Introduction
Retinoblastoma (RB) is the most common type of
malignancy in young children and originates from immature cells
within the retina (1). RB has a
very high mortality rate and leads to a large number of
cancer-associated deaths, particularly in developing countries
(2). Increased efforts have been
made in the treatment of RB in the past years; however, these
developments are very limited and the 5-year survival of patients
with RB remains quite poor (3).
The majority of patients with RB are diagnosed at an advanced
stage, and this is accompanied by metastasis, which is a major
cause of RB-associated malignancy (4). Therefore, there is an urgent
requirement to identify novel diagnostic markers and develop
effective therapeutic targets for RB intervention.
MicroRNAs (miRNAs/miRs) are a class of small
noncoding RNAs that regulate gene expression at the
post-transcriptional level by associating with the 3′-untranslated
region (UTR) of target mRNAs to accelerate their degradation
(5,6). Numerous reports have demonstrated
that miRNAs are involved in a variety of physiological processes,
including cell proliferation, development, survival and
differentiation (7–10). Accumulating studies have also
indicated that miRNAs may serve as oncogenes or tumor suppressors
to regulate tumor cell proliferation, migration and invasion
(11–14). For instance, miRNA-320a inhibits
tumor proliferation and invasion by targeting c-Myc in human
hepatocellular carcinoma (15).
Numerous miRNAs have been reported to regulate the progression of
RB, including miR-29a (12),
miR106b (13), miRNA-382 (16) and miRNA-320 (17); however, the functions of miRNAs in
RB are yet to be determined and the roles of most miRNAs in RB
remain unknown.
miR-93-5p has been recently reported to promote the
proliferation and metastasis of numerous human cancers, including
hepatocellular carcinoma (18),
non-small cell lung cancer (11)
and gastric cancer (19); however,
the functions of miR-93-5p in RB remain elusive. In the present
study, it was revealed that miR-93-5p expression was significantly
upregulated in RB tissues compared with in normal tissues.
Additionally, inhibition of miR-93-5p markedly suppressed the
proliferation, migration and invasion while inducing cellular
apoptosis of RB cells. Mechanistically, the present study
demonstrated that phosphatase and tensin homolog (PTEN), an
inhibitor of the phosphoinositide 3-kinase (PI3K)/protein kinase B
(AKT) signaling pathway (20), was
a target of miR-93-5p in RB cells. In addition, the expression of
PTEN was downregulated and inversely correlated with that of
miR-93-5p in RB cells. Furthermore, the present study revealed that
knockdown of PTEN significantly rescued the proliferation,
migration and invasion of RB cells transfected with miR-93-5p
inhibitors. It was reported that miR-93-5p knockdown promoted PTEN
expression and consequently inhibited the activation of the
PI3K/AKT pathway in the present study. In conclusion, the results
of the present study demonstrated that miR-93-5p may be associated
with the progression of RB by modulating the PTEN/PI3K/AKT
signaling pathway.
Patients and methods
Patient samples
A total of 23 human RB (16 males, 7 females; age,
15–51 years) and 12 normal retina tissue samples (8 males, 4
females; age, 22–61 years) were collected between February 2013 to
May 2016, and were provided by the Weifang Medical University
(Weifang, China). All tissue samples were harvested at surgery,
immediately frozen in liquid nitrogen and stored at −80°C until RNA
extraction. The present study was approved by the Ethics Committee
of Weifang Medical University. All of the experiments were
conducted following the obtainment of written informed consent from
each patient or their family.
Cell culture and transfection
The human RB cell lines, including HXO-Rb44, Y79 and
SO-RB50, and the normal human retinal pigment epithelial cell line,
ARPE-19, were purchased from the American Type Culture Collection
(Manassas, VA, USA). All cancer cells were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 100 IU/ml penicillin, 100 mg/ml streptomycin, 20 mM
glutamine and 10% heat-inactivated fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.). The normal human retinal pigment
epithelial cell line ARPE-19 was cultured in Dulbecco's modified
Eagles medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.) with
10% FBS, 5 mg/ml transferrin, 5 mg/ml insulin, 100 ng/ml
hydrocortisone and 10 mM HEPES. All cells were cultured in a
humidified atmosphere of 5% CO2 at 37°C.
A total of 1×106 cells were transfected
with 50 nM miR-93-5p mimics, mimic control, miR-93-5p inhibitors,
negative control (NC) inhibitor, PTEN small interfering RNA (siRNA)
or control siRNA using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. miR-93-5p mimics (5′-CAAAGUGCUGUUCGUGCAGGUAG-3′), mimic
control (5′-ACAUCUGCGUAAGAUUCGAGUCUA-3′), miR-93-5p inhibitors
(5′-CTACCTGCACGAACAGCACTTTG-3′), NC inhibitor
(5′-UCACAACCUCCUAGAAAGAGUAGA-3′), PTEN siRNA
(5′-AGAUGUUAGUGACAAUGAACC-3′) and control siRNA
(5′-AATTCTCCGAACGTGTCACGT-3′) were obtained from Shanghai
GenePharma Co., Ltd. (Shanghai, China). Following transfection,
cells were cultured for 48 h and transfection efficiency was
validated using reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) as described below, prior to subsequent
experimentation.
RT-qPCR
Total RNA was extracted from tumor tissue or
cultured cell lines using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocols. Total RNA was reverse transcribed into cDNA using the
PrimeScript™ RT reagent kit (Takara Biotechnology Co., Ltd.,
Dalian, China) with gDNA Eraser (Takara Biotechnology Co., Ltd.),
according to the manufacturer's protocols; 1 mg total RNA was
reverse transcribed for each sample. qPCR was conducted using the
TaqMan™ MicroRNA Assay kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) for miRNAs and the Fast SYBR™ Green Master Mix
(Applied Biosystems; Thermo Fisher Scientific Inc.) for mRNAs, on
an Applied Biosystems Real-Time PCR machine (Thermo Fisher
Scientific, Inc.). miR-93-5p and PTEN expression levels were
normalized to U6 and GAPDH, respectively using the
2−ΔΔCq method (21).
The thermocycling conditions were as follows: Denaturation at 95°C
for 10 min; followed by 40 cycles of denaturation at 95°C for 15
sec and elongation at 60°C for 1 min. The primer sequences were as
follows: miR-93-5p forward, 5′-ACACTCCAGCTGGGCAAAGTGCTGTTCGTGC-3′
and reverse, 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCTACCTGC-3′;
PTEN forward, 5′-TCCCAGACATGACAGCCATC-3′ and reverse,
5′-TGCTTTGAATCCAAAAACCTTACT-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′; and GAPDH forward,
5′-ATGTTGCAACCGGGAAGGAA-3′ and reverse,
5′-AGGAAAAGCATCACCCGGAG-3′.
