Introduction
Although the medical approaches have developed
rapidly, acute myocardial infarction (AMI) remains a principal
cause of mortality worldwide (1).
AMI induces a complex pathological processes including the death of
cardiomyocytes, inflammation, fibrosis and hypertrophic growth of
cardiomyocytes, which may progress to heart failure (2). Currently, the most effective
therapies for AMI are vascular reflow to the ischemic region by
thrombolysis, percutaneous coronary intervention and coronary
artery bypass surgery (3).
As the ischemic area remains hypoxic, the heart
continues to lose cardiomyocytes, which decreases its contractile
function. The condition can be modeled by the permanent ligation of
coronary artery in rodent animals. When the myocardium is
reperfused for the reoxygenation of ischemic myocardium, the
reoxygenation can induce reperfusion injury through cell injury
pathways, including free radical production, mitochondrial injury,
activation of pro-apoptotic pathway and calcium dysregulation
(4). Reoxygenation also induces
cardiac remodeling by the transdifferentiation of cardiac
fibroblasts into myofibroblasts (4–6).
These side effects of reperfusion may increase the risk of
mortality (7,8). Therefore, a comprehensive
understanding of myocardial biology in ischemia-reperfusion (IR) is
important.
Weighted gene co-expression network analysis (WGCNA)
has been successfully used in systems biology studies to
investigate the intrinsic organization of transcriptomes (9). This approach provides an effective
method to examine the gene expression patterns, calculate the
adjacency of genes, construct gene networks and assess the
importance of genes within the network (10). It has been applied for construction
of gene co-expression network modules, the identification of hub
genes and identifying the connection of diagnostic genes and
diseases (10–12). However, WGCNA has not been applied
to identify the network and hub genes associated with IR myocardium
from days 2 to 7, to the best of the authors' knowledge.
In the present study, the microarray data on IR
remodeling were downloaded from National Center for Biotechnology
Information (NCBI) (13) and
differentially expressed genes (DEGs) identified by pairwise
comparison. Subsequently, WGCNA was performed to investigate the
expression modules and key genes for each module. The functional
annotation of each module revealed the biological associations with
dynamic features of myocardium from days 2 to 7.
Materials and methods
Analysis of differentially expressed
genes
The microarray data of rat IR were downloaded from
the NCBI Gene Expression Omnibus database (accession number:
GSE4105) and normalized by MAS5 (13). All the samples were divided into 4
groups, including day 2 (2d)-sham, 2d-IR, day 7 (7d)-sham and
7d-IR. DEGs were analyzed between two subsequent groups. DEGs
between any two consecutive stages were determined by the limma
3.34.9 package in R (version 3.1.1) (14). The threshold of the significance of
DEG was set as fold change 31.5 and P<0.05. A total of 3,321
DEGs were obtained with the cutoff set as P<0.05.
Construction of gene co-expression
networks
To study the functional organization of the rat IR
heart, DEGs between any two consecutive stages were selected and
weighted co-expression network analysis performed by using the
WGCNA R package (15). First,
missing values and outlier samples were checked by an unsupervised
hierarchical clustering analysis. Then, a weighted adjacency matrix
was created and the soft threshold power (β) set at 10 to analyze
scale-free topology. The power β determines the co-expression
similarity for the adjacency matrix. The power 10 was chosen, for
which the scale-free topology fitting index R2 and mean
connectivity may reach a plateau (Fig.
