Introduction
As an important part of the immune system,
antimicrobial peptides (AMPs) can physically destroy microbial
membranes and induce their cleavage, indicating their potential as
substitutes for traditional antibiotics (1). Following the identification of LL-37,
the only natural human cathelicidin (an AMP subfamily), in 1995
(2,3), its presence has been detected in
various types of cells and epithelia, including intestinal
epithelial cells (4), mast cells
(5), monocytes (6) and lymphocytes (7). Natural human AMPs are the first line
of defense against local infection, serving an important role in
the invasion of systemic pathogens into local areas or local wound
infections (8,9). As the peptide chain of LL-37 is too
long and difficult to synthesize, it is not a routine therapeutic
drug for bacterial infections or other inflammatory diseases in the
clinic; however, the cost of producing short-chain AMPs is
relatively low. Furthermore, the removal of hydrophobic amino acids
from the N-terminus of natural LL-37 can reduce its toxicity to
eukaryotic cells, as well as reduce its interactions with human
plasma proteins (10,11). Thus, short-chain AMPs have
attracted the attention of researchers. Compared with natural
LL-37, KR-20 and KS-30, two derivatives of LL-37, exhibit improved
antimicrobial properties (12,13).
KR-12 is the shortest derivative of LL-37 with antimicrobial
activity (10,11). Compared with LL-37 and its other
derivatives, the cost of KR-12 synthesis is lower, and its
cytotoxicity is reduced; for example, LL-37, but not KR-12, induces
hemolytic effectsin human erythrocytes (14). As an analogue of KR-12 (amino acid
sequence: KRIVQRIKDFLR-NH2), KR-12-a6 (amino acid
sequence: LRIVKLILKWLR-NH2) has similar antibacterial
properties and biocompatibility (15); also, as a highest hydrophobic
analogue of KR-12, KR-12-a6 displays greater inhibition of
lipopolysaccharide (LPS)-stimulated tumor necrosis factor-α
production and higher LPS-binding activity than KR-12 (15). Therefore, KR-12-a6 may be a
potential therapeutic drug for the treatment of clinical infectious
diseases, including osteomyelitis.
Osteomyelitis is a common infectious disease that
can cause serious symptoms locally or systemically, such as severe
pain, high fever, swelling and tenderness around the infection
site, posing a great challenge to clinical treatment (16). In addition to the surgical removal
of infected bone tissues, local or systemic application of
antibiotics is the most common treatment (17); however, with the increasing use of
antibiotics to treat bone infections in the clinic, bacterial
resistance to antibiotics has gradually developed (18). In addition, osteolysis caused by
infection cannot be treated with traditional antibiotics.
Gentamicin (19) and vancomycin
(20), two commonly used
antibiotics in the treatment of bone-related infections, have been
shown to inhibit the viability of osteoblasts and reduce their
numbers. Recently, certain small polypeptides have been reported to
promote bone integration while controlling infection (21). Therefore, drugs with favorable
antibacterial properties and pro-osseointegration abilities are of
great importance for the treatment of clinical bone infections.
Human bone marrow mesenchymal stem cells (hBMSCs)
play a key role in the process of bone regeneration (22–24).
hBMSCs can differentiate into osteoblasts during bone formation,
and are the main source of bone progenitor cells (25). When local infections are
controlled, hBMSCs are activated and differentiate into osteoblasts
to repair the local osteolysis (26). However, when hBMSCs fail to
completely repair the local bone defect caused by infection, local
osteoporosis and even pathological fracture can result (27). At present, drugs that inhibit
osteoclast activity and bone resorption, such as vitamin D
analogues, calcitonin and estrogen, are commonly used in the
treatment of osteoporosis (28).
However, these drugs do not promote osteogenic differentiation or
exhibit antibacterial properties. Considering the increased
resistance to antibiotics in the clinic and the ideal antibacterial
properties of AMPs, it was proposed that AMPs may exhibit positive
therapeutic effects for the treatment of osteomyelitis (29). Therefore, the present study
examined whether its analogue KR-12-a6 could also promote the
osteogenic differentiation of hBMSCs and investigated the
underlying mechanisms.
Materials and methods
Isolation and culture of hBMSCs
Bone marrow was obtained from the iliac crest marrow
aspirates of 3 healthy donors (one 34-year old male, two females at
19 and 38-year's old) undergoing iliac crest bone transplantation
at Jingzhou Central Hospital, Tongji Medical College of Huazhong
University of Science and Technology (Jingzhou, China) between
January and July 2018. The procedure was performed following
approval from the Ethics Committee of Huazhong University of
Science and Technology and after obtaining donors' informed
consent. The collected bone marrow was treated with heparin
anticoagulant and then diluted in α-minimal essential medium
(α-MEM; Gibco; Thermo Fisher Scientific, Inc.) (30). hBMSCs were then isolated by density
gradient centrifugation (400 × g for 20 min at room temperature)
with Ficoll-Paque (GE Healthcare) and plastic adherence, purified
by discarding suspended cells through exchanging medium, and then
grown in α-MEM supplemented with 20% fetal bovine serum (Atlanta
Biologicals; Bio-Techne Corporation), 100 U/ml penicillin
(Invitrogen; Thermo Fisher Scientific, Inc.), 100 µg/ml
streptomycin (Invitrogen; Thermo Fisher Scientific, Inc.), and 2 mM
L-glutamine (Invitrogen; Thermo Fisher Scientific, Inc.). Cells
were then cultured at 37°C with 5% humidified CO2. The
hBMSCs were passaged and maintained at low density. Adherent cells
reaching 80% confluence were harvested by using 0.25% trypsin at
37°C for 3 min. Then, 1×105 cells were replated, and the
remainder was used for the analysis of gene expression.
Osteoblast differentiation
hBMSCs were plated in 6-well plates in standard
growth medium. At 80% confluence, the medium was replaced with
osteogenic medium consisting of α-MEM + 10% fetal calf serum
(Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin supplemented with 10 mM β-glycerophosphate
(Calbiochem; Merck KGaA), 50 µg/ml L-ascorbic acid (Wako
Chemicals GmbH), 10 nM dexamethasone (Sigma-Aldrich; Merck KGaA),
and 10 nM calcitriol (Sigma-Aldrich; Merck KGaA). The medium was
replaced every 3 days. The control cells were grown in standard
growth medium. Cell pellets were harvested for RNA isolation at
different time points following induction.
Alkaline phosphatase (ALP) staining
and quantitative analysis
hBMSCs were cultured in 24-well plates at
2×104 cells/cm2. At 80% confluence, cells
were induced to differentiate into osteoblasts using StemPro™
Osteogenesis Differentiation kit (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with KR-12-a6 (synthesized and purified by GL
Biochem (Shanghai) Ltd.) at 0, 20, 30, 40, 60, or 80 µg/ml
at 37°C for 7 days. For ALP staining, hBMSCs in each group were
washed twice with PBS at the end of the 7-day KR-12-a6-treatment
period, fixed with 4% paraformaldehyde for 30 s, and then stained
using the ALP staining kit (Maokang Biotechnology, Shanghai)
according to the manufacturer's instructions. Images were acquired
by an inverted phase contrast microscope (CKX41, Olympus) under
×100 magnification. At least five fields per sample were randomly
selected and observed. To quantitatively analyze ALP activity,
hBMSCs after the 7-day KR-12-a6-treatment period were washed twice
with PBS and lysed with Triton X-100 (1%) for 15 min. Then, the
activity was measured by Alkaline Phosphatase Assay kit (Beyotime
Institute of Biotechnology) according to the manufacturer's
instructions. The final estimation was based on the absorbance at
405 nm measured by a spectrophotometer.