Cell Counting Kit-8 (CCK-8)
proliferation assays
Transfected cells were collected at 24 h
post-transfection and seeded into 96-well plates at a density of
3×103 cells per well. Following culturing for 24, 48 and
72 h, a CCK-8 assay (Beyotime Institute of Biotechnology, Shanghai,
China) was performed according to the manufacturer's protocols. In
brief, 10 µl CCK-8 reagent was added to each well; the cells were
incubated at 37°C with 5% CO2 for 2 h. Absorbance was
determined at a wavelength of 450 nm using an ELx808 absorbance
reader (BioTek Instruments, Inc., Winooski, VT, USA). Each assay
was performed in triplicate and repeated three times.
Transwell assays
Transwell chamber assays were performed to analyze
cell invasion and migration. Matrigel was employed to pre-coat the
membrane of Transwell chambers to simulate a matrix barrier for the
invasion assay. The transfected cells in the log phase were seeded
on the upper champers at a density of 2×105 cells/well
in 200 µl serum-free RPMI-1640 medium. A total of 600 µl RPMI-1640
medium with 10% FBS was added to the lower chamber to stimulate
cell migration and invasion. Following 24 h of incubation at 37°C,
cells that had migrated or invaded to the lower chamber were fixed
with paraformaldehyde for 5 min at 25°C and stained with 0.1%
crystal violet for 5 min at 25°C. The images of cells were obtained
under a TS100 inverted light microscope (Nikon Corporation, Tokyo,
Japan) at magnification, ×200; the cell number was counted in five
selected randomly fields per membrane.
Western blot analysis
Y79 and SO-RB50 cells were lysed in cold
radioimmunoprecipitation assay buffer, and protein concentration
was determined using a Pierce™ BCA Protein Assay kit (both Thermo
Fisher Scientific, Inc.). Protein (40 µg/lane) was separated via
10% SDS-PAGE and then transferred to a polyvinylidene difluoride
(PVDF) membrane (Thermo Fisher Scientific, Inc.). The membrane was
blocked using 5% non-fat milk in PBS (Thermo Fisher Scientific,
Inc.) containing 0.1% Tween-20 (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) at room temperature for 2 h. Subsequently, the
PVDF membrane was incubated with rabbit anti-PTEN (1:1,000; cat.
no. ab32199; Abcam, Cambridge, MA, USA), rabbit anti-phosphorylated
(p)-AKT (1:1,000; cat. no. 4060; Cell Signaling Technology, USA),
rabbit anti-AKT (1:1,000; cat. no. ab18785) and rabbit anti-GAPDH
(1:1,000; cat. no. ab9485; both Abcam) primary antibodies at room
temperature for 2 h. Following washing with PBS for 10 min, the
PVDF membrane was incubated with horseradish peroxidase-conjugated
goat anti-rabbit secondary antibodies (1:5,000; cat. no. ab7090;
Abcam) at room temperature for 1 h. Membranes were then washed with
PBS for 10 min and the protein bands were visualized using the
Pierce™ ECL Western Blotting Substrate kit (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Protein densitometry was performed using ImageJ Software version
1.41 (National Institutes of Health, Bethesda, MD, USA).
Cell apoptosis assays
Cell apoptosis was determined using an Annexin
V-fluorescein isothiocyanate (FITC) Apoptosis Detection kit
(Nanjing KeyGen Biotech Co., Ltd., Nanjing, China), according to
the manufacturer's protocols. Briefly, RB cells were harvested 48 h
following transfection and then washed with PBS. Then,
5×104 cells were resuspended in 500 µl of binding buffer
containing 5 µl of Annexin V-FITC and 5 µl of propidium iodide (PI)
for 10 min at 4°C. Subsequently, apoptosis was determined using a
flow cytometer. The data were analyzed with FlowJo software version
5.7.2 (FlowJo LLC, Ashland, OR, USA). The total percentage Annexin
V-positive cells indicated apoptotic cells.
Luciferase assay
TargetScan version 7.1 (http://www.targetscan.org/index.html) was used to
predict potential miR-93-5p targets. The wild-type (WT) 3′-UTR
sequence of PTEN or the mutant (Mut) 3′-UTR sequence of PTEN was
amplified by PCR and incorporated into the pGL3 control vector
(Promega Corporation, Madison, WI, USA) between MluI and
XhoI restriction sites, to obtain the WT-PTEN-3′-UTR or
Mut-PTEN-3′-UTR, respectively. A total of 2×104
cells/well were seeded into 24-well plates the day prior to
transfection and then cells were cotransfected with 1 mg WT or Mut
PTEN-3′-UTR along with 50 nM miR-93-5p mimics or mimic control
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). At 48 h following cotransfection, the luciferase
activity for the WT or Mut PTEN 3′-UTR was measured using the
Dual-Luciferase® Reporter Assay system (Promega
Corporation) and normalized to Renilla luciferase
activity.