1). It suggested that β=10 was the minimum value for the
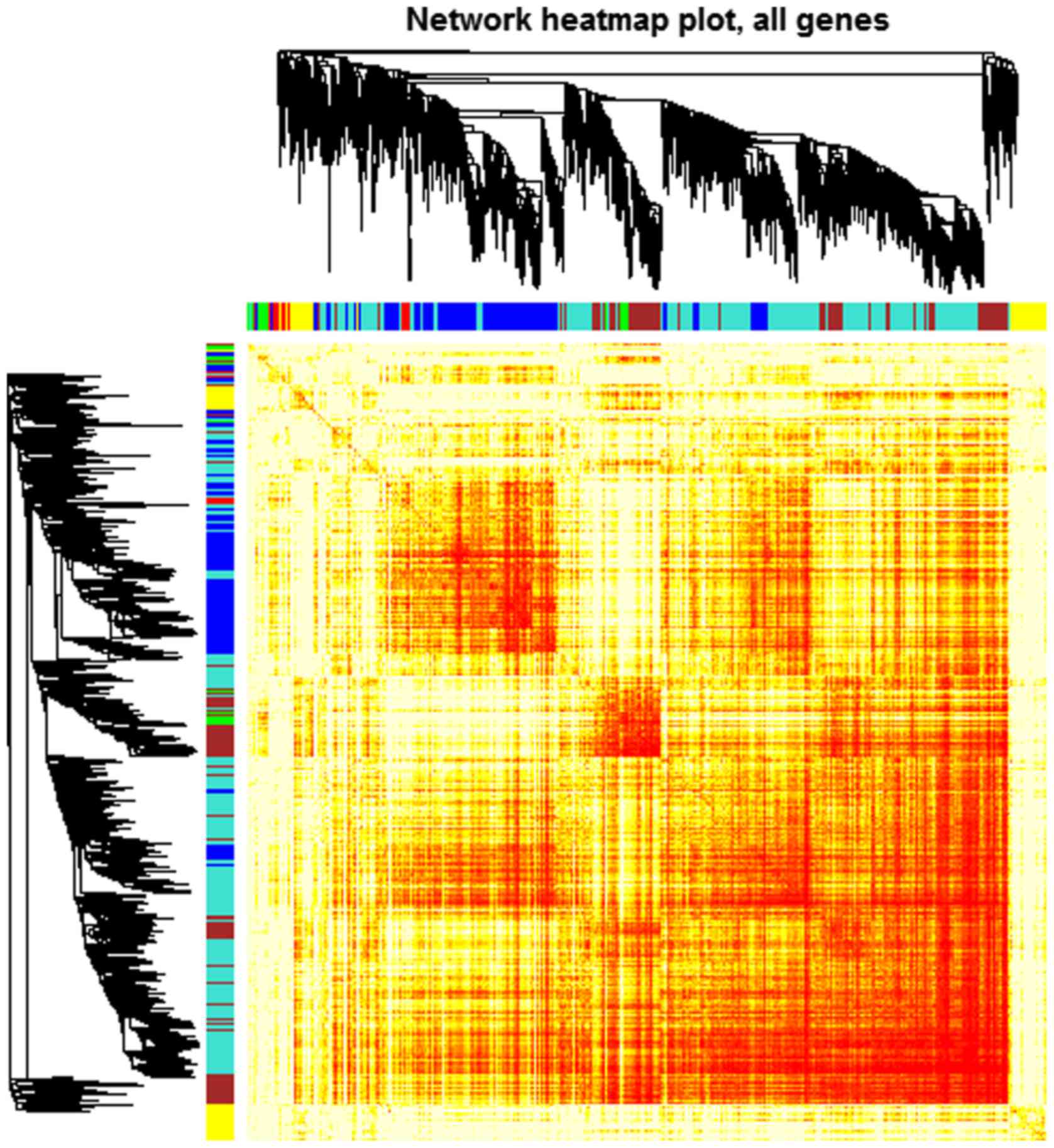
scale-free topology criterion. Modules were identified by the
following parameters: ‘power =10, minModuleSize =30, mergeCutHeight
=0.25’. A total of 6 modules containing all the DEGs were
identified. Genes of each module are demonstrated in the
topological overlap heatmap (Fig.
2).
Modules were labeled using different colors and
shown as a hierarchical clustering dendrogram. The weighted network
was visualized using a heatmap plot. Gene expression patterns of
DEGs in each module were analyzed by hierarchical clustering
analysis.
Gene Ontology (GO) and pathway
analysis
To identify the biological process and signaling
pathways in each module, GO (16)
and Kyoto Encyclopedia of Genes and Genomes (KEGG) (17) pathways were enriched by David
online tools (david.ncifcrf.gov) (18,19).
The threshold of significant enrichment was set as P<0.05.
Protein-protein interaction (PPI)
network analysis
To determine the interaction between DEGs in each
module, they were mapped to Search Tool for the Retrieval of
Interacting Genes (STRING; version 10.5), an online tool for
evaluating protein-protein interaction (20). Interactions with a combined score
>0.4 were considered significant. The protein-protein networks
were constructed by Cytoscape software (version 3.2.0; http://www.cytoscape.org/) (21). Modules of protein-protein
interaction were identified by Molecular Complex Detection (MCODE;
version 1.5) (22) with the
following cutoff: Nodes of cluster >4, Score of MCODE >3.