Alizarin red staining and quantitative
analysis
Assessment of ex vivo mineralization was
performed by employing alizarin red staining. hBMSCs at the density
of 5×104 cells/well in 6-well plates underwent
osteoblast differentiation in medium supplemented with KR-12-a6 at
0, 20, 30, 40, 60, or 80 µg/ml at 37°C for 21 days. Then
cells were washed in PBS, fixed in 70% ethanol at −20°C for 1 h,
and rinsed in dH2O. The cultures were stained with 40 mM
alizarin red (Sigma-Aldrich; Merck KGaA) at pH 4.2 for 10 min at
room temperature with rotation. Cells were then rinsed twice with
dH2O, followed by washing three times with PBS to reduce
nonspecific staining. Images were acquired by an inverted phase
contrast microscope (CKX41, Olympus Corporation) under ×100
magnification. At least five fields per sample were randomly
selected and observed. For further quantitative analysis, a 10%
chlorinated 16 alkyl pyridine solution of sodium phosphate (pH 7.0)
was added into each sample to dissolve the dye, and the absorbance
was measured at 620 nm by a spectrophotometer.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
hBMSCs at the density of 5×104 cells/well
in 12-well plates were treated with KR-12-a6 at 0, 20, 30, and 40
µg/ml and then cultured at 37°C for 3, 7, and 14 days. On
day 3, 7, and 14 post-KR-12-a6 treatment, hBMSCs were harvested in
Buffer RLT using an RNeasy Mini kit (Qiagen, Inc.). Total RNA was
prepared according to standard protocols. Complementary DNAs
(cDNAs) were synthesized from 0.5 µg total RNA using a QuantiTect
Reverse Transcription kit (Qiagen, Inc.) according to the
manufacturer's protocols. qPCR was conducted using a QuantiTect
SYBR Green PCR kit (Qiagen, Inc.) and an ABI PRISM 7900HT SDS
instrument (Applied Biosystems; Thermo Fisher Scientific, Inc.).
qPCR was performed as follows: 2 min activation at 95°C, then 40
cycles of 15 sec denaturation at 95°C and 15 sec annealing at 62°C,
and a final 20-sec extension at 68°C. All reactions were in a final
volume of 20 µl consisting of 2X master mix (10 µl), forward and
reverse primers (2 µl), cDNA, and RNase H2O, and each
sample was run in triplicate. The specificity of PCR products was
confirmed at the end of each run via melting curve analysis. All
signals were normalized to β-actin, and the quantification
cycle was determined. RT-qPCR was quantified by 2−∆∆Cq
method (31). Oligonucleotide
primers used for RT-qPCR are presented in Table I. β-actin was used as the
housekeeping gene to normalize gene expression levels.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Forward primer | Reverse primer |
|---|
| RUNX2 |
5′-GCCACCACTCACTACCACACCTA-3′ |
5′-TCCTGACGAAGTGCCATAGTAGAGATAT-3′ |
| ALP |
5′-GGACCATTCCCACGTCTTCAC-3′ |
5′-CCTTGTAGCCAGGCCCATTG-3′ |
| COL1A1 |
5′-TGGGAGGAAGCAAAAGACTC-3′ |
5′-GGGTCATTTCCACATGCTTT-3′ |
| BSP |
5′-TGCCTTGAGCCTGCTTCC-3′ |
5′-GCAAAATTAAAGCAGTCTTCATTTTG-3′ |
| BMP2 |
5′-AACACTGTGCGCAGCTTCC-3′ |
5′-CTCCGGGTTGTTTTCCCAC-3′ |
| OSX |
5′-CCCCACCTCTTGCAACCA-3′ |
5′-CCCCACCTATTGCAACCA-3′ |
| OPN |
5′-GCCGACCAAGGAAAACTCACT-3′ |
5′-GGCACAGGTGATGCCTAGGA-3′ |
| OCN |
5′-CCCAGGCGCTACCTGTATCAA-3′ |
5′-GGTCAGCCAACTCGTCACAGTC-3′ |
| β-actin |
5′-TGGCACCCAGCACAATGAA-3′ |
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′ |
Western blot analysis
hBMSCs at the density of 5×104 cells/well
in 12-well plates were treated with KR-12-a6 at 0, 20, 30, and 40
µg/ml and then cultured at 37°C for 7 days. On day 7
post-KR-12-a6 treatment, hBMSCs were harvested and lysed in
immunoprecipitation lysis buffer (10 mM Tris, 0.15 M NaCl, 1%
NP-40, and 10% glycerol, pH 7.4, at 22°C) containing protease and
phosphatase inhibitor cocktails (Sigma-Aldrich; Merck KGaA). Cell
lysates were then centrifuged at 15,000 × g for 45 min at 4°C. The
supernatants were collected, and the protein concentration was
determined by BCA assay with a Bio-Rad Model 680 Plate Reader.
Then, 40 µg/sample of protein was separated via 7.5% SDS-PAGE and
transferred onto nitrocellulose membranes. The membranes were
blocked with 5% non-fat milk for 1 h at 4°C, and then incubated
with primary antibodies targeting phosphorylated (p)-Smad1/5 (cat.
no. 9516S, Cell Signaling Technology, 1:500 dilution), Smad1/5
(cat. no. sc-6201, Santa Cruz Biotechnology, 1:200 dilution), or
β-actin (cat. no. sc-1616, Santa Cruz Biotechnology, 1:200
dilution) overnight at 4°C. The following day, membranes were
washed and incubated with bovine anti-rabbit IgG-HRP secondary
antibody (cat. no. sc-2370, Santa Cruz Biotechnology, 1:1,000
dilution) or bovine anti-goat IgG-HRP secondary antibody (cat. no.
sc-2350, Santa Cruz Biotechnology, 1:1,000 dilution) for 1 h at
room temperature, and immunoreactive bands were detected and
visualized by ECL reagent (EMD Millipore) and a FluorChem FC
Imaging system (Alpha Innotech). The protein expression was then
quantified by AlphaEase FC StandAlone software (version 6.0.0.14,
Alpha Innotech).
LDN-212854 treatment of hBMSCs
hBMSCs at the density of 5×104 cells/well
in 12-well plates were pre-treated with 10 µM LDN-212854 (Selleck
Chemicals, Shanghai) or DMSO (vehicle control) at 37°C for 14 h,
then washed 3 times with plain culture medium. hBMSCs continued to
be cultured in medium containing 1 µM LDN-212854 alone or 1
µM LDN-212854 in combination with 40 µg/ml KR-12-a6
at 37°C for 7 days. After washing for times with PBS, hBMSCs were
harvested and subjected to western blot analysis.
Statistical analysis
Each experiment was repeated at least three times,
data are expressed as the mean ± standard deviation. One-way
analysis of variance (ANOVA) followed by Tukey's post hoc test was
used to determine statistical significance. All statistical
analyses were performed using SPSS 19.0 (IBM Corp.). P<0.05 was
considered to indicate a statistically significant difference.