Statistical analysis
SPSS statistical software for Windows version 19.0
(IBM Corp., Armonk, NY, USA) and GraphPad Prism version 5.01
(GraphPad Software, Inc., La Jolla, CA, USA) software were used for
statistical analysis. All experiments were repeated at least three
times and data are represented as the mean ± standard deviation
from at least three independent experiments. A Student's t-test and
one-way analysis of variance followed by a Tukey's post hoc test
was used to compare two or multiple groups, respectively, for
statistical significance. Spearman's rank correlation analysis was
performed to analyze correlation between miR-93-5p and PTEN
expression levels. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-93-5p is highly expressed in RB
tissues and cell lines
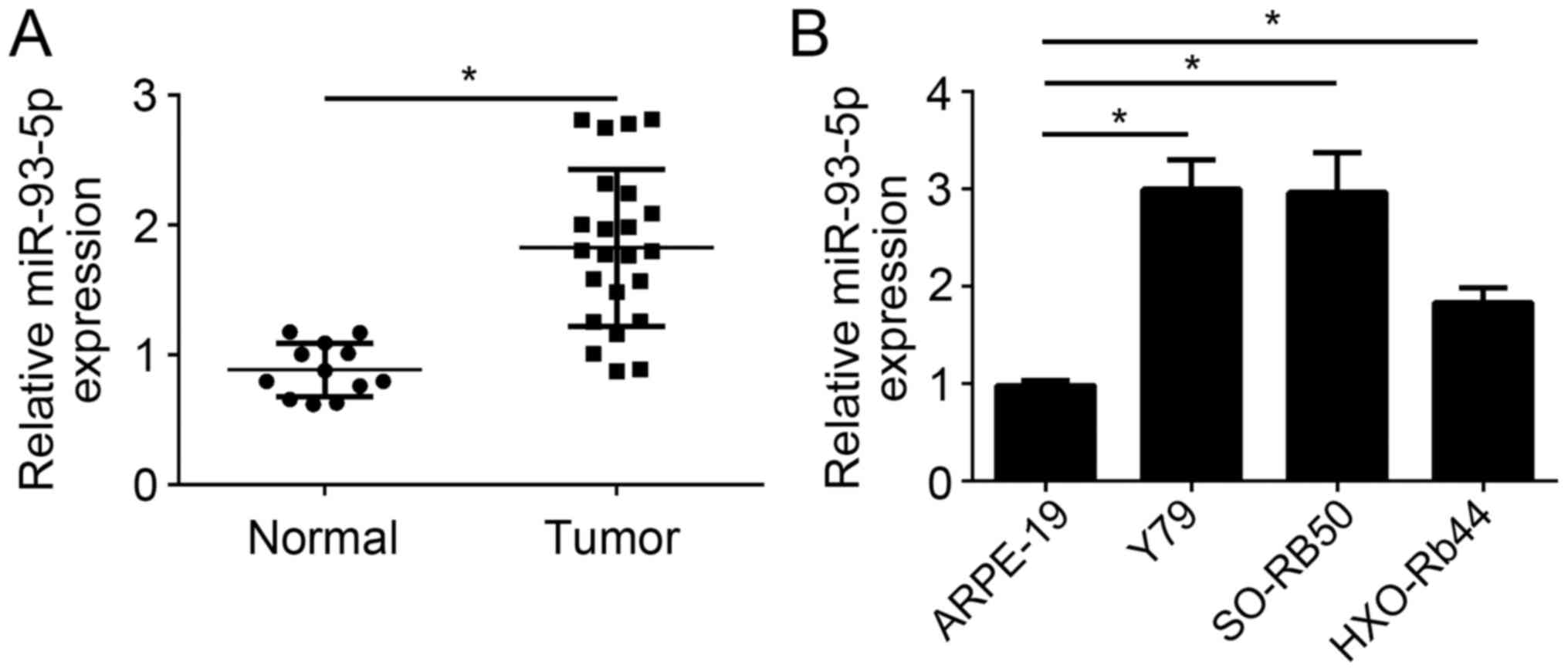
The present study analyzed the expression levels of
miR-93-5p in RB tissues by RT-qPCR. It was observed that the
expression levels of miR-93-5p were significantly upregulated in RB
tissues (n=23) compared with in normal tissues (n=12) (P<0.05;
Fig. 1A). Furthermore, RT-qPCR
analysis also indicated that miR-93-5p expression levels were also
significantly upregulated in the RB cell lines, including Y79,
SO-RB50 and HXO-Rb44, compared with in ARPE-19 cells (P<0.05;
Fig. 1B). miR-93-5p expression
levels were highest in Y79 and SO-RB50 cells; therefore, these two
cell lines were selected for following experiments. These results
demonstrated that miR-93-5p expression was upregulated in RB cells,
indicating that miR-93-5p may be involved in the progression of
RB.
Knockdown of miR-93-5p inhibits RB
cell proliferation and promotes apoptosis
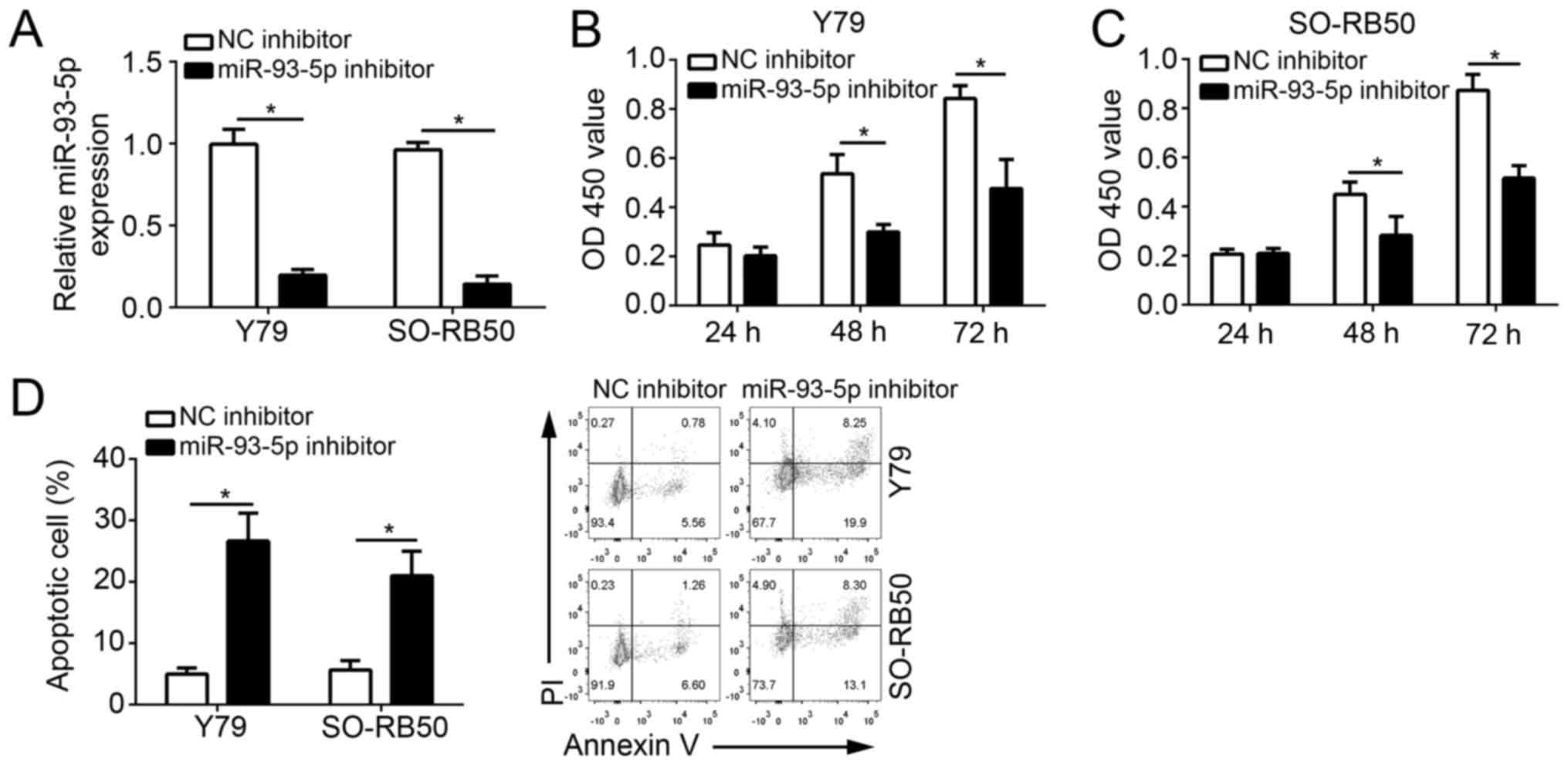
To investigate the function of miR-93-5p, the
expression of miR-93-5p was downregulated in Y79 and SO-RB50 cells
via transfection with a miR-93-5p inhibitor. RT-qPCR analysis
indicated that miR-93-5p expression levels were significantly
downregulated in Y79 and SO-RB50 cells compared with in cells
transfected with the NC inhibitor (P<0.05; Fig. 2A). In addition, CCK-8 assays were
performed to analyze cell proliferation. As presented in Fig. 2B and C, miR-93-5p knockdown
significantly suppressed the proliferation of Y79 and SO-RB50 cells
at 48 and 72 h following transfection compared with in the NC
inhibitor group (P<0.05). Furthermore, cell apoptosis was
determined by Annexin V/PI staining. The results demonstrated that
miR-93-5p knockdown significantly increased the percentage of Y79
and SO-RB50 apoptotic cells (P<0.05; Fig. 2D). The results suggested that
miR-93-5p promoted the malignant behavior of RB cells.