Results
Functional annotation of DEGs in each
module
Significantly enriched biological process and KEGG
pathways in each module are shown in Fig. 3. Different modules represented
distinct gene expression patterns following IR. The blue module
denoted the DEGs enriched between 7d-IR and other groups (Fig. 3A). ‘Extracellular matrix
organization’, ‘wound healing’, ‘angiogenesis’, ‘canonical Wnt
signaling’ and ‘PI3K-AKT signaling’ were significantly enriched in
this module (Figs. 4A and 5A). In the brown module, DEGs were
enriched between IR and sham group (Fig. 3B). ‘Regulation of cell
proliferation’, ‘regulation of cell shape’, ‘intergrin mediated
signaling pathway’, ‘endocytosis’, ‘Fc gamma R-mediated
phagocytosis’, ‘cardiac contraction’, ‘hypertrophic
cardiomyopathy’, ‘adrenergic signaling’ and ‘sphingolipid signaling
pathway’ were significantly enriched in this module (Figs. 4C and 5C). In the green module, DEGs were
enriched between 2d-IR and other groups (Fig. 3C). ‘Response to organic cyclic
compound’, ‘epithelial cell proliferation’, ‘response to wounding’,
and ‘amino acid metabolism’ were enriched significantly in this
module (Figs. 4D and 5D). In the red module, DEGs were enriched
between days 2 and 7 (Fig. 3D).
‘Chromatin remodeling’, ‘neural tube closure’, and ‘neural tube
formation’ were enriched significantly in this module (Fig. 4F). In the turquoise module, the
dynamic pattern of DEGs is shown in Fig. 3E. ‘Fatty acid β-oxidation’,
‘tricarboxylic acid cycle’, ‘ATP metabolic process’ and ‘oxidative
phosphorylation’ were enriched significantly in this module
(Figs. 4E and 5E). In the yellow module, DEGs were
enriched between sham-7 day and other groups (Fig. 3F). ‘Ion transmembrane transport’,
‘regulation of body fluid levels’, ‘adrenergic signaling’ and
‘cGMP-PKG signaling’ pathways were significantly enriched (Figs. 4B and 5B).
Identification of the key genes in
each module
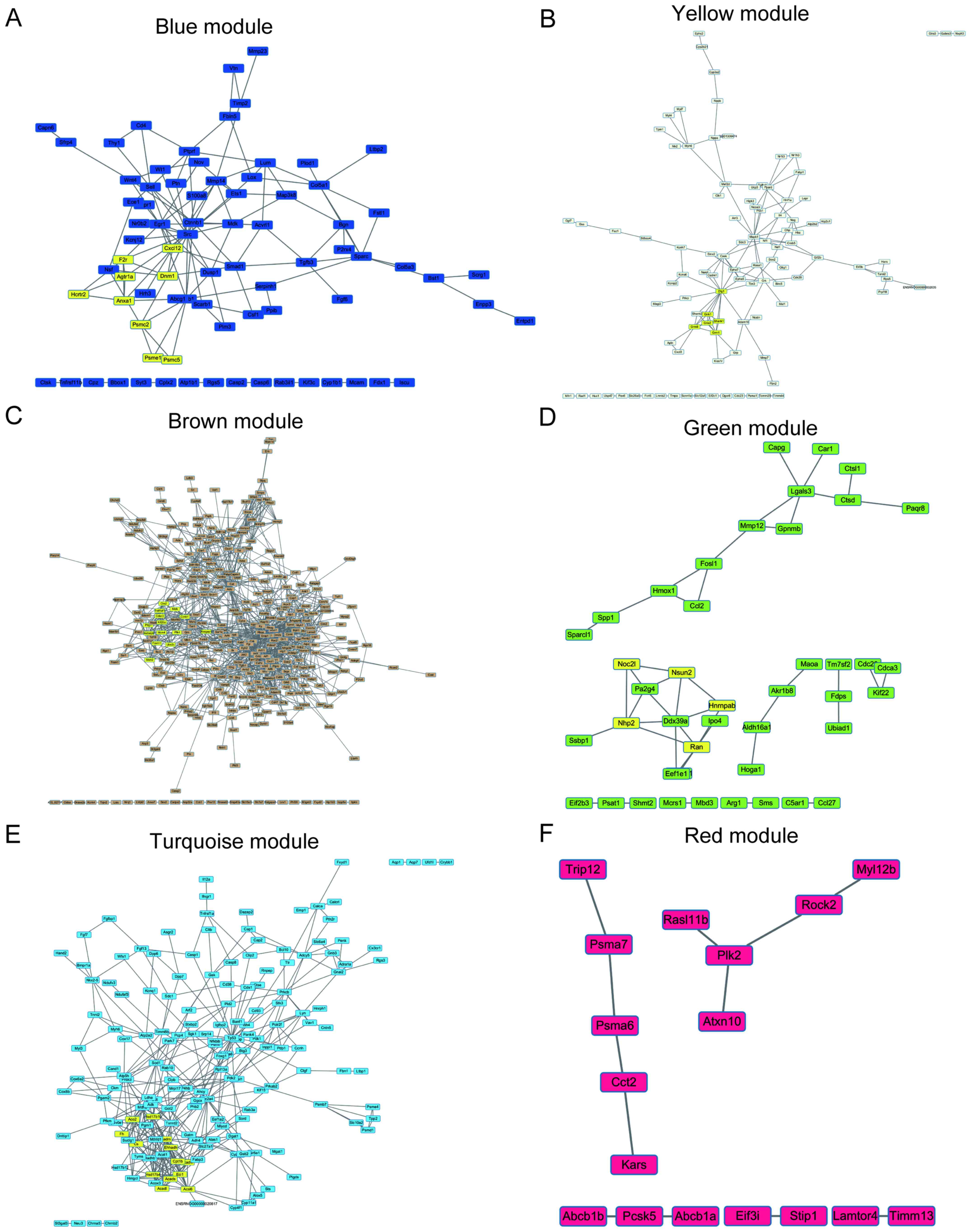
To elucidate the interaction between the hub genes
in each module, PPI networks of hub genes were constructed using
Cytoscape software. To explore core modules of protein-protein
interactions, PPI modules were identified by MCODE.
The protein interacting with maximum number of DEGs