Results
Intensity of ALP and alizarin red
staining increases with elevated concentrations of KR-12-a6 in
hBMSCs
To determine the effects of KR-12-a6 on the
osteogenic differentiation of hBMSCs, ALP staining and alizarin red
staining were performed. With increasing concentrations of
KR-12-a6, the intensity of ALP (Fig.
1) and alizarin red staining (Fig.
2) also increased. KR-12-a6 at 40 µg/ml exhibited the
strongest effects on both ALP (Fig.
1) and alizarin red staining (Fig.
2), whereas KR-12-a6 at 60 and 80 µg/ml did not notably
increase the staining intensity compared with KR-12-a6 at 40
µg/ml.
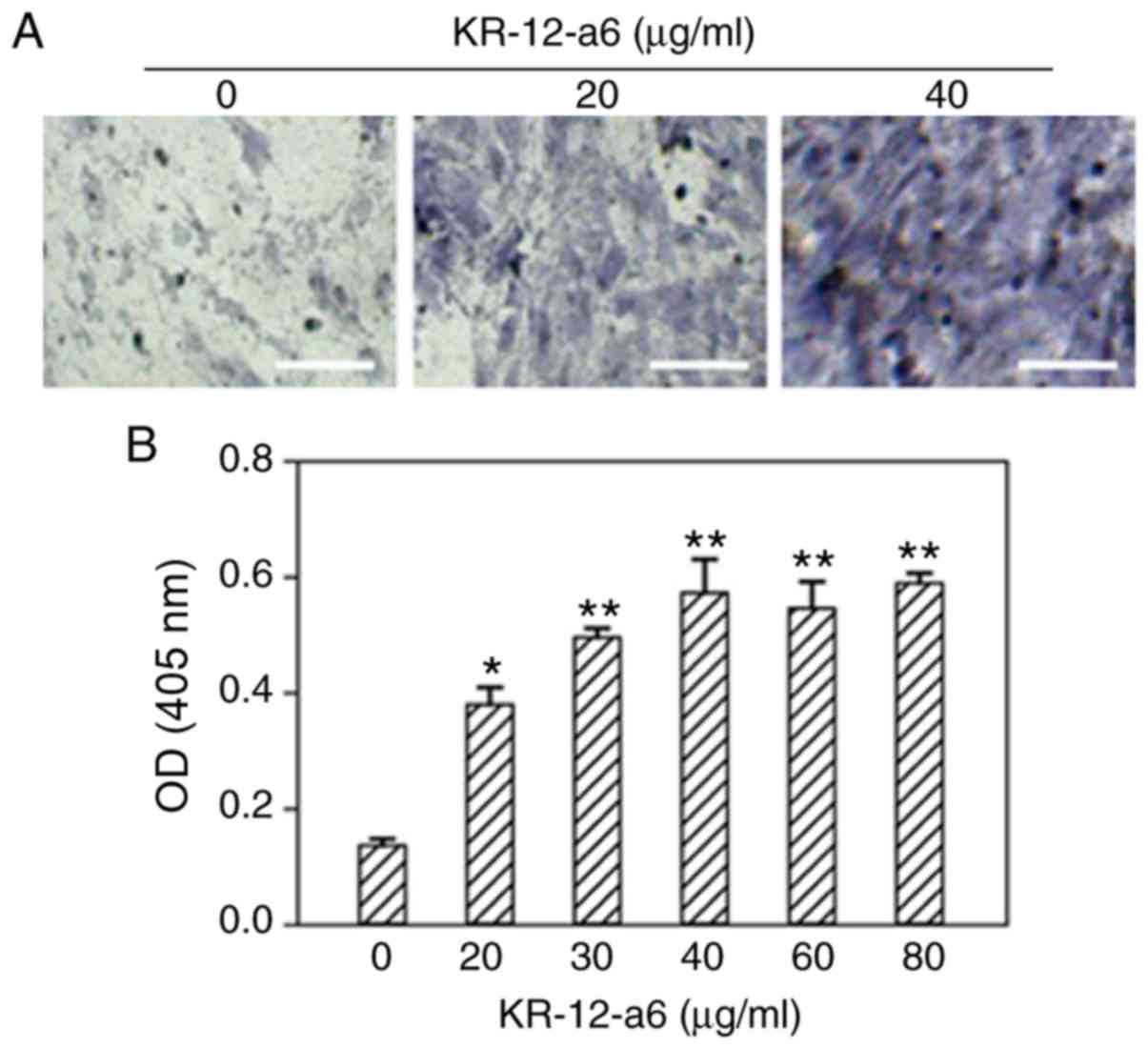 | Figure 1.Osteogenic ALP staining of hBMSCs and
quantitative analysis following KR-12-a6 treatment. ALP staining
was performed following KR-12-a6 stimulation at different
concentrations (0, 20, 30, 40, 60, and 80 µg/ml) in hBMSCs. (A)
Representative images following stimulation with 0, 20, and 40
µg/ml KR-12-a6. (B) Optical density of staining at 405 nm. Data are
presented asthe mean ± SD (n=4). *P<0.05, **P<0.01 vs.
KR-12-a6 at 0 µg/ml. Scale bar=100 µm. ALP, alkaline phosphate; OD,
optical density; hBMSC, human bone marrow mesenchymal stem
cell. |
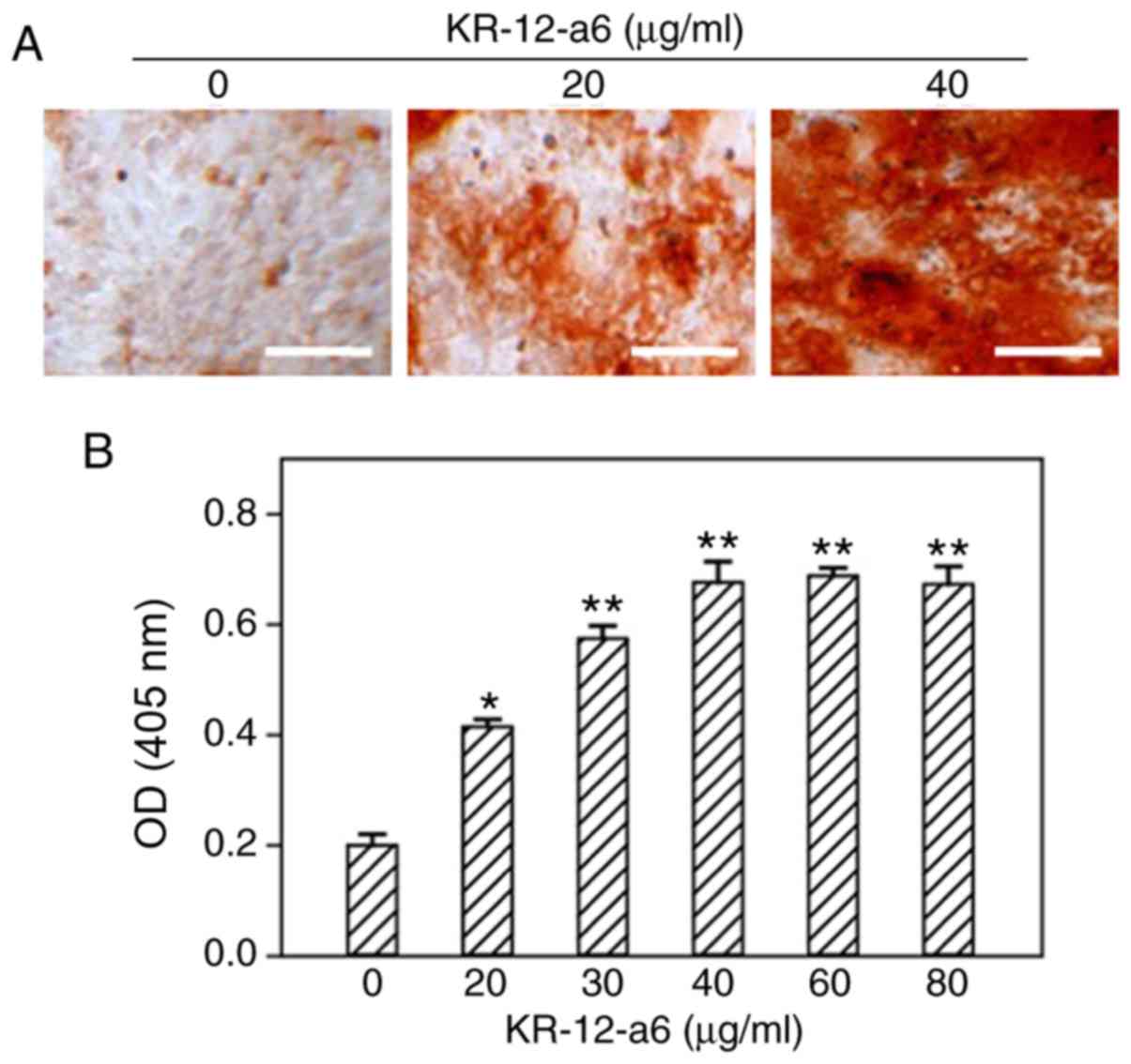 | Figure 2.Osteogenic alizarin red staining of
hBMSCs and quantitative analysis following KR-12-a6 treatment.