miR-93-5p knockdown suppresses RB cell
migration and invasion
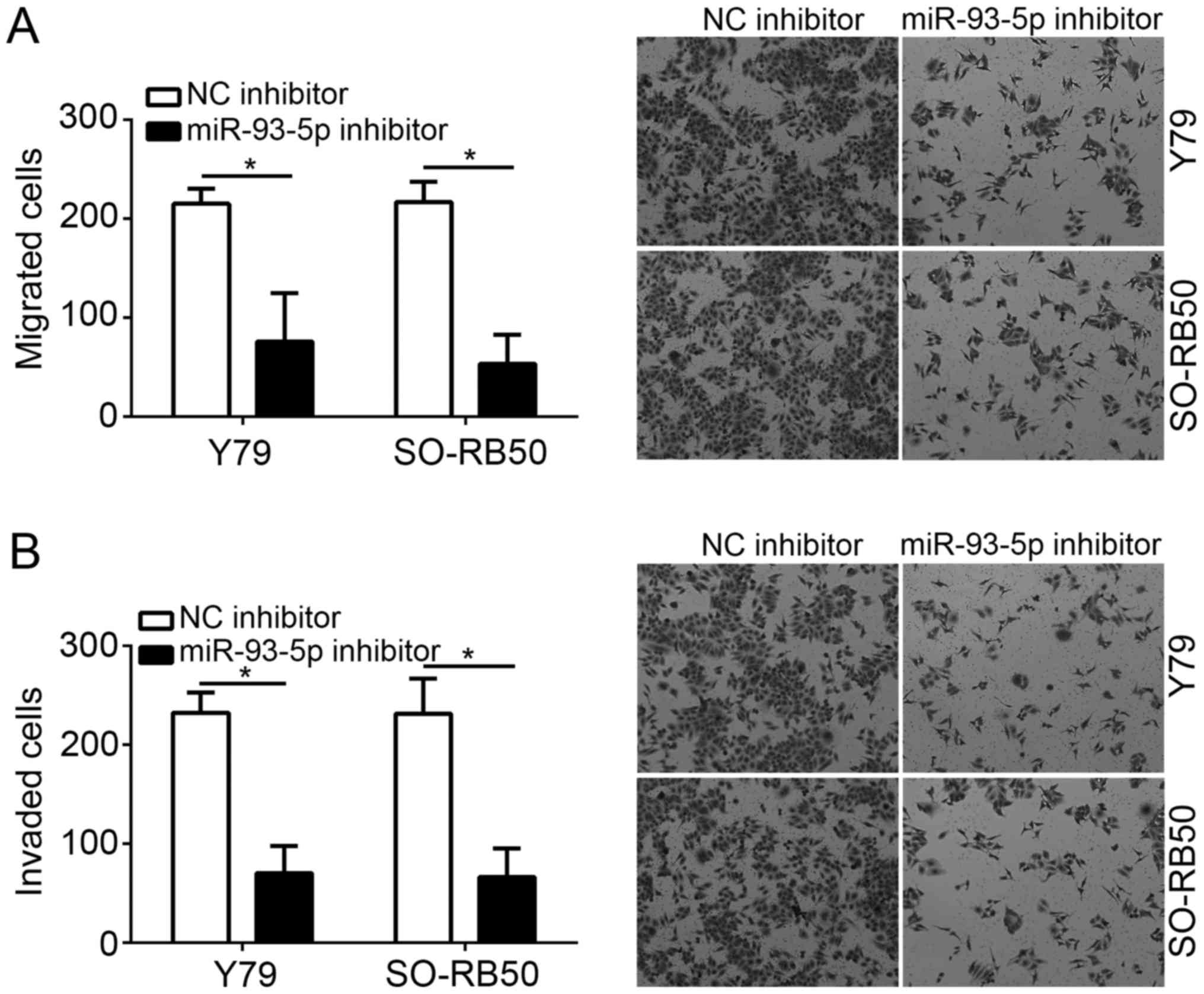
To analyze the effects of miR-93-5p on tumor
metastasis, the cell migration and invasive abilities of RB cells
transfected with miR-93-5p or control inhibitors were investigated
via Transwell assays. The results indicated that miR-93-5p
knockdown significantly suppressed the migration and invasive
abilities of Y79 and SO-RB50 cells compared with in the NC
inhibitor group (P<0.05; Fig. 3A
and B).