was defined as the key gene in each module. As shown in Fig. 6, the key genes of each module are
SRC proto-oncogene, non-receptor tyrosine kinase (Scr) for
the blue module, discs large MAGUK scaffold protein 1 (Dlg1)
for the yellow module, ATP citrate lyase (Acly) for the
brown module, RAN, member RAS oncogene family (Ran) for the
green module, tumor protein p53 (TP53) for the turquoise
module and polo like kinase 2 (Plk2) for the red module. The
most significant cluster of each network was highlighted for yellow
color in each module.
Discussion
The aim of the present study was to identify the
genes and networks associated with the heart remodeling process
following IR. Distinct modules, biological process, KEGG pathways
and co-expression networks during heart remodeling were identified
and the results of the present study provided insights into the
regulation of heart remodeling following IR.
A total of 6 modules were identified by constructing
co-expression networks. Hierarchical clustering analysis revealed
that each module represented a different dynamic gene expression
pattern and a different biological function. The blue module
represented the specific physiological features of the heart
following 7 days' IR, associated with ‘canonical Wnt signaling’,
‘TGFβ receptor signaling’ and ‘PI3K-AKT signaling pathway’. It has
been reported that canonical Wnt signaling, TGFβ and PI3K-AKT
signaling pathways are involved in regulation of cardiac
fibroblasts growth and extracellular matrix (ECM) remodeling
(23–26).
The brown module represented the common
physiological features of the heart following IR, which are
associated with phagocytosis. The green module represented the
specific physiology of the heart after 2 days' IR, associated with
amino acid metabolism. Apoptosis-associated genes were
significantly upregulated in the day 2 IR heart, including heme
oxygenase 1, FOS like 1, AP-1 transcription factor subunit and
eukaryotic translation elongation factor 1 epsilon 1. The red
module represented the physiological state of the heart between
days 2 and 7, which is associated with neural development. The
turquoise module represented the gradual process of heart
remodeling which is closely associated with fatty acid metabolism.
It has been documented that during heart remodeling and heart
failure, the energy metabolism shifts from the fatty acid
metabolism to glucose metabolism (27). In this module, fatty acid
metabolism genes including acyl-CoA oxidase 3, pristanoyl,
carnitine palmitoyltransferase 1B, acyl-CoA synthetase long chain
family member 6, hydroxyacyl-CoA dehydrogenase and acyl-CoA
dehydrogenase very long chain were gradually and significantly
downregulated. The yellow module represented the specific
physiological state of the normal heart at day 7, which is closely
associated with the cGMP-PKG signaling pathway. The cGMP/PKG
pathway is known to inhibit cardiac structural remodeling (28). All these results suggested heart
remodeling following IR is a complex process, involving ECM, neural
development, apoptosis and energy metabolism shift.
Hub genes were also identified in each module,
including Src for the blue module, Dlg1 for the
yellow module, Acly for the brown module, Ran for the
green module, TP53 for turquoise module and Plk2 for
red module. Src, upregulated in the heart 7 days following
IR, is a non-receptor tyrosine kinase that serves roles in numerous
biological process including cell adhesion, cell cycle and cell
migration (29,30). In the heart, Src is
activated by AngII and contributes to pathophysiology of cardiac
remodeling and hypertrophy (23).
It has been suggested that Src may be a candidate target for
heart remodeling diseases (31).
Dlg1, upregulated in the day 7 normal heart, encodes a
multi-domain scaffolding protein, taking part in skeletogenesis,
cytoskeleton organization and endothelia proliferation (32,33).
Although there have not been any studies on its function in the
heart, to the best of the authors' knowledge, it may be a target to
improve heart function based on its function in other cell types.
Acly, upregulated in the IR heart, is an ATP citrate lyase
which is responsible for the synthesis of cytosolic acetyl-CoA
(34). Acly-mediated
dimethyl α-ketoglutarate prevents autophagy of heart and reduces
heart contractile performance (35). Acly may be an adaptive
response in the IR heart and may be used for the inhibition of
autophagy in the heart. Ran, a member of RAS superfamily,
was upregulated in the day 2 IR heart. Although there have not been
any studies on its function in the heart, to the best of the
authors' knowledge, previous studies have suggested that Ran
controls microtubule organization during the apoptotic process
(36,37). Ran can perform a similar
function in cardiomyocytes (38),
which should to be studied in the future. TP53, gradually
upregulated in the IR heart, is key for heart disease progression.
A number of studies have suggested that TP53 serves an
important role in heart remodeling and heart failure, and is
responsible for suppression of hypoxia inducible factor 1-induced
angiogenesis, apoptosis of cardiomyocytes and fatty acid metabolism
(39–41). Plk2, upregulated in the day
7 heart, is a member of the serine/threonine protein kinases.
Plk2 mediates antioxidant signaling and is essential for
preventing p53-dependent necrotic cell death in cancer cells and so
may possess therapeutic implications for heart diseases (42,43).
Taken together, the results of the current study
revealed the functional modules for heart remodeling following IR
using a WGCNA method and identified key genes in each module by
protein-protein interaction network. The results demonstrated that
heart remodeling following IR is a complex process. A number of
biological processes and signaling pathways are involved. The
results of the present study may provide useful targets for heart
disease treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
Study design: NG, NZ, LY and ZL; data analysis: JW,
FL, YW and XC; manuscript preparation: NG. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
WGCNA
|
weighted gene co-expression network
analysis
|
|
PPI
|
protein-protein interaction
network
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
DEGs
|
differentially expressed genes
|
|
AMI
|
acute myocardial infarction
|
|
ECM
|
extracellular matrix
|
References
|
1
|
GBD 2015 Disease and Injury Incidence and
Prevalence Collaborators: Global, regional, and national incidence,
prevalence, and years lived with disability for 310 diseases and
injuries, 1990–2015: A systematic analysis for the global burden of
disease study 2015. Lancet. 388:1545–1602. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ojha N, Roy S, Radtke J, Simonetti O,
Gnyawali S, Zweier JL, Kuppusamy P and Sen CK: Characterization of
the structural and functional changes in the myocardium following
focal ischemia-reperfusion injury. Am J Physiol Heart Circ Physiol.
294:H2435–H2443. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aversano T, Aversano LT, Passamani E,
Knatterud GL, Terrin ML, Williams DO and Forman SA; Atlantic
Cardiovascular Patient Outcomes Research Team (C-PORT), :
Thrombolytic therapy vs primary percutaneous coronary intervention
for myocardial infarction in patients presenting to hospitals
without on-site cardiac surgery: A randomized controlled trial.
Jama. 287:1943–1951. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sen CK, Khanna S and Roy S: Perceived
hyperoxia: Oxygen-induced remodeling of the reoxygenated heart.
Cardiovasc Res. 71:280–288. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Roy S, Khanna S, Bickerstaff AA,
Subramanian SV, Atalay M, Bierl M, Pendyala S, Levy D, Sharma N,
Venojarvi M, et al: Oxygen sensing by primary cardiac fibroblasts:
A key role of p21(Waf1/Cip1/Sdi1). Circ Res. 92:264–271. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Roy S, Khanna S, Wallace WA, Lappalainen
J, Rink C, Cardounel AJ, Zweier JL and Sen CK: Characterization of
perceived hyperoxia in isolated primary cardiac fibroblasts and in
the reoxygenated heart. J Biol Chem. 278:47129–47135. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McNamara MT and Higgins CB: Magnetic
resonance imaging of chronic myocardial infarcts in man. AJR Am J
Roentgenol. 146:315–320. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Orn S, Manhenke C, Anand IS, Squire I,
Nagel E, Edvardsen T and Dickstein K: Effect of left ventricular
scar size, location, and transmurality on left ventricular
remodeling with healed myocardial infarction. Am J Cardiol.
99:1109–1114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang B and Horvath S: A general framework
for weighted gene co-expression network analysis. Stat Appl Genet
Mol Biol. 4:172005. View Article : Google Scholar
|
|
10
|
Maschietto M, Tahira AC, Puga R, Lima L,
Mariani D, Bda Paulsen S, Belmonte-de-Abreu P, Vieira H, Krepischi
AC, Carraro DM, et al: Co-expression network of
neural-differentiation genes shows specific pattern in
schizophrenia. BMC Med Genomics. 8:232015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo Y and Xing Y: Weighted gene
co-expression network analysis of pneumocytes under exposure to a
carcinogenic dose of chloroprene. Life Sci. 151:339–347. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tian F, Zhao J, Fan X and Kang Z: Weighted
gene co-expression network analysis in identification of
metastasis-related genes of lung squamous cell carcinoma based on
the cancer genome atlas database. J Thorac Dis. 9:42–53. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Roy S, Khanna S, Kuhn DE, Rink C, Williams
WT, Zweier JL and Sen CK: Transcriptome analysis of the
ischemia-reperfused remodeling myocardium: Temporal changes in
inflammation and extracellular matrix. Physiol Genomics.
25:364–374. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: Limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
da Huang W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
da Huang W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Szklarczyk D, Morris JH, Cook H, Kuhn M,
Wyder S, Simonovic M, Santos A, Doncheva NT, Roth A, Bork P, et al:
The STRING database in 2017: Quality-controlled protein-protein
association networks, made broadly accessible. Nucleic Acids Res.
45:D362–D368. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Haendeler J and Berk BC: Angiotensin II
mediated signal transduction. Important role of tyrosine kinases.
Regul Pept. 95:1–7. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
McMullen JR, Shioi T, Zhang L, Tarnavski
O, Sherwood MC, Kang PM and Izumo S: Phosphoinositide
3-kinase(p110alpha) plays a critical role for the induction of
physiological, but not pathological, cardiac hypertrophy. Proc Natl
Acad Sci USA. 100:12355–12360. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Raaf L, Noll C, Mel Cherifi H, Samuel JL,
Delcayre C, Delabar JM, Benazzoug Y and Janel N: Myocardial
fibrosis and TGFB expression in hyperhomocysteinemic rats. Mol Cell
Biochem. 347:63–70. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shanaki M, Hossein-Nezhad A, Meshkani R,
Beigy M, Shirzad M, Pasalar P and Golmohammadi T: Effects of
resveratrol on crosstalk between canonical B-Catenin/Wnt and FOXO
pathways in coronary artery disease patients with metabolic
syndrome: A case control study. Iran J Pharm Res. 15:547–559.
2016.PubMed/NCBI
|
|
27
|
Mueller-Hennessen M, Sigl J, Fuhrmann JC,
Witt H, Reszka R, Schmitz O, Kastler J, Fischer JJ, Müller OJ,
Giannitsis E, et al: Metabolic profiles in heart failure due to
non-ischemic cardiomyopathy at rest and under exercise. ESC Heart
Fail. 4:178–189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen J, Wang D, Wang F, Shi S, Chen Y,
Yang B, Tang Y and Huang C: Exendin-4 inhibits structural
remodeling and improves Ca2+ homeostasis in rats with
heart failure via the GLP-1 receptor through the eNOS/cGMP/PKG
pathway. Peptides. 90:69–77. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ziehr J, Sheibani N and Sorenson CM:
Alterations in cell-adhesive and migratory properties of proximal
tubule and collecting duct cells from bcl-2 -/- mice. Am J Physiol
Renal Physiol. 287:F1154–F1163. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chetty S, Engquist EN, Mehanna E, Lui KO,
Tsankov AM and Melton DA: A Src inhibitor regulates the cell cycle
of human pluripotent stem cells and improves directed
differentiation. J Cell Biol. 210:1257–1268. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pandey P, Hawkes W, Hu J, Megone WV,
Gautrot J, Anilkumar N, Zhang M, Hirvonen L, Cox S, Ehler E, et al:
Cardiomyocytes sense matrix rigidity through a combination of
muscle and non-muscle myosin contractions. Dev Cell. 44(326–336):
e3232018.
|
|
32
|
Laprise P, Viel A and Rivard N: Human
homolog of disc-large is required for adherens junction assembly
and differentiation of human intestinal epithelial cells. J Biol
Chem. 279:10157–10166. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rivera C, Simonson SJ, Yamben IF, Shatadal
S, Nguyen MM, Beurg M, Lambert PF and Griep AE: Requirement for
Dlgh-1 in planar cell polarity and skeletogenesis during vertebrate
development. PLoS One. 8:e544102013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Elshourbagy NA, Near JC, Kmetz PJ, Wells
TN, Groot PH, Saxty BA, Hughes SA, Franklin M and Gloger IS:
Cloning and expression of a human ATP-citrate lyase cDNA. Eur J
Biochem. 204:491–499. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mariño G, Pietrocola F, Kong Y, Eisenberg
T, Hill JA, Madeo F and Kroemer G: Dimethyl α-ketoglutarate
inhibits maladaptive autophagy in pressure overload-induced
cardiomyopathy. Autophagy. 10:930–932. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dallol A, Hesson LB, Matallanas D, Cooper
WN, O'Neill E, Maher ER, Kolch W and Latif F: RAN GTPase is a
RASSF1A effector involved in controlling microtubule organization.
Curr Biol. 19:1227–1232. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Moss DK, Wilde A and Lane JD: Dynamic
release of nuclear RanGTP triggers TPX2-dependent microtubule
assembly during the apoptotic execution phase. J Cell Sci.
122:644–655. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang X, Zhang D, Zhang H, Huang Y and
Teng M: Corrigendum: Role of Ran-regulated nuclear-cytoplasmic
trafficking of pVHL in the regulation of microtubular
stability-mediated HIF-1α in hypoxic cardiomyocytes. Sci Rep.
5:113072015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fujita T and Ishikawa Y: Apoptosis in
heart failure. -The role of the β-adrenergic receptor-mediated
signaling pathway and p53-mediated signaling pathway in the
apoptosis of cardiomyocytes. Circ J. 75:1811–1818. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Oka T, Morita H and Komuro I: Novel
molecular mechanisms and regeneration therapy for heart failure. J
Mol Cell Cardiol. 92:46–51. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Teng H, Sui X, Zhou C, Shen C, Yang Y,
Zhang P, Guo X and Huo R: Fatty acid degradation plays an essential
role in proliferation of mouse female primordial germ cells via the
p53-dependent cell cycle regulation. Cell Cycle. 15:425–431. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li J, Ma W, Wang PY, Hurley PJ, Bunz F and
Hwang PM: Polo-like kinase 2 activates an antioxidant pathway to
promote the survival of cells with mitochondrial dysfunction. Free
Radic Biol Med. 73:270–277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Matsumoto T, Wang PY, Ma W, Sung HJ,
Matoba S and Hwang PM: Polo-like kinases mediate cell survival in
mitochondrial dysfunction. Proc Natl Acad Sci USA. 106:14542–14546.
2009. View Article : Google Scholar : PubMed/NCBI
|