Alizarin red staining was performed following KR-12-a6 stimulation
at different concentrations (0, 20, 30, 40, 60, and 80 µg/ml) in
hBMSCs. (A) Representative images following stimulation with 0, 20,
and 40 µg/ml KR-12-a6. (B) Optical density of staining at 620 nm.
Data are presented as the mean ± SD (n=4). *P<0.05, **P<0.01
vs. KR-12-a6 at 0 µg/ml. Scale bar=100 µm. OD, optical density;
hBMSC, human bone marrow mesenchymal stem cell. |
mRNA expression of osteoblastic
differentiation-associated genes increases after KR-12-a6
stimulation at different time pointsin hBMSCs
The mRNA expression levels of osteoblastic
differentiation-associated genes, including RUNX2 (encoding
runt-related transcription factor 2; Fig. 3A), ALP (Fig. 3B), COL1A1 (encoding type 1
collagen alpha 1 chain; Fig. 3C),
BSP (encoding bone sialoprotein; Fig. 3D), BMP2 (encoding bone
morphogenic protein 2; Fig. 3E),
OSX (encoding osterix; Fig.
3F), OCN (encoding osteocalcin; Fig. 3G) and OPN (encoding
osteopontin; Fig. 3H), were
determined via RT-qPCR analysis following treatment of hBMSCs with
KR-12-a6 for 3, 7 or 14 days. The mRNA levels of RUNX2 and
ALP increased in a dose-dependent manner as early as 3 days
post-KR-12-a6 treatment. The mRNA expression of COL1A1, BSP
and BMP2 was significantly upregulated from day 7
post-KR-12-a6 treatment compared with the control. In contrast, the
mRNA levels of OSX, OCN and OPN were only
significantly upregulated at day 14 following KR-12-a6
stimulation.
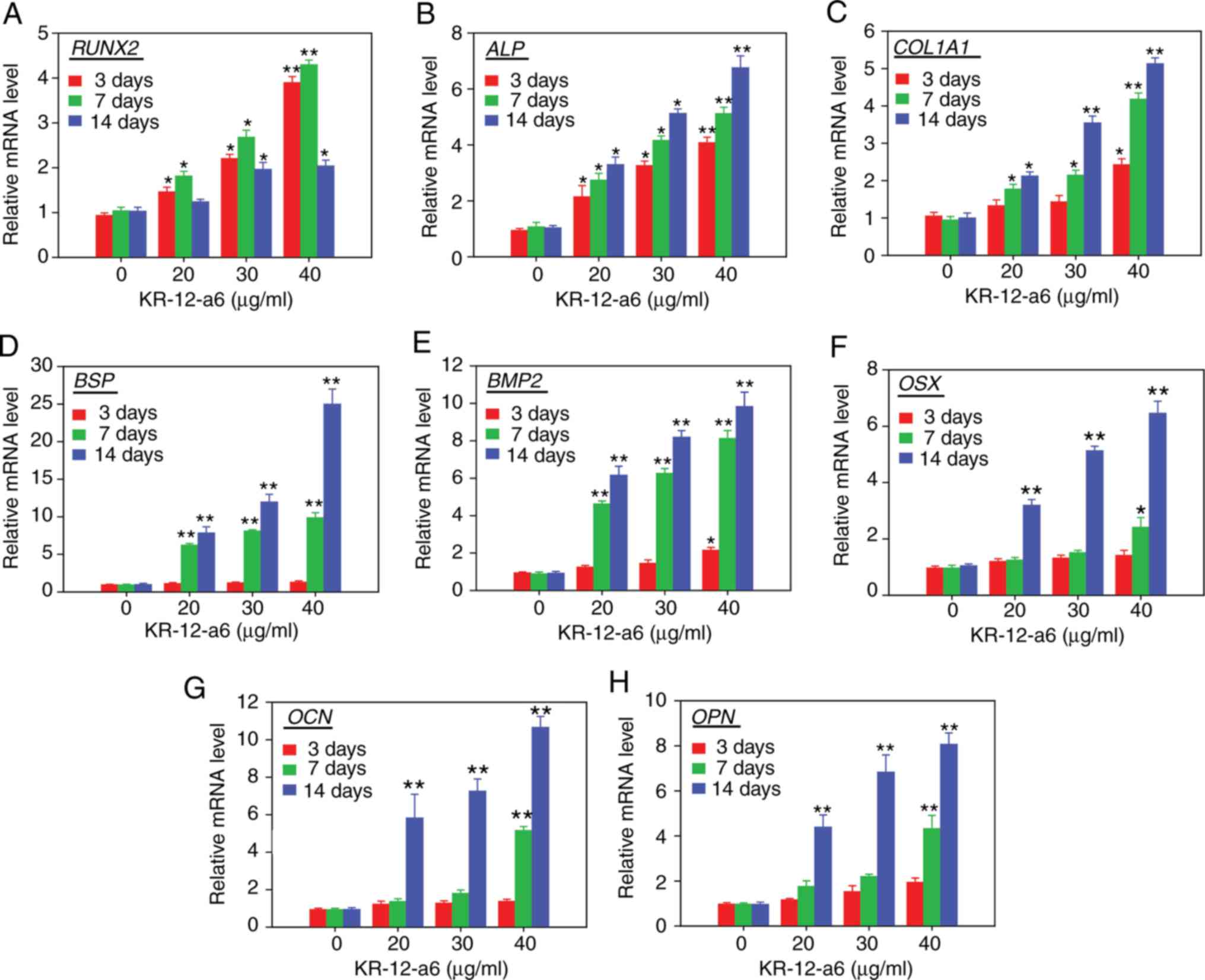 | Figure 3.Effects of KR-12-a6 on the mRNA
expression of osteogenic differentiation markers. Human bone marrow
mesenchymal stem cells were treated with KR-12-a6 at concentrations
of 0, 20, 30 or 40 µg/ml, and the mRNA levels of (A) RUNX2,
(B) ALP, (C) COL1A1, (D) BSP, (E) BMP2,
(F) OSX, (G) OCN and (H) OPN were determined
via reverse transcription-quantitative PCR on days 3, 7 and 14
post-KR-12-a6 treatment. Data are presented as the mean ± SD (n=4).