PTEN is a target of miR-93-5p
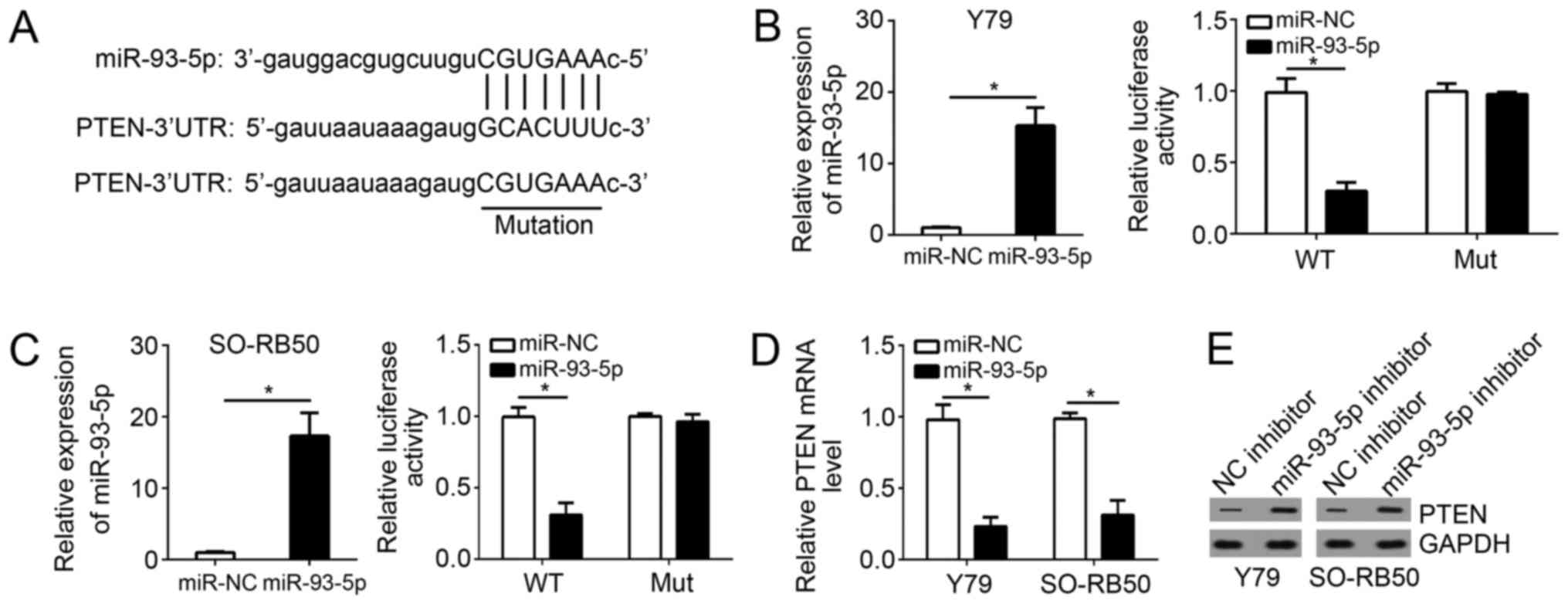
To further investigate the mechanism of miR-93-5p in
RB cells, bioinformatics analysis was conducted. The present study
reported that PTEN was a potential target of miR-93-5p. A potential
binding site of miR-93-5p was determined in the 3′-UTR of PTEN mRNA
(Fig. 4A). To verify this
prediction, luciferase reporter assays were conducted with RB cells
cotransfected with miR-93-5p mimics or control and WT-PTEN-3′-UTR
or Mut-PTEN-3′-UTR. Firstly, RT-qPCR was performed to confirm
successful miR-93-5p overexpression (Fig. 4B and C). Next, the luciferase
reporter assay revealed that miR-93-5p overexpression significantly
inhibited the luciferase activity of the WT-PTEN-3′-UTR compared
with in cells cotransfected with control (P<0.05); however, that
of the Mut-PTEN-3′-UTR in Y79 and SO-RB50 cells were unaffected
(Fig. 4B and C). To directly
analyze the effects of miR-93-5p on PTEN expression, miR-93-5p
mimic or control was transfected into RB cells, which demonstrated
that overexpression of miR-93-5p significantly reduced the mRNA
expression levels of PTEN in Y79 and SO-RB50 cells (Fig. 4D). Consistently, knockdown of
miR-93-5p markedly upregulated the protein expression levels of
PTEN in Y79 and SO-RB50 cells (Fig.
4E). The data demonstrated that PTEN may be a direct target of
miR-93-5p in RB cells.
PTEN expression is inversely
correlated with miR-93-5p expression in RB tissues
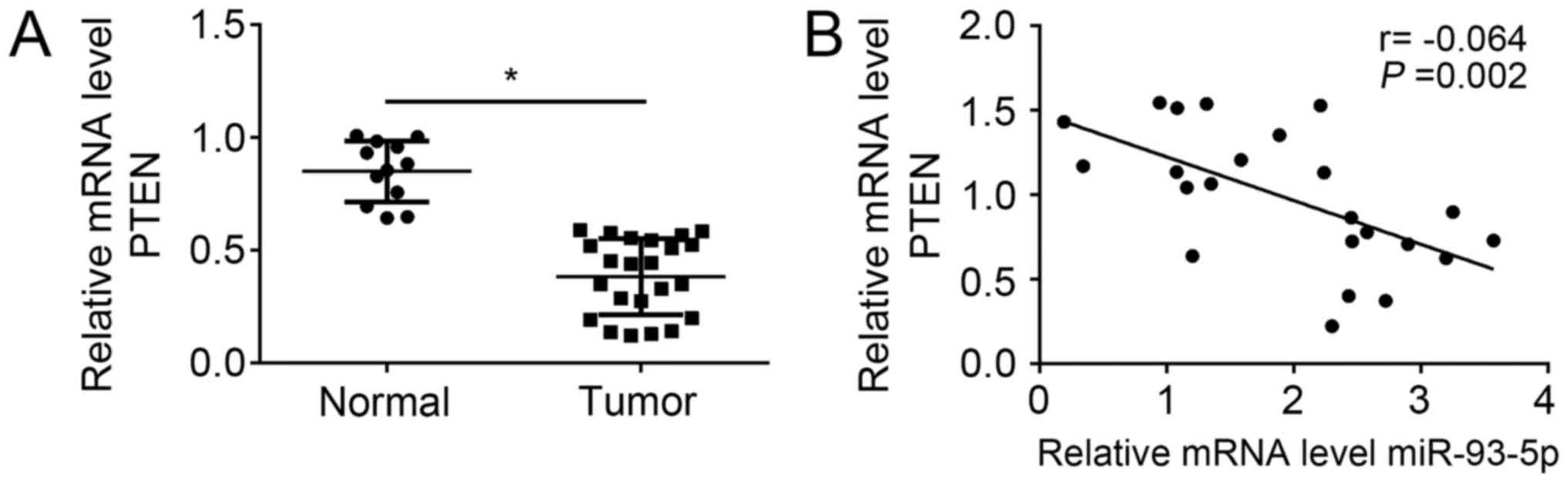
The present study investigated the expression levels
of PTEN in RB tissues by RT-qPCR. The results revealed that PTEN
expression levels were significantly downregulated in RB tissues
compared with in normal tissues (Fig.
5A). In addition, a statistically significant inverse
correlation was observed via Spearman's correlation analysis
between miR-93-5p and PTEN levels in RB tissues (Fig. 5B).
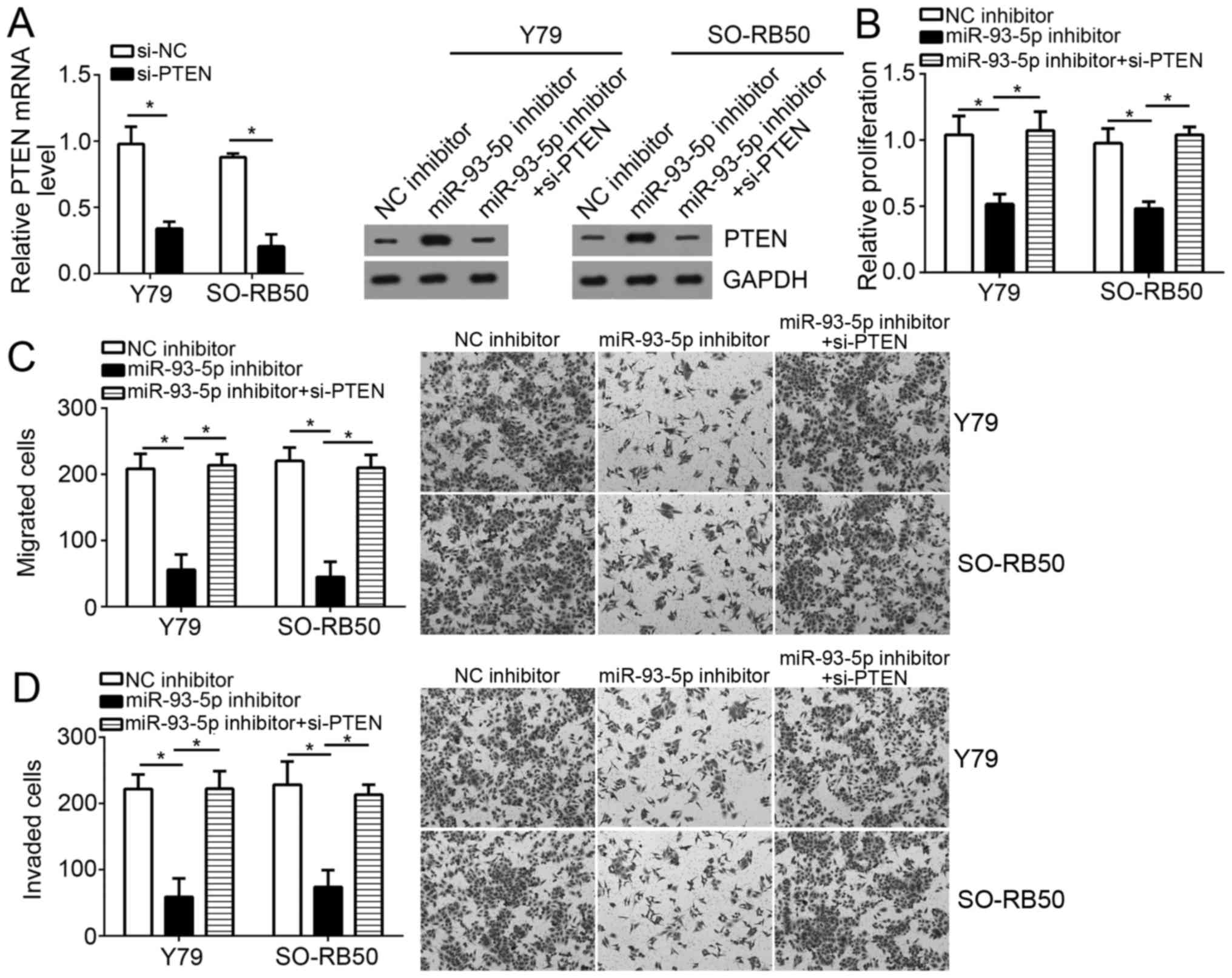
PTEN knockdown reverses the inhibitory
effects of miR-93-5p depletion on the proliferation, migration and
invasion of RB cells
As miR-93-5p was determined to be significantly
overexpressed in tumor cells (Fig.
1B), a rescue assay using miR-93-5p inhibitors was conducted;
the expression levels of PTEN were downregulated in
miR-93-5p-depleted RB cells via transfection with a specific siRNA
against PTEN. RT-qPCR confirmed the efficiency of PTEN siRNA
transfection (Fig. 6A). Western
blotting indicated that the expression levels of PTEN were markedly
downregulated compared with in miR-93-5p-downregulated RB cells
(Fig. 6A). Notably, knockdown of
PTEN significantly rescued the suppressive effects of miR-93-5p
inhibition on RB cell proliferation, migration and invasion
(P<0.05; Fig. 6B-D). These
results indicated that miR-93-5p may exerts its oncogenic roles
within RB cells by regulating PTEN expression, at least partly.
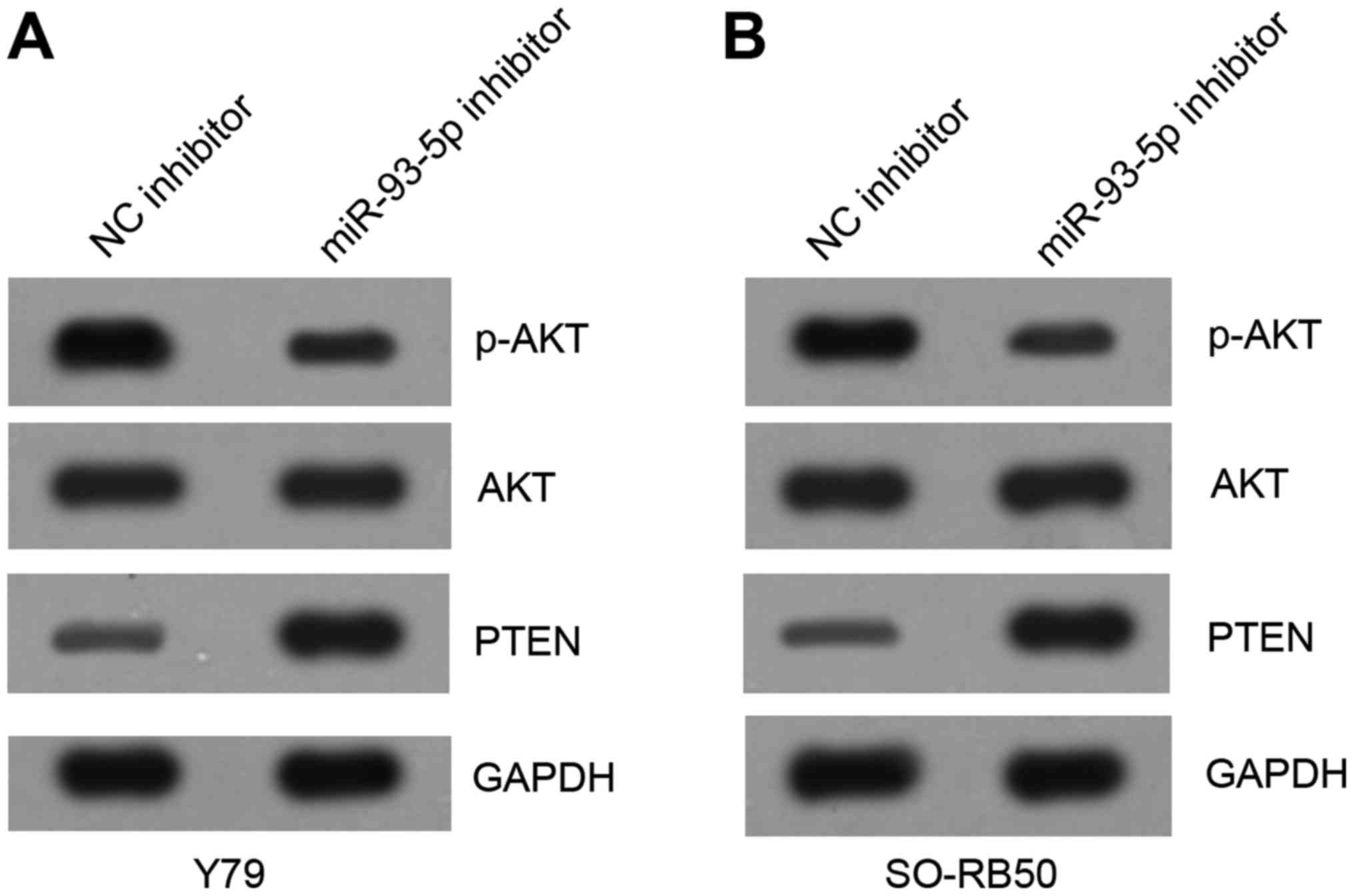
miR-93-5p knockdown suppresses the
PI3K/AKT signaling pathway in RB cells
PTEN has been demonstrated to be a negative
regulator of the PI3K/AKT signaling pathway, which is abnormally
activated in numerous types of tumors, including RB (22). The present study investigated
whether miR-93-5p may exert an effect on the PI3K/AKT signaling
pathway. Western blotting results indicated that knockdown of
miR-93-5p notably upregulated the protein levels of PTEN and
inhibited that of p-AKT compared with in the control group
(Fig. 7A and B), which suggested
that miR-93-5p positively activates the PI3K/AKT signaling pathway
in RB cells. In conclusion, the present study demonstrated that
miR-93-5p may exert an oncogenic role in RB via regulation of the
PTEN/PI3K/AKT signaling pathway.
Discussion
In the past decade, miRNAs have been widely
demonstrated to serve essential functions in the initiation and
progression of RB via regulating the expression of specific genes
(23). For instance, Liu et
al (24) reported that the
proliferation, migration and invasion of human RB cells were
significantly suppressed by miR-124 in a signal transducer and
activator of transcription 3-dependent manner. Lei et al
(25) reported that miR-101 was
downregulated in RB tissues and suppressed tumor cell growth and
proliferation by inhibiting the expression of enhancer of zeste
homolog 2. Liu et al (26)
demonstrated that miR-34a was downregulated in RB tissues and
enhanced tumor cell chemosensitivity and promoted cell death by
targeting high mobility group box 1 to suppress autophagy. Wang
et al (27) reported that
miRNA-183 inhibited the proliferation, migration and invasion of RB
cells downregulating the expression of low-density lipoprotein
receptor-related protein 6. Martin et al (28) revealed that miR-449 significantly
inhibited the proliferation and induced the apoptosis of RB cells.
Additionally, Wang et al (29) reported that miR-365b-3p was
downregulated in RB tissues and was associated with cell cycle
arrest, and enhanced tumor cell apoptosis by targeting paired box
protein Pax-6. In the present study, it was observed that miR-93-5p
was significantly upregulated in RB tissues and cell lines.
Functional experiments demonstrated that miR-93-5p knockdown
inhibited the proliferation, migration and invasion, but induced
the apoptosis of RB cells. The results of the present study
indicated that miR-93-5p serves a crucial role in the progression
of RB.
miR-93-5p has been demonstrated to serve as an
oncogene and promote tumor growth and metastasis in numerous types
of cancer, including ovarian carcinoma (30), endometrial carcinoma (31), triple-negative breast cancer
(32), gastric cancer (19), non-small cell lung cancer (11) and hepatocellular carcinoma
(18). However, to the best of our
knowledge, the physiological functions and underlying molecular
mechanism of miR-93-5p in RB have not been investigated. In the
present study, it was reported that miR-93-5p expression was
significantly upregulated in RB specimens compared with in normal
retinal tissues. Consistently, the expression levels of miR-93-5p
were higher in RB cell lines than that of ARPE-19 cells.
Additionally, the present study revealed that knockdown of
miR-93-5p suppressed the proliferation, migration and invasion of
RB cells, but enhanced cellular apoptosis. These results suggested
that miR-93-5p serves as an oncogene in RB.
Increasing evidence has indicated that miRNAs exert
biological functions in cancer cells by suppressing the expression
of target genes (31,33). Bioinformatics analysis using
TargetScan indicated that PTEN may be a potential target of
miR-93-5p as determined in the present study. In addition, the
interaction between PTEN and miR-93-5p in RB cells was demonstrated
by luciferase reporter assays, RT-qPCR and western blot analysis in
the present study. PTEN has been reported to suppress tumor growth
and metastasis in various cancers by regulating proliferation,
migration and invasion, and the cell cycle (20). Importantly, accumulating evidence
has indicated that PTEN is a negative regulator of the PI3K/AKT
signaling pathway, in which aberrant activation leads to the
progression of several types of cancer, such as RB (16,34);
however, the mechanism underlying the regulation of PTEN in RB
requires further investigation. In the present study, the
inhibition of miR-93-5p was associated with significantly enhanced
PTEN expression in RB cells; opposing results were observed when
PTEN expression was downregulated. In addition, the present study
reported that miR-93-5p expression levels were inversely correlated
with PTEN mRNA expression in RB tissues. Knockdown of PTEN
partially reversed the effects of miR-93-5p depletion on RB cell
proliferation, migration and invasion. Additionally, the results of
the present study also demonstrated that knockdown of miR-93-5p
inhibited the activation of the PI3K/AKT pathway in RB cells. These
observations suggest that miR-93-5p exhibited an oncogenic role in
RB via the targeting of PTEN. Compared to the previous study by Wei
et al (34), the present
study revealed the relationship between PTEN and miR-93-5p in RB
progression.
In conclusion, the present study reported that
miR-93-5p may promoted the progression of RB by regulating cell
proliferation, apoptosis, migration and invasion in a PTEN/PI3K/AKT
signaling pathway-dependent manner. These results suggest that
miR-93-5p may be a potential therapeutic target for the treatment
of RB; however, further investigation is required.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YC and MG made substantial contributions to the
design of the present study, analyzed and interpreted the results,
and wrote the manuscript. FX and PW performed the experiments,
including apoptosis detection and luciferase reporter assays. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
For the use of human samples, the present study was
approved by the Institutional Ethics Committee of Weifang Medical
University and all enrolled patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shields CL and Shields JA: Retinoblastoma
management: Advances in enucleation, intravenous chemoreduction,
and intra-arterial chemotherapy. Curr Opin Ophthalmol. 21:203–212.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jabbour P, Chalouhi N, Tjoumakaris S,
Gonzalez LF, Dumont AS, Chitale R, Rosenwasser R, Bianciotto CG and
Shields C: Pearls and pitfalls of intraarterial chemotherapy for
retinoblastoma. J Neurosurg Pediatr. 10:175–181. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Houston SK, Murray TG, Wolfe SQ and
Fernandes CE: Current update on retinoblastoma. Int Ophthalmol
Clin. 51:77–91. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kivelä T: The epidemiological challenge of
the most frequent eye cancer: Retinoblastoma, an issue of birth and
death. Br J Ophthalmol. 93:1129–1131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen K and Rajewsky N: The evolution of
gene regulation by transcription factors and microRNAs. Nat Rev
Genet. 8:93–103. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Du T and Zamore PD: Beginning to
understand microRNA function. Cell Res. 17:661–663. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen CZ, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Brennecke J, Hipfner DR, Stark A, Russell
RB and Cohen SM: bantam encodes a developmentally regulated
microRNA that controls cell proliferation and regulates the
proapoptotic gene hid in Drosophila. Cell. 113:25–36. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Geng S and Zhou W: Influence of extrinsic
inputs and synaptic gains on dynamics of Wendling's neural mass
model: A bifurcation analysis. J Integr Neurosci. 15:463–483. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu P, Vernooy SY, Guo M and Hay BA: The
Drosophila microRNA Mir-14 suppresses cell death and is required
for normal fat metabolism. Curr Biol. 13:790–795. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang W, Bai J, Liu D, Wang S, Zhao N, Che
R and Zhang H: MiR-93-5p up-regulation is involved in non-small
cell lung cancer cells proliferation and migration and poor
prognosis. Gene. 647:13–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu S, Zhang X, Hu C, Wang Y and Xu C:
miR-29a inhibits human retinoblastoma progression by targeting
STAT3. Oncol Rep. 39:739–746. 2018.PubMed/NCBI
|
|
13
|
Yang G, Fu Y, Zhang L, Lu X and Li Q:
miR106b regulates retinoblastoma Y79 cells through Runx3. Oncol
Rep. 38:3039–3043. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shen Y, Ye YF, Ruan LW, Bao L, Wu MW and
Zhou Y: Inhibition of miR-660-5p expression suppresses tumor
development and metastasis in human breast cancer. Genet Mol Res.
16:2017. View Article : Google Scholar
|
|
15
|
Xie F, Yuan Y, Xie L, Ran P, Xiang X,
Huang Q, Qi G, Guo X, Xiao C and Zheng S: miRNA-320a inhibits tumor
proliferation and invasion by targeting c-Myc in human
hepatocellular carcinoma. Onco Targets Ther. 10:885–894. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Song D, Diao J, Yang Y and Chen Y:
MicroRNA382 inhibits cell proliferation and invasion of
retinoblastoma by targeting BDNF-mediated PI3K/AKT signalling
pathway. Mol Med Rep. 16:6428–6436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao Y, Zhang S and Zhang Y: MicroRNA-320
inhibits cell proliferation, migration and invasion in
retinoblastoma by targeting specificity protein 1. Mol Med Rep.
16:2191–2198. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang X, Liao Z, Bai Z, He Y, Duan J and
Wei L: MiR-93-5p promotes cell proliferation through
down-regulating PPARGC1A in hepatocellular carcinoma cells by
bioinformatics analysis and experimental verification. Genes
(Basel). 9:E512018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li L, Zhao J, Huang S, Wang Y, Zhu L, Cao
Y, Xiong J and Deng J: MiR-93-5p promotes gastric cancer-cell
progression via inactivation of the Hippo signaling pathway. Gene.
641:240–247. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Waniczek D, Śnietura M, Lorenc Z,
Nowakowska-Zajdel E and Muc-Wierzgoń M: Assessment of PI3K/AKT/PTEN
signaling pathway activity in colorectal cancer using quantum
dot-conjugated antibodies. Oncol Lett. 15:1236–1240.
2018.PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Di Fiore R, Drago-Ferrante R, D'Anneo A,
Augello G, Carlisi D, De Blasio A, Giuliano M, Tesoriere G and
Vento R: In human retinoblastoma Y79 cells okadaic
acid-parthenolide co-treatment induces synergistic apoptotic
effects, with PTEN as a key player. Cancer Biol Ther. 14:922–931.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang Y and Mei Q: miRNA signature
identification of retinoblastoma and the correlations between
differentially expressed miRNAs during retinoblastoma progression.
Mol Vis. 21:1307–1317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu S, Hu C, Wang Y, Shi G, Li Y and Wu H:
miR-124 inhibits proliferation and invasion of human retinoblastoma
cells by targeting STAT3. Oncol Rep. 36:2398–2404. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lei Q, Shen F, Wu J, Zhang W, Wang J and
Zhang L: MiR-101, downregulated in retinoblastoma, functions as a
tumor suppressor in human retinoblastoma cells by targeting EZH2.
Oncol Rep. 32:261–269. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu K, Huang J, Xie M, et al: MIR34A
regulates autophagy and apoptosis by targeting HMGB1 in the
retinoblastoma cell. Autophagy. 10:442–452. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang J, Wang X, Li Z, Liu H and Teng Y:
MicroRNA-183 suppresses retinoblastoma cell growth, invasion and
migration by targeting LRP6. FEBS J. 281:1355–1365. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Martin A, Jones A, Bryar PJ, et al:
MicroRNAs-449a and −449b exhibit tumor suppressive effects in
retinoblastoma. Biochem Biophys Res Commun. 440:599–603. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang J, Wang X, Wu G, Hou D and Hu Q:
MiR-365b-3p, down-regulated in retinoblastoma, regulates cell cycle
progression and apoptosis of human retinoblastoma cells by
targeting PAX6. FEBS Lett. 587:1779–1786. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen X, Chen S, Xiu YL, Sun KX, Zong ZH
and Zhao Y: RhoC is a major target of microRNA-93-5P in epithelial
ovarian carcinoma tumorigenesis and progression. Mol Cancer.
14:312015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen S, Chen X, Sun KX, Xiu YL, Liu BL,
Feng MX, Sang XB and Zhao Y: MicroRNA-93 promotes
epithelial-mesenchymal transition of endometrial carcinoma cells.
PLoS One. 11:e01657762016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shyamasundar S, Lim JP and Bay BH: miR-93
inhibits the invasive potential of triple-negative breast cancer
cells in vitro via protein kinase WNK1. Int J Oncol. 49:2629–2636.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ivkovic Catela T, Voss G, Cornella H and
Ceder Y: microRNAs as cancer therapeutics: A step closer to
clinical application. Cancer Lett. 407:113–122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wei D, Miao Y, Yu L, Wang D and Wang Y:
Downregulation of microRNA198 suppresses cell proliferation and
invasion in retinoblastoma by directly targeting PTEN. Mol Med Rep.
18:595–602. 2018.PubMed/NCBI
|