico M, Reutens AT, Fu M,
Watanabe G, Lee RJ, Kitsis RN, Henglein B, Avantaggiati M,
Somasundaram K, et al: Activation of the cyclin D1 gene by the
E1A-associated protein p300 through AP-1 inhibits cellular
apoptosis. J Biol Chem. 274:34186–34195. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xia H, Diebold D, Nho R, Perlman D,
Kleidon J, Kahm J, Avdulov S, Peterson M, Nerva J, Bitterman P and
Henke C: Pathological integrin signaling enhances proliferation of
primary lung fibroblasts from patients with idiopathic pulmonary
fibrosis. J Exp Med. 205:1659–1672. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
White ES, Atrasz RG, Hu B, Phan SH,
Stambolic V, Mak TW, Hogaboam CM, Flaherty KR, Martinez FJ, Kontos
CD and Toews GB: Negative regulation of myofibroblast
differentiation by PTEN (Phosphatase and Tensin Homolog Deleted on
chromosome 10). Am J Respir Crit Care Med. 173:112–121. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sonoda Y, Matsumoto Y, Funakoshi M,
Yamamoto D, Hanks SK and Kasahara T: Anti-apoptotic role of Focal
Adhesion Kinase (FAK). Induction of inhibitor-of-apoptosis proteins
and apoptosis suppression by the overexpression of FAK in a human
leukemic cell line, HL-60. J Biol Chem. 275:16309–16315. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Alexandrov K, Rojas M and Satarug S: The
critical DNA damage by benzo(a)pyrene in lung tissues of smokers
and approaches to preventing its formation. Toxicol Lett.
198:63–68. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Podechard N, Lecureur V, Le Ferrec E,
Guenon I, Sparfel L, Gilot D, Gordon JR, Lagente V and Fardel O:
Interleukin-8 induction by the environmental contaminant
benzo(a)pyrene is aryl hydrocarbon receptor-dependent and leads to
lung inflammation. Toxicol Lett. 177:130–137. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Babbin BA, Laukoetter MG, Nava P, Koch S,
Lee WY, Capaldo CT, Peatman E, Severson EA, Flower RJ, Perretti M,
et al: Annexin A1 regulates intestinal mucosal injury,
inflammation, and repair. J Immunol. 181:5035–5044. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nadeem A, Masood A and Siddiqui N:
Oxidant-antioxidant imbalance in asthma: Scientific evidence,
epidemiological data and possible therapeutic options. Ther Adv
Respir Dis. 2:215–235. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Leoni G, Alam A, Neumann PA, Lambeth JD,
Cheng G, McCoy J, Hilgarth RS, Kundu K, Murthy N, Kusters D, et al:
Annexin A1, formyl peptide receptor, and NOX1 orchestrate
epithelial repair. J Clin Invest. 123:443–454. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park EJ, Yi J, Chung KH, Ryu DY, Choi J
and Park K: Oxidative stress and apoptosis induced by titanium
dioxide nanoparticles in cultured BEAS-2B cells. Toxicol Lett.
180:222–229. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Truong-Tran AQ, Grosser D, Ruffin RE,
Murgia C and Zalewski PD: Apoptosis in the normal and inflamed
airway epithelium: Role of zinc in epithelial protection and
procaspase-3 regulation. Biochem Pharmacol. 66:1459–1468. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Stambolic V, Suzuki A, de la Pompa JL,
Brothers GM, Mirtsos C, Sasaki T, Ruland J, Penninger JM,
Siderovski DP and Mak TW: Negative regulation of PKB/Akt-dependent
cell survival by the tumor suppressor PTEN. Cell. 95:29–39. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tiozzo C, De Langhe S, Yu M, Londhe VA,
Carraro G, Li M, Li C, Xing Y, Anderson S, Borok Z, et al: Deletion
of Pten expands lung epithelial progenitor pools and confers
resistance to airway injury. Am J Respir Crit Care Med.
180:701–712. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
You D, Xin J, Volk A, Wei W, Schmidt R,
Scurti G, Nand S, Breuer EK, Kuo PC, Breslin P, et al: FAK mediates
a compensatory survival signal parallel to PI3K-AKT in PTEN-null
T-ALL cells. Cell Rep. 10:2055–2068. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sonoda Y, Kasahara T, Yokota-Aizu E, Ueno
M and Watanabe S: A suppressive role of p125FAK protein tyrosine
kinase in hydrogen peroxide-induced apoptosis of T98G cells.
Biochem Biophys Res Commun. 241:769–774. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wu W: DLX4 homeoprotein promotes PI3K/Akt
anti-apoptosis pathway in ER negative breast cancer cells through
upregulation of VEGFA. Dissert Thes-Gradwork. 2010.
|
|
49
|
Wang XQ, Yao RQ, Liu X, Huang JJ, Qi DS
and Yang LH: Quercetin protects oligodendrocyte precursor cells
from oxygen/glucose deprivation injury in vitro via the activation
of the PI3K/Akt signaling pathway. Brain Res Bull. 86:277–284.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Qin S, Inazu T, Takata M, Kurosaki T,
Homma Y and Yamamura H: Cooperation of tyrosine kinases p72syk and
p53/56lyn regulates calcium mobilization in chicken B cell oxidant
stress signaling. Eur J Biochem. 236:443–449. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Suzuki Y, Ohsugi K and Ono Y: Oxidative
stress triggers tyrosine phosphorylation in B cells through a
redox- and inflammatory cytokine-sensitive mechanism. Immunology.
87:396–401. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sturrock A, Huecksteadt TP, Norman K,
Sanders K, Murphy TM, Chitano P, Wilson K, Hoidal JR and Kennedy
TP: Nox4 mediates TGF-beta1-induced retinoblastoma protein
phosphorylation, proliferation, and hypertrophy in human airway
smooth muscle cells. Am J Physiol Lung Cell Mol Physiol.
292:L1543–L1555. 2007. View Article : Google Scholar : PubMed/NCBI
|