*P<0.05, **P<0.01 vs. KR-12-a6 at 0 µg/ml. ALP,
alkaline phosphatase; BMP, bone morphogenic protein;
BSP, bone sialoprotein; COL1A1, type 1 collagen α1
chain; OCN, osteocalcin; OPN, osteopontin;
OSX, osterix; RUNX2, runt-related transcription
factor 2. |
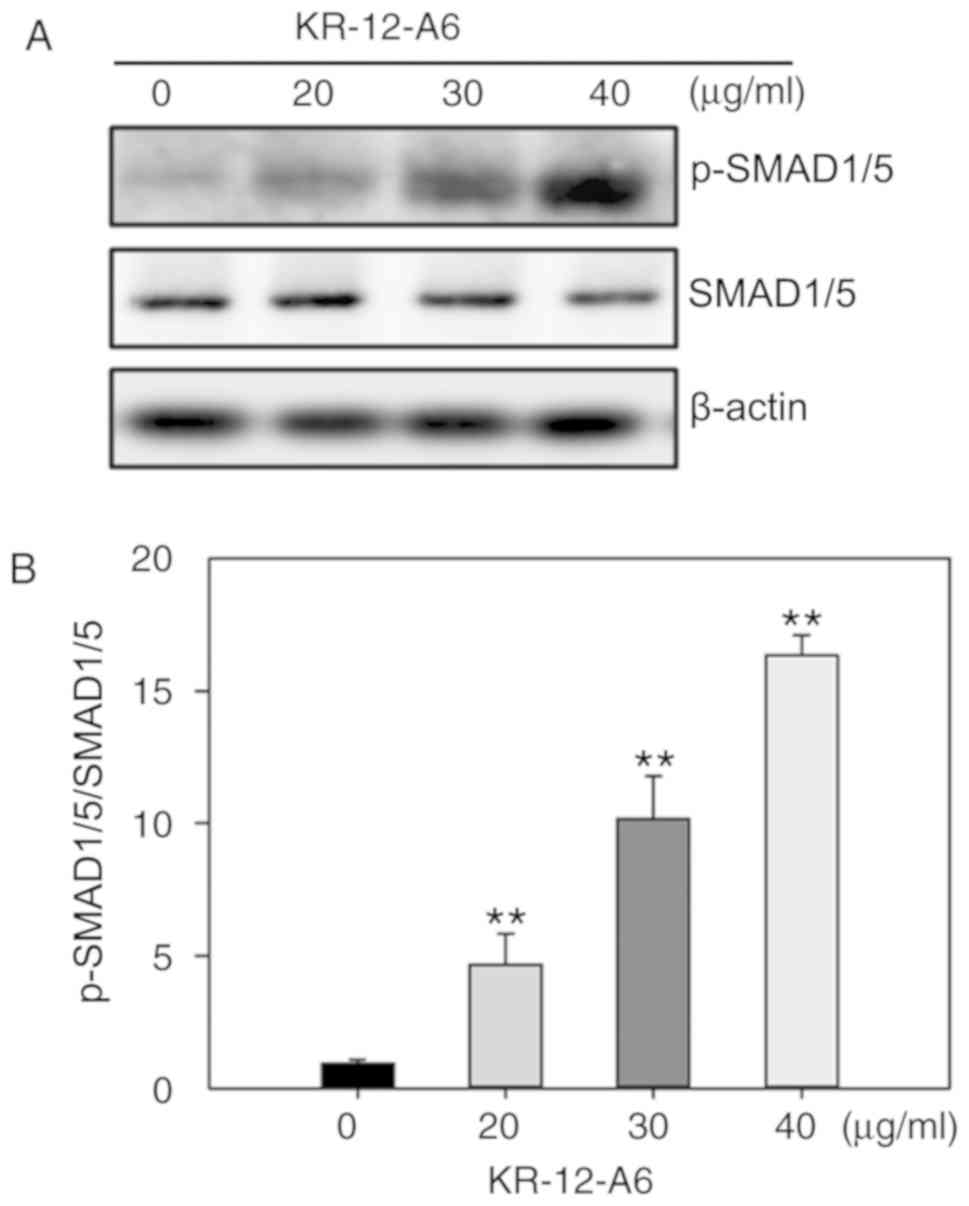
BMP/SMAD signaling is activated during
KR-12-a6-induced hBMSC osteogenic differentiation
As a significant elevation of BMP2 mRNA was
observed in Fig. 3E, it was next
investigated as to whether BMP/SMAD signaling was involved in
KR-12-a6-induced hBMSC osteogenic differentiation. The activation
of SMAD signaling was examined via western blotting following
KR-12-a6-induced hBMSC osteogenesis. The results showed that
KR-12-a6 promoted the phosphorylation of Smad1/5 in a
dose-dependent manner following 7 days of KR-12-a6 treatment
(Fig. 4A and B) and exhibited the
maximum activation at 40 µg/ml. These results suggested that
KR-12-a6 activated BMP/SMAD signaling in a dose-dependent
manner.
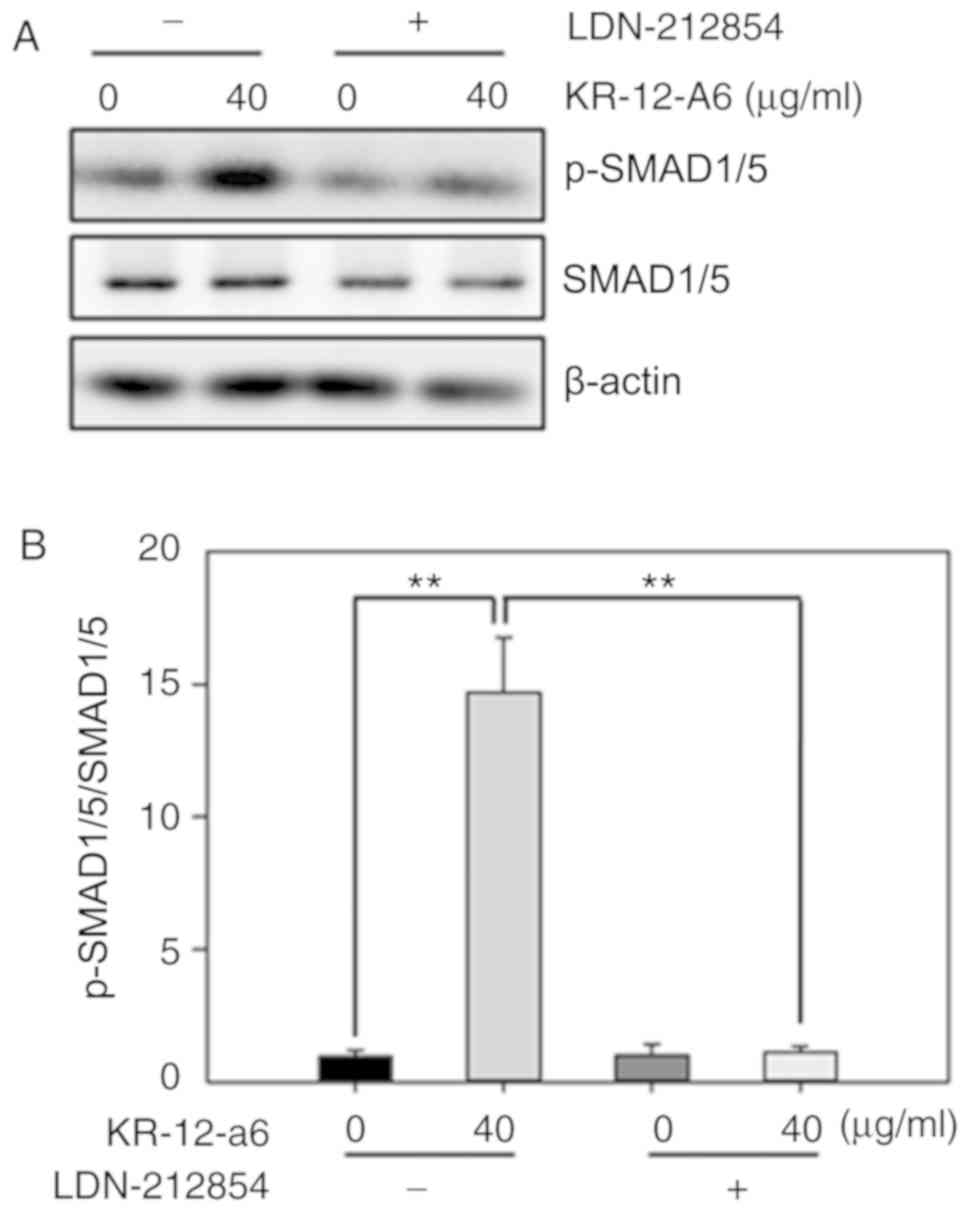
Inhibition of BMP/SMAD signaling
suppresses KR-12-a6-induced osteogenic differentiation of
hBMSCs
To further elucidate the role of BMP/SMAD signaling
in osteoblast differentiation, LDN-212854, a novel BMP inhibitor
that exhibits greater selectivity for BMP compared with the TGF-β
type I receptors, was used to suppress BMP/SMAD signaling. Western
blotting was performed to observe the changes of several Smad
proteins after 7 days of KR-12-a6 treatment with or without
LDN-212854 (Fig. 5). The results
showed that KR-12-a6 at 40 µg/ml significantly promoted the
phosphorylation of Smad1/5 in hBMSCs, which was consistent with the
results in Fig. 4. However, the
use of LDN-212854 significantly reduced Smad1/5 phosphorylation in
KR-12-a6-treated hBMSCs (Fig. 5).
RT-qPCR analysis was then performed to further examine the effects
of LDN-212854 (Fig. 6). The
results showed that LDN-212854, which inhibited BMP/SMAD signaling,
significantly suppressed the mRNA expression of several
osteoblastic differentiation-associated genes, including
RUNX2 (Fig. 6A), ALP
(Fig. 6B), COL1A1 (Fig. 6C), BSP (Fig. 6D), BMP2 (Fig. 6E), OSX (Fig. 6F), OCN (Fig. 6G), and OPN (Fig. 6H) in hBMSCs at day 7 post-KR-12-a6
treatment. Collectively, these results indicated that BMP/SMAD
signaling exerts a positive role in KR-12-a6-induced hBMSC
osteogenesis.
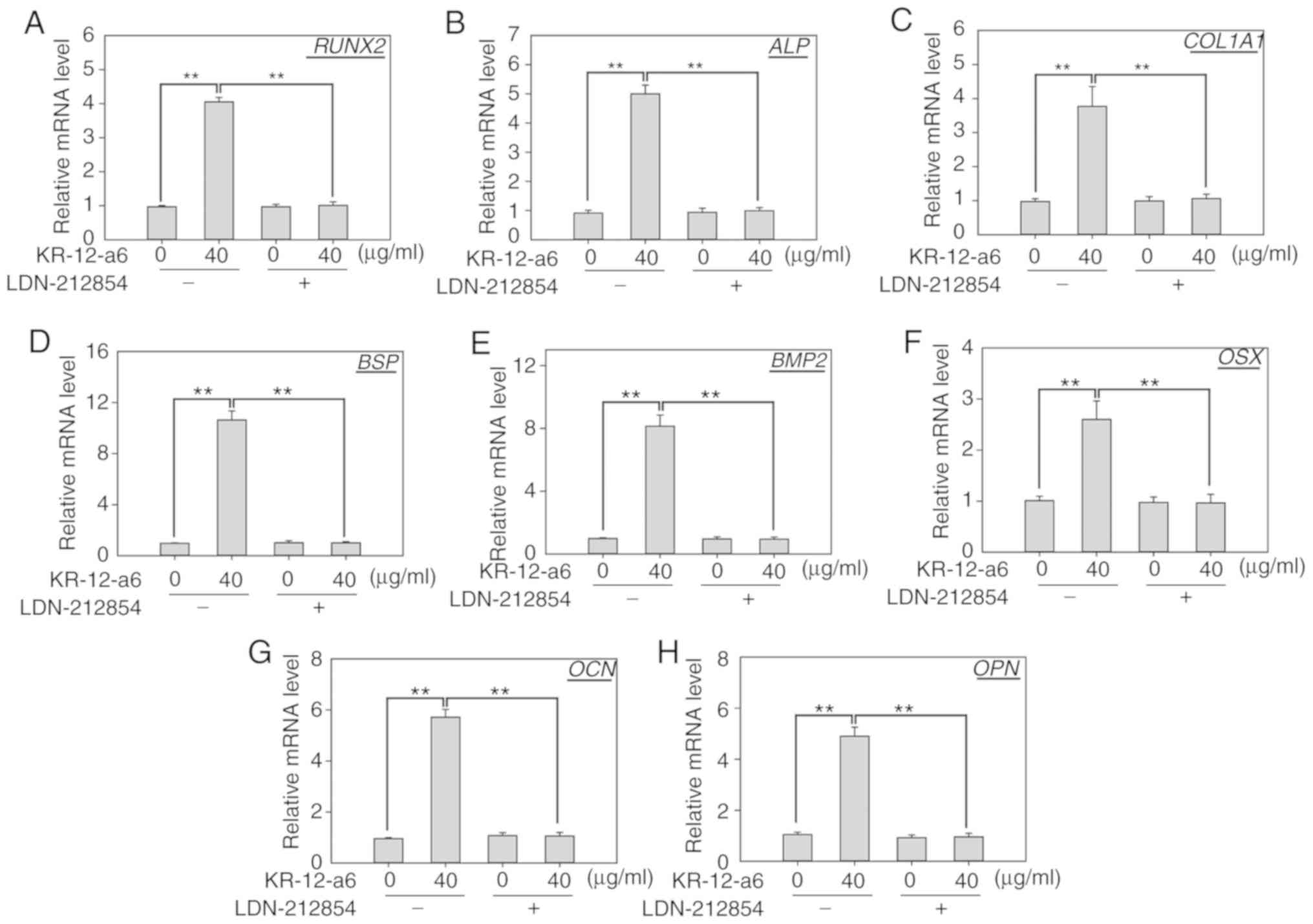 | Figure 6.Effects of LDN-212854 on the mRNA
expression of osteogenic differentiation-associated genes in
KR-12-a6-treated hBMSCs. Reverse transcription-quantitative PCR was
performed to examine them RNA expression of (A) RUNX2, (B)
ALP, (C) COL1A1, (D) BSP, (E) BMP2, (F)
OSX, (G) OCN and (H) OPN in hBMSCs on day 7
following the various treatments: Control, KR-12-a6, LDN-212854,
and KR-12-a6+LDN-212854. Data are presented as the mean ± SD (n=4).
**P<0.01 vs. KR-12-a6 at 40 µg/ml without LDN-212854.
ALP, alkaline phosphatase; BMP, bone morphogenic
protein; BSP, bone sialoprotein; COL1A1, type 1
collagen alpha 1 chain; hBMSC, human bone marrow mesenchymal stem
cell; OCN, osteocalcin; OPN, osteopontin; OSX,
osterix; RUNX2, runt-related transcription factor 2. |
Discussion
Bone infection and osteolysis are common clinical
symptoms of osteomyelitis, which requires local or systemic
treatment with antibiotics (32).
With the rise of drug-resistant bacteria in clinical bone
infection, traditional antibiotics have become less effective
(18–20). Due to their low drug resistance and
excellent antimicrobial properties, AMPs have received increasing
attention. AMPs are an important part of the innate immune system,
and are regarded as potential substitutes for traditional
antibiotics (33). Antimicrobial
peptide LL-37 is an important antimicrobial substance that is
naturally synthesized in the human body (7,13).
It is the first line of defense against local infection and
systemic pathogen invasion; more importantly, it does not lead to
bacterial resistance (9). However,
because of its long amino acid sequence, LL-37 is difficult to
develop as a clinical drug for infectious diseases (10,11).
As the shortest active fragment of LL-37, KR-12 has the advantages
of low cost for synthesis and low cytotoxicity; it is thus
predicted to serve an important role in the treatment of infections
caused by drug-resistant bacteria (10,11).
As an analogue of KR-12, KR-12-a6 has the same antibacterial
properties, and is also a potential drug for the clinical treatment
of osteomyelitis (15).
Antibiotics that are commonly used to control
infection do not promote the formation of new bones; rather, they
inhibit the local osteogenic microenvironment and show no effect on
the clinical treatment of infection-associated osteolysis (19,20).
Studies have shown that antibiotics, including gentamicin and
vancomycin, inhibit osteoblast proliferation and differentiation
(19,20). In contrast, certain small peptides
can control infection while promoting new bone formation (21). Various small AMPs and their
analogues may have the ability to promote bone differentiation
while also possessing antibacterial properties. Therefore, treating
osteomyelitis with AMPs may promote bone repair following
infection-induced osteolysis while also controlling the infection.
Due to the ability to differentiate into osteoblasts, hBMSCs have
been widely used in the study of osteogenic differentiation
(24). To verify whether KR-12-a6,
a KR-12 analogue, can also affect the osteogenic differentiation of
hBMSCs, primary hBMSCs were selected as experimental subjects in
this study.
The results showed that KR-12-a6 enhanced the
osteogenic differentiation and mineralization of hBMSCs in a
dose-dependent manner. KR-12-a6 at 40 µg/ml significantly promoted
osteogenic differentiation, indicating that KR-12-a6can effectively
promote osteogenic differentiation at high concentrations. To
accurately evaluate the expression levels of osteoblast-associated
genes after KR-12-a6 stimulation, several osteoblast-associated
genes were detected at different stages of osteogenic
differentiation. During the first 7 days of osteogenesis, the mRNA
levels of RUNX2, ALP, COL1A1, BSP and BMP2 increased
significantly. In contrast, the late-stage markers of osteogenesis,
including OSX, OCN and OPN, were upregulated during
the second week of osteogenic differentiation. RT-qPCR analysis at
different stages revealed that KR-12-a6 enhanced the expression of
osteoblast-associated genes in hBMSCs; these effects were more
notable at high concentrations of KR-12-a6. These findings
suggested that KR-12-a6 may promote the osteogenic differentiation
of hBMSCs in vitro. Further mechanistic studies revealed
that KR-12-a6 significantly promoted the phosphorylation of
Smad1/5. When LDN-212854, a selective BMP inhibitor, was used to
block BMP/SMAD signaling, the activation of p-Smad1/5 was
suppressed. Furthermore, the mRNA expression of several osteogenic
differentiation-associated genes was also inhibited in
KR-12-a6-treated hBMSCs, suggesting the involvement of BMP/SMAD
signaling in the KR-12-a6-induced osteogenic differentiation of
hBMSCs. However, the current study showed certain limitations.
First, results obtained with the use of primary hBMSCs could not
fully represent the biophysiological events that happen in
vivo and the translational relevance of the study should be
strictly confirmed in animal studies in vivo. Second, the
activation of BMP/SMAD signaling might not be the only mechanism
through which KR-12-a6 enhanced osteogenesis, other potential
signaling pathways involved in osteogenesis should also be
investigated.
In conclusion, KR-12-a6 promoted the osteogenic
differentiation of hBMSCs in a dose-dependent manner, and BMP/SMAD
signaling was involved in the process. In addition, considering the
potential for the use of AMPs in the treatment of osteomyelitis and
other infections, discovering more peptides with antibacterial
properties and bone formation regulation is of clear importance for
clinical treatment. KR-12-a6 may be an effective drug for the
prevention and treatment of local osteomyelitis and infectious
osteolysis.
Acknowledgements
The authors would like to thank Dr Yan C. Cheng
(senior scientist, Center for Biomedical Research, the Rockefeller
University, United States) for his valuable opinions in the
discussion of the present study.
Funding
This work was supported by the Shaanxi Province Key
Research & Development Projects (grant no. 2017kw-043) and the
Talent Support Program of Air Force Military Medical University
‘Project Ling Yun’ (grant no. cyjhsll).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LS and LF designed the research and wrote the
manuscript; LF and PJ performed experiments and analyzed data; YH
and HL collected and analyzed data; LS and LF revised the
manuscript and approved the final submission. All authors discussed
the results and reviewed the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Huazhong University of Science and Technology.
Informed consents were obtained from all donors.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bechinger B and Gorr SU: Antimicrobial
peptides: Mechanisms of action and resistance. J Dent Res.
96:254–260. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Larrick JW, Hirata M, Balint RF, Lee J,
Zhong J and Wright SC: Human CAP18: A novel antimicrobial
lipopolysaccharide-binding protein. Infect Immun. 63:1291–1297.
1995.PubMed/NCBI
|
|
3
|
Cowland JB, Johnsen AH and Borregaard N:
hCAP-18, a cathelin/pro-bactenecin-like protein of human neutrophil
specific granules. FEBS Lett. 368:173–176. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chakraborty K, Ghosh S, Koley H,
Mukhopadhyay AK, Ramamurthy T, Saha DR, Mukhopadhyay D,
Roychowdhury S, Hamabata T, Takeda Y and Das S: Bacterial exotoxins
downregulate cathelicidin (hCAP-18/LL-37) and human beta-defensin 1
(HBD-1) expression in the intestinal epithelial cells. Cell
Microbiol. 10:2520–2537. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Agier J, Brzezińska-Błaszczyk E,
Żelechowska P, Wiktorska M, Pietrzak J and Różalska S: Cathelicidin
LL-37 affects surface and intracellular toll-like receptor
expression in tissue mast cells. J Immunol Res. 2018:73571622018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chamilos G, Gregorio J, Meller S, Lande R,
Kontoyiannis DP, Modlin RL and Gilliet M: Cytosolic sensing of
extracellular self-DNA transported into monocytes by the
antimicrobial peptide LL37. Blood. 120:3699–2707. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Agerberth B, Charo J, Werr J, Olsson B,
Idali F, Lindbom L, Kiessling R, Jörnvall H, Wigzell H and
Gudmundsson GH: The human antimicrobial and chemotactic peptides
LL-37 and alpha-defensins are expressed by specific lymphocyte and
monocyte populations. Blood. 96:3086–3093. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dorschner RA, Pestonjamasp VK, Tamakuwala
S, Ohtake T, Rudisill J, Nizet V, Agerberth B, Gudmundsson GH and
Gallo RL: Cutaneous injury induces the release of cathelicidin
anti-microbial peptides active against group A
Streptococcus. J Invest Dermatol. 117:91–97. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Duplantier AJ and van Hoek ML: The human
cathelicidin antimicrobial peptide LL-37 as a potential treatment
for polymicrobial infected wounds. Front Immunol. 4:1432013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang G: Structures of human host defense
cathelicidin LL-37 and its smallest antimicrobial peptide KR-12 in
lipid micelles. J Biol Chem. 283:32637–32643. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mishra B, Epand RF, Epand RM and Wang G:
Structural location determines functional roles of the basic amino
acids of KR-12, the smallest antimicrobial peptide from human
cathelicidin LL-37. RSC Adv. Nov 14–2013.(Epub ahead of print).
doi: 10.1039/C3RA42599A. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rico-Mata R, De Leon-Rodriguez LM and
Avila EE: Effect of antimicrobial peptides derived from human
cathelicidin LL-37 on Entamoeba histolytica trophozoites.
Exp Parasitol. 133:300–306. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Feng X, Sambanthamoorthy K, Palys T and
Paranavitana C: The human antimicrobial peptide LL-37 and its
fragments possess both antimicrobial and antibiofilm activities
against multidrug-resistant Acinetobacter baumannii.
Peptides. 49:131–137. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo Y, McLean DT, Linden GJ, McAuley DF,
McMullan R and Lundy FT: The naturally occurring host defense
peptide, LL-37, and its truncated mimetics KE-18 and KR-12 have
selected biocidal and antibiofilm activities against Candida
albicans, Staphylococcus aureus, and Escherichia coli in
vitro. Front Microbiol. 8:5442017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jacob B, Park IS, Bang JK and Shin SY:
Short KR-12 analogs designed from human cathelicidin LL-37
possessing both antimicrobial and antiendotoxic activities without
mammalian cell toxicity. J Pept Sci. 19:700–707. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Geurts J, Hohnen A, Vranken T and Moh P:
Treatment strategies for chronic osteomyelitis in low- and
middle-income countries: Systematic review. Trop Med Int Health.
22:1054–1062. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mortazavi MM, Khan MA, Quadri SA, Suriya
SS, Fahimdanesh KM, Fard SA, Hassanzadeh T, Taqi MA, Grossman H and
Tubbs RS: Cranial osteomyelitis: A comprehensive review of modern
therapies. World Neurosurg. 111:142–153. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fily F, Ronat JB, Malou N, Kanapathipillai
R, Seguin C, Hussein N, Fakhri RM and Langendorf C: Post-traumatic
osteomyelitis in Middle East war-wounded civilians: Resistance to
first-line antibiotics in selected bacteria over the decade
2006–2016. BMC Infect Dis. 19:1032019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ince A, Schütze N, Karl N, Löhr JF and
Eulert J: Gentamicin negatively influenced osteogenic function in
vitro. Int Orthop. 31:223–228. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mantripragada VP and Jayasuriya AC: Effect
of dual delivery of antibiotics (vancomycin and cefazolin) and
BMP-7 from chitosan microparticles on Staphylococcus
epidermidis and pre-osteoblasts in vitro. Mater Sci Eng C Mater
Biol Appl. 67:409–417. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Choe H, Narayanan AS, Gandhi DA, Weinberg
A, Marcus RE, Lee Z, Bonomo RA and Greenfield EM: Immunomodulatory
peptide IDR-1018 decreases implant infection and preserves
osseointegration. Clin Orthop Relat Res. 473:2898–2907. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zou W, Greenblatt MB, Brady N, Lotinun S,
Zhai B, de Rivera H, Singh A, Sun J, Gygi SP, Baron R, et al: The
microtubule-associated protein DCAMKL1 regulates osteoblast
function via repression of Runx2. J Exp Med. 210:1793–1806. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mansour A, Abou-Ezzi G, Sitnicka E,
Jacobsen SE, Wakkach A and Blin-Wakkach C: Osteoclasts promote the
formation of hematopoietic stem cell niches in the bone marrow. J
Exp Med. 209:537–549. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Crane JL and Cao X: Bone marrow
mesenchymal stem cells and TGF-β signaling in bone remodeling. J
Clin Invest. 124:466–472. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li B: MicroRNA regulation in osteogenic
and adipogenic differentiation of bone mesenchymal stem cells and
its application in bone regeneration. Curr Stem Cell Res Ther.
13:26–30. 2018.PubMed/NCBI
|
|
26
|
Bidwell JP, Alvarez MB, Hood M Jr and
Childress P: Functional impairment of bone formation in the
pathogenesis of osteoporosis: The bone marrow regenerative
competence. Curr Osteoporos Rep. 11:117–125. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shen GS, Zhou HB, Zhang H, Chen B, Liu ZP,
Yuan Y, Zhou XZ and Xu YJ: The GDF11-FTO-PPARγ axis controls the
shift of osteoporotic MSC fate to adipocyte and inhibits bone
formation during osteoporosis. Biochim Biophys Acta Mol Basis Dis.
1864:3644–3654. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pavone V, Testa G, Giardina SMC, Vescio A,
Restivo DA and Sessa G: Pharmacological therapy of osteoporosis: A
systematic current review of literature. Front Pharmacol.
8:8032017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Faber C, Stallmann HP, Lyaruu DM, Joosten
U, von Eiff C, van NieuwAmerongen A and Wuisman PI: Comparable
efficacies of the antimicrobial peptide human lactoferrin 1–11 and
gentamicin in a chronic methicillin-resistant Staphylococcus
aureus osteomyelitis model. Antimicrob Agents Chemother.
49:2438–2444. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xing K, Huang G, Hua S, Xu G and Li M:
Systematic review of randomized controlled trials on antibiotic
treatment for osteomyelitis in diabetes. Diabet Med. 36:546–556.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sierra JM, Fusté E, Rabanal F, Vinuesa T
and Viñas M: An overview of antimicrobial peptides and the latest
advances in their development. Expert Opin Biol Ther. 17:663–676.
2017. View Article : Google Scholar : PubMed/NCBI